
Dissecting Tumor-Immune Microenvironment in Breast Cancer at a Spatial and Multiplex Resolution


 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
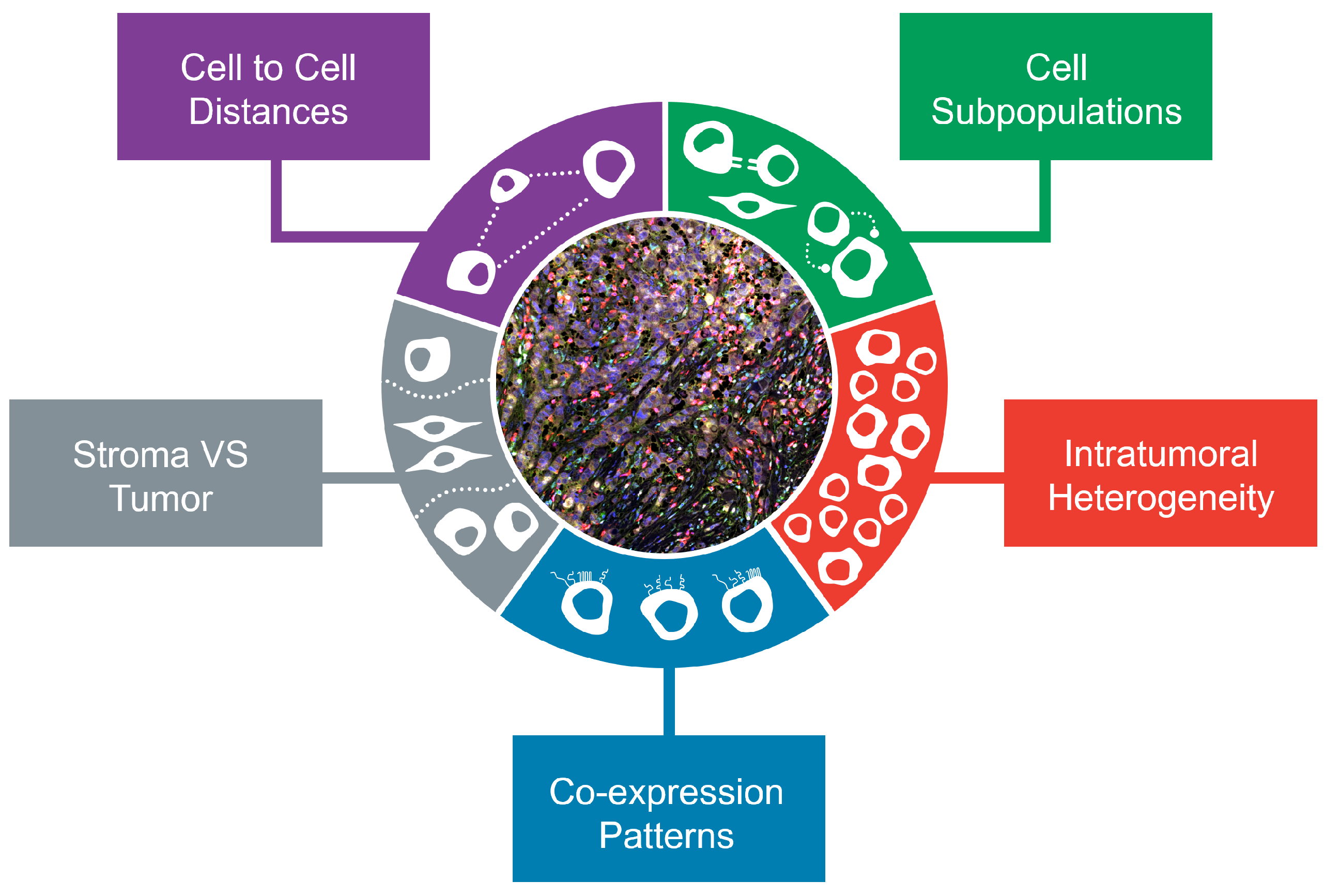
2. Multiplex In Situ Methods: A Plethora of Assays Platforms and Analysis Tools
2.1. Antibody Conjugated with Chromogen
2.1.1. Discovery Ultra
2.1.2. Multiplexed Immunohistochemical Consecutive Staining on Single Slide (MICSSS)
2.2. Antibody Conjugated with Fluorophore

{kind=link}
| Assay | D.Ultra [32] | MICSSS [33] | Vectra/Polaris [35] | Opal [39] | CODEX [41,42] | InSituPlex [43] | GeoMx DSP [44,45] | MIBI-TOF [46,47] | Cy-TOF [48] |
|---|---|---|---|---|---|---|---|---|---|
| Vendor | Roche | Remark et al. | Akoya Biosciences | Roche and Akoya Biosciences | Akoya Biosciences | Ultivue | Nanostring | IonPath | Bodenmiller et al. |
| Antibody Conjugate | chromogen | chromogen | fluorescent | Opal fluorescent | DNA barcode | DNA barcode | DNA barcode | Metal isotope | Metal isotope |
| Tissue disruption | no | no | no | no | no | no | no | Yes (adjustable) | yes |
| Biomarkers | 5 | 10+ | ∼9 | ∼8 | 40+ | ∼5 | 70+ | 30+ | 30+ |
| Staining cycles | Iterative | Iterative | Iterative | Iterative | Iterative | 1 | 1 | 1 | 1 |
| Scanning | camera | camera | Vectra Polaris | Vectra Polaris | CODEX fluidics | compatible with various systems | GeoMx | TOF spectrometer | TOF spectrometer |
| Profiling area | Whole slide | Whole slide | adjustable (660 m → whole slide) | adjustable (660 μm → whole slide) | 660 m | NA | adjustable (280 μm → 800 m) | 800 m | 1 mm |
| Software | NA | NA | InForm | InForm QuPath | CODEX analysis manager & viewer | compatible with various systems | GeoMx data analysis suite | MIBIAnalysis | HistoCat, Cell Profiler |
| Resolution | Human vision | Human vision | 0.25–0.9 m | 0.25–0.9 m | 0.26 m | NA | 10 m | 0.26 m | 1 m |
| Spatial information | yes | yes | yes | yes | yes | yes | limited | yes | yes |
| Single-cell information | yes | yes | yes | yes | yes | yes | limited | yes | yes |
| Disadvantages | Human Vision Limits, Semi- Quantitative | time- consuming, coverslip removal errors | costly, limited marker numbers | costly, limited marker numbers | costly, time- consuming, difficult cell segmentation, difficult panel validation | limited marker numbers | costly, limited spatial information, limited single cell information | special training, costly, time- consuming, difficult cell segmentation, difficult panel validation | special training, costly, time- consuming, difficult cell segmentation, difficult panel validation |
2.3. Antibody Conjugated with DNA-Barcode
2.3.1. CO-Detection by indEXing (CODEX®) & InSituPlex®
2.3.2. GeoMx® Digital Spatial Profiling
2.4. Antibody Conjugated with Metal Isotope: Mass Spectrometry Immunohistochemistry (MS-IHC)
Imaging Mass Cytometry with Time of Flight (Cy-TOF) and Multiplex Ion Beam Imaging with Time of Flight (MIBI-TOF)
2.5. Image Analysis Software for Multiplex Profiling Assays
3. Applications of Multiplex Methods in Breast Cancer: TIME Composition, Spatiotemporal Heterogeneity and Prognostic Implications
3.1. Overview of Multiplex In Situ Evaluation of Immune Response Heterogeneity in BC
3.2. Spatial Heterogeneity of Breast Cancer TIME
3.2.1. Intratumoral Spatial Heterogeneity: Regions of Interest, Tissue Compartments, Cell-to-Cell Distances
3.2.2. Spatial Heterogeneity of Immune Regulatory Proteins: The PD-L1 Paradigm
| Author | Journal | Year | Tissue Type | Disease Setting | Treatment | BC Type | Pts No | Assay | Panel | Scanning | Software | ROIs Number, Area (per Patient) | Tissue Segmentation | Prognosis/ Prediction | Spatial Heterogeneity |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fluorescent Multiplex Immunohistochemistry | |||||||||||||||
| Angelis [62] | CCR | 2019 | WTS | Neo-adjuvant | Lapatinib trastuzumab | HER2+ BC | 29 | mIF | CD4, CD8, CD20, FoxP3, CD68, CK, DAPI | Vectra | InForm | 5, 2 mm | (CK+/DAPI+) (CK-/DAPI+) | Higher Stromal CD4, Tumoral CD4,CD20: Higher pCR | CD8 enriched In tumoral, CD4, CD8, CD20 Positive correlation |
| Brown [80] | CCR | 2014 | WTS | Neo-adjuvant | Taxane, anthracycline trastuzumab | all | 87 | mIF | CD3, CD8, CD20, CK, DAPI | PM-2000 hardware (HistoRx) | AQUA | At least 3 CK enriched | (CD3+/DAPI+) (CD3-/DAPI-) | Higher stromal CD4, CD8, CD20: Higher pCR | TNBC Higher CD20, CD8 densities |
| Graeser [61] | JIC | 2020 | WTS | Neo-adjuvant | Paclitaxel, gemcitabine, carboplatin | TNBC | 66 | mIF | CD4, CD8, CD73, PD1, PD-L1, CK7, DAPI | Vectra/Polaris | InForm | Whole slide | (CK7+/DAPI+) (CK7-/DAPI+) | On treatment CD4+PD1+: Higher pCR | NA |
| Griguolo [60] | Npj precision oncology | 2021 | WTS | Neo-adjuvant | Lapatinib, trastuzumab | HER2+ BC | 65 | MCISSS | CD3, CD4, CD8 Foxp3, Ki67, panCK, hematoxylin | NanoZoomer 2.0HT (Hamamatsu Photonics, Japan) | VISIOPHARM | Whole slide | (CK+/HTX+) (CK-/HTX-) | On treatment TumoralCD8+ higher pCR | Ki67+CD3+ Higher densities closer to cancer cells, enriched in HR- |
| Kearney [63] | SABCS | 2021 | WTS | Neo-adjuvant | Anthracycline Trastuzumab pertuzumab | HR+/ HER2+ BC | 28 | mIF | CD3, CD8, CD68, FOXP3, Pan-CK, DAPI | NA | HALO | NA | CK+ CK- | Higher CD3+CD8-FoxP3- higher pCR | NA |
| Yam [71] | CCR | 2021 | WTS | Neo-adjuvant | doxorubicin, cyclo-phosphamide, paclitaxel | TNBC | 102 | mIF | PDL1, PD1, CD3, CD68, Pan-CK, DAPI | Vectra 3.0 | InForm | NA | CK+ CK- | Higher CD3+/CD68+ ratio, CD3-cancer proximity higher pCR | PD-L1 expression: tumor (more common) and stromal compartments, CD3+ PD1+ rare population |
| Janiszewska [83] | JCIinsight | 2021 | WTS | Neo-adjuvant | Trastuzumab emtansine, pertuzumab | HER2+ BC | 20 | mIF | 20 | NA | NA | NA | CK+ CK- | pCR: higher CD8, RD:GZM+ mf closer to cancer, immune cells less proximal to vimentin+ cells | NA |
| Egelston [67] | JCIinsight | 2019 | WTS | Adjuvant | doxorubicin, cyclophospamide hamide, paclitaxel | TNBC | 25 | mIF | CD8, CD103, CD69, pan-CK, DAPI | Vectra 3.0 | InForm | multiple | (CK+/DAPI+) (CK-/DAPI+) | Higher Tumoral CD8+CD103+: Better RFS | CD8+CD103+ Enriched closer to cancer cells |
| Millar [84] | Cancers | 2020 | TMA | Adjuvant | CMF, anthracycline | all | 485 | mIF | CD3, CD8, CD20, CD68, Fox P3, PD-1, PD-L1, PanCK, DAPI | Vectra/Polaris | InForm | 1, 780 μm | (CK+/DAPI+) (CK-/DAPI+) | Combined Stromal lower CD8, CD20: Shorter OS | NA |
| Garaud [79] | JCIinsight | 2019 | WTS | Adjuvant | Chemotherapy | HER2+ BC, TNBC | 249 | mIF | CD4, CD8, CD20, FoxP3, CD68, CK, DAPI | Vectra/Polaris | InForm | NA | NA | Higher CD20: Favorable prognosis | Description of Lymphocytic aggregates |
| Costa [81] | Cancer Cell | 2018 | NA | NA | NA | TNBC | NA | mIF | CD25, FAP, PDL2, OX40L, DAPI | HistoFluor microscope | manual | NA | NA | NA | FAP+PDL2+ OX40L+: T cell exhaustion |
| Wortman [73] | Npj Breast Cancer | 2021 | WTS | Adjuvant | Chemotherapy | TNBC | 36 | mIF | CD3, CD4, CD8, FOXP3, CD20, DAPI, PanCK | Vectra 3.0 | InForm | NA | NA | Higher tumoral Distribution CD3,CD20: Better RFS | CD3, CD20: Description of Aggregates |
| Mani [72] | Breast cancer research | 2016 | WTS | NA | Surgery | NA | 31 | mIF | CK, DAPI, CD3, CD8, CD20 | NA | AQUA | 6–50 | NA | NA | Approximately 70% of spatial Heterogeneity of B cells: Within same tissue section |
| O’Meara [85] | SABCS | 2021 | NA | NA | NA | all | 132 | mIF | CD8, FoxP3, PD1, PDL1, CK | Vectra | NA | NA | NA | NA | HR+ Immune mark.: Correlates with Grade, Recurrence score |
| Shimada [86] | SABCS | 2021 | NA | NA | NA | HR+ BC | 5 | mIF | 9-21 proteins | CyteFinder microscope | MCMICRO | NA | Tumor Stroma Immune cells | NA | Description Of cold, hot Excluded TIME |
| Noel [66] | JCI | 2021 | WTS | Adjuvant | Chemotherapy, HER2 targeted | HER2+ BC, TNBC | 48 | mIF | CD4, CD20, PD-1, ICOS, Ki-67, Foxp3 | Vectra/Polaris | NA | NA | NA | Active TLS: Favorable prognosis | Active TLS: Combined CD4+PD1+, CD20+ki67+, GZMB+CD8+ |
| Bedard [68] | SABCS | 2021 | NA | Metastatic | INT230-6 (cisplatin and vinblastine) pembrolizumab | TNBC | 3 | mIF | CD4, CD8, FoxP3 | NA | NA | NA | NA | NA | Metastasis Lower CD4,CD8 Higher FoxP3 |
| Zhu [87] | J.Immun. Cancer | 2019 | WTS | Metastatic | NA | all | 5 | mIF | CD8, FoxP3, CD68, CD20, CK, DAPI | NA | NA | NA | NA | NA | Metastasis Higher CD68 |
| He [69] | Plos One | 2020 | WTS | Metastatic | Chemotherapy | TNBC | 10 | mIF | CD4, CD8, FOXP3, CD20, CD33, PD1, CK, DAPI | Vectra | InForm | NA | CK+ CK- | Higher PD1+CD8+ PD1+CD4+ Better outcome | Metastasis Lower Str. CD8+, PD1+CD8+ |
| Ultivue InSituPlex | |||||||||||||||
| Ahmed [70] | CCR | 2020 | WTS | Neo-adjuvant | nab-paclitaxel, doxorubicin, cyclophosphamide, durvalumab | TNBC | 45 | InSitu Plex | CD8,CD68,PDL1, CK, Sox10, Hoechst | PM2000 microscope (HistoRx) | AQUA | NA | CK+, CK- CD68-, CK- CD68+ | pCR vs RD: higher baseline PDL1 expression and similar CD68 densities | PD-L1 expression: on both stromal and tumoral compartments |
| GeoMx Digital Spatial Profiling | |||||||||||||||
| Carter [75] | SABCS | 2020 | WTS | Neo-adjuvant | taxane | HR+/ HER2- BC | 39 | DSP | 58 proteins | GeoMx | NA | 6, 600 m | (CK+/SYTO13+) (CK-/SYTO13+) | After NAT: LAG3 expression: Residual disease | Most immune Markers: Stromal enrichment |
| McNamara [77] | Nature Cancer | 2018 | WTS | Neo-adjuvant | Trastuzumab lapatinib | HER2+ BC | 28 | DSP | 40 proteins | GeoMx | NA | 4, 464–666 m | CK+ CK- | On treatment Expression CD45, CD56 Higher pCR | Compartment enrichment Tumoral CD56, B7H4 PDL1, IDO Stromal CD3, CD4, CD8, CD68 |
| Carter [76] | SABCS | 2020 | TMA | adjuvant | Chemotherapy | TNBC | 167 | DSP | 58 proteins | GeoMx | NA | 1, 600 m | (CK+/SYTO13+) (CK-/SYTO13+) | Stromal LAG3 Higher PFS | Compartment Enrichment Tumoral CD27, HLADR, IDO1 PDL1 Stromal PDL1 |
| Stewart [74] | Scientific reports | 2020 | WTS | Adjuvant | Chemotherapy | TNBC | 10 | DSP | 39 proteins | GeoMx | NA | 6, 300 m | CK+ CK- | No relapse: Tumoral HLA-DR, IDO1, B2M Stromal CD45, CD4, PD-1 | Most immune Markers Stromal |
| Kulasinghe [88] | Frontiers In Oncology | 2022 | TMA | Adjuvant | Chemotherapy | TNBC | 24 | DSP | 68 proteins | GeoMx | NA | 1, NA | CK+ CK- | Responders Tumoral higher GZMA, STING, Fibronectin Lower CD80 Stromal Lower 4-1BB | NA |
| Carter [89] | CCR | 2021 | TMA | NA | Chemotherapy untreated | TNBC | 184 | DSP | 58 proteins | GeoMx | GeoMx software | 1, 600 m | CK+, CK-/SYTO13+ | NA | PD-L1+: immunologically hotter (mf, T cells, checkpoints (IDO-1), antigen presentation, STING) |
| Leon-Ferre [90] | SABCS | 2021 | TMA | NA | Surgery | TNBC | 111 | DSP | 58 proteins | GeoMx | NA | 1, NA | CK+ CK- | NA | LAR vs non-LAR TNBC Lower Stromal CD45, CD14, IDO1 Tumoral CD45, B7H3 |
| Schlam [78] | JTM | 2021 | WTS | Metastatic | Chemotherapy HER2 targeted | HER2+ BC | 8 | DSP | 70 proteins | GeoMx | NA | 2 | CK+ CK- | NA | Metastasis Lower CD3, CD8 Tim-3, CD27, 4-1BB |
| Mass Spectrometry Immunohistochemistry | |||||||||||||||
| Bianchini [91] | SABCS | 2021 | WTS | Neo-adjuvant | atezolizumab, carboplatin, nab-paclitaxel | TNBC | 243 | IMC | 43 proteins | TOF | NA | NA | NA | Higher pCR: density APC PDL1+IDO+, spatial correlation of CK+ and CD8+PD1+, CD8+GZMB+, CD20+ | NA |
| Keren [28] | Cell | 2018 | WTS | NA | Surgery | TNBC | 41 | MIBI- TOF | 36 proteins | TOF | NA | 1, 800 μm | NA | Compartmentalized pattern longer OS | Hot tumors Were divided Into “Mixed”, “Compartmentalized” |
3.2.3. Inter-Patient Spatial Heterogeneity: Characterization of Spatial Organization of BC TIME
3.3. Temporal Heterogeneity of Breast Cancer TIME
3.3.1. Longitudinal Evaluation under Treatment Pressure at the Neoadjuvant BC Setting
3.3.2. Intra-Patient Heterogeneity: From Primary Breast Cancer to Metastatic Progression
3.4. Prognostic and Predictive Implications of Multiplexed Spatial Profiling in Breast Cancer
3.5. AI-Assisted Evaluation of TIME on H&E Breast Cancer Tissue Images
4. Discussion
5. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassilakopoulos, T.P.; Chatzidimitriou, C.; Asimakopoulos, J.V.; Arapaki, M.; Tzoras, E.; Angelopoulou, M.K.; Konstantopoulos, K. Immunotherapy in Hodgkin Lymphoma: Present Status and Future Strategies. Cancers 2019, 11, 1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocco, D.; Della Gravara, L.; Battiloro, C.; Gridelli, C. The role of combination chemo-immunotherapy in advanced non-small cell lung cancer. Expert Rev. Anticancer Ther. 2019, 19, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Carlino, M.S.; Larkin, J.; Long, G.V. Immune checkpoint inhibitors in melanoma. Lancet 2021, 398, 1002–1014. [Google Scholar] [CrossRef]
- André, T.; Shiu, K.K.; Kim, T.W.; Jensen, B.V.; Jensen, L.H.; Punt, C.; Smith, D.; Garcia-Carbonero, R.; Benavides, M.; Gibbs, P. Pembrolizumab in microsatellite-instability-high advanced colorectal cancer. N. Engl. J. Med. 2020, 383, 2207–2218. [Google Scholar] [CrossRef]
- Postow, M.A.; Sidlow, R.; Hellmann, M.D. Immune-related adverse events associated with immune checkpoint blockade. N. Engl. J. Med. 2018, 378, 158–168. [Google Scholar] [CrossRef]
- Schmid, P.; Adams, S.; Rugo, H.S.; Schneeweiss, A.; Barrios, C.H.; Iwata, H.; Diéras, V.; Hegg, R.; Im, S.A.; Shaw Wright, G. Atezolizumab and nab-paclitaxel in advanced triple-negative breast cancer. N. Engl. J. Med. 2018, 379, 2108–2121. [Google Scholar] [CrossRef]
- Miles, D.; Gligorov, J.; André, F.; Cameron, D.; Schneeweiss, A.; Barrios, C.; Xu, B.; Wardley, A.M.; Kaen, D.; Andrade, L. Primary results from IMpassion131, a double-blind placebo-controlled randomised phase III trial of first-line paclitaxel (PAC)+/-atezolizumab (atezo) for unresectable locally advanced/metastatic triple-negative breast cancer (mTNBC). Ann. Oncol. 2021, 32, 994–1004. [Google Scholar] [CrossRef]
- Cortes, J.; Cescon, D.W.; Rugo, H.S.; Nowecki, Z.; Im, S.A.; Yusof, M.M.; Gallardo, C.; Lipatov, O.; Barrios, C.H.; Holgado, E. Pembrolizumab plus chemotherapy versus placebo plus chemotherapy for previously untreated locally recurrent inoperable or metastatic triple-negative breast cancer (KEYNOTE-355): A randomised, placebo-controlled, double-blind, phase 3 clinical trial. Lancet 2020, 396, 1817–1828. [Google Scholar] [CrossRef]
- Schmid, P.; Cortes, J.; Pusztai, L.; McArthur, H.; Kümmel, S.; Bergh, J.; Denkert, C.; Park, Y.H.; Hui, R.; Harbeck, N. Pembrolizumab for early triple-negative breast cancer. N. Engl. J. Med. 2020, 382, 810–821. [Google Scholar] [CrossRef]
- Mittendorf, E.A.; Zhang, H.; Barrios, C.H.; Saji, S.; Jung, K.H.; Hegg, R.; Koehler, A.; Sohn, J.; Iwata, H.; Telli, M.L. Neoadjuvant atezolizumab in combination with sequential nab-paclitaxel and anthracycline-based chemotherapy versus placebo and chemotherapy in patients with early-stage triple-negative breast cancer (IMpassion031): A randomised, double-blind, phase 3 trial. Lancet 2020, 396, 1090–1100. [Google Scholar] [CrossRef]
- Loi, S.; Sirtaine, N.; Piette, F.; Salgado, R.; Viale, G.; Van Eenoo, F.; Rouas, G.; Francis, P.; Crown, J.; Hitre, E. Prognostic and predictive value of tumor-infiltrating lymphocytes in a phase III randomized adjuvant breast cancer trial in node-positive breast cancer comparing the addition of docetaxel to doxorubicin with doxorubicin-based chemotherapy: BIG 02-98. J. Clin. Oncol. 2013, 31, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Foukakis, T.; Lövrot, J.; Matikas, A.; Zerdes, I.; Lorent, J.; Tobin, N.; Suzuki, C.; Brage, S.E.; Carlsson, L.; Einbeigi, Z. Immune gene expression and response to chemotherapy in advanced breast cancer. Br. J. Cancer 2018, 118, 480–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matikas, A.; Lövrot, J.; Ramberg, A.; Eriksson, M.; Lindsten, T.; Lekberg, T.; Hedenfalk, I.; Loman, N.; Bergh, J.; Hatschek, T. Dynamic evaluation of the immune infiltrate and immune function genes as predictive markers for neoadjuvant chemotherapy in hormone receptor positive, HER2 negative breast cancer. Oncoimmunology 2018, 7, e1466017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denkert, C.; von Minckwitz, G.; Darb-Esfahani, S.; Lederer, B.; Heppner, B.I.; Weber, K.E.; Budczies, J.; Huober, J.; Klauschen, F.; Furlanetto, J. Tumour-infiltrating lymphocytes and prognosis in different subtypes of breast cancer: A pooled analysis of 3771 patients treated with neoadjuvant therapy. Lancet Oncol. 2018, 19, 40–50. [Google Scholar] [CrossRef]
- Hofman, P.; Badoual, C.; Henderson, F.; Berland, L.; Hamila, M.; Long-Mira, E.; Lassalle, S.; Roussel, H.; Hofman, V.; Tartour, E. Multiplexed immunohistochemistry for molecular and immune profiling in lung cancer - Just about ready for prime-time? Cancers 2019, 11, 283. [Google Scholar] [CrossRef] [Green Version]
- Cristescu, R.; Mogg, R.; Ayers, M.; Albright, A.; Murphy, E.; Yearley, J.; Sher, X.; Liu, X.Q.; Lu, H.; Nebozhyn, M. Pan-tumor genomic biomarkers for PD-1 checkpoint blockade-based immunotherapy. Science 2018, 362, eaar3593. [Google Scholar] [CrossRef] [Green Version]
- Zerdes, I.; Karafousia, V.; Mezheyeuski, A.; Stogiannitsi, M.; Kuiper, R.; Moreno Ruiz, P.; Rassidakis, G.; Bergh, J.; Hatschek, T.; Foukakis, T. Discordance of PD-L1 Expression at the Protein and RNA Levels in Early Breast Cancer. Cancers 2021, 13, 4655. [Google Scholar] [CrossRef]
- Chrétien, S.; Zerdes, I.; Bergh, J.; Matikas, A.; Foukakis, T. Beyond PD-1/PD-L1 inhibition: What the future holds for breast cancer immunotherapy. Cancers 2019, 11, 628. [Google Scholar] [CrossRef] [Green Version]
- Reticker-Flynn, N.E.; Engleman, E.G. Cancer systems immunology. eLife 2020, 9, e53839. [Google Scholar] [CrossRef]
- Salgado, R.; Denkert, C.; Demaria, S.; Sirtaine, N.; Klauschen, F.; Pruneri, G.; Wienert, S.; Van den Eynden, G.; Baehner, F.L.; Pénault-Llorca, F. The evaluation of tumor-infiltrating lymphocytes (TILs) in breast cancer: Recommendations by an International TILs Working Group 2014. Ann. Oncol. 2015, 26, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Dieci, M.V.; Radosevic-Robin, N.; Fineberg, S.; Van den Eynden, G.; Ternes, N.; Penault-Llorca, F.; Pruneri, G.; D’Alfonso, T.M.; Demaria, S.; Castaneda, C. Update on tumor-infiltrating lymphocytes (TILs) in breast cancer, including recommendations to assess TILs in residual disease after neoadjuvant therapy and in carcinoma in situ: A report of the International Immuno-Oncology Biomarker Working Group on Breast Cancer. Semin. Cancer Biol. 2018, 52, 16–25. [Google Scholar] [PubMed]
- Savas, P.; Salgado, R.; Denkert, C.; Sotiriou, C.; Darcy, P.K.; Smyth, M.J.; Loi, S. Clinical relevance of host immunity in breast cancer: From TILs to the clinic. Nat. Rev. Clin. Oncol. 2016, 13, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Kos, Z.; Roblin, E.; Kim, R.S.; Michiels, S.; Gallas, B.D.; Chen, W.; van de Vijver, K.K.; Goel, S.; Adams, S.; Demaria, S. Pitfalls in assessing stromal tumor infiltrating lymphocytes (sTILs) in breast cancer. npj Breast Cancer 2020, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.S.; Mellman, I. Elements of cancer immunity and the cancer-immune set point. Nature 2017, 541, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Provenzano, E.; Dawson, S.J.; Blows, F.; Liu, B.; Shah, M.; Earl, H.; Poole, C.; Hiller, L.; Dunn, J. Association between CD8+ T-cell infiltration and breast cancer survival in 12 439 patients. Ann. Oncol. 2014, 25, 1536–1543. [Google Scholar] [CrossRef] [PubMed]
- Berry, S.; Giraldo, N.A.; Green, B.F.; Cottrell, T.R.; Stein, J.E.; Engle, E.L.; Xu, H.; Ogurtsova, A.; Roberts, C.; Wang, D. Analysis of multispectral imaging with the AstroPath platform informs efficacy of PD-1 blockade. Science 2021, 372, eaba2609. [Google Scholar] [CrossRef]
- Keren, L.; Bosse, M.; Marquez, D.; Angoshtari, R.; Jain, S.; Varma, S.; Yang, S.R.; Kurian, A.; Van Valen, D.; West, R. A structured tumor-immune microenvironment in triple negative breast cancer revealed by multiplexed ion beam imaging. Cell 2018, 174, 1373–1387.e19. [Google Scholar] [CrossRef] [Green Version]
- Gruosso, T.; Gigoux, M.; Manem, V.S.K.; Bertos, N.; Zuo, D.; Perlitch, I.; Saleh, S.M.I.; Zhao, H.; Souleimanova, M.; Johnson, R.M. Spatially distinct tumor immune microenvironments stratify triple-negative breast cancers. J. Clin. Investig. 2019, 129, 1785–1800. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Gao, R.; Sei, E.; Brandt, R.; Hartman, J.; Hatschek, T.; Crosetto, N.; Foukakis, T.; Navin, N.E. Chemoresistance evolution in triple-negative breast cancer delineated by single-cell sequencing. Cell 2018, 173, 879–893. e13. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Stein, J.E.; Rimm, D.L.; Wang, D.W.; Bell, J.M.; Johnson, D.B.; Sosman, J.A.; Schalper, K.A.; Anders, R.A.; Wang, H. Comparison of biomarker modalities for predicting response to PD-1/PD-L1 checkpoint blockade: A systematic review and meta-analysis. JAMA Oncol. 2019, 5, 1195–1204. [Google Scholar] [CrossRef] [PubMed]
- Morrison, L.E.; Lefever, M.R.; Behman, L.J.; Leibold, T.; Roberts, E.A.; Horchner, U.B.; Bauer, D.R. Brightfield multiplex immunohistochemistry with multispectral imaging. Lab. Investig. 2020, 100, 1124–1136. [Google Scholar] [CrossRef] [PubMed]
- Remark, R.; Merghoub, T.; Grabe, N.; Litjens, G.; Damotte, D.; Wolchok, J.D.; Merad, M.; Gnjatic, S. In-depth tissue profiling using multiplexed immunohistochemical consecutive staining on single slide. Sci. Immunol. 2016, 1, aaf6925. [Google Scholar] [CrossRef] [PubMed]
- Loos, C.M.v.d. Multiple immunoenzyme staining: Methods and visualizations for the observation with spectral imaging. J. Histochem. Cytochem. 2008, 56, 313–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Z.; Puri, S.; Moudgil, T.; Wood, W.; Hoyt, C.C.; Wang, C.; Urba, W.J.; Curti, B.D.; Bifulco, C.B.; Fox, B.A. Multispectral imaging of formalin-fixed tissue predicts ability to generate tumor-infiltrating lymphocytes from melanoma. J. Immunother. Cancer 2015, 3, 47. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.R.; Izar, B.; Wang, S.; Yapp, C.; Mei, S.; Shah, P.M.; Santagata, S.; Sorger, P.K. Highly multiplexed immunofluorescence imaging of human tissues and tumors using t-CyCIF and conventional optical microscopes. eLife 2018, 7, e31657. [Google Scholar] [CrossRef]
- Gerdes, M.J.; Sevinsky, C.J.; Sood, A.; Adak, S.; Bello, M.O.; Bordwell, A.; Can, A.; Corwin, A.; Dinn, S.; Filkins, R.J. Highly multiplexed single-cell analysis of formalin-fixed, paraffin-embedded cancer tissue. Proc. Natl. Acad. Sci. USA 2013, 110, 11982–11987. [Google Scholar] [CrossRef] [Green Version]
- Taube, J.M.; Akturk, G.; Angelo, M.; Engle, E.L.; Gnjatic, S.; Greenbaum, S.; Greenwald, N.F.; Hedvat, C.V.; Hollmann, T.J.; Juco, J. The Society for Immunotherapy in Cancer statement on best practices for multiplex immunohistochemistry (IHC) and immunofluorescence (IF) staining and validation. J. Immunother. Cancer 2020, 8, e000155. [Google Scholar] [CrossRef]
- Viratham Pulsawatdi, A.; Craig, S.G.; Bingham, V.; McCombe, K.; Humphries, M.P.; Senevirathne, S.; Richman, S.D.; Quirke, P.; Campo, L.; Domingo, E.; et al. A robust multiplex immunofluorescence and digital pathology workflow for the characterisation of the tumour immune microenvironment. Mol. Oncol. 2020, 14, 2384–2402. [Google Scholar] [CrossRef]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef] [Green Version]
- Goltsev, Y.; Samusik, N.; Kennedy-Darling, J.; Bhate, S.; Hale, M.; Vazquez, G.; Black, S.; Nolan, G.P. Deep profiling of mouse splenic architecture with CODEX multiplexed imaging. Cell 2018, 174, 968–981.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schürch, C.M.; Bhate, S.S.; Barlow, G.L.; Phillips, D.J.; Noti, L.; Zlobec, I.; Chu, P.; Black, S.; Demeter, J.; McIlwain, D.R. Coordinated cellular neighborhoods orchestrate antitumoral immunity at the colorectal cancer invasive front. Cell 2020, 182, 1341–1359.e19. [Google Scholar] [CrossRef] [PubMed]
- Manesse, M.; Patel, K.K.; Bobrow, M.; Downing, S.R. The InSituPlex® staining method for multiplexed immunofluorescence cell phenotyping and spatial profiling of tumor FFPE samples. Methods Mol. Biol. 2020, 2055, 585–592. [Google Scholar] [PubMed]
- McCart Reed, A.E.; Bennett, J.; Kutasovic, J.R.; Kalaw, E.; Ferguson, K.; Yeong, J.; Simpson, P.T.; Lakhani, S.R. Digital spatial profiling application in breast cancer: A user’s perspective. Virchows Arch. 2020, 477, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Decalf, J.; Albert, M.L.; Ziai, J. New tools for pathology: A user’s review of a highly multiplexed method for in situ analysis of protein and RNA expression in tissue. J. Pathol. 2019, 247, 650–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelo, M.; Bendall, S.C.; Finck, R.; Hale, M.B.; Hitzman, C.; Borowsky, A.D.; Levenson, R.M.; Lowe, J.B.; Liu, S.D.; Zhao, S. Multiplexed ion beam imaging of human breast tumors. Nat. Med. 2014, 20, 436–442. [Google Scholar] [CrossRef] [Green Version]
- Keren, L.; Bosse, M.; Thompson, S.; Risom, T.; Vijayaragavan, K.; McCaffrey, E.; Marquez, D.; Angoshtari, R.; Greenwald, N.F.; Fienberg, H. MIBI-TOF: A multiplexed imaging platform relates cellular phenotypes and tissue structure. Sci. Adv. 2019, 5, eaax5851. [Google Scholar] [CrossRef] [Green Version]
- Giesen, C.; Wang, H.A.; Schapiro, D.; Zivanovic, N.; Jacobs, A.; Hattendorf, B.; Schüffler, P.J.; Grolimund, D.; Buhmann, J.M.; Brandt, S. Highly multiplexed imaging of tumor tissues with subcellular resolution by mass cytometry. Nat. Methods 2014, 11, 417–422. [Google Scholar] [CrossRef]
- Du, Z.; Lin, J.R.; Rashid, R.; Maliga, Z.; Wang, S.; Aster, J.C.; Izar, B.; Sorger, P.K.; Santagata, S. Qualifying antibodies for image-based immune profiling and multiplexed tissue imaging. Nat. Protoc. 2019, 14, 2900–2930. [Google Scholar] [CrossRef]
- Nairn, R. Standardization in immunofluorescence. Clin. Exp. Immunol. 1968, 3, 465. [Google Scholar]
- Mansfield, J. Multispectral imaging: A review of its technical aspects and applications in anatomic pathology. Vet. Pathol. 2014, 51, 185–210. [Google Scholar] [CrossRef] [PubMed]
- Bendall, S.C.; Simonds, E.F.; Qiu, P.; El-ad, D.A.; Krutzik, P.O.; Finck, R.; Bruggner, R.V.; Melamed, R.; Trejo, A.; Ornatsky, O.I. Single-cell mass cytometry of differential immune and drug responses across a human hematopoietic continuum. Science 2011, 332, 687–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, H.; Bolen, J.; Schuetter, L.; Massion, P.; Hoyt, C.C.; VandenBerg, S.; Esserman, L.; Borowsky, A.D.; Campbell, M.J. Characterizing the Tumor Immune Microenvironment with Tyramide-Based Multiplex Immunofluorescence. J. Mammary Gland Biol. Neoplasia 2020, 25, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Yaghoobi, V.; Martinez-Morilla, S.; Liu, Y.; Charette, L.; Rimm, D.L.; Harigopal, M. Advances in quantitative immunohistochemistry and their contribution to breast cancer. Expert Rev. Mol. Diagn. 2020, 20, 509–522. [Google Scholar] [CrossRef]
- Parra, E.R.; Francisco-Cruz, A.; Wistuba, I.I. State-of-the-art of profiling immune contexture in the era of multiplexed staining and digital analysis to study paraffin tumor tissues. Cancers 2019, 11, 247. [Google Scholar] [CrossRef] [Green Version]
- Stack, E.C.; Foukas, P.G.; Lee, P.P. Multiplexed tissue biomarker imaging. J. Immunother. Cancer 2016, 4, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Shiao, S.L.; Yoshida, E.J.; Swartwood, S.; Huang, F.; Doche, M.E.; Chung, A.P.; Knudsen, B.S.; Gertych, A. Data integration from pathology slides for quantitative imaging of multiple cell types within the tumor immune cell infiltrate. Diagn. Pathol. 2017, 12, 69. [Google Scholar] [CrossRef]
- Sideras, K.; Galjart, B.; Vasaturo, A.; Pedroza-Gonzalez, A.; Biermann, K.; Mancham, S.; Nigg, A.L.; Hansen, B.E.; Stoop, H.A.; Zhou, G. Prognostic value of intra-tumoral CD8+/FoxP3+ lymphocyte ratio in patients with resected colorectal cancer liver metastasis. J. Surg. Oncol. 2018, 118, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Soferman, R. RSIP Vision, Visual Intelligence for MedTech. Available online: https://rsipvision.com/about-us/ (accessed on 11 April 2022).
- Griguolo, G.; Serna, G.; Pascual, T.; Fasani, R.; Guardia, X.; Chic, N.; Paré, L.; Pernas, S.; Muñoz, M.; Oliveira, M. Immune microenvironment characterisation and dynamics during anti-HER2-based neoadjuvant treatment in HER2-positive breast cancer. NPJ Precis. Oncol. 2021, 5, 23. [Google Scholar] [CrossRef]
- Graeser, M.; Feuerhake, F.; Gluz, O.; Volk, V.; Hauptmann, M.; Jozwiak, K.; Christgen, M.; Kuemmel, S.; Grischke, E.M.; Forstbauer, H. Immune cell composition and functional marker dynamics from multiplexed immunohistochemistry to predict response to neoadjuvant chemotherapy in the WSG-ADAPT-TN trial. J. Immunother. Cancer 2021, 9, e002198. [Google Scholar] [CrossRef]
- De Angelis, C.; Nagi, C.; Hoyt, C.C.; Liu, L.; Roman, K.; Wang, C.; Zheng, Y.; Veeraraghavan, J.; Sethunath, V.; Nuciforo, P. Evaluation of the predictive role of tumor immune infiltrate in patients with HER2-positive breast cancer treated with neoadjuvant anti-HER2 therapy without chemotherapy. Clin. Cancer Res. 2020, 26, 738–745. [Google Scholar] [CrossRef] [PubMed]
- Kearney, M.R.; Vanguri, R.; Wang, Q.; Fenn, K.; Guo, H.; Marks, D.; Hibshoosh, H.; Kalinsky, K.M.; Connolly, E. Characterization of the tumor microenvironment in patients with hormone receptor positive, HER2 negative early breast cancer. In Proceedings of the San Antonio Breast Cancer Symposium 2021, San Antonio, TX, USA, 7–11 December 2021; Volume P2-07-09. [Google Scholar]
- Zerdes, I.; Simonetti, M.; Matikas, A.; Harbers, L.; Acs, B.; Boyaci, C.; Zhang, N.; Salgkamis, D.; Agartz, S.; Moreno-Ruiz, P. Interplay between copy number alterations and immune profiles in the early breast cancer Scandinavian Breast Group 2004-1 randomized phase II trial: Results from a feasibility study. npj Breast Cancer 2021, 7, 144. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, K.; Kim, I.; Chun, B.; Pucilowska, J.; Redmond, W.L.; Urba, W.J.; Martel, M.; Wu, Y.; Campbell, M.; Sun, Z. Multiplex immunofluorescence to measure dynamic changes in tumor-infiltrating lymphocytes and PD-L1 in early-stage breast cancer. Breast Cancer Res. 2021, 23, 2. [Google Scholar] [CrossRef] [PubMed]
- Noël, G.; Fontsa, M.L.; Garaud, S.; De Silva, P.; de Wind, A.; Van den Eynden, G.G.; Salgado, R.; Boisson, A.; Locy, H.; Thomas, N. Functional Th1-oriented T follicular helper cells that infiltrate human breast cancer promote effective adaptive immunity. J. Clin. Investig. 2021, 131, e139905. [Google Scholar] [CrossRef]
- Egelston, C.A.; Avalos, C.; Tu, T.Y.; Rosario, A.; Wang, R.; Solomon, S.; Srinivasan, G.; Nelson, M.S.; Huang, Y.; Lim, M.H. Resident memory CD8+ T cells within cancer islands mediate survival in breast cancer patients. JCI Insight 2019, 4, e130000. [Google Scholar] [CrossRef] [Green Version]
- Bedard, P.; Siu, L.L.; Thomas, J.; Hanna, D.; Olszanski, A.J.; Azad, N.; Whalen, G.; Ingham, M.; Mahmood, S.; Bender, L.H.; et al. Safety and efficacy of INT230-6, a potential first-in-class intratumoral therapy, in monotherapy and in combination with pembrolizumab: Results from the IT-01 study [KEYNOTE-A10] in subjects with locally advanced, unresectable and metastatic breast cancer. In Proceedings of the San Antonio Breast Cancer Symposium 2021, San Antonio, TX, USA, 7–11 December 2021; Volume P5-16-13. [Google Scholar]
- He, T.F.; Yost, S.E.; Frankel, P.H.; Dagis, A.; Cao, Y.; Wang, R.; Rosario, A.; Tu, T.Y.; Solomon, S.; Schmolze, D. Multi-panel immunofluorescence analysis of tumor infiltrating lymphocytes in triple negative breast cancer: Evolution of tumor immune profiles and patient prognosis. PLoS ONE 2020, 15, e0229955. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, F.S.; Gaule, P.; McGuire, J.; Patel, K.; Blenman, K.; Pusztai, L.; Rimm, D.L. PD-L1 protein expression on both tumor cells and macrophages are associated with response to neoadjuvant durvalumab with chemotherapy in triple-negative breast cancer. Clin. Cancer Res. 2020, 26, 5456–5461. [Google Scholar] [CrossRef]
- Yam, C.; Yen, E.Y.; Chang, J.T.; Bassett, R.L.; Alatrash, G.; Garber, H.; Huo, L.; Yang, F.; Philips, A.V.; Ding, Q.Q.; et al. Immune phenotype and response to neoadjuvant therapy in triple-negative breast cancer. Clin. Cancer Res. 2021, 27, 5365–5375. [Google Scholar] [CrossRef]
- Mani, N.L.; Schalper, K.A.; Hatzis, C.; Saglam, O.; Tavassoli, F.; Butler, M.; Chagpar, A.B.; Pusztai, L.; Rimm, D.L. Quantitative assessment of the spatial heterogeneity of tumor-infiltrating lymphocytes in breast cancer. Breast Cancer Res. 2016, 18, 78. [Google Scholar] [CrossRef] [Green Version]
- Wortman, J.C.; He, T.F.; Solomon, S.; Zhang, R.Z.; Rosario, A.; Wang, R.; Tu, T.Y.; Schmolze, D.; Yuan, Y.; Yost, S.E. Spatial distribution of B cells and lymphocyte clusters as a predictor of triple-negative breast cancer outcome. npj Breast Cancer 2021, 7, 84. [Google Scholar] [CrossRef]
- Stewart, R.L.; Matynia, A.P.; Factor, R.E.; Varley, K.E. Spatially-resolved quantification of proteins in triple negative breast cancers reveals differences in the immune microenvironment associated with prognosis. Sci. Rep. 2020, 10, 6598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, J.M.; Boughey, J.C.; He, J.; Suman, V.J.; Wang, X.; Kachergus, J.M.; Kalari, K.R.; Wang, L.; Weinshilboum, R.; Moyer, A.M.; et al. Abstract PD7-05: Neoadjuvant chemotherapy selectively alters spatially-defined immune landscapes in clinical luminal B HR+/HER2- breast cancers: Analysis of the breast cancer genome guided therapy study (BEAUTY). Cancer Res. 2021, 81, PD7-05. [Google Scholar] [CrossRef]
- Carter, J.M.; Chumsri, S.; Hillman, D.W.; Zahrieh, D.M.; Ma, Y.; Wang, X.; Kachergus, J.M.; Boughey, J.C.; Liu, M.C.; Kalari, K.R.; et al. Abstract PS16-01: Intra-epithelial tumor immune landscapes are associated with clinical outcomes in early-stage triple-negative breast cancer. Cancer Res. 2021, 81, PS16-01. [Google Scholar] [CrossRef]
- McNamara, K.L.; Caswell-Jin, J.L.; Joshi, R.; Ma, Z.; Kotler, E.; Bean, G.R.; Kriner, M.; Zhou, Z.; Hoang, M.; Beechem, J. Spatial proteomic characterization of HER2-positive breast tumors through neoadjuvant therapy predicts response. Nat. Cancer 2021, 2, 400–413. [Google Scholar] [CrossRef] [PubMed]
- Schlam, I.; Church, S.E.; Hether, T.D.; Chaldekas, K.; Hudson, B.M.; White, A.M.; Maisonet, E.; Harris, B.T.; Swain, S.M. The tumor immune microenvironment of primary and metastatic HER2? positive breast cancers utilizing gene expression and spatial proteomic profiling. J. Transl. Med. 2021, 19, 480. [Google Scholar] [CrossRef] [PubMed]
- Garaud, S.; Buisseret, L.; Solinas, C.; Gu-Trantien, C.; de Wind, A.; Van den Eynden, G.; Naveaux, C.; Lodewyckx, J.N.; Boisson, A.; Duvillier, H. Tumor-infiltrating B cells signal functional humoral immune responses in breast cancer. JCI Insight 2019, 5, e129641. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.R.; Wimberly, H.; Lannin, D.R.; Nixon, C.; Rimm, D.L.; Bossuyt, V. Multiplexed quantitative analysis of CD3, CD8, and CD20 predicts response to neoadjuvant chemotherapy in breast cancer. Clin. Cancer Res. 2014, 20, 5995–6005. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.; Kieffer, Y.; Scholer-Dahirel, A.; Pelon, F.; Bourachot, B.; Cardon, M.; Sirven, P.; Magagna, I.; Fuhrmann, L.; Bernard, C. Fibroblast heterogeneity and immunosuppressive environment in human breast cancer. Cancer Cell 2018, 33, 463–479.e10. [Google Scholar] [CrossRef] [Green Version]
- Zerdes, I.; Matikas, A.; Bergh, J.; Rassidakis, G.Z.; Foukakis, T. Genetic, transcriptional and post-translational regulation of the programmed death protein ligand 1 in cancer: Biology and clinical correlations. Oncogene 2018, 37, 4639–4661. [Google Scholar] [CrossRef] [Green Version]
- Janiszewska, M.; Stein, S.; Metzger Filho, O.; Eng, J.; Kingston, N.L.; Harper, N.W.; Rye, I.H.; Alečković, M.; Trinh, A.; Murphy, K.C.; et al. The impact of tumor epithelial and microenvironmental heterogeneity on treatment responses in HER2+ breast cancer. JCI Insight 2021, 6, e147617. [Google Scholar] [CrossRef]
- Millar, E.; Browne, L.; Slapetova, I.; Shang, F.; Ren, Y.; Bradshaw, R.; Ann Brauer, H.; O’Toole, S.; Beretov, J.; Whan, R. TILs immunophenotype in breast cancer predicts local failure and overall survival: Analysis in a large radiotherapy trial with long-term follow-up. Cancers 2020, 12, 2365. [Google Scholar] [CrossRef] [PubMed]
- O’Meara, T.A.; Keenan, T.E.; Waks, A.G.; Felt, K.D.; Sharma, B.; Rodig, S.; Hughes, M.; Lin, N.U.; Agudo, J.; Guerriero, J.L.; et al. Multiplexed immunofluorescence staining of intra-tumoral immune cell populations and associations with immunohistochemical, clinical, and pathologic variables in breast cancer. In Proceedings of the San Antonio Breast Cancer Symposium 2021, San Antonio, TX, USA, 7–11 December 2021; Volume P1-04-05. [Google Scholar]
- Shimada, K.; Cui, Y.X.; Goldberg, J.S.; Pastorello, R.; Davis, J.; Vallius, T.; Kania, L.; Patel, A.; Moore, M.; Ogayo, E.R.; et al. Integrative analysis of single-cell transcriptomic and spatial profiles characterized distinct tumor microenvironment phenotypes in hormone receptor positive (HR+) breast cancer. In Proceedings of the San Antonio Breast Cancer Symposium 2021, San Antonio, TX, USA, 7–11 December 2021; Volume P4-04-06. [Google Scholar]
- Zhu, L.; Narloch, J.L.; Onkar, S.; Joy, M.; Broadwater, G.; Luedke, C.; Hall, A.; Kim, R.; Pogue-Geile, K.; Sammons, S. Metastatic breast cancers have reduced immune cell recruitment but harbor increased macrophages relative to their matched primary tumors. J. Immunother. Cancer 2019, 7, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulasinghe, A.; Monkman, J.; Shah, E.T.; Matigian, N.; Adams, M.N.; O’Byrne, K. Spatial Profiling Identifies Prognostic Features of Response to Adjuvant Therapy in Triple Negative Breast Cancer (TNBC). Front. Oncol. 2021, 11, 798296. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.M.; Polley, M.Y.C.; Leon-Ferre, R.A.; Sinnwell, J.; Thompson, K.J.; Wang, X.; Ma, Y.; Zahrieh, D.; Kachergus, J.M.; Solanki, M.; et al. Characteristics and Spatially Defined Immune (micro) landscapes of Early-stage PD-L1–positive Triple-negative Breast Cancer. Clin. Cancer Res. 2021, 27, 5628–5637. [Google Scholar] [CrossRef] [PubMed]
- Leon-Ferre, R.A.; Carter, J.M.; Zahrieh, D.M.; Hillman, D.W.; Chumsri, S.; Ma, Y.; Kachergus, J.M.; Wang, X.; Boughey, J.C.; Liu, M.C.; et al. Digital spatial profiling of immune-related proteins in luminal androgen receptor (LAR) vs. non-LAR triple-negative breast cancer (TNBC). In Proceedings of the San Antonio Breast Cancer Symposium 2021, San Antonio, TX, USA, 7–11 December 2021; Volume P1-04-01. [Google Scholar]
- Bianchini, G.; Wang, X.Q.; Danenberg, E.; Huang, C.S. Single-cell spatial analysis by imaging mass cytometry and immunotherapy response in triple-negative breast cancer (TNBC) in the NeoTRIPaPDL1 trial. In Proceedings of the San Antonio Breast Cancer Symposium 2021, San Antonio, TX, USA, 7–11 December 2021. [Google Scholar]
- Page, D.B.; Pucilowska, J.; Sanchez, K.G.; Conrad, V.K.; Conlin, A.K.; Acheson, A.K.; Perlewitz, K.S.; Imatani, J.H.; Aliabadi-Wahle, S.; Moxon, N.; et al. A phase Ib study of preoperative, locoregional IRX-2 cytokine immunotherapy to prime immune responses in patients with early-stage breast cancer. Clin. Cancer Res. 2020, 26, 1595–1605. [Google Scholar] [CrossRef] [PubMed]
- Jackson, H.W.; Fischer, J.R.; Zanotelli, V.R.; Ali, H.R.; Mechera, R.; Soysal, S.D.; Moch, H.; Muenst, S.; Varga, Z.; Weber, W.P. The single-cell pathology landscape of breast cancer. Nature 2020, 578, 615–620. [Google Scholar] [CrossRef]
- Ali, H.R.; Jackson, H.W.; Zanotelli, V.R.; Danenberg, E.; Fischer, J.R.; Bardwell, H.; Provenzano, E.; Rueda, O.M.; Chin, S.F.; Aparicio, S. Imaging mass cytometry and multiplatform genomics define the phenogenomic landscape of breast cancer. Nat. Cancer 2020, 1, 163–175. [Google Scholar] [CrossRef]
- Xiao, Y.; Ma, D.; Zhao, S.; Suo, C.; Shi, J.; Xue, M.Z.; Ruan, M.; Wang, H.; Zhao, J.; Li, Q. Multi-omics profiling reveals distinct microenvironment characterization and suggests immune escape mechanisms of triple-negative breast cancer. Clin. Cancer Res. 2019, 25, 5002–5014. [Google Scholar] [CrossRef] [Green Version]
- DeNardo, D.G.; Brennan, D.J.; Rexhepaj, E.; Ruffell, B.; Shiao, S.L.; Madden, S.F.; Gallagher, W.M.; Wadhwani, N.; Keil, S.D.; Junaid, S.A. Leukocyte complexity predicts breast cancer survival and functionally regulates response to chemotherapy. Cancer Discov. 2011, 1, 54–67. [Google Scholar] [CrossRef] [Green Version]
- Boman, C.; Zerdes, I.; Mårtensson, K.; Bergh, J.; Foukakis, T.; Valachis, A.; Matikas, A. Discordance of PD-L1 status between primary and metastatic breast cancer: A systematic review and meta-analysis. Cancer Treat. Rev. 2021, 99, 102257. [Google Scholar] [CrossRef]
- Rizzo, A.; Ricci, A.D. Biomarkers for breast cancer immunotherapy: PD-L1, TILs and beyond. Expert Opin. Investig. Drugs 2021, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Helmink, B.A.; Reddy, S.M.; Gao, J.; Zhang, S.; Basar, R.; Thakur, R.; Yizhak, K.; Sade-Feldman, M.; Blando, J.; Han, G. B cells and tertiary lymphoid structures promote immunotherapy response. Nature 2020, 577, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Cabrita, R.; Lauss, M.; Sanna, A.; Donia, M.; Larsen, M.S.; Mitra, S.; Johansson, I.; Phung, B.; Harbst, K.; Vallon-Christersson, J. Tertiary lymphoid structures improve immunotherapy and survival in melanoma. Nature 2020, 577, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Petitprez, F.; de Reyniès, A.; Keung, E.Z.; Chen, T.W.W.; Sun, C.M.; Calderaro, J.; Jeng, Y.M.; Hsiao, L.P.; Lacroix, L.; Bougoüin, A. B cells are associated with survival and immunotherapy response in sarcoma. Nature 2020, 577, 556–560. [Google Scholar] [CrossRef]
- Acs, B.; Hartman, J. Next generation pathology: Artificial intelligence enhances histopathology practice. J. Pathol. 2020, 250, 7–8. [Google Scholar] [CrossRef]
- Amgad, M.; Stovgaard, E.S.; Balslev, E.; Thagaard, J.; Chen, W.; Dudgeon, S.; Sharma, A.; Kerner, J.K.; Denkert, C.; Yuan, Y. Report on computational assessment of tumor infiltrating lymphocytes from the International Immuno-Oncology Biomarker Working Group. npj Breast Cancer 2020, 6, 16. [Google Scholar] [CrossRef]
- Bai, Y.; Cole, K.; Martinez-Morilla, S.; Ahmed, F.S.; Zugazagoitia, J.; Staaf, J.; Bosch, A.; Ehinger, A.; Niméus, E.; Hartman, J. An Open Source, Automated Tumor Infiltrating Lymphocyte Algorithm for Prognosis in Triple-Negative Breast Cancer. Clin. Cancer Res. 2021, 27, 5557–5565. [Google Scholar] [CrossRef]
- Ali, H.; Dariush, A.; Thomas, J.; Provenzano, E.; Dunn, J.; Hiller, L.; Vallier, A.L.; Abraham, J.; Piper, T.; Bartlett, J. Lymphocyte density determined by computational pathology validated as a predictor of response to neoadjuvant chemotherapy in breast cancer: Secondary analysis of the ARTemis trial. Ann. Oncol. 2017, 28, 1832–1835. [Google Scholar] [CrossRef]
- Ali, H.R.; Dariush, A.; Provenzano, E.; Bardwell, H.; Abraham, J.E.; Iddawela, M.; Vallier, A.L.; Hiller, L.; Dunn, J.A.; Bowden, S.J. Computational pathology of pre-treatment biopsies identifies lymphocyte density as a predictor of response to neoadjuvant chemotherapy in breast cancer. Breast Cancer Res. 2016, 18, 21. [Google Scholar] [CrossRef]
- Park, Y.H.; Lal, S.; Lee, J.E.; Choi, Y.L.; Wen, J.; Ram, S.; Ding, Y.; Lee, S.H.; Powell, E.; Lee, S.K. Chemotherapy induces dynamic immune responses in breast cancers that impact treatment outcome. Nat. Commun. 2020, 11, 6175. [Google Scholar] [CrossRef]
- Nederlof, I.; Hajizadeh, S.; Sobhani, F.; Raza, S.E.A.; Desmedt, C.; Salgado, R.F.; Kok, M.; Yuan, Y.; Horlings, H. Spatial analysis of lymphocytes and fibroblasts identifies biological relevant patterns in estrogen receptor positive breast cancer. ESMO Breast Cancer Congress 2020. Available online: https://oncologypro.esmo.org/meeting-resources/esmo-breast-cancer-congress/spatial-analysis-of-lymphocytes-and-fibroblasts-identifies-biological-relevant-patterns-in-estrogen-receptor-positive-breast-cancer (accessed on 11 April 2022).
- Crosetto, N.; Bienko, M.; Van Oudenaarden, A. Spatially resolved transcriptomics and beyond. Nat. Rev. Genet. 2015, 16, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.M.; Asselin-Labat, M.L.; Nguyen, Q.; Berthelet, J.; Tan, X.; Wimmer, V.C.; Merino, D.; Rogers, K.L.; Naik, S.H. Spatial omics and multiplexed imaging to explore cancer biology. Nat. Methods 2021, 18, 997–1012. [Google Scholar] [CrossRef] [PubMed]
- Allam, M.; Cai, S.; Coskun, A.F. Multiplex bioimaging of single-cell spatial profiles for precision cancer diagnostics and therapeutics. NPJ Precis. Oncol. 2020, 4, 11. [Google Scholar] [CrossRef]
- Andersson, A.; Larsson, L.; Stenbeck, L.; Salmén, F.; Ehinger, A.; Wu, S.Z.; Al-Eryani, G.; Roden, D.; Swarbrick, A.; Borg, A. Spatial deconvolution of HER2-positive breast cancer delineates tumor-associated cell type interactions. Nat. Commun. 2021, 12, 6012. [Google Scholar] [CrossRef]
- Nederlof, I.; De Bortoli, D.; Bareche, Y.; Nguyen, B.; De Maaker, M.; Hooijer, G.K.; Buisseret, L.; Kok, M.; Smid, M.; Van den Eynden, G.G. Comprehensive evaluation of methods to assess overall and cell-specific immune infiltrates in breast cancer. Breast Cancer Res. 2019, 21, 151. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.X.; Song, M.; Maecker, H.T.; Gnjatic, S.; Patton, D.; Lee, J.J.; Adam, S.J.; Moravec, R.; Liu, X.S.; Cerami, E. Network for biomarker immunoprofiling for cancer immunotherapy: Cancer Immune Monitoring and Analysis Centers and Cancer Immunologic Data Commons (CIMAC-CIDC). Clin. Cancer Res. 2021, 27, 5038–5048. [Google Scholar] [CrossRef]
- Bergholtz, H.; Carter, J.M.; Cesano, A.; Cheang, M.C.U.; Church, S.E.; Divakar, P.; Fuhrman, C.A.; Goel, S.; Gong, J.; Guerriero, J.L. Best Practices for Spatial Profiling for Breast Cancer Research with the GeoMx® Digital Spatial Profiler. Cancers 2021, 13, 4456. [Google Scholar] [CrossRef]
- Blank, C.U.; Haining, W.N.; Held, W.; Hogan, P.G.; Kallies, A.; Lugli, E.; Lynn, R.C.; Philip, M.; Rao, A.; Restifo, N.P. Defining T cell exhaustion. Nat. Rev. Immunol. 2019, 19, 665–674. [Google Scholar] [CrossRef]
- Hickey, J.W.; Tan, Y.; Nolan, G.P.; Goltsev, Y. Strategies for accurate cell type identification in CODEX multiplexed imaging data. Front. Immunol. 2021, 12, 727626. [Google Scholar] [CrossRef]
- McKinley, E.T.; Sui, Y.; Al-Kofahi, Y.; Millis, B.A.; Tyska, M.J.; Roland, J.T.; Santamaria-Pang, A.; Ohland, C.L.; Jobin, C.; Franklin, J.L. Optimized multiplex immunofluorescence single-cell analysis reveals tuft cell heterogeneity. JCI Insight 2017, 2, e93487. [Google Scholar] [CrossRef] [PubMed]
- Grys, B.T.; Lo, D.S.; Sahin, N.; Kraus, O.Z.; Morris, Q.; Boone, C.; Andrews, B.J. Machine learning and computer vision approaches for phenotypic profiling. J. Cell Biol. 2017, 216, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Caicedo, J.C.; Cooper, S.; Heigwer, F.; Warchal, S.; Qiu, P.; Molnar, C.; Vasilevich, A.S.; Barry, J.D.; Bansal, H.S.; Kraus, O. Data-analysis strategies for image-based cell profiling. Nat. Methods 2017, 14, 849–863. [Google Scholar] [CrossRef] [PubMed]
- Boisson, A.; Noël, G.; Saiselet, M.; Rodrigues-Vitória, J.; Thomas, N.; Fontsa, M.L.; Sofronii, D.; Naveaux, C.; Duvillier, H.; Craciun, L. Fluorescent Multiplex Immunohistochemistry Coupled With Other State-Of-The-Art Techniques to Systematically Characterize the Tumor Immune Microenvironment. Front. Mol. Biosci. 2021, 8, 673042. [Google Scholar] [CrossRef] [PubMed]

|
Software (Company) | Analyses/Capacity |
|---|---|
| Inform [56] (Akoya Biosciences) | multispectral unmixing, tissue and cell segmentation, cellular phenotyping |
| MultiOmyx [37] (Neo Genomics) | quantitative analysis at the cell-level, spatial analytics |
| HALO [57] (Indica Labs) | immune cell population contexture analysis segmentation and classification |
| Visiopharm [58] | cellular identification spatial profiling |
| RSIPVision [59] | nuclei detection, segmentation and cellular colocalization |
| QuPath [40] | cell segmentation and classification |
| Multiplexing Level | |
|---|---|
| High | Challenging panel validation, increase in image analysis labor/difficult cell phenotyping |
| Panel Validation | |
| Section thickness | Affects staining intensity and tissue autofluorescence |
| Staining sequence of primary antibodies | Can deal with epitope instability and cross reaction of primary antibodies |
| Fluorophores, Co-localization in the same cellular compartment, Low abundance epitopes | Selection of spectrally separated fluorophores; selection of more intense fluorophores |
| Staining pattern | For each antibody, staining pattern in multiplex image should be identical to single-plex immunohistochemistry |
| Regions of interest | |
| Number | Whole tissue section resemblance: as many as possible evaluation |
| Prior selection | Potential selection bias |
| Statistical analysis | |
| Optimal statistical method | Hierarchical linear modeling: statistical power improvement over t-test. |
| Cut-off | Biomarkers expressed by various cell types: establishment of single positivity threshold is difficult |
| Image Analysis | |
| Storage | Extreme data sizes |
| Cell phenotyping | Difficult cellular segmentation: 1. Mixed cells with different shapes and sizes 2. High multiplexing level |
| Distance analysis | Densities can be confounding factor: Possible solution proximity analysis between areas with similar levels of cellular infiltration |
| Bias | Inter-center and inter-vendor variability, as well as intra- and inter-observer variability either during tissue sample preparation or during image acquisition. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tzoras, E.; Zerdes, I.; Tsiknakis, N.; Manikis, G.C.; Mezheyeuski, A.; Bergh, J.; Matikas, A.; Foukakis, T. Dissecting Tumor-Immune Microenvironment in Breast Cancer at a Spatial and Multiplex Resolution. Cancers 2022, 14, 1999. https://doi.org/10.3390/cancers14081999
Tzoras E, Zerdes I, Tsiknakis N, Manikis GC, Mezheyeuski A, Bergh J, Matikas A, Foukakis T. Dissecting Tumor-Immune Microenvironment in Breast Cancer at a Spatial and Multiplex Resolution. Cancers. 2022; 14(8):1999. https://doi.org/10.3390/cancers14081999
Chicago/Turabian StyleTzoras, Evangelos, Ioannis Zerdes, Nikos Tsiknakis, Georgios C. Manikis, Artur Mezheyeuski, Jonas Bergh, Alexios Matikas, and Theodoros Foukakis. 2022. "Dissecting Tumor-Immune Microenvironment in Breast Cancer at a Spatial and Multiplex Resolution" Cancers 14, no. 8: 1999. https://doi.org/10.3390/cancers14081999