
Comprehensive Analysis Identifies Ameloblastin-Related Competitive Endogenous RNA as a Prognostic Biomarker for Testicular Germ Cell Tumour

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Preparation
2.2. Immunofluorescence of Testis Tissues from Rat
2.3. The RNAseq Data Analysis of AMBN
2.4. Differential Gene Expression Analysis
2.5. Survival Analysis and Construction of Gene-Specific Prognosis Models for TGCT
2.6. Immune Infiltrate Levels Related to AMBN
2.7. Functional Enrichment Analysis
2.8. Establishment of the ceRNA Network Related to AMBN in TGCT
2.9. Methylation and Expression Analysis of GFAP
2.10. Statistical Analysis
3. Results
3.1. Down-Regulation of AMBN Expression and Clinical Value in TGCT
3.2. Analysis of Differentially Expressed Genes (DEGs)
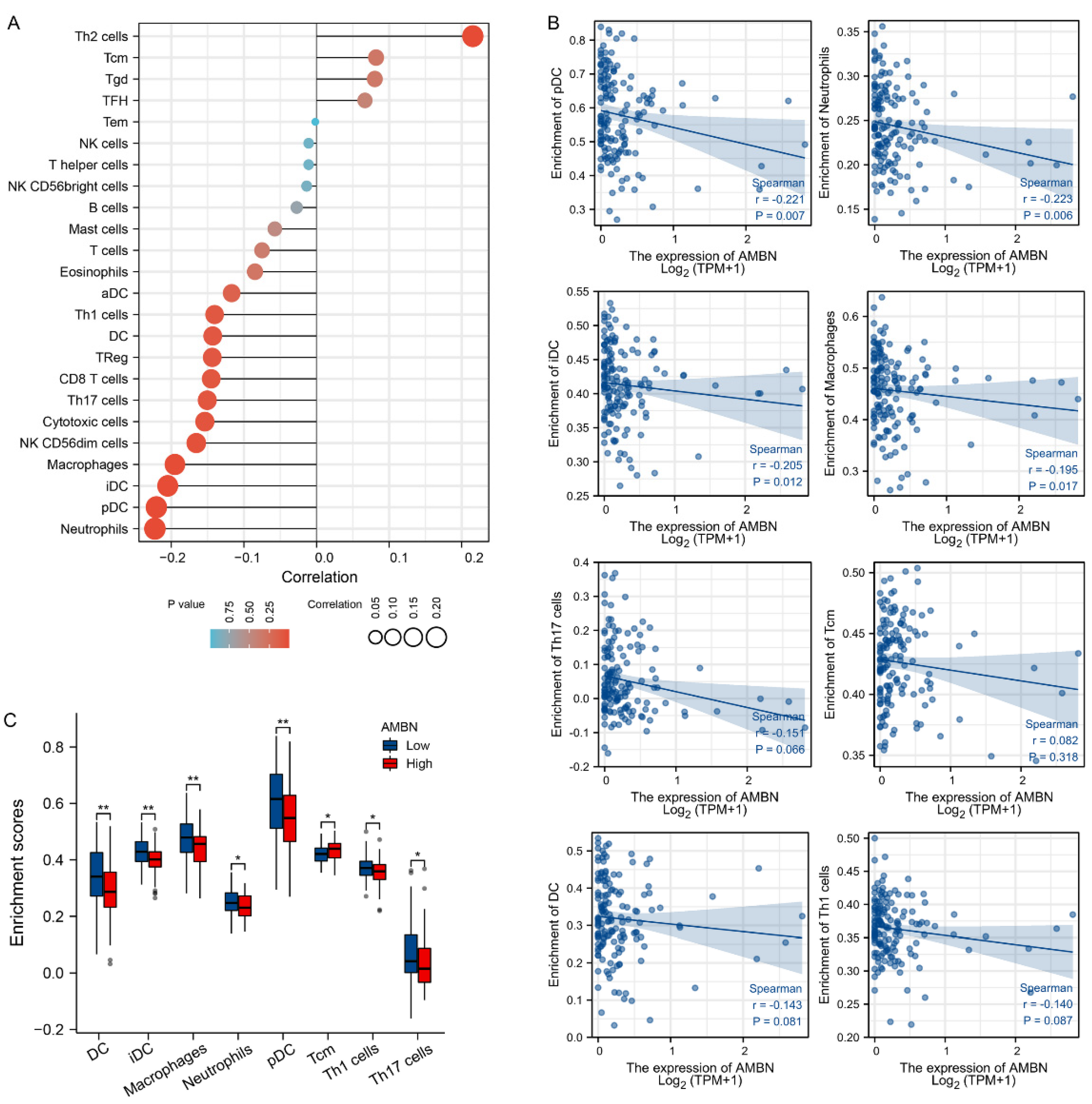
3.3. Correlation between Immune Cell Infiltration and AMBN mRNA Expression in TGCT
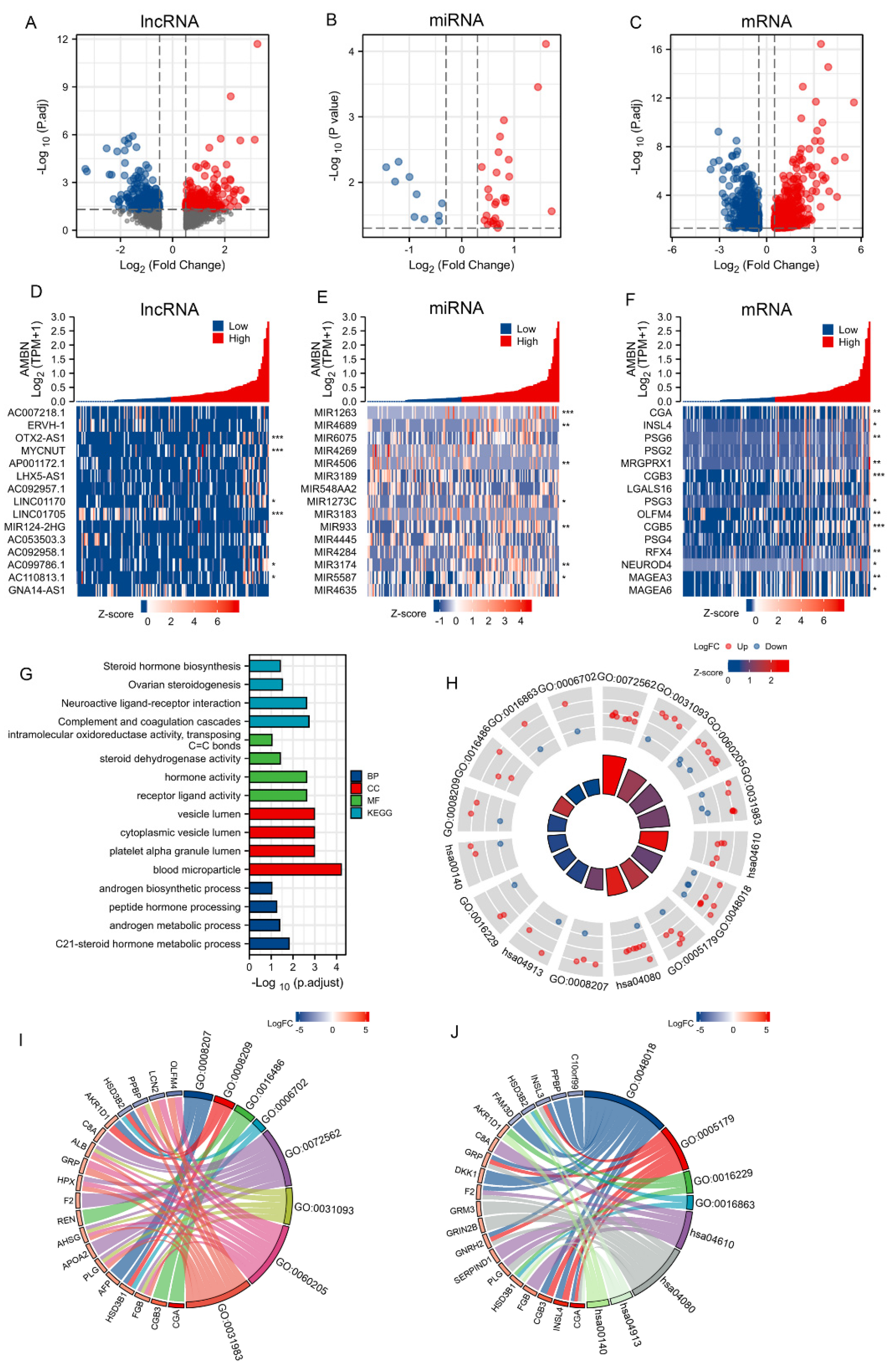
3.4. Construction of a lncRNA-miRNA-mRNA Triplet Regulatory Network and Its Functional Enrichment
3.5. Validation of the ceRNA Network Model
3.6. Prognostic Analysis of the ceRNA Network Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Richardson, L.C.; Neri, A.J.; Tai, E.; Glenn, J.D. Testicular cancer: A narrative review of the role of socioeconomic position from risk to survivorship. Urol. Oncol. Semin. Orig. Investig. 2012, 30, 95–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batool, A.; Karimi, N.; Wu, X.-N.; Chen, S.-R.; Liu, Y.-X. Testicular germ cell tumor: A comprehensive review. Cell. Mol. Life Sci. 2019, 76, 1713–1727. [Google Scholar] [CrossRef] [PubMed]
- Gillessen, S.; Sauvé, N.; Collette, L.; Daugaard, G.; de Wit, R.; Albany, C.; Tryakin, A.; Fizazi, K.; Stahl, O.; Gietema, J.A.; et al. Predicting Outcomes in Men With Metastatic Nonseminomatous Germ Cell Tumors (NSGCT): Results From the IGCCCG Update Consortium. J. Clin. Oncol. 2021, 39, 1563–1574. [Google Scholar] [CrossRef] [PubMed]
- Ylönen, O.; Jyrkkiö, S.; Pukkala, E.; Syvänen, K.; Boström, P.J. Time trends and occupational variation in the incidence of testicular cancer in the Nordic countries. Br. J. Urol. 2018, 122, 384–393. [Google Scholar] [CrossRef] [Green Version]
- Oing, C.; Kollmannsberger, C.; Oechsle, K.; Bokemeyer, C. Investigational targeted therapies for the treatment of testicular germ cell tumors. Expert Opin. Investig. Drugs 2016, 25, 1033–1043. [Google Scholar] [CrossRef]
- Albers, P.; Albrecht, W.; Algaba, F.; Bokemeyer, C.; Cohn-Cedermark, G.; Fizazi, K.; Horwich, A.; Laguna, M.P.; Nicolai, N.; Oldenburg, J. Guidelines on Testicular Cancer: 2015 Update. Eur. Urol. 2015, 68, 1054–1068. [Google Scholar] [CrossRef]
- Suarez-Carmona, M.; Lesage, J.; Cataldo, D.; Gilles, C. EMT and inflammation: Inseparable actors of cancer progression. Mol. Oncol. 2017, 11, 805–823. [Google Scholar] [CrossRef]
- Bahcecioglu, G.; Basara, G.; Ellis, B.W.; Ren, X.; Zorlutuna, P. Breast cancer models: Engineering the tumor microenvironment. Acta Biomater. 2020, 106, 1–21. [Google Scholar] [CrossRef]
- Nallanthighal, S.; Heiserman, J.P.; Cheon, D.-J. The Role of the Extracellular Matrix in Cancer Stemness. Front. Cell Dev. Biol. 2019, 7, 86. [Google Scholar] [CrossRef]
- Provenzano, P.P.; Eliceiri, K.W.; Campbell, J.M.; Inman, D.R.; White, J.G.; Keely, P.J. Collagen reorganization at the tumor-stromal interface facilitates local invasion. BMC Med. 2006, 4, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 2020, 11, 5120. [Google Scholar] [CrossRef] [PubMed]
- Krebsbach, P.H.; Lee, S.K.; Matsuki, Y.; Kozak, C.A.; Yamada, K.; Yamada, Y. Full-length Sequence, Localization, and Chromosomal Mapping of Ameloblastin. J. Biol. Chem. 1996, 271, 4431–4435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atsawasuwan, P.; Lu, X.; Ito, Y.; Chen, Y.; Gopinathan, G.; Evans, C.; Kulkarni, A.; Gibson, C.; Luan, X.; Diekwisch, T.G. Expression and Function of Enamel-related Gene Products in Calvarial Development. J. Dent. Res. 2013, 92, 622–628. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Fukumoto, S.; Yamada, Y.; A Evans, C.; Diekwisch, T.G.; Luan, X. Ameloblastin, an Extracellular Matrix Protein, Affects Long Bone Growth and Mineralization. J. Bone Miner. Res. 2016, 31, 1235–1246. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Diekwisch, T.G.H.; Luan, X. Structure and function of ameloblastin as an extracellular matrix protein: Adhesion, calcium binding, and CD63 interaction in human and mouse. Eur. J. Oral Sci. 2011, 119, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Sonoda, A.; Iwamoto, T.; Nakamura, T.; Fukumoto, E.; Yoshizaki, K.; Yamada, A.; Arakaki, M.; Harada, H.; Nonaka, K.; Nakamura, S.; et al. Critical Role of Heparin Binding Domains of Ameloblastin for Dental Epithelium Cell Adhesion and Ameloblastoma Proliferation. J. Biol. Chem. 2009, 284, 27176–27184. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.; Wu, Y.-P.; Yin, H.-B.; Xue, X.-Y.; Gou, X. Molecular network-based identification of competing endogenous RNAs and mRNA signatures that predict survival in prostate cancer. J. Transl. Med. 2018, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef]
- Geisler, S.; Coller, J. RNA in unexpected places: Long non-coding RNA functions in diverse cellular contexts. Nat. Rev. Mol. Cell Biol. 2013, 14, 699–712. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Fu, H.; Wu, Y.; Zheng, X. Function of lncRNAs and approaches to lncRNA-protein interactions. Sci. China Life Sci. 2013, 56, 876–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, W.-X.; Koirala, P.; Mo, Y.-Y. LncRNA-mediated regulation of cell signaling in cancer. Oncogene 2017, 36, 5661–5667. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Tian, H.; Cao, Y.-X.; He, X.; Chen, L.; Song, X.; Ping, P.; Huang, H.; Sun, F. Downregulation of miR-320a/383-sponge-like long non-coding RNA NLC1-C (narcolepsy candidate-region 1 genes) is associated with male infertility and promotes testicular embryonal carcinoma cell proliferation. Cell Death Dis. 2015, 6, e1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bo, H.; Zhu, F.; Liu, Z.; Deng, Q.; Liu, G.; Li, R.; Zhu, W.; Tan, Y.; Liu, G.; Fan, J.; et al. Integrated analysis of high-throughput sequencing data reveals the key role of LINC00467 in the invasion and metastasis of testicular germ cell tumors. Cell Death Discov. 2021, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhandari, V.; Hoey, C.; Liu, L.Y.; LaLonde, E.; Ray, J.; Livingstone, J.; Lesurf, R.; Shiah, Y.-J.; Vujcic, T.; Huang, X.; et al. Molecular landmarks of tumor hypoxia across cancer types. Nat. Genet. 2019, 51, 308–318. [Google Scholar] [CrossRef]
- Climent, M.; Viggiani, G.; Chen, Y.-W.; Coulis, G.; Castaldi, A. MicroRNA and ROS Crosstalk in Cardiac and Pulmonary Diseases. Int. J. Mol. Sci. 2020, 21, 4370. [Google Scholar] [CrossRef]
- Esquela-Kerscher, A.; Slack, F. Oncomirs—MicroRNAs with a role in cancer. Nat. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Karreth, F.A.; Pandolfi, P.P. ceRNA Cross-Talk in Cancer: When ce-bling Rivalries Go Awry. Cancer Discov. 2013, 3, 1113–1121. [Google Scholar] [CrossRef] [Green Version]
- Vivian, J.; Rao, A.A.; Nothaft, F.A.; Ketchum, C.; Armstrong, J.; Novak, A.; Pfeil, J.; Narkizian, J.; Deran, A.D.; Musselman-Brown, A.; et al. Toil enables reproducible, open source, big biomedical data analyses. Nat. Biotechnol. 2017, 35, 314–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusumbe, A.P.; Ramasamy, S.; Starsichova, A.; Adams, R.H. Sample preparation for high-resolution 3D confocal imaging of mouse skeletal tissue. Nat. Protoc. 2015, 10, 1904–1914. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-Seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef] [Green Version]
- Bindea, G.; Mlecnik, B.; Tosolini, M.; Kirilovsky, A.; Waldner, M.; Obenauf, A.C.; Angell, H.; Fredriksen, T.; Lafontaine, L.; Berger, A.; et al. Spatiotemporal Dynamics of Intratumoral Immune Cells Reveal the Immune Landscape in Human Cancer. Immunity 2013, 39, 782–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Paraskevopoulou, M.D.; Vlachos, I.S.; Karagkouni, D.; Georgakilas, G.; Kanellos, I.; Vergoulis, T.; Zagganas, K.; Tsanakas, P.; Floros, E.; Dalamagas, T.; et al. DIANA-LncBase v2: Indexing microRNA targets on non-coding transcripts. Nucleic Acids Res. 2016, 44, D231–D238. [Google Scholar] [CrossRef] [Green Version]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. miRWalk: An online resource for prediction of microRNA binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines, Indicates that Thousands of Human Genes are MicroRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Z.; Pan, X.; Yang, Y.; Huang, Y.; Shen, H.-B. The lncLocator: A subcellular localization predictor for long non-coding RNAs based on a stacked ensemble classifier. Bioinformatics 2018, 34, 2185–2194. [Google Scholar] [CrossRef]
- Enright, A.J.; John, B.; Gaul, U.; Tuschl, T.; Sander, C.; Marks, D.S. MicroRNA targets in Drosophila. Genome Biol. 2003, 5, R1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.; Jeschke, J.; Van Criekinge, W.; Van Engeland, M.; De Meyer, T. MEXPRESS update 2019. Nucleic Acids Res. 2019, 47, W561–W565. [Google Scholar] [CrossRef]
- Schuebel, K.E.; Chen, W.; Cope, L.; Glöckner, S.C.; Suzuki, H.; Yi, J.-M.; Chan, T.A.; Van Neste, L.; Van Criekinge, W.; Bosch, S.V.D.; et al. Comparing the DNA Hypermethylome with Gene Mutations in Human Colorectal Cancer. PLoS Genet. 2007, 3, e157. [Google Scholar] [CrossRef]
- Oldenburg, J.; Berney, D.; Bokemeyer, C.; Climent, M.; Daugaard, G.; Gietema, J.; De Giorgi, U.; Haugnes, H.; Huddart, R.; Leão, R.; et al. Testicular seminoma and non-seminoma: ESMO-EURACAN Clinical Practice Guideline for diagnosis, treatment and follow-up. Ann. Oncol. 2022, 33, 362–375. Available online: https://pubmed.ncbi.nlm.nih.gov/35065204/ (accessed on 15 September 2021). [CrossRef]
- Ando, T.; Kudo, Y.; Iizuka, S.; Tsunematsu, T.; Umehara, H.; Shrestha, M.; Matsuo, T.; Kubo, T.; Shimose, S.; Arihiro, K.; et al. Ameloblastin induces tumor suppressive phenotype and enhances chemosensitivity to doxorubicin via Src-Stat3 inactivation in osteosarcoma. Sci. Rep. 2017, 7, 40187. [Google Scholar] [CrossRef] [Green Version]
- Gajewski, T.F.; Schreiber, H.; Fu, Y.-X. Innate and adaptive immune cells in the tumor microenvironment. Nat. Immunol. 2013, 14, 1014–1022. [Google Scholar] [CrossRef] [Green Version]
- Tamburstuen, M.V.; Reppe, S.; Spahr, A.; Sabetrasekh, R.; Kvalheim, G.; Slaby, I.; Syversen, U.; Lyngstadaas, S.P.; Reseland, J.E. Ameloblastin promotes bone growth by enhancing proliferation of progenitor cells and by stimulating immunoregulators. Eur. J. Oral Sci. 2010, 118, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, M.; Okinaga, T.; Ariyoshi, W.; Kitamura, C.; Nishihara, T. Ameloblastin Upregulates Inflammatory Response through Induction of IL-1β in Human Macrophages. J. Cell. Biochem. 2017, 118, 3308–3317. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, N.; Nakayama, H.; Miyazaki, E.; Hayashi, Y.; Toi, M.; Hiroi, M.; Enzan, H. Distribution and role of CD34-positive stromal cells and myofibroblasts in human normal testicular stroma. Histol. Histopathol. 2004, 19, 743–751. [Google Scholar]
- Sawai, N.; Koike, K.; Ito, S.; Mwamtemi, H.H.; Kurokawa, Y.; Kinoshita, T.; Sakashita, K.; Higuchi, T.; Takeuchi, K.; Shiohara, M.; et al. Neutrophilic cell production by combination of stem cell factor and thrombopoietin from CD34(+) cord blood cells in long-term serum-deprived liquid culture. Blood 1999, 93, 509–518. [Google Scholar] [CrossRef]
- Clanchy, F.I.; Hamilton, J.A. The development of macrophages from human CD34+ haematopoietic stem cells in serum-free cultures is optimized by IL-3 and SCF. Cytokine 2013, 61, 33–37. [Google Scholar] [CrossRef]
- Tamburstuen, M.V.; Reseland, J.E.; Spahr, A.; Brookes, S.J.; Kvalheim, G.; Slaby, I.; Snead, M.L.; Lyngstadaas, S.P. Ameloblastin expression and putative autoregulation in mesenchymal cells suggest a role in early bone formation and repair. Bone 2011, 48, 406–413. [Google Scholar] [CrossRef] [Green Version]
- Parker, C.; Milosevic, M.; Panzarella, T.; Banerjee, D.; Jewett, M.; Catton, C.; Tew-George, B.; Gospodarowicz, M.; Warde, P. The prognostic significance of the tumour infiltrating lymphocyte count in stage I testicular seminoma managed by surveillance. Eur. J. Cancer 2002, 38, 2014–2019. [Google Scholar] [CrossRef]
- Solinas, C.; Chanzá, N.M.; Awada, A.; Scartozzi, M. The immune infiltrate in prostate, bladder and testicular tumors: An old friend for new challenges. Cancer Treat. Rev. 2016, 53, 138–145. [Google Scholar] [CrossRef]
- Suzuki, K.; Shin, T.; Shimomura, Y.; Iwahata, T.; Okada, H. Spermatogenesis in tumor-bearing testes in germ cell testicular cancer patients. Hum. Reprod. 2015, 30, 2853–2858. [Google Scholar] [CrossRef] [Green Version]
- Saint, F.; Leroy, X.; Graziana, J.P.; Moukassa, D.; Gosselin, B.; Biserte, J.; Chopin, D.; Rigot, J.M. Dendritic Cell Infiltration in A Patient with Seminomatous Germ Cell Tumor of the Testis: Is There a Relationship with Infertility and Tumor Stage? J. Urol. 2002, 167, 1643–1647. [Google Scholar] [CrossRef]
- Cochran, J.S.; Walsh, P.C.; Porter, J.C.; Nicholson, T.C.; Madden, J.D.; Peters, P.C. The Endocrinology of Human Chorionic Gonadotropin-Secreting Testicular Tumors: New Methods in Diagnosis. J. Urol. 1975, 114, 549–555. [Google Scholar] [CrossRef]
- Salemi, M.; Lanza, G.; Mogavero, M.P.; Cosentino, F.I.I.; Borgione, E.; Iorio, R.; Ventola, G.M.; Marchese, G.; Salluzzo, M.G.; Ravo, M.; et al. A Transcriptome Analysis of mRNAs and Long Non-Coding RNAs in Patients with Parkinson’s Disease. Int. J. Mol. Sci. 2022, 23, 1535. [Google Scholar] [CrossRef] [PubMed]
- Marston, J.L.; Greenig, M.; Singh, M.; Bendall, M.L.; Duarte, R.R.; Feschotte, C.; Iñiguez, L.P.; Nixon, D.F. SARS-CoV-2 infection mediates differential expression of human endogenous retroviruses and long interspersed nuclear elements. JCI Insight 2021, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, M.; Kollmannsberger, C.; Nappi, L. Emerging Role of Biomarkers in Testicular Germ Cell Tumors. Curr. Oncol. Rep. 2022, 24, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Pan, X.; Xu, P.; Mi, Y.; Wang, W.; Wu, X.; He, Q.; Liu, X.; Tang, W.; An, H.-X. Plasma microRNA alterations between EGFR-activating mutational NSCLC patients with and without primary resistance to TKI. Oncotarget 2017, 8, 88529–88536. [Google Scholar] [CrossRef] [Green Version]
- Middeldorp, J.; Hol, E.M. GFAP in health and disease. Prog. Neurobiol. 2011, 93, 421–443. [Google Scholar] [CrossRef]
- Jung, C.S.; Foerch, C.; Schanzer, A.; Heck, A.; Plate, K.H.; Seifert, V.; Steinmetz, H.; Raabe, A.; Sitzer, M. Serum GFAP is a diagnostic marker for glioblastoma multiforme. Brain 2007, 130, 3336–3341. [Google Scholar] [CrossRef]
- Tian, J.; Song, T.; Wang, H.; Wang, W.; Zhang, Z.; Yan, R. Thalidomide alleviates bone cancer pain by down-regulating expressions of NF-κB and GFAP in spinal astrocytes in a mouse model. Int. J. Neurosci. 2019, 129, 896–903. [Google Scholar] [CrossRef]
- Hainfellner, J.A.; Voigtländer, T.; Ströbel, T.; Mazal, P.R.; Maddalena, A.S.; Aguzzi, A.; Budka, H. Fibroblasts can express glial fibrillary acidic protein (GFAP) in vivo. J. Neuropathol. Exp. Neurol. 2001, 60, 449–461. [Google Scholar] [CrossRef] [Green Version]
- Davidoff, M.S.; Middendorff, R.; Köfüncü, E.; Müller, D.; Ježek, D.; Holstein, A.-F. Leydig cells of the human testis possess astrocyte and oligodendrocyte marker molecules. Acta Histochem. 2002, 104, 39–49. [Google Scholar] [CrossRef]
- Omirinde, J.; Olukole, S.; Oke, B. Nerve and glial cell expressions in the testes and epididymides of different age groups of cane rat (Thryonomys swinderianus). J. Microsc. Ultrastruct. 2021, 9, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Restrepo, A.; Smith, C.A.; Agnihotri, S.; Shekarforoush, M.; Kongkham, P.N.; Seol, H.J.; Northcott, P.; Rutka, J.T. Epigenetic regulation of glial fibrillary acidic protein by DNA methylation in human malignant gliomas. Neuro-Oncology 2010, 13, 42–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Zhang, Y.-F.; Zhang, Z.-P.; Fu, L.-A.; Cao, X.-L.; Zhang, Y.-Z.; Guo, C.-J.; Yan, X.-C.; Yang, Q.-C.; Hu, Y.-Y.; et al. miR-342-5p Regulates Neural Stem Cell Proliferation and Differentiation Downstream to Notch Signaling in Mice. Stem Cell Rep. 2017, 8, 1032–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molla, M.; Descroix, V.; Aïoub, M.; Simon, S.; Castañeda, B.; Hotton, D.; Bolaños, A.; Simon, Y.; Lezot, F.; Goubin, G.; et al. Enamel Protein Regulation and Dental and Periodontal Physiopathology in Msx2 Mutant Mice. Am. J. Pathol. 2010, 177, 2516–2526. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hou, Y.; Chen, W.; Wang, J.; Xie, W.; Zhang, X.; Zeng, L. KIF9-AS1, LINC01272 and DIO3OS lncRNAs as novel biomarkers for inflammatory bowel disease. Mol. Med. Rep. 2017, 17, 2195–2202. [Google Scholar] [CrossRef] [Green Version]
- Hung, J.; Scanlon, J.P.; Mahmoud, A.D.; Rodor, J.; Ballantyne, M.; Fontaine, M.A.; Temmerman, L.; Kaczynski, J.; Connor, K.L.; Bhushan, R.; et al. Novel Plaque Enriched Long Noncoding RNA in Atherosclerotic Macrophage Regulation (PELATON). Arter. Thromb. Vasc. Biol. 2020, 40, 697–713. [Google Scholar] [CrossRef]
- Fu, H.; Zhang, Z.; Zhang, Z.; Qiu, H. LncRNA Pelaton, a Ferroptosis Suppressor and Prognositic Signature for GBM. SSRN 4001767. 2021. Available online: https://ssrn.com/abstract=4001767 (accessed on 15 September 2021).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Total (N) | Univariate Analysis | Multivariate Analysis | ||
|---|---|---|---|---|---|
| Hazard Ratio (95% CI) | p Value | Hazard Ratio (95% CI) | p Value | ||
| AMBN | 139 | 1.590 (0.468–5.400) | 0.458 | ||
| Pathologic stage | 132 | ||||
| Stage I | 106 | Reference | |||
| Stage II & Stage III | 26 | 0.210 (0.064–0.691) | 0.010 | 11,576,935.157 (1,535,060.618–87,309,534.264) | <0.001 |
| Clinical stage | 130 | ||||
| Stage I | 98 | Reference | |||
| Stage II & Stage III | 32 | 0.202 (0.071–0.576) | 0.003 | 0.000 (0.000–0.000) | <0.001 |
| Radiation therapy | 137 | ||||
| No | 113 | Reference | |||
| Yes | 24 | 0.873 (0.386–1.978) | 0.745 | ||
| Primary therapy outcome | 88 | ||||
| PR & PD | 14 | Reference | |||
| CR | 74 | 9.339 (1.264–68.994) | 0.029 | 155,601,452.260 (0.000–Inf) | 0.997 |
| Race | 134 | ||||
| Asian | 4 | Reference | |||
| Black or African American | 6 | 5.045 (0.520–48.936) | 0.163 | 6,123,473,977.844 (714,602,666.954–52,472,423,195.932) | <0.001 |
| White | 124 | 1.192 (0.163–8.728) | 0.863 | 246,966,750.563 (28,820,747.782–2,116,273,191.320) | <0.001 |
| Age | 139 | ||||
| <=30 | 67 | Reference | |||
| >30 | 72 | 0.697 (0.373–1.301) | 0.257 | ||
| Serum tumor markers(S) | 125 | ||||
| S0 | 43 | Reference | |||
| S1 | 41 | 1.944 (0.782–4.835) | 0.153 | 3.655 (1.527–8.750) | 0.004 |
| S2 | 36 | 2.908 (1.190–7.106) | 0.019 | 5.402 (2.230–13.087) | <0.001 |
| S3 | 5 | 3.993 (0.827–19.283) | 0.085 | 1.000 (1.000–1.000) | |
| Lymphovascular invasion | 135 | ||||
| No | 79 | Reference | |||
| Yes | 56 | 1.364 (0.733–2.538) | 0.327 | ||
| Testicular intratubular germ cell neoplasia | 130 | ||||
| Absent | 71 | Reference | |||
| Present | 59 | 0.729 (0.383–1.386) | 0.335 | ||
| History of undescended testis | 132 | ||||
| No | 109 | Reference | |||
| Yes | 23 | 0.560 (0.219–1.434) | 0.227 | ||
| Family history of testicular cancer | 122 | ||||
| No | 107 | Reference | |||
| Yes | 15 | 3.009 (1.407–6.435) | 0.005 | 1.637 (0.621–4.314) | 0.319 |
| Laterality | 134 | ||||
| Left | 74 | Reference | |||
| Right | 60 | 1.231 (0.639–2.371) | 0.534 | ||
| Pathologic T stage | 138 | ||||
| T1 | 80 | Reference | |||
| T2 & T3 | 58 | 1.158 (0.621–2.160) | 0.645 | ||
| Pathologic N stage | 64 | ||||
| N0 | 51 | Reference | |||
| N1 & N2 | 13 | 0.108 (0.015–0.793) | 0.029 | 1.000 (0.133–7.542) | 1.000 |
| Pathologic M stage | 124 | ||||
| M0 | 120 | Reference | |||
| M1 | 4 | 0.000 (0.000–Inf) | 0.996 | ||
| Clinical T stage | 118 | ||||
| T1 | 68 | Reference | |||
| T2 & T3 | 50 | 0.832 (0.433–1.596) | 0.579 | ||
| Clinical N stage | 113 | ||||
| N0 | 83 | Reference | |||
| N1 & N2 & N3 | 30 | 0.212 (0.074–0.608) | 0.004 | 1.000 (0.133–7.542) | 1.000 |
| Clinical M stage | 133 | ||||
| M0 | 125 | Reference | |||
| M1 | 8 | 0.624 (0.150–2.600) | 0.517 | ||
| Characteristic | Low Expression of GFAP | High Expression of GFAP | p |
|---|---|---|---|
| n | 67 | 67 | |
| Pathologic stage, n (%) | 0.303 | ||
| Stage I | 51 (40.2%) | 50 (39.4%) | |
| Stage II | 6 (4.7%) | 6 (4.7%) | |
| Stage III | 4 (3.1%) | 10 (7.9%) | |
| Clinical stage, n (%) | 0.370 | ||
| Stage I | 49 (39.2%) | 44 (35.2%) | |
| Stage II | 9 (7.2%) | 8 (6.4%) | |
| Stage III | 5 (4%) | 10 (8%) | |
| Primary therapy outcome, n (%) | 0.868 | ||
| PD | 0 (0%) | 1 (1.2%) | |
| SD | 0 (0%) | 0 (0%) | |
| PR | 5 (6%) | 8 (9.6%) | |
| CR | 31 (37.3%) | 38 (45.8%) | |
| Age, n (%) | 0.057 | ||
| ≤30 | 26 (19.4%) | 38 (28.4%) | |
| >30 | 41 (30.6%) | 29 (21.6%) | |
| Serum tumour markers(S), n (%) | 0.041 | ||
| S0 | 28 (23.3%) | 15 (12.5%) | |
| S1 | 16 (13.3%) | 22 (18.3%) | |
| S2 | 12 (10%) | 22 (18.3%) | |
| S3 | 2 (1.7%) | 3 (2.5%) | |
| Lymphovascular invasion, n (%) | 0.023 | ||
| No | 45 (34.6%) | 30 (23.1%) | |
| Yes | 21 (16.2%) | 34 (26.2%) | |
| Testicular intratubular germ cell neoplasia, n (%) | 0.246 | ||
| Absent | 38 (30.4%) | 30 (24%) | |
| Present | 25 (20%) | 32 (25.6%) | |
| History of undescended testis, n (%) | 0.258 | ||
| No | 47 (37%) | 57 (44.9%) | |
| Yes | 14 (11%) | 9 (7.1%) | |
| Family history of testicular cancer, n (%) | 1.000 | ||
| No | 52 (44.1%) | 52 (44.1%) | |
| Yes | 7 (5.9%) | 7 (5.9%) | |
| Laterality, n (%) | 0.938 | ||
| Left | 35 (27.1%) | 37 (28.7%) | |
| Right | 29 (22.5%) | 28 (21.7%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geng, T.; Heyward, C.A.; Chen, X.; Zheng, M.; Yang, Y.; Reseland, J.E. Comprehensive Analysis Identifies Ameloblastin-Related Competitive Endogenous RNA as a Prognostic Biomarker for Testicular Germ Cell Tumour. Cancers 2022, 14, 1870. https://doi.org/10.3390/cancers14081870
Geng T, Heyward CA, Chen X, Zheng M, Yang Y, Reseland JE. Comprehensive Analysis Identifies Ameloblastin-Related Competitive Endogenous RNA as a Prognostic Biomarker for Testicular Germ Cell Tumour. Cancers. 2022; 14(8):1870. https://doi.org/10.3390/cancers14081870
Chicago/Turabian StyleGeng, Tianxiang, Catherine Anne Heyward, Xi Chen, Mengxue Zheng, Yang Yang, and Janne Elin Reseland. 2022. "Comprehensive Analysis Identifies Ameloblastin-Related Competitive Endogenous RNA as a Prognostic Biomarker for Testicular Germ Cell Tumour" Cancers 14, no. 8: 1870. https://doi.org/10.3390/cancers14081870