
lncRNAs UC.145 and PRKG1-AS1 Determine the Functional Output of DKK1 in Regulating the Wnt Signaling Pathway in Gastric Cancer

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture
2.2. Sample Collection and Preparation
2.3. Antibodies and Reagents
2.4. Total RNA Extraction and Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction
2.5. Methylation-Specific PCR
2.6. Chromatin Immunoprecipitation Assay
2.7. Gene Expression Profiling
2.8. Small Interfering RNA (siRNA) Transfection
2.9. Construction of lncRNA Overexpression Vector
2.10. Cell Viability Analysis
2.11. Apoptosis Analysis
2.12. Invasion and Migration Analysis
2.13. Colony Formation Assay
2.14. Western Blot Analysis
2.15. RNA Immunoprecipitation
2.16. Statistical Analysis
3. Results
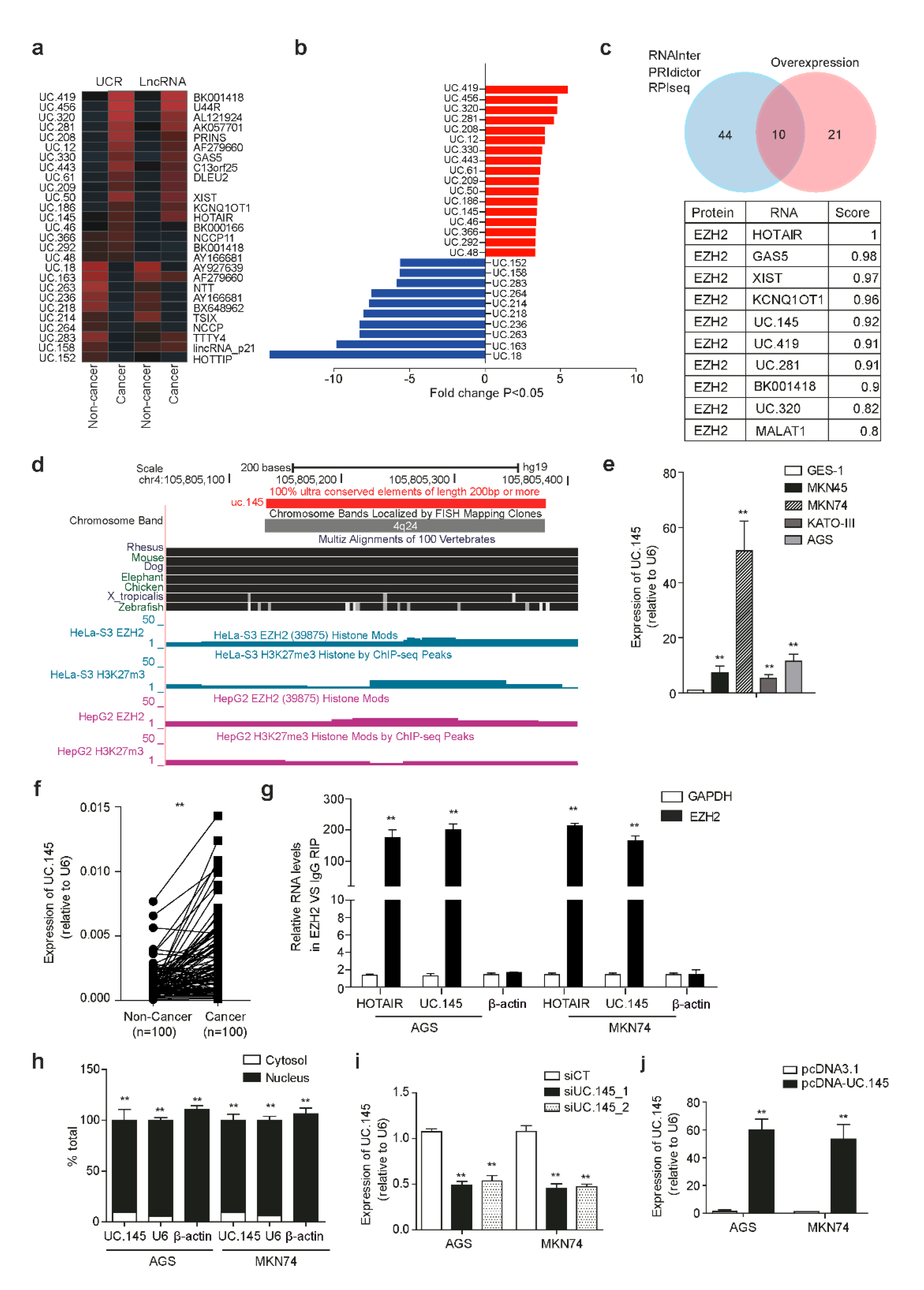
3.1. Characteristics of T-UCRs in GC
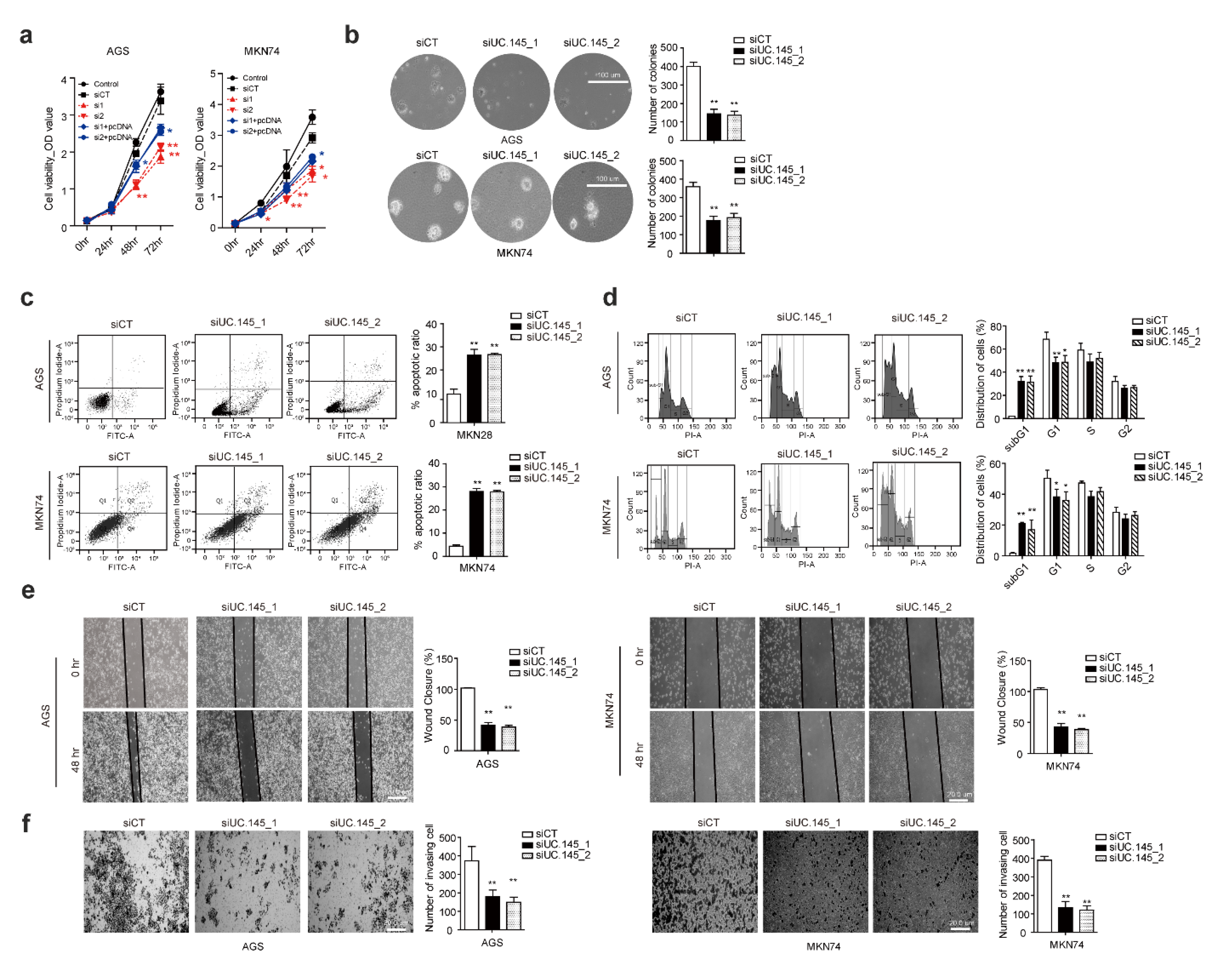
3.2. UC.145 Regulates In Vitro Proliferation of GC Cells
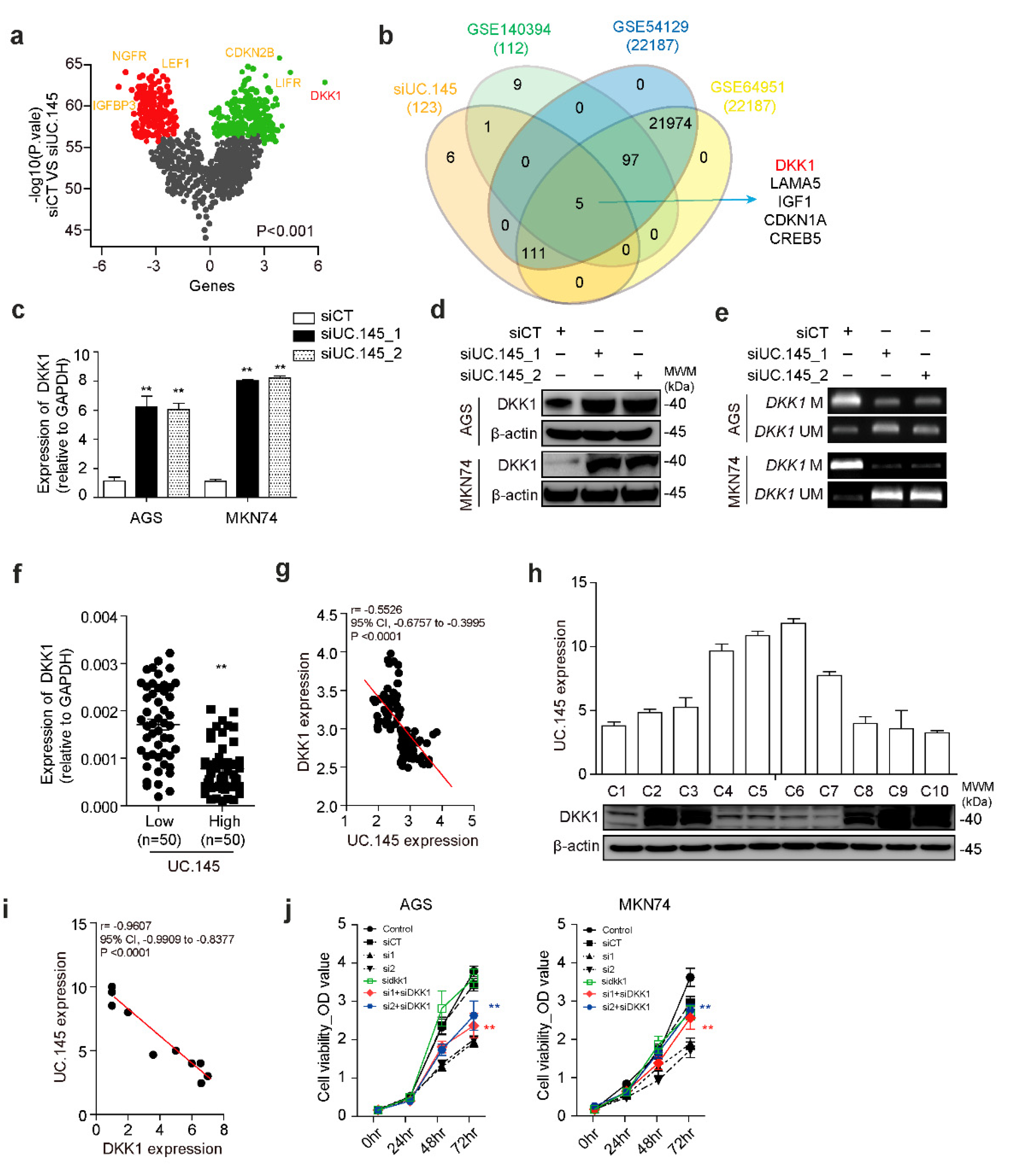
3.3. DKK1 Is a Target of UC.145 in GC
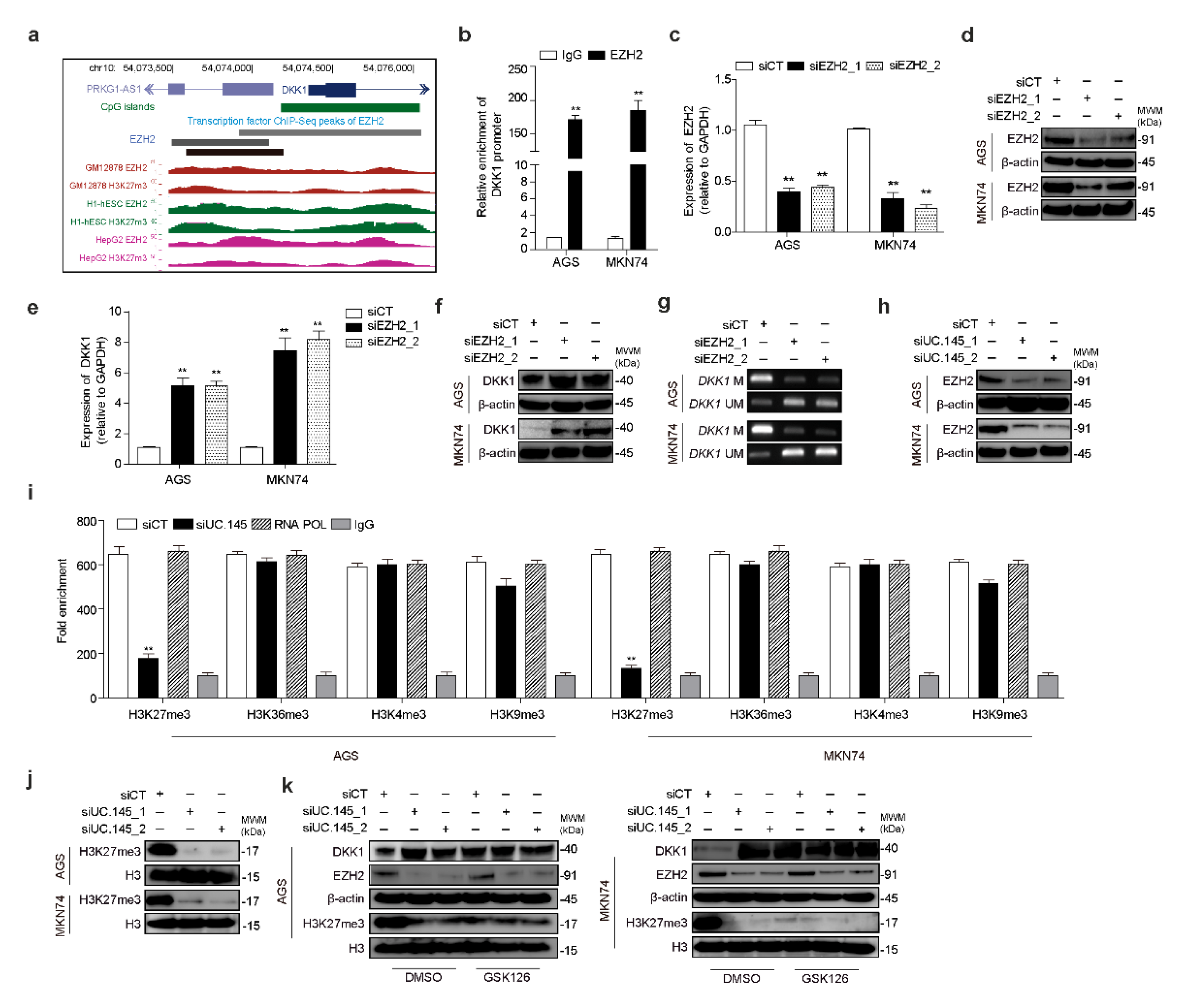
3.4. UC.145 Regulates DKK1 by Interacting with EZH2
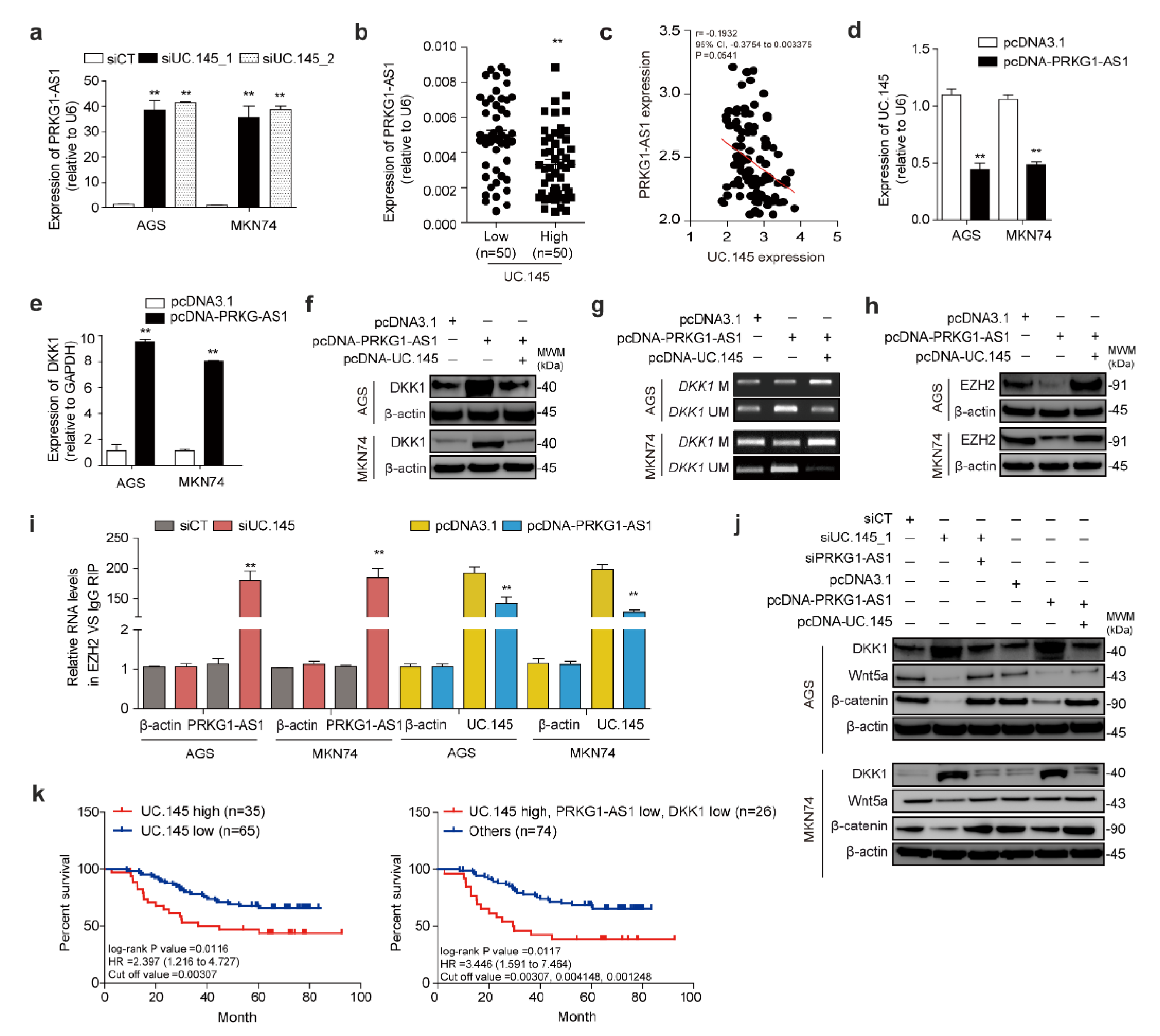
3.5. Novel lncRNA PRKG1-AS1, Which Is Controlled by UC.145, Regulates DKK1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alzeeb, G.; Arzur, D.; Trichet, V.; Talagas, M.; Corcos, L.; Le Jossic-Corcos, C. Gastric cancer cell death analyzed by live cell imaging of spheroids. Sci. Rep. 2022, 12, 1488. [Google Scholar] [CrossRef]
- Arnes, M.; Casas Tinto, S. Aberrant Wnt signaling: A special focus in CNS diseases. J. Neurogenet. 2017, 31, 216–222. [Google Scholar] [CrossRef]
- Niida, A.; Hiroko, T.; Kasai, M.; Furukawa, Y.; Nakamura, Y.; Suzuki, Y.; Sugano, S.; Akiyama, T. DKK1, a negative regulator of Wnt signaling, is a target of the beta-catenin/TCF pathway. Oncogene 2004, 23, 8520–8526. [Google Scholar] [CrossRef] [Green Version]
- Vargas, J.Y.; Fuenzalida, M.; Inestrosa, N.C. In vivo activation of Wnt signaling pathway enhances cognitive function of adult mice and reverses cognitive deficits in an Alzheimer’s disease model. J. Neurosci. 2014, 34, 2191–2202. [Google Scholar] [CrossRef] [Green Version]
- Hirata, H.; Hinoda, Y.; Nakajima, K.; Kawamoto, K.; Kikuno, N.; Ueno, K.; Yamamura, S.; Zaman, M.S.; Khatri, G.; Chen, Y.; et al. Wnt antagonist DKK1 acts as a tumor suppressor gene that induces apoptosis and inhibits proliferation in human renal cell carcinoma. Int. J. Cancer 2011, 128, 1793–1803. [Google Scholar] [CrossRef]
- Yaccoby, S.; Ling, W.; Zhan, F.; Walker, R.; Barlogie, B.; Shaughnessy, J.D., Jr. Antibody-based inhibition of DKK1 suppresses tumor-induced bone resorption and multiple myeloma growth in vivo. Blood 2007, 109, 2106–2111. [Google Scholar] [CrossRef]
- Forget, M.A.; Turcotte, S.; Beauseigle, D.; Godin-Ethier, J.; Pelletier, S.; Martin, J.; Tanguay, S.; Lapointe, R. The Wnt pathway regulator DKK1 is preferentially expressed in hormone-resistant breast tumours and in some common cancer types. Br. J. Cancer 2007, 96, 646–653. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Sancho, J.M.; Aguilera, O.; Garcia, J.M.; Pendas-Franco, N.; Pena, C.; Cal, S.; Garcia de Herreros, A.; Bonilla, F.; Munoz, A. The Wnt antagonist DICKKOPF-1 gene is a downstream target of beta-catenin/TCF and is downregulated in human colon cancer. Oncogene 2005, 24, 1098–1103. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Wu, J.; Chen, B.; Li, M.; Tian, Y.; He, M.; Xue, J.; Wang, J.; Bai, S.; Sharma, A.; et al. Serum Dickkopf-1 (DKK1) is significantly lower in patients with ling cancer but is rapidly normalized after treatment. Am. J. Transl. Res. 2014, 6, 850. [Google Scholar]
- Jia, X.; Li, N.; Peng, C.; Deng, Y.; Wang, J.; Deng, M.; Lu, M.; Yin, J.; Zheng, G.; Liu, H.; et al. miR-493 mediated DKK1 down-regulation confers proliferation, invasion and chemo-resistance in gastric cancer cells. Oncotarget 2016, 7, 7044–7054. [Google Scholar] [CrossRef]
- Gao, C.; Xie, R.; Ren, C.; Yang, X. Dickkopf-1 expression is a novel prognostic marker for gastric cancer. J. Biomed. Biotechnol. 2012, 2012, 804592. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Duan, X.L.; Qi, X.L.; Meng, L.; Xu, Y.S.; Wu, T.; Dai, P.G. Concurrent Hypermethylation of SFRP2 and DKK2 Activates the Wnt/beta-Catenin Pathway and Is Associated with Poor Prognosis in Patients with Gastric Cancer. Mol. Cells 2017, 40, 45–53. [Google Scholar] [CrossRef] [Green Version]
- You, B.-H.; Yoon, J.-H.; Kang, H.; Lee, E.K.; Lee, S.K.; Nam, J.-W. HERES, a lncRNA that regulates canonical and noncanonical Wnt signaling pathways via interaction with EZH2. Proc. Natl. Acad. Sci. USA 2019, 116, 24620–24629. [Google Scholar] [CrossRef] [Green Version]
- Fattahi, S.; Kosari-Monfared, M.; Golpour, M.; Emami, Z.; Ghasemiyan, M.; Nouri, M.; Akhavan-Niaki, H. LncRNAs as potential diagnostic and prognostic biomarkers in gastric cancer: A novel approach to personalized medicine. J. Cell. Physiol. 2020, 235, 3189–3206. [Google Scholar] [CrossRef]
- Chandra Gupta, S.; Nandan Tripathi, Y. Potential of long non-coding RNAs in cancer patients: From biomarkers to therapeutic targets. Int. J. Cancer 2017, 140, 1955–1967. [Google Scholar] [CrossRef]
- Ferdin, J.; Nishida, N.; Wu, X.; Nicoloso, M.S.; Shah, M.Y.; Devlin, C.; Ling, H.; Shimizu, M.; Kumar, K.; Cortez, M.A.; et al. HINCUTs in cancer: Hypoxia-induced noncoding ultraconserved transcripts. Cell Death Differ. 2013, 20, 1675–1687. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Fang, W.; Sun, L.; Lu, Y.; Dou, L.; Huang, X.; Tang, W.; Yu, L.; Li, J. Ultraconserved element uc.372 drives hepatic lipid accumulation by suppressing miR-195/miR4668 maturation. Nat. Commun. 2018, 9, 612. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.X.; Peng, J.C.; Xu, A.T.; Zhao, D.; Qiao, Y.Q.; Wang, T.R.; Shen, J.; Ran, Z.H. Noncoding Transcribed Ultraconserved Region (T-UCR) uc.261 Participates in Intestinal Mucosa Barrier Damage in Crohn’s Disease. Inflamm. Bowel Dis. 2016, 22, 2840–2852. [Google Scholar] [CrossRef]
- Terracciano, D.; Terreri, S.; de Nigris, F.; Costa, V.; Calin, G.A.; Cimmino, A. The role of a new class of long noncoding RNAs transcribed from ultraconserved regions in cancer. Biochim. Biophys. Acta Rev. Cancer 2017, 1868, 449–455. [Google Scholar] [CrossRef]
- Fabris, L.; Calin, G.A. Understanding the Genomic Ultraconservations: T-UCRs and Cancer. Int. Rev. Cell Mol. Biol. 2017, 333, 159–172. [Google Scholar] [CrossRef]
- Pereira Zambalde, E.; Mathias, C.; Rodrigues, A.C.; de Souza Fonseca Ribeiro, E.M.; Fiori Gradia, D.; Calin, G.A.; Carvalho de Oliveira, J. Highlighting transcribed ultraconserved regions in human diseases. Wiley Interdiscip. Rev. RNA 2020, 11, e1567. [Google Scholar] [CrossRef]
- Simon, J.A.; Lange, C.A. Roles of the EZH2 histone methyltransferase in cancer epigenetics. Mutat. Res. 2008, 647, 21–29. [Google Scholar] [CrossRef]
- He, L.J.; Cai, M.Y.; Xu, G.L.; Li, J.J.; Weng, Z.J.; Xu, D.Z.; Luo, G.Y.; Zhu, S.L.; Xie, D. Prognostic significance of overexpression of EZH2 and H3k27me3 proteins in gastric cancer. Asian Pac. J. Cancer Prev. 2012, 13, 3173–3178. [Google Scholar] [CrossRef] [Green Version]
- Guttman, M.; Rinn, J.L. Modular regulatory principles of large non-coding RNAs. Nature 2012, 482, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Chen, X.X.; Li, W.X.; Wu, X.Q.; Huang, C.; Xie, J.; Zhao, Y.X.; Meng, X.M.; Li, J. EZH2-mediated repression of Dkk1 promotes hepatic stellate cell activation and hepatic fibrosis. J. Cell. Mol. Med. 2017, 21, 2317–2328. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Liu, Y.; Mai, X.; Lu, S.; Jin, L.; Tai, X. STAT1-induced upregulation of LINC00467 promotes the proliferation migration of lung adenocarcinoma cells by epigenetically silencing DKK1 to activate Wnt/beta-catenin signaling pathway. Biochem. Biophys. Res. Commun. 2019, 514, 118–126. [Google Scholar] [CrossRef]
- Ritie, L.; Spenle, C.; Lacroute, J.; Bolcato-Bellemin, A.L.; Lefebvre, O.; Bole-Feysot, C.; Jost, B.; Klein, A.; Arnold, C.; Kedinger, M.; et al. Abnormal Wnt and PI3Kinase signaling in the malformed intestine of lama5 deficient mice. PLoS ONE 2012, 7, e37710. [Google Scholar] [CrossRef]
- Tiffen, J.C.; Gunatilake, D.; Gallagher, S.J.; Gowrishankar, K.; Heinemann, A.; Cullinane, C.; Dutton-Regester, K.; Pupo, G.M.; Strbenac, D.; Yang, J.Y.; et al. Targeting activating mutations of EZH2 leads to potent cell growth inhibition in human melanoma by derepression of tumor suppressor genes. Oncotarget 2015, 6, 27023. [Google Scholar] [CrossRef] [Green Version]
- Hippo, Y.; Taniguchi, H.; Tsutsumi, S.; Machida, N.; Chong, J.M.; Fukayama, M.; Kodama, T.; Aburatani, H. Global Gene Expression Analysis of Gastric Cancer by Oligonucleotide Microarrays. Cancer Res. 2002, 62, 233–240. [Google Scholar]
- Li, F.; Yoshizawa, J.M.; Kim, K.M.; Kanjanapangka, J.; Grogan, T.R.; Wang, X.; Elashoff, D.E.; Ishikawa, S.; Chia, D.; Liao, W.; et al. Discovery and Validation of Salivary Extracellular RNA Biomarkers for Noninvasive Detection of Gastric Cancer. Clin. Chem. 2018, 64, 1513–1521. [Google Scholar] [CrossRef] [Green Version]
- Meephansan, J.; Ungpraphakorn, N.; Ponnikorn, S.; Suchonwanit, P.; Poovorawan, Y. Efficacy of 1,550-nm Erbium-Glass Fractional Laser Treatment and Its Effect on the Expression of Insulin-Like Growth Factor 1 and Wnt/beta-Catenin in Androgenetic Alopecia. Dermatol. Surg. 2018, 44, 1295–1303. [Google Scholar] [CrossRef]
- Gu, Z.; Tan, W.; Feng, G.; Meng, Y.; Shen, B.; Liu, H.; Cheng, C. Wnt/beta-catenin signaling mediates the senescence of bone marrow-mesenchymal stem cells from systemic lupus erythematosus patients through the p53/p21 pathway. Mol. Cell. Biochem. 2014, 387, 27–37. [Google Scholar] [CrossRef]
- Jamali, A.E.; Freund, C.; Rechner, C.; Scheidereit, C.; Dietz, R.; Bergmann, M.W. Reoxygenation after severe hypoxia induces cardiomyocyte hypertrophy in vitro activation of CREB downstream of GSK3β. FASEB J. 2004, 18, 1096–1098. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Hou, G.; Yang, J.; Chen, W.; Guo, L.; Mao, Q.; Ge, J.; Zhang, X. SOX9-activated PXN-AS1 promotes the tumorigenesis of glioblastoma by EZH2-mediated methylation of DKK1. J. Cell. Mol. Med. 2020, 24, 6070–6082. [Google Scholar] [CrossRef] [Green Version]
- Vire, E.; Brenner, C.; Deplus, R.; Blanchon, L.; Fraga, M.; Didelot, C.; Morey, L.; Van Eynde, A.; Bernard, D.; Vanderwinden, J.M.; et al. The Polycomb group protein EZH2 directly controls DNA methylation. Nature 2006, 439, 871–874. [Google Scholar] [CrossRef]
- Lujambio, A.; Portela, A.; Liz, J.; Melo, S.A.; Rossi, S.; Spizzo, R.; Croce, C.M.; Calin, G.A.; Esteller, M. CpG island hypermethylation-associated silencing of non-coding RNAs transcribed from ultraconserved regions in human cancer. Oncogene 2010, 29, 6390–6401. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.C.; Shen, J.; Ran, Z.H. Transcribed ultraconserved region in human cancers. RNA Biol. 2013, 10, 1771–1777. [Google Scholar] [CrossRef] [Green Version]
- Huarte, M. The emerging role of lncRNAs in cancer. Nat. Med. 2015, 21, 1253–1261. [Google Scholar] [CrossRef]
- Yang, Z.; Guo, X.; Li, G.; Shi, Y.; Li, L. Long noncoding RNAs as potential biomarkers in gastric cancer: Opportunities and challenges. Cancer Lett. 2016, 371, 62–70. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, J.-h.; Byun, H.; Ivan, C.; Calin, G.A.; Jung, D.; Lee, S. lncRNAs UC.145 and PRKG1-AS1 Determine the Functional Output of DKK1 in Regulating the Wnt Signaling Pathway in Gastric Cancer. Cancers 2022, 14, 2369. https://doi.org/10.3390/cancers14102369
Yoon J-h, Byun H, Ivan C, Calin GA, Jung D, Lee S. lncRNAs UC.145 and PRKG1-AS1 Determine the Functional Output of DKK1 in Regulating the Wnt Signaling Pathway in Gastric Cancer. Cancers. 2022; 14(10):2369. https://doi.org/10.3390/cancers14102369
Chicago/Turabian StyleYoon, Jung-ho, Hyojoo Byun, Cristina Ivan, George A. Calin, Dahyun Jung, and Sangkil Lee. 2022. "lncRNAs UC.145 and PRKG1-AS1 Determine the Functional Output of DKK1 in Regulating the Wnt Signaling Pathway in Gastric Cancer" Cancers 14, no. 10: 2369. https://doi.org/10.3390/cancers14102369