
Long Non-Coding RNA (lncRNA) in Oral Squamous Cell Carcinoma: Biological Function and Clinical Application
by
 , ,
, ,
Jianfei Tang
1,2,3,4,5 ,
,
Xiaodan Fang
1,2,3,4,5, 
Juan Chen
1,2,3,4,5,
Haixia Zhang
6,* and
Zhangui Tang
1,2,3,4,5,* 1
Hunan Key Laboratory of Oral Health Research, Central South University, Changsha 410008, China
2
Hunan 3D Printing Engineering Research Center of Oral Care, Central South University, Changsha 410008, China
3
Hunan Clinical Research Center of Oral Major Diseases and Oral Health, Central South University, Changsha 410008, China
4
Xiangya Stomatological Hospital, Central South University, Changsha 410008, China
5
Xiangya School of Stomatology, Central South University, Changsha 410008, China
6
The Oncology Department of Xiangya Second Hospital, Central South University, Changsha 410008, China
*
Authors to whom correspondence should be addressed.
Cancers 2021, 13(23), 5944; https://doi.org/10.3390/cancers13235944
Submission received: 9 October 2021
/
Revised: 22 November 2021
/
Accepted: 24 November 2021
/
Published: 26 November 2021
(This article belongs to the Collection Regulatory and Non-Coding RNAs in Cancer Epigenetic Mechanisms)
Abstract
:Simple Summary
Increasing evidence has revealed the regulatory roles of long non-coding RNAs (lncRNAs) in the initiation and progress of oral squamous cell carcinoma (OSCC). As some novel lncRNA-targeted techniques combined with immune checkpoint therapies have emerged, they provide a new strategy for OSCC treatment. This review summarizes current knowledge regarding the involvement of lncRNAs in OSCC along with their possible use as diagnostic and prognostic biomarker and therapeutic targets.
Abstract
Oral squamous cell carcinoma (OSCC) is a type of malignancy with high mortality, leading to poor prognosis worldwide. However, the molecular mechanisms underlying OSCC carcinogenesis have not been fully understood. Recently, the discovery and characterization of long non-coding RNAs (lncRNAs) have revealed their regulatory importance in OSCC. Abnormal expression of lncRNAs has been broadly implicated in the initiation and progress of tumors. In this review, we summarize the functions and molecular mechanisms regarding these lncRNAs in OSCC. In addition, we highlight the crosstalk between lncRNA and tumor microenvironment (TME), and discuss the potential applications of lncRNAs as diagnostic and prognostic tools and therapeutic targets in OSCC. Notably, we also discuss lncRNA-targeted therapeutic techniques including CRISPR-Cas9 as well as immune checkpoint therapies to target lncRNA and the PD-1/PD-L1 axis. Therefore, this review presents the future perspectives of lncRNAs in OSCC therapy, but more research is needed to allow the applications of these findings to the clinic.
1. Introduction
OSCC originates in the surface of oral mucosal epithelium. It is one of the most commonly diagnosed lethal malignancies with poor prognosis worldwide. According to the Global Cancer Statistics 2020, about 53,260 new cases and 10,750 deaths of oral cancer were calculated in the USA, accounting for approximately 4% of all male cancer cases [1], and the most recent data indicate that the overall cases will rise to 510,948 in 2035 [2]. To date, surgical treatment combined with radiation and chemotherapy remains the primary approach for management of OSCC [3]. Advancements in treatment strategies such as drug development and computer-assisted surgery were expected to lead to an improved survival in OSCC patients. However, OSCC remains an incurable malignance and treatment-associated outcomes are still unchanged. In addition, most patients with OSCC are diagnosed at advanced stages and no early screening strategy has proven to be effective. Therefore, to further improve efficiency of diagnosis and acquire a better prognosis, comprehensive investigation aiming to elaborate molecular mechanisms of OSCC and to discover novel diagnostic tools and precision therapeutic approaches are still urgently needed.
Long non-coding RNAs (lncRNAs) is a novel class of RNAs with a length longer than 200 nucleotides. These RNAs cannot encode proteins or peptides [4,5]. In 1989, a study found that H19 was the first lncRNA that is highly expressed in mouse embryo and neonatal liver. The mouse H19 gene shared high homology (77%) with human H19 gene, though the specific function of the gene remains unclear [6]. Therefore, lncRNAs were mainly considered as “junk transcripts” that do not generate any function for many years. The functional research of lncRNAs has endured a long history. Luisa et al. confirmed that the H19 gene acts as a trans regulator gene of the imprinted gene network (IGN) such as Igf2 and other imprinted genes in the mouse embryo [7]. Up to now, lncRNA microarray and whole-genome transcriptome have identified more than 50,000 lncRNAs, some of which have been functionally characterized and revealed tissue-specific expression patterns [8]. With regard to their biological role in multiple human cancers including OSCC, many studies have shown that lncRNAs perform modulatory functions that influence cell biological behaviors, immune response, and transformed phenotype in cells. For example, lncRNA HOTAIR was reported as an oncogene that was pervasively overexpressed in most solid cancers including oral cancer and acts to promote OSCC invasion and metastasis [9]. THRIL is an immunoregulatory lncRNA that was shown to regulate the expression of pro-inflammatory cytokine TNF-α in Kawasaki disease and other immune-related inflammatory diseases through interacting with hnRNPL forming an RNA—protein complex to bind TNF-α promoter [10]. A study reported that the lncRNA loc100506114 contributes to the functional transformation of fibroblasts to cancer-associated fibroblasts (CAFs) in OSCC [11]. Moreover, lncRNAs are richly distributed in body fluids including blood, urine, saliva even exosomes, therefore they could be regarded as a type of non-invasive biomarker [12]. LncRNAs are functional transcripts that will help to identify various cancer characteristics and hallmarks, and further become an attractive potential therapeutic target.
In general, precise diagnosis and individual treatment is the ultimate direction in which we would like to explore, and lncRNAs have significant potential clinical values for diagnosis and treatment of OSCC. Therefore, this review summarizes the functions and molecular mechanisms of lncRNA in OSCC. In addition, we highlight the crosstalk between lncRNA and the tumor microenvironment, and discuss the potential applications that lncRNAs can serve including diagnostic and prognostic tools as well as therapeutic targets in OSCC.
2. Roles of LncRNA in OSCC
In the past few decades, due to the emergence of high-throughput sequencing technologies, exponential growth in the number of lncRNAs with aberrant expression have been confirmed by RNA-Seq and lncRNA-microarray profiling in various cancers [13]. Fang et al. performed RNA-Seq to profile lncRNA expression in five pairs of OSCC tissues and adjacent-normal tissues; 2915 lncRNAs were significantly differentially expressed, and of these lncRNAs, 11 were associated with OSCC metastasis [14]. Based on The Cancer Genome Atlas (TCGA) database, RNA sequencing analysis of 523 oral cancer samples in India by Ganesan Arunkumar identified 11 dysregulated lncRNA in OSCC that are closely related to tobacco chewing/smoking history [15]. Evidence also showed that different functional studies revealed the role of lncRNAs in oncogenesis, tumor-suppression, and chemoresistance as well as governing virtually every physiological cell process (Figure 1). Abnormal lncRNAs are involved in many aspects of cancer cell processes including cell proliferation, apoptosis, invasion and metastasis [16], epithelial–mesenchymal transition (EMT), and drug resistance [17]. Moreover, lncRNAs even affect the outcome of patients such as lymph node metastasis, distal metastasis, and postoperative recurrence [18]. Thus, in this part, we summarize the current dysregulated lncRNAs in OSCC and elaborate on the functions of these lncRNAs in OSCC (Table 1).
2.1. Oncogenic Function of LncRNAs in OSCC
According to previous studies, a majority of dysregulated lncRNAs exhibit a trend of upregulated expression and function as an oncogene in promoting malignant biological behaviors in OSCC including cell proliferation, migration, metastasis, and angiogenesis. However, lncRNAs inhibited apoptosis and the process of the cell cycle. For example, many lncRNAs such as H19 [47], OIP5-AS1 [85], DNM3OS [14], AFAP1-AS1 [21], ADAMTS9-AS2 [20], LINC00668 [67], and BANCR [23] were confirmed to be overexpressed in OSCC cells and promoted tumor development by enhancing the proliferation and migration in vitro and in vivo. Beyond this, lncRNAs were also involved in the regulation of the cell cycle and inhibiting apoptosis in OSCC. For example, when the lncRNA LEF1-AS1 was silenced, it caused the arrest of the G0/G1 cell cycle and suppressed cell proliferation and growth in vitro via inactivation of the Hippo signaling pathway [59]. Furthermore, some certain lncRNAs were verified to promote OSCC invasion, metastasis, and angiogenesis [62,103]. For example, MALAT-1 is closely related to the growth and metastasis of OSCC cells and through the regulation of target small proline-rich protein (SPRR) in order to promote distant metastasis [77]. LINC00319 is downstream of Chemokine ligand 18 (CCL18), and overexpression of LINC00319 regulated the expression of VEGFA and MMP-9 to promote the angiogenic ability of OSCC cells [62]. Similarly, knockdown of FOXCUT led to the downregulation of angiogenesis factor VEGFA in Tca8113 and SCC9 cells, which indicated the potential function for FOXCUT in angiogenesis of OSCC [43].
2.2. Tumor-Suppressor Function of LncRNAs in OSCC
LncRNA can be used not only as an oncogene to promote the occurrence and development of tumors, but also as a suppressing factor to inhibit the growth and metastasis [104]. According to previous studies, lncRNA NKILA has been reported to be a tumor suppressor, which has been negatively correlated with metastasis and prognosis in breast cancer [105]. Consistent with the results in breast cancer, Huang et al. confirmed that NKILA expression levels in tongue squamous cell carcinoma (TSCC) was expressed significantly less. High expression of NKILA represses EMT and migration in Tscca and CAL27 cells via activation of the NF-κB/Twist signaling pathway to regulate the biological process of TSCC [84]. Growth-arrest-specific transcript 5 (GAS5) is another representative lncRNA that has been widely reported as a tumor suppressor in many cancers; Zeng et al. also confirmed that GAS5 functions as a tumor suppressor in OSCC via the miR-21/PTEN axis to inhibit tumor cell proliferation, migration, invasion, and EMT [46]. Moreover, maternally expressed gene 3 (MEG3) is an acknowledged tumor suppressor that has widely been investigated in cancers including OSCC. The overexpression of MEG3 decreases proliferation and migration of SCC15 while inducing CAL27 apoptosis. Mechanistically, MEG3 could exert the tumor-suppressor function not only by blocking the WNT/β-catenin signaling pathway, but also by acting as a miRNA sponge of miR-548d-3p to modulate the JAK–STAT signaling pathway [80,81]. Furthermore, other lncRNAs such as FALEC was also reported as a tumor-suppressor with low expression in OSCC, and the overexpression of FALEC significantly repressed OSCC cell proliferation and migration both in vitro and in vivo, and this predicts a good prognosis in OSCC patients [40]. Compared to the oncogenic functions of lncRNAs, the studies of lncRNA action as a tumor suppressor in order to exert inhibitive functions in OSCC are limited. Therefore, relevant research in OSCC requires comprehensive investigation.
2.3. OSCC LncRNAs Regulate Chemoresistance and Radiosensitivity
Adjuvant radiation or chemotherapy plus radiation has been the primary approach for the treatment of OSCC patients depending on the disease stage [3]. Cisplatin (CDDP) is a platinum-based drug that is commonly used as an efficient adjuvant treatment for OSCC patients. However, cisplatin-resistance is a headache in chemotherapy, resulting in tumor relapse and poor prognosis [53]. Emerging evidence showed that lncRNAs may function as vital regulators of chemoresistance in OSCC [17,106]. For instance, the upregulation of HOXA11-AS significantly increased resistance to cisplatin and tumor cell growth, and the knockdown of HOMA11-AS markedly enhanced CDDP-mediated tumor inhibition in vivo [53]. Likewise, Lin et al. reported the increased expression of CILA1 in cisplatin-resistant OSCC cells lines, and the silence of CILA1 significantly inhibited the migration, invasion, and EMT, while increasing the sensitivity to chemotherapy of these cells [36]. Furthermore, another study shown by Fang et al. indicated that UCA1 increased the proliferation of OSCC cells and induced cisplatin resistance by modulating the expression of the miR-184 target gene SF1 [101]. Notably, Zhang et al. confirmed that overexpression of lncRNA ANRIL can be induced by paracrine action of CAF-derived Midkine, thereby enhancing the proliferation and resistance to cisplatin of tumor cells [22]. Interestingly, Wang et al. confirmed that silencing HOTAIR significantly enhanced sensitivity to CDDP while inhibiting tumor cell autophagy [51]. Moreover, other lncRNAs such as KCNQ1OT1 [56,57]. LHFPL3-AS1 [60] and PVT-1 [90] were also reported to be involved in promoting cisplatin-resistance in OSCC cells.
In addition to chemoresistance, radiosensitivity is another tremendous challenge among the comprehensive therapies for OSCC. Accumulating evidence also revealed the crucial roles of lncRNAs in radiotherapy. For example, Gou et al. observed that lncRNA BLACAT1 was associated with low radiosensitivity and poor outcomes of HNSCC patients, and they further confirmed that the knockdown of BLACAT1 in SCC25 cells markedly improved the radiosensitivity by regulating PSEN1 [24]. Likewise, another example is LINC00473, which is highly expressed in OSCC cells. LINC00473 knockdown in these cells significantly enhanced the sensitivity of radiotherapy by modulating the Wnt/β-catenin signaling pathway [65].
3. Molecular Mechanism of LncRNAs in OSCC
In the past few years, research has revealed that lncRNAs could potentially be involved in the tumorigenesis of cancer, and identifying the molecular mechanisms of lncRNAs within OSCC is still a challenge. In general, a myriad of studies have confirmed that lncRNAs exert regulatory functions, mainly via the following mechanisms: (1) epigenetic regulation; (2) transcriptional regulation; and (3) post-transcriptional regulation [107,108]. Mechanistic studies published to date indicate that the dysregulated lncRNA in OSCC may exert their biological functions through these molecular mechanisms (Figure 1).
3.1. Epigenetic Regulation
Epigenetic regulation is a complex process, which is mediated by DNA and histone modifications, and are crucial for transcription machinery during gene expression [107,109]. Recent studies have clearly suggested that epigenetic changes can contribute to the development of several human malignancies and that lncRNAs play a vital role in this context. A battery of lncRNAs have been reported to mediate chromatin remodeling and DNA modification by acting as a molecular scaffold, thereby regulating the expression of genes before finally affecting cancer development [110].
A typical example is the association with polycomb repressive complexes 2 (PRC2), which is composed of EED, SUZ12, and EZH2 [111]. It can catalyze H3K27 trimethylation, causing chromatin compaction and thereby affecting the transcriptional state. To date, lncRNA HOTAIR has been shown to promote migration, invasion, and poor survival in OSCC. HOTAIR could act as a molecular scaffold to recruit EZH2 and H3K27me3, thereby repressing E-cadherin expression [9]. Another study verified that lncRNA FALEC could also recruit PRC2 component EZH2, causing H3K27me3 trimethylation and ECM1 silencing, leading to the inhibition of proliferation and migration in OSCC cells [40]. Furthermore, dysregulation of chromatin modification can modulate lncRNA transcription, cancer initiation, and malignant progression. One recent study indicated that lncRNA PLAC2 is transcriptionally activated by CBP-mediated H3K27 acetylation at the promoter region and promoted OSCC progression via activation of the Wnt/β-catenin signaling pathway [89]. There are still other patterns of epigenetic modifications mediated by lncRNAs in OSCC urging further exploration.
3.2. Transcriptional Regulation
In addition to the above features of lncRNA epigenetic regulation, lncRNAs could also regulate gene expression in a transcription-dependent manner, which leads to cancer progression [111]. Recent studies indicate that lncRNAs can regulate transcription to recruit TFs or by directly binding to the gene promoters, thereby regulating different malignant biologic behaviors of tumors. On one hand, lncRNAs interact with key cancer associated TFs to regulate the activity of transcription. One study reported that tumor suppressor lncRNA NKILA hindered OSCC migration and invasion by interacting with NF-κB and ultimately lowering Twist and E-cadherin [84]. On the other hand, by interacting with DNA-binding proteins including CTCF, lncRNAs can facilitate their binding and expression at a targeted gene region. The DNA-binding protein CTCF can mediate chromatin interaction and DNA looping [112]. This type of mechanism can be observed in LINC00941, which drives CTCF recruitments to promote CAPRIN2 expression, thereby accelerating cell proliferation and colony formation in OSCC [69]. However, in recent studies, scholars have reported that some enhancers can also produce enhancer RNA (eRNA) through transcription, which can modulate genes that are far away from a specific direction, but the specific relationship between enhancer genomic loci and gene expression regulation remains unclear, especially in OSCC; related studies need to be further explored [107].
3.3. Post-Transcriptional Regulation
Emerging reports have indicated that lncRNA functions as a post-transcriptional regulator in the gene expression process. The mechanisms of lncRNA in post-transcriptional regulation mainly include the alteration of mRNA splicing, miRNA sponge, mRNA stability, protein translation, and RNA editing, even in the regulation of signaling pathways [4]. lncRNA regulated phosphorylation of critical signaling molecules have been reported to contribute to cancer progression. For instance, lncRNA LEF1-AS1 could interact with the LATS1 protein, leading to attenuated YAP1 phosphorylation that ultimately promotes cell proliferation and migration in OSCC. Further studies have demonstrated that depletion of lncRNA LEF1-AS1 results in upregulation of cytoplasmic YAP1 expression and downregulation of the nuclear YAP1 level, suggesting the involvement of lncRNA LEF1-AS1 in the regulation of phosphorylation of the crucial players of the Hippo signaling pathway [59]. On the other hand, lncRNAs regulate mRNA stability via post-translational modifications that support cancer development. A recent study showed that LINC00284 facilitates the mRNA stability of KAZN by binding with RBP FUS and promoting cell proliferation and migration in OSCC [61]. Notably, lncRNA FOXC1 associates and forms a lncRNA–mRNA duplex with FOXCUT and thereby increases its stability to promote OSCC cell migration [43]. Likewise, lncRNA CEBPA-AS1 could bind with CEBPA mRNA, thus leading to OSCC cell proliferation, invasion, and migration [35].
In addition, lncRNA was shown to function as a competing endogenous RNA (ceRNA) and regulate gene expression at the posttranscriptional level, consequently influencing cancer progression [113]. In detail, lncRNA could use its miRNA response elements (MREs) in mRNA binding site as a natural decoy for miRNA and inhibits the ex-pression of miRNA on the target gene (mRNA) [113,114,115]. Recently accumulating evidence has confirmed the role of lncRNA in regulating the pathogenesis of OSCC via miRNA sponge. For example, lncRNA RC3H2 facilitates OSCC tumor growth and metastasis by acting as a ceRNA for miRNA-101-3p. Mechanistically, lncRNA RC3H2 can compete with miRNA-101-3p for binding with the target EZH2 to promote OSCC malignant behavior [92]. Another ceRNA, lncRNA H19, acts by competitively sponging miR-138 and upregulating vimentin and N-cadherin expression, leading to EMT in OSCC [47]. Additionally, lncRNA KCNQ1OT1 promotes cisplatin resistance of OSCC by functioning as a molecular sponge for miR-211-5p to activate Ezrin/Fak/Src signaling [56]. Other lncRNAs such as LTSCCAT [76], OIP5-AS1 [85], DNM3OS [14], SNHG16 [95], HOTTIP [52], HCP5 [50], JPX [55], and AFAP1-AS1 [21] were also reportedly involved in promoting the progression of OSCC via the ceRNA mechanism. In summary, lncRNAs could act as ceRNAs to sponge various miRNAs to participate in post-transcription, thus promoting OSCC malignant development.
Collectively, it is important to note that individual lncRNAs could exert their functions through different modes of post-transcriptional regulation simultaneously. The regulation of other posttranslational modifications including pre-mRNA alternative splicing by lncRNAs has not been substantiated in OSCC. Future work will be aimed at elucidating the mechanism of action of lncRNAs that use other posttranslational pathways in contributing to OSCC development.
4. The Crosstalk between LncRNA and Tumor Microenvironment in OSCC
TME is mainly composed of parenchyma cells, immune cells, peripheral extracellular matrix (ECM) as well as some signal molecules [116]. TME of OSCC is characterized by hypoxia, chronic inflammation, and immunosuppression. This surrounding environment is regarded as an intricate physical and biochemical system, which is involved in tumor onset, progression, metastasis, and influences the prognosis of treatment. In particular, recent emerging studies have indicated that abnormal expression of certain lncRNAs is strongly associated with hypoxia, metabolism, immune cells, and CAFs; therefore, these lncRNAs have a crucial role in TME (Figure 2).
4.1. LncRNA and Hypoxic, Metabolic TME in OSCC
Hypoxia is a common and important feature in the TME and is tightly linked to cancer development and aggressive phenotypes [117]. Under hypoxic TME, the vital regulator Hypoxia-induced factor-1 alpha (HIF-1α) mediates tumor growth, invasiveness, and metastasis, contributing to aggressive phenotypes in various cancers including OSCC [82]. Despite hypoxic response signaling having been extensively explored, the involvement of lncRNAs in the hypoxic response has become a new focus of cancer research. It is well established that 56 hypoxia-associated lncRNAs (HALs) have led to cancer progression [117]. HALs have been analyzed in OSCC research and found to be associated with worse outcome and clinicopathological characteristics [82]. Zhu et al. found that lncHAS2-AS1 was substantially increased in OSCC. In response to hypoxic conditions, HIF-1α drove the expression of lncHAS2-AS1, which caused HAS2 accumulation. Moreover, the study also demonstrated that lncHAS2-AS1 promoted the EMT and invasion potential in OSCC [49]. Notably, expression of HALs also regulate HIF-1α activity. Shih et al. showed that the expression level of lncHIFCAR is significantly elevated in OSCC, which is associated with tumor grade as well as poor overall survival and recurrence-free survival. In response to hypoxic conditions, lncHIFCAR directly binds with HIF-1α and further facilitates HIF-1α target expression, leading to increased invasion, metastasis, metabolic reprogramming, and sphere-forming ability in vitro and in vivo [82].
Notably, recent studies have revealed that some lncRNAs were involved in reprogramming metabolism, especially modulating glycolysis in OSCC cells, resulting in the progression of malignant behaviors such as proliferation and metastasis [39,86,118]. A representative example is lncRNA-p23154. It enhances metastasis in OSCC through binding with the promoter region of miR-378a-3p in the nucleus in order to promote glucose transporter 1(GLUT1) expression, leading to enhanced glycolysis [86]. Moreover, ELF3-AS1 is another typical lncRNA, which is upregulated in OSCC; the silencing or forced expression of ELF3-AS1 in OSCC cells resulted in the same trend of GLUT1 expression; ELF3-AS1 and GLUT1 overexpression leads to a significantly increased proliferation rate of OSCC cells and glucose uptake, which means that upregulation of ELF3-AS1 promotes the proliferation of OSCC cells, and may positively regulate GLUT1 to affect glucose metabolism [39]. Likewise, Yang et al. reported that H-19 was associated with glycolysis in oral CAFs via the interaction of H19-derived miR-675-5p binding to PFKFB3, resulting in the proliferation and migration of OSCC cells [118].
4.2. LncRNA and Cancer-Associated Fibroblasts
CAFs are important and abundant components within TME, which interact closely with tumor cells by a paracrine mode of action, thus contributing to tumor initiation and malignant progression [119,120]. It has been reported that lncRNAs are involved in and sustain this interaction, and this effect on the TME has aroused extensive attention.
Under the action of lncRNA, normal fibroblasts (NFs) were activated and acquired CAF phenotype, which in turn promotes many aggressive features in cancer including OSCC. Ding et al. revealed that upregulated FLJ22447 maintains the stromal phenotype of CAF via IL-33, thus promoting tumor proliferation [42]. Moreover, other examples of lncRNAs playing a key role in CAFs within OSCC are TIRY and loc100506114, and have been described as being elevated in CAFs when compared to the adjacent NFs. Two studies demonstrated that TIRY and loc100506114 participated in the functional transformation of human NFs to the phenotype of CAFs, which supports tumor cell growth, invasion, and metastasis [11,97].
Interestingly, CAFs have been reported to induce upregulation of lncRNA in tumor cells. In this regard, Ding et al. showed that OSCC cells expressed FLJ22447 in an exosomal manner to activate adjacent NFs, thereby facilitating elevated FLJ22447 to obtain the phenotype of CAFs, which form a positive feedback loop to promote OSCC development, which are associated with short survival and poor prognosis in OSCC patients [42]. In another example, Midkine secreted by CAFs promoted the upregulation of lncRNA ANRIL in OSCCs. Such lncRNA was reported to enhance cisplatin-based chemoresistance [22]. Additionally, Yang et al. reported lncRNA H19 as potential epithelial–mesenchymal common targets (EMCTs) and is involved in regulating glycolysis, proliferation, and migration in oral CAFs via the miR-675-5p/PFKFB3 pathway [118]. Altogether, lncRNA has been shown to play a role in the crosstalk between CAFs and OSCC cells and may be regarded as a possible therapeutic target or predictive biomarker.
4.3. LncRNA and Cancer-Associated Immune Cells in OSCC
Infiltrated immune cells such as T cells, tumor-associated macrophages (TAMs), dendritic cells (DCs), and natural killer cells (NKs) are also key components of TME [3]. The reciprocal crosstalk between cancer cells and immune cells shapes the pro-tumorigenic microenvironment in a way that renders it to escape immune surveillance and suited for immune tolerance [121]. LncRNAs are reported to participate in various processes of immune response within TME to promote tumor progression [122]. In OSCC patients, Feng et al. found that lncRNA SLC16A1-AS1 was positively correlated with resting NK cells, M1 macrophages, activated mast cells, and activated memory CD4 T cells by bioinformatics analysis, but negatively correlated with plasma cells, T follicular cells, resting mast cells, and Tregs [93]. Likewise, Li et al. reported that LINC02195 was an immune-related lncRNA that was upregulated in OSCC cells. A positive correlation can be seen between increasing number of infiltrating CD8+T, CD4+T cells, and LINC02195 by bioinformation analysis. In addition, LINC02195 acts as a regulator, which was closely associated with high expression of the HLA I gene, thereby regulating the MHC I protein to show the potential function in affecting immunosurveillance [75]. Moreover, the silence of lncRNA TUG1 substantially enhanced NK cells, killing sensitivity in OSCC cells [123]. Notably, lncRNA LBX1-AS1 was significantly upregulated in the exosomes, which are derived from RBPJ overexpressed-macrophages in OSCC, and lncRNA LBX-AS1 inhibits tumor growth and invasion. Furthermore, this effect can be attenuated by lncRNA LBX1-AS1 knockdown [58]. In addition, lncRNAs can impact the function and cytotoxicity of T cells via regulation of the expression of molecules on the surface of the tumor cells and directly inducing cell death or enhancing T cell exhaustion in the TME [124]. For example, LNC-SOX5, which is associated with carcinogenesis of tongue carcinoma, has also been reported to function on the regulating cytotoxicity of CD8+ T cells in colorectal cancer [54,125], while at present, the function of LNC-SOX5 in regulating the CD8+ T cell cytotoxicity effect in OSCC remains unclear. Furthermore, Liu et al. also reported that lncRNA FOXD2-AS1 was involved in regulating the proliferation and functions of antigen-presenting cells (APCs) to inhibit the adaptive immunity in OSCC [44]. The above evidence indicates the significance of lncRNAs in immunotherapy and it could be the potential immunotherapy target. However, only a few lncRNAs have been reported in the crosstalk with immune cells in OSCC currently, and comprehensive study remains deficient.
4.4. Extracellular Vesicles: Exosome-Associated LncRNAs in TME
Exosomes refer specifically to a kind of extracellular vesicle (EV) that is secreted by most eukaryotic cells [126]. Currently, due to their unique functions in mediating intercellular communication and activating signaling as extracellular messengers, exosomes as effective signaling molecules have been broadly investigated in cancers [127,128]. The diverse cargo in which exosomes carry such as lncRNA are released from exosomes and will dynamically change depending on the cell type. These lncRNAs are involved in regulating tumor metastasis, angiogenesis, immunosuppression, drug resistance, and metabolism [129,130], thereby enhancing the interaction between cancer cells and surrounding cells [131]. For instance, in the studies of OSCC, FLJ22447 and LBX1-AS1 were representative exosome-associated lncRNAs that were derived from CAFs and macrophages, respectively. Both of these are strongly associated with OSCC progression, recurrence, and poor prognosis [42,58]. Analysis of FLJ22447 derived from CAFs revealed it as a medium in which to interact with surrounding stromal fibroblasts to exert the modulatory function in activating CAFs via IL-33, thus inducing the proliferation of OSCC cells [42]. In contrast, combined analysis of LBX1-AS1, which is secreted from RBPJ overexpressed macrophages in OSCC patients, the overexpression of exosome-associated lncRNA LBX1-AS1 upregulated tumor suppressor gene FOXO3 to inhibit the proliferation and invasion of OSCC cells [58]. These studies suggested that exosomes, especially exosome-associated lncRNA, probably exert modulation functions in OSCC development and could be a potential biomarker for OSCC diagnosis and therapy. However, exosome-associated lncRNA has been a novel topic in recent years. Some lncRNAs such as LINC01133 have been reported to act as a tumor repressive gene in OSCC cells [71], while LINC01133, derived from exosomes only, have been reported in pancreatic tumors and bladder urothelial carcinoma [132,133].
Current research has already revealed the roles of exosome-associated lncRNAs from different cells in OSCC patients. Therefore, exosome-associated lncRNAs might be potential biomarkers for the diagnosis and therapy of OSCC. Furthermore, compared with conventional targeting vectors, exosomes have lower systemic toxicity and higher stability, and exhibit nonimmunogenic properties [134]. For this purpose, the introduction of engineered lncRNAs into OSCC cells or tissues might represent a new and efficient approach for future cancer therapy.
5. Translational Potential of LncRNAs in OSCC
The presence of numerous lncRNAs that may have roles in cancer progression and outcomes have important clinical implications. On one hand, lncRNAs are stable and widely distributed in various tissues and body fluids including blood, saliva, and urine, making them a promising noninvasive biomarker for cancer diagnosis and prognosis [135]. On the other hand, lncRNAs have highly tissue-specific expression patterns and are functionally characterized, which contribute to the hallmarks of cancer [136]. Therefore, they are potential therapeutic targets. Here, we present the translational potential of lncRNAs in OSCC (Figure 3).
5.1. LncRNAs as a Novel Diagnostic and Prognostic Tools
Studies have confirmed that body fluids can detect the dysregulated lncRNA from primary tumors [135]. A representative lncRNA is prostate Cancer Antigen 3(PCA3, also known as DD3), which is derived from the patient’s urine and is widely applied in the diagnosis of prostate cancer due to its high specificity and sensitivity [137,138,139]. With regard to OSCC, Jia et al. confirmed that the expression profile of plasma lncRNAs in OSCC patients are changed by microarray analysis [140], which suggested the potential diagnostic value of circulating lncRNAs in OSCC. For instance, the level of CASC2 significantly decreased in the plasma of OSCC patients with local recurrence while it increased in those without recurrence; receiver operating characteristic curves (ROC) showed a significant value in detecting the expression of plasma CASC2 for OSCC diagnosis (the area under the ROC curves (AUC) = 0.8445) [28]. CASC15 is another lncRNA that was upregulated in the plasma of patients with OSCC, and this feature can also be applied to distinguish OSCC patients from oral ulcer patients [26]. In parallel, a study by Shao et al. identified lncAC007271.3, a kind of serum lncRNAs with a high expression level in OSCC patients when compared to the classic tumor markers SCCA, the ROC curves illustrated that the level of serum AC007271.3 could effectively discriminate between OSCC patients and controls (AUC = 0.873; 95% confidence interval (CI), 0.815–0.931; p < 0.001) with high sensitivity and specificity (77.6% and 84.5%, respectively) [141]. Moreover, a study by Tang et al. revealed that saliva in its entirety contains detectable amounts of certain lncRNAs such as HOTAIR and MALAT1 [12]. Interestingly, HOTAIR was differentially expressed in the saliva of OSCC patients with metastasis compared to those who without metastasis [12], which indicates the prospect of detecting lncRNAs in saliva to act as a rapid and noninvasive tool for OSCC diagnosis.
LncRNAs have also been indicated to be closely related to a series of clinic pathological parameters such as lymph metastasis and local recurrence in OSCC and may serve as valuable predictive biomarkers. The expression level of LINC00152 is increased in OSCC tissues and is positively correlated with cervical lymph node metastasis, higher TNM stage, and postoperative recurrence [8]. Similarly, other lncRNAs such as H19, CCAT2, and TUG1 have also been reported to have high expression levels in OSCC and are associated with the TNM stage and pathological grade [34,47,99]. In particular, one study reported that LINC-RoR was associated with cellular differentiation in OSCC, and was highly expressed in tumors with undifferentiated pathology and served as a predictor to therapeutic response [15]. Furthermore, a study by Jin et al. reported increased TIRY expression in OSCC tissues. Authors used ROC and diagnostic evaluation tests to reveal that OSCC patients with different risk of recurrence or metastasis within one year could be distinguished by TIRY expression (AUC = 0.897), which indicates the potential diagnostic value of TIRY [97].
In addition to the diagnostic potential of lncRNAs, previous studies have already confirmed that lncRNAs are associated with the survival time of OSCC patients and regarded as prognostic biomarkers. For example, MALAT1 was highly expressed in OSCC tissues and Zhou et al. verified that the lower expression of MALAT1 in OSCC patients had a better survival rate computed by Kaplan–Meier analysis [78]. Likewise, Yao et al. observed markedly upregulated BANCR expression in OSCC tissues; more importantly, a multivariate proportional hazards (COX) regression analysis revealed that in addition to lymph node metastasis, BANCR expression level was independently associated with poor overall survival (OS) and disease-free survival (DFS), which suggests BANCR was an independent prognostic factor in OSCC patients [23]. The same as BANCR, Yang et al. reported that the average OS of OSCC patients with low CASC9 expression was longer than those with high CASC9 expression by Kaplan-analysis, and COX regression analysis also revealed that the CASC9 expression level was an independent predictor of the OSCC prognosis [29]. In addition, similar reports of other lncRNAs such as LINC01234 [73,74], colon cancer–associated transcript 2 (CCAT2) [87], FOXD2-AS1 [44] and FTH1P3 [45] were also confirmed to be unregulated in OSCC tissues and associated with low OS, which indicated the poor prognostic capability.
However, tissue biopsy is still considered the gold standard for cancer diagnosis, even though it is an invasive procedure. The main advantages of lncRNAs as a biomarker for cancer diagnosis and prognosis is due to the high stability, high sensitivity, specificity, and non-invasive nature during body fluid circulation [135]. Due to the features of specificity in diseases and cell types, it is much easier to detect and make lncRNAs that are suitable for diagnosis in cancer patients. LncRNAs serve as valuable biomarkers in applications such as diagnosis and prognosis and have been shown to have tremendous potential in the future.
5.2. LncRNAs as Therapeutic Agents or Targets
Given the fact that the expression of lncRNAs is tissue/cell specific, and the carcinogenic roles of these lncRNAs are diverse, lncRNAs show promise as attractive targets for drug development and significant implication in clinical application for cancer treatment. Therapeutic approaches that accurately target lncRNAs may exert anti-tumor effects. Emerging advanced lncRNA-based techniques such as antisense oligonucleotides (ASOs), small interfering RNAs (siRNAs), short hairpin RNAs (shRNAs), aptamers, and CRISPR-Cas9, have revealed that targeting specific lncRNAs can effectively inhibit OSCC growth, migration, and invasion in vitro and in vivo [142].
Conventional tools such as siRNAs and shRNAs have already been extensively employed to study the mechanisms and therapy of lncRNAs. For example, intratumor delivery of MALAT1-targeted siRNA substantially inhibits tumor growth in the Tscca xenograft mouse model [78]. Knockdown of lncRNA LINC00460 by siRNA-based targeting method siLINC00460 led to inhibition of tumor metastasis in lung metastasis models in OSCC [63]. Furthermore, Wang et al. also demonstrated that tail vein injection of an shRNA specifically targeting lnc-p23154 significantly reduced tumor metastasis in the OSCC mouse model [86]. Interestingly, shRNA-mediated knockdown of lncRNA KCNQ1OT1 can increase cisplatin sensitivity as well as decrease tumor burden in OSCC xenografts, suggesting a novel potential approach for the reversion of cisplatin resistance in OSCC [56]. However, studies on siRNA technology specificity have shown off-target effects [143].
ASO techniques are the most powerful approach to target lncRNAs. For example, ASO-mediated FOXD2-AS1 silence has been shown to inhibit tumor growth in a OSCC mouse [44]. Furthermore, Li et al. observed that tumor growth and metastasis was significantly suppressed after AC104041.1 specific LNA-ASO in patient-derived xenograft (PDX) models generated from HNSCC patients [19]. However, off-target effects can be observed when targeting lncRNAs using ASO therapeutics, which can be seen in the generally low abundance of lncRNAs in vivo [142]. Despite these obstacles, protein-coding genes have been targeted using ASOs. An ASO targeting STAT3 (AZD9150) was tested for antitumor activity in patients with refractory lymphoma and lung cancer (NCT01563302) [144]. Therefore, the ASO technique of targeting lncRNAs for cancer treatment may be hopeful. Surprisingly, the CRISPR–Cas9 system has transformed the way lncRNAs are studied and has offered new opportunities for therapeutic targeting of lncRNAs in cancer research [145]. For instance, Zhang et al. confirmed that genomic deletion of lncRNA XIST using the CRISPR/Cas9 system reduced the tumor formation in tongue cancer [146]. In parallel, according to a study by Chang et al., CRISPR/Cas9 targeting MIR31HG markedly presented a reduction in oncogenicity of OSCC, which shows the potential therapeutic efficacy by targeting MIR31HG [83]. Thus, CRISPR/Cas9 may open a bright pathway for tumor therapy for gene-level tumor treatments considering its higher stability and lower off-target effect. So far, no lncRNA-targeting therapeutics have entered clinical development, and personalized and targeted therapy based on CRISPR/Cas9 is likely the future of tumor therapy.
Beyond the above mentioned statements, anti-PD-1/PD-L1 therapy has recently been approved for use in recurrent and/or metastatic OSCC patients, however, quite a number of patients are resistant to immune therapy [147]. Therapy targeting these lncRNAs may regulate the resistance within patients with OSCC to anti-PD-1/PD-L1 immunotherapy. LncRNA as a potential upstream regulator can target the PD-1/PD-L1 axis, thereby leading to marked anti-tumor activity [148]. Studies have reported that SNHG20 promotes esophageal squamous cell carcinoma growth and metastasis by activating the ATM/JAK/PD-L1 pathway [149]. GATA3-AS1 enhances the ubiquitination of PD-L1 by upregulating COPS5, thus promoting the immune escape of breast cancer cells [150]. Remarkably, interventional methods targeting UCA1 and anti-PD-1 treatment enhance therapy efficacy for bladder cancer [151]. More importantly, Ma and their team reported IFNα-induced lncMX1–215 markedly suppressed proliferation and metastasis capacity in OSCC. LncMX1-215 negatively regulated PD-L1 expression to inhibit immune escape [48]. The results can direct us toward a potential chemotherapy regime and suggests that targeting lncRNAs can be used to design more effective immune therapies by targeting the PD-1/PD-L1 axis. However, comprehensive investigation should further support the strategy.
6. Perspectives and Conclusions
Despite significant efforts, clinical trials testing novel treatment strategies, personalized medicine and non-invasive, specific biomarkers are the final goals to improving the survival rate of OSCC patients. It is worth noting that outstanding questions and challenges remain.
First, even though the pro-cancer and anti-cancer roles of lncRNAs are revealed in OSCC, we need to clarify different expression levels of lncRNAs in OSCC patients with diverse clinic stages, thereby providing better clues for early diagnosis and discriminating tumor progression. Second, lncRNA can easily be detected in saliva, particularly in OSCC metastatic patients. Therefore, lncRNAs derived from saliva that act as the symbolic body fluid in the oral cavity should be considered for biomarker viability. Third, lncRNAs act as regulators that are associated with TME of OSCC in diverse aspects such as hypoxic conditions, metabolic reprogramming, CAFs, immune cells, and exosomes. However, the explorations of their underlying mechanisms have only begun to scratch the surface and comprehensive studies are still deficient. Fourth, due to the poor conservation of lncRNAs in different species [136], whether or not the experimental results obtained from animal models in vivo can be extended and applied to humans needs further clinical trials to prove their efficacy. More importantly, due to some of the existing challenges in lncRNA therapeutics such as the hurdles of specificity, delivery, tolerability, and the unpredictable off-target effects of lncRNA-based techniques [143], there is still a substantial amount of information to explore. Potential solutions have been mentioned to improve these problems such as chemical modification for optimizing hepatotoxicity and off-target effects of ASOs or instead of stable viral transduction [142,143]. Some substantial progress in oligonucleotide technique is also being explored such as locked nucleic acids (LNAs) with better character of stability and low toxicity [13]. Furthermore, the novel gene-editing technology CRISPR/Cas9 system also showed tremendous potential in clinical application due to its operability and economical features [145].
Notably, lncRNAs are involved in regulating the PD-1/PD-L1 axis, which plays an important role in immunotherapy in cancer patients. However, no lncRNA-targeting therapeutics to modulate the PD-1/PD-L1 axis have entered clinical trials. Therefore, the detailed mechanisms of lncRNA modulation of the PD-1/PD-L1 axis are still poorly understood in OSCC. Further investigations will help us understand that targeting lncRNAs in combination with anti-PD-1/PD-L1 may prove to be a broadly applicable new strategy in tumor immunotherapy.
Taken together, as described in this review, we summarize the functions and potential molecular mechanisms of dysregulated lncRNAs in OSCC as well as the crosstalk with the tumor microenvironment. In particular, we emphasized the translational potential of lncRNAs in the diagnosis and treatment in the future, especially the emerging lncRNA-targeted therapeutic techniques including CRISPR-Cas9 as well as immune checkpoint therapies to target lncRNA and the PD-1/PD-L1 axis. Finally, we truly believe that we have only made the first step toward an in-depth understanding of the functions of lncRNAs in OSCC and this review presents the future perspectives of lncRNAs in OSCC therapy.
Author Contributions
Conceptualization, J.T., Z.T. and H.Z.; Investigation, J.T.; Visualization, J.T., J.C.; Supervision, X.F.; Validation, X.F.; Funding acquisition, Z.T. and H.Z.; Project administration, Z.T. and H.Z.; Writing—Original Draft Preparation, J.T.; Writing—Review & Editing, X.F. and J.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Hunan Key Laboratory of Oral Health Research (2019TPI039 to Z.T.), and Hunan Provincial Natural Science Foundation of China (2021JJ30975 to H.Z.).
Conflicts of Interest
All the authors declare no conflicts of interest.
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Primers 2020, 6, 92. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Brannan, C.; Dees, E.; Ingram, R.; Tilghman, S. The product of the H19 gene may function as an RNA. Mol. Cell Biol. 1990, 10, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Gabory, A.; Ripoche, M.A.; Le Digarcher, A.; Watrin, F.; Ziyyat, A.; Forne, T.; Jammes, H.; Ainscough, J.F.; Surani, M.A.; Journot, L.; et al. H19 acts as a trans regulator of the imprinted gene network controlling growth in mice. Development 2009, 136, 3413–3421. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Liu, Y.; Guo, C.; Zhang, S.; Gong, Z.; Tang, Y.; Yang, L.; He, Y.; Lian, Y.; Li, X.; et al. Upregulated long non-coding RNA LINC00152 expression is associated with progression and poor prognosis of tongue squamous cell carcinoma. J. Cancer 2017, 8, 523–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Zhang, L.; Zhang, L.; Wang, Y.; Li, H.; Ren, X.; Wei, F.; Yu, W.; Liu, T.; Wang, X.; et al. Long non-coding RNA HOTAIR promotes tumor cell invasion and metastasis by recruiting EZH2 and repressing E-cadherin in oral squamous cell carcinoma. Int. J. Oncol. 2015, 46, 2586–2594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Chao, T.C.; Chang, K.Y.; Lin, N.; Patil, V.S.; Shimizu, C.; Head, S.R.; Burns, J.C.; Rana, T.M. The long noncoding RNA THRIL regulates TNFalpha expression through its interaction with hnRNPL. Proc. Natl. Acad. Sci. USA 2014, 111, 1002–1007. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Song, Y.; Li, D.; Liu, X.; Pan, Y.; Ding, L.; Shi, G.; Wang, Y.; Ni, Y.; Hou, Y. Cancer-associated fibroblasts promote tumor progression by lncRNA-mediated RUNX2/GDF10 signaling in oral squamous cell carcinoma. Mol. Oncol. 2021, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Wu, Z.; Zhang, J.; Su, B. Salivary lncRNA as a potential marker for oral squamous cell carcinoma diagnosis. Mol. Med. Rep. 2013, 7, 761–766. [Google Scholar] [CrossRef] [Green Version]
- Huarte, M. The emerging role of lncRNAs in cancer. Nat. Med. 2015, 21, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Tang, Z.; Zhang, H.; Quan, H. Long non-coding RNA DNM3OS/miR-204-5p/HIP1 axis modulates oral cancer cell viability and migration. J. Oral. Pathol. Med. 2020, 49, 865–875. [Google Scholar] [CrossRef]
- Arunkumar, G.; Deva Magendhra Rao, A.K.; Manikandan, M.; Arun, K.; Vinothkumar, V.; Revathidevi, S.; Rajkumar, K.S.; Rajaraman, R.; Munirajan, A.K. Expression profiling of long non-coding RNA identifies linc-RoR as a prognostic biomarker in oral cancer. Tumour. Biol. 2017, 39, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Jiang, E.; Shao, Z.; Shang, Z. Long noncoding RNAs in the metastasis of oral squamous cell carcinoma. Front. Oncol. 2020, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Gao, L.; Ma, X.; Huang, J.; Chen, J.; Zeng, L.; Ashby, C.; Zou, C.; Chen, Z. Long non-coding RNAs regulate drug resistance in cancer. Mol. Cancer 2020, 19, 54. [Google Scholar] [CrossRef]
- Chandra Gupta, S.; Nandan Tripathi, Y. Potential of long non-coding RNAs in cancer patients: From biomarkers to therapeutic targets. Int. J. Cancer 2017, 140, 1955–1967. [Google Scholar] [CrossRef]
- Li, M.; Ding, X.; Zhang, Y.; Li, X.; Zhou, H.; Yang, L.; Li, Y.; Yang, P.; Zhang, X.; Hu, J.; et al. Antisense oligonucleotides targeting lncRNA AC104041.1 induces antitumor activity through Wnt2B/β-catenin pathway in head and neck squamous cell carcinomas. Cell Death Dis. 2020, 11, 672. [Google Scholar] [CrossRef]
- Li, Y.; Wan, Q.; Wang, W.; Mai, L.; Sha, L.; Mashrah, M.; Lin, Z.; Pan, C. LncRNA ADAMTS9-AS2 promotes tongue squamous cell carcinoma proliferation, migration and EMT via the miR-600/EZH2 axis. Biomed. Pharm. 2019, 112, 1–9. [Google Scholar] [CrossRef]
- Li, M.; Yu, D.; Li, Z.; Zhao, C.; Su, C.; Ning, J. Long noncoding RNA AFAP1AS1 facilitates the growth and invasiveness of oral squamous cell carcinoma by regulating the miR145/HOXA1 axis. Oncol. Rep. 2021, 45, 1094–1104. [Google Scholar] [CrossRef]
- Zhang, D.; Ding, L.; Li, Y.; Ren, J.; Shi, G.; Wang, Y.; Zhao, S.; Ni, Y.; Hou, Y. Midkine derived from cancer-associated fibroblasts promotes cisplatin-resistance via up-regulation of the expression of lncRNA ANRIL in tumour cells. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.; Kong, F.; Zhang, S.; Wang, G.; She, P.; Zhang, Q. Long non-coding RNA BANCR promotes proliferation and migration in oral squamous cell carcinoma via MAPK signaling pathway. J. Oral Pathol. Med. 2021, 50, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Gou, C.; Han, P.; Li, J.; Gao, L.; Ji, X.; Dong, F.; Su, Q.; Zhang, Y.; Liu, X. Knockdown of lncRNA BLACAT1 enhances radiosensitivity of head and neck squamous cell carcinoma cells by regulating PSEN1. Br. J. Radiol. 2020, 93, 20190154. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ma, J.; Chen, J.; Huang, H. LncRNA CACS15 regulates tongue squamous cell carcinoma cell behaviors and predicts survival. BMC Oral Health 2019, 19, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Guo, B.; Zhu, Y.; Xu, W.; Ning, S.; Liu, L. Up-regulation of plasma lncRNA CACS15 distinguished early-stage oral squamous cell carcinoma patient. Oral Dis. 2020, 26, 1619–1624. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Z.; Ma, L.; Gong, Z.; Xue, L.; Wang, Q. Long non-coding RNA CASC15 promotes tongue squamous carcinoma progression through targeting miR-33a-5p. Environ. Sci. Pollut. Res. Int. 2018, 25, 22205–22212. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Wu, W. Downregulation of lncRNA CASC2 promotes the postoperative local recurrence of early oral squamous cell carcinoma. Eur. Arch. Oto-Rhino-Laryngol. 2019, 276, 605–610. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, D.; Liu, H.; Yang, K. Increased expression of lncRNA CASC9 promotes tumor progression by suppressing autophagy-mediated cell apoptosis via the AKT/mTOR pathway in oral squamous cell carcinoma. Cell Death Dis. 2019, 10, 41. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Xu, H.; Sun, G.; Zhang, Y. LncRNA CASC9 affects cell proliferation, migration, and invasion of tongue squamous cell carcinoma via regulating miR-423-5p/SOX12 axes. Cancer Manag. Res. 2020, 12, 277–287. [Google Scholar] [CrossRef] [Green Version]
- Li, G.H.; Ma, Z.H.; Wang, X. Long non-coding RNA CCAT1 is a prognostic biomarker for the progression of oral squamous cell carcinoma via miR-181a-mediated Wnt/beta-catenin signaling pathway. Cell Cycle 2019, 18, 2902–2913. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, G.; Murugan, A.K.; Prasanna Srinivasa Rao, H.; Subbiah, S.; Rajaraman, R.; Munirajan, A.K. Long non-coding RNA CCAT1 is overexpressed in oral squamous cell carcinomas and predicts poor prognosis. Biomed. Rep. 2017, 6, 455–462. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Shen, Z. Knockdown of long non-coding RNA (lncRNA) colon cancer-associated transcript-1 (CCAT1) suppresses oral squamous cell carcinoma proliferation, invasion, and migration by inhibiting the discoidin domain receptor 2 (DDR2)/ERK/AKT axis. Med. Sci. Monit. 2020, 26, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Hu, X.; Shang, C.; Zhong, M.; Guo, Y. Silencing of long non-coding RNA CCAT2 depressed malignancy of oral squamous cell carcinoma via Wnt/beta-catenin pathway. Tumour. Biol. 2017, 39, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Ma, Y.; Hu, X.; Song, R.; Zhu, L.; Zhong, M. Long non-coding RNA CEBPA-AS1 correlates with poor prognosis and promotes tumorigenesis via CEBPA/Bcl2 in oral squamous cell carcinoma. Cancer Biol. Ther. 2018, 19, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Sun, L.; Xie, S.; Zhang, S.; Fan, S.; Li, Q.; Chen, W.; Pan, G.; Wang, W.; Weng, B.; et al. Chemotherapy-induced long non-coding RNA 1 promotes metastasis and chemo-resistance of TSCC via the wnt/beta-catenin signaling pathway. Mol. Ther. 2018, 26, 1494–1508. [Google Scholar] [CrossRef] [Green Version]
- Lv, T.; Liu, H.; Wu, Y.; Huang, W. Knockdown of lncRNA DLEU1 inhibits the tumorigenesis of oral squamous cell carcinoma via regulation of miR1495p/CDK6 axis. Mol. Med. Rep. 2021, 23, 1–11. [Google Scholar] [CrossRef]
- Nishiyama, K.; Maruyama, R.; Niinuma, T.; Kai, M.; Kitajima, H.; Toyota, M.; Hatanaka, Y.; Igarashi, T.; Kobayashi, J.I.; Ogi, K.; et al. Screening for long noncoding RNAs associated with oral squamous cell carcinoma reveals the potentially oncogenic actions of DLEU1. Cell Death Dis. 2018, 9, 826. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.; Li, Z.; Gan, Z.; Yang, Z.; Wu, Z.; Rong, M. LncRNA ELF3-AS1 is involved in the regulation of oral squamous cell carcinoma cell proliferation by reprogramming glucose metabolism. OncoTargets Ther. 2019, 12, 6857–6863. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Xie, T.; Qiu, X.; Sun, X.; Chen, J.; Huang, Z.; Zheng, X.; Wang, Z.; Zhao, J. Long noncoding RNA FALEC inhibits proliferation and metastasis of tongue squamous cell carcinoma by epigenetically silencing ECM1 through EZH2. Aging 2019, 11, 4990–5007. [Google Scholar] [CrossRef]
- Zhang, N.; Zeng, L.; Wang, S.; Wang, R.; Yang, R.; Jin, Z.; Tao, H. LncRNA FER1L4 promotes oral squamous cell carcinoma progression via targeting miR-133a-5p/Prx1 axis. OncoTargets Ther. 2021, 14, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Ren, J.; Zhang, D.; Li, Y.; Huang, X.; Hu, Q.; Wang, H.; Song, Y.; Ni, Y.; Hou, Y. A novel stromal lncRNA signature reprograms fibroblasts to promote the growth of oral squamous cell carcinoma via LncRNA-CAF/interleukin-33. Carcinogenesis 2018, 39, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.P.; Yao, J.; Luo, W.; Feng, F.K.; Ma, J.T.; Ren, Y.P.; Wang, D.L.; Bu, R.F. The expression and functional role of a FOXC1 related mRNA-lncRNA pair in oral squamous cell carcinoma. Mol. Cell Biochem. 2014, 394, 177–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Zhou, W.; Lin, C.; Wang, X.; Zhang, X.; Zhang, Y.; Yang, R.; Chen, W.; Cao, W. Dysregulation of FOXD2-AS1 promotes cell proliferation and migration and predicts poor prognosis in oral squamous cell carcinoma: A study based on TCGA data. Aging 2020, 13, 2379–2396. [Google Scholar] [CrossRef]
- Zhang, C.Z. Long non-coding RNA FTH1P3 facilitates oral squamous cell carcinoma progression by acting as a molecular sponge of miR-224-5p to modulate fizzled 5 expression. Gene 2017, 607, 47–55. [Google Scholar] [CrossRef]
- Zeng, B.; Li, Y.; Jiang, F.; Wei, C.; Chen, G.; Zhang, W.; Zhao, W.; Yu, D. LncRNA GAS5 suppresses proliferation, migration, invasion, and epithelial-mesenchymal transition in oral squamous cell carcinoma by regulating the miR-21/PTEN axis. Exp. Cell Res. 2019, 374, 365–373. [Google Scholar] [CrossRef]
- Hong, Y.; He, H.; Sui, W.; Zhang, J.; Zhang, S.; Yang, D. Long non-coding RNA H19 promotes cell proliferation and invasion by acting as a ceRNA of miR138 and releasing EZH2 in oral squamous cell carcinoma. Int. J. Oncol. 2018, 52, 901–912. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Chang, H.; Yang, W.; Lu, Y.; Hu, J.; Jin, S. A novel IFNalpha-induced long noncoding RNA negatively regulates immunosuppression by interrupting H3K27 acetylation in head and neck squamous cell carcinoma. Mol. Cancer 2020, 19, 4. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Wang, S.; Chen, J.; Wang, Z.; Liang, X.; Wang, X.; Jiang, J.; Lang, J.; Li, L. Long noncoding RNA HAS2-AS1 mediates hypoxia-induced invasiveness of oral squamous cell carcinoma. Mol. Carcinog. 2017, 56, 2210–2222. [Google Scholar] [CrossRef]
- Zhao, J.; Bai, X.; Feng, C.; Shang, X.; Xi, Y. Long Non-Coding RNA HCP5 Facilitates Cell Invasion And Epithelial-Mesenchymal Transition In Oral Squamous Cell Carcinoma By miR-140-5p/SOX4 Axis. Cancer Manag. Res. 2019, 11, 10455–10462. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liu, W.; Wang, P.; Li, S. RNA interference of long noncoding RNA HOTAIR suppresses autophagy and promotes apoptosis and sensitivity to cisplatin in oral squamous cell carcinoma. J. Oral Pathol. Med. 2018, 47, 930–937. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Tang, Y.; Tang, J.; Liu, Z.; Wang, X. Downregulation of lncRNA HOTTIP suppresses the proliferation, migration, and invasion of oral tongue squamous cell carcinoma by regulation of HMGA2-Mediated wnt/beta-catenin pathway. Cancer Biother. Radiopharm. 2020, 35, 720–730. [Google Scholar] [CrossRef]
- Wang, X.; Li, H.; Shi, J. LncRNA HOXA11-AS promotes proliferation and cisplatin resistance of oral squamous cell carcinoma by suppression of miR-214-3p expression. Biomed. Res. Int. 2019, 2019, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Ye, S.; Wang, J.; Gu, Z.; Zhang, Y.; Zhang, C.; Ma, X. HuR stabilizes lnc-Sox5 mRNA to promote tongue carcinogenesis. Biochemistry 2017, 82, 438–445. [Google Scholar] [CrossRef]
- Yao, Y.; Chen, S.; Lu, N.; Yin, Y.; Liu, Z. LncRNA JPX overexpressed in oral squamous cell carcinoma drives malignancy via miR-944/CDH2 axis. Oral Dis. 2021, 27, 924–933. [Google Scholar] [CrossRef]
- Zhang, S.; Ma, H.; Zhang, D.; Xie, S.; Wang, W.; Li, Q.; Lin, Z.; Wang, Y. LncRNA KCNQ1OT1 regulates proliferation and cisplatin resistance in tongue cancer via miR-211-5p mediated Ezrin/Fak/Src signaling. Cell Death Dis. 2018, 9, 742. [Google Scholar] [CrossRef] [PubMed]
- Qiao, C.; Qiao, T.; Jin, H.; Liu, L.; Zheng, M.; Wang, Z. LncRNA KCNQ1OT1 contributes to the cisplatin resistance of tongue cancer through the KCNQ1OT1/miR-124-3p/TRIM14 axis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Ai, Y.; Wei, H.; Wu, S.; Tang, Z.; Li, X.; Zou, C. Exosomal LncRNA LBX1-AS1 derived from RBPJ overexpressed-macrophages inhibits oral squamous cell carcinoma progress via miR-182-5p/FOXO3. Front. Oncol. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Zhang, C.; Bao, C.; Zhang, X.; Lin, X.; Pan, D.; Chen, Y. Knockdown of lncRNA LEF1-AS1 inhibited the progression of oral squamous cell carcinoma (OSCC) via Hippo signaling pathway. Cancer Biol. Ther. 2019, 20, 1213–1222. [Google Scholar] [CrossRef]
- Li, J.; Xu, X.; Zhang, D.; Lv, H.; Lei, X. LncRNA LHFPL3-AS1 promotes oral squamous cell carcinoma growth and cisplatin resistance through targeting miR-362-5p/CHSY1 pathway. OncoTargets Ther. 2021, 14, 2293–2300. [Google Scholar] [CrossRef]
- Yan, D.; Wu, F.; Peng, C.; Wang, M. Silencing of LINC00284 inhibits cell proliferation and migration in oral squamous cell carcinoma by the miR-211-3p/MAFG axis and FUS/KAZN axis. Cancer Biol. Ther. 2021, 22, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Liu, J.; Li, S.; Jia, B.; Huang, Z.; Shen, J.; Luo, H.; Zhao, J. CCL18-induced LINC00319 promotes proliferation and metastasis in oral squamous cell carcinoma via the miR-199a-5p/FZD4 axis. Cell Death Dis. 2020, 11, 777. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Cao, W.; Wu, K.; Qin, X.; Wang, X.; Li, Y.; Yu, B.; Zhang, Z.; Wang, X.; Yan, M.; et al. LncRNA LINC00460 promotes EMT in head and neck squamous cell carcinoma by facilitating peroxiredoxin-1 into the nucleus. J. Exp. Clin. Cancer Res. 2019, 38, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, K.; Wang, X.; Yu, H.; Yu, Z.; Wang, D.; Xu, X. LINC00460 facilitated tongue squamous cell carcinoma progression via the miR-320b/IGF2BP3 axis. Oral Dis. 2021, 1–13. [Google Scholar] [CrossRef]
- Han, P.; Ji, X.; Zhang, M.; Gao, L. Upregulation of lncRNA LINC00473 promotes radioresistance of HNSCC cells through activating Wnt/β-catenin signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7305–7313. [Google Scholar] [CrossRef]
- Ding, J.; Yang, C.; Yang, S. LINC00511 interacts with miR-765 and modulates tongue squamous cell carcinoma progression by targeting LAMC2. J. Oral Pathol. Med. 2018, 47, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Z. Long intergenic non-coding RNA 668 regulates VEGFA signaling through inhibition of miR-297 in oral squamous cell carcinoma. Biochem. Biophys. Res. Commun. 2017, 489, 404–412. [Google Scholar] [CrossRef]
- Yu, J.; Liu, Y.; Gong, Z.; Zhang, S.; Guo, C.; Li, X.; Tang, Y.; Yang, L.; He, Y.; Wei, F.; et al. Overexpression long non-coding RNA LINC00673 is associated with poor prognosis and promotes invasion and metastasis in tongue squamous cell carcinoma. Oncotarget 2017, 8, 16621–16632. [Google Scholar] [CrossRef] [Green Version]
- Ai, Y.; Wu, S.; Zou, C.; Wei, H. LINC00941 promotes oral squamous cell carcinoma progression via activating CAPRIN2 and canonical WNT/beta-catenin signaling pathway. J. Cell. Mol. Med. 2020, 24, 10512–10524. [Google Scholar] [CrossRef]
- Jia, B.; Dao, J.; Han, J.; Huang, Z.; Sun, X.; Zheng, X.; Xiang, S.; Zhou, H.; Liu, S. LINC00958 promotes the proliferation of TSCC via miR-211-5p/CENPK axis and activating the JAK/STAT3 signaling pathway. Cancer Cell Int. 2021, 21, 147. [Google Scholar] [CrossRef]
- Kong, J.; Sun, W.; Zhu, W.; Liu, C.; Zhang, H.; Wang, H. Long noncoding RNA LINC01133 inhibits oral squamous cell carcinoma metastasis through a feedback regulation loop with GDF15. J. Surg. Oncol. 2018, 118, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Yang, H.; Li, Y.; Guo, W.; Zhang, Y.; Shen, H.; Xing, L.; Li, Y.; Wu, W.; Zhang, X. Long non-coding RNA LINC01137 contributes to oral squamous cell carcinoma development and is negatively regulated by miR-22-3p. Cell. Oncol. 2021, 44, 595–609. [Google Scholar] [CrossRef]
- Huang, W.; Cao, J.; Peng, X. LINC01234 facilitates growth and invasiveness of oral squamous cell carcinoma through regulating the miR-637/NUPR1 axis. Biomed. Pharmacother. 2019, 120, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Jian, X.; Xu, P.; Zhu, R.; Wang, Y. Linc01234 promotes cell proliferation and metastasis in oral squamous cell carcinoma via miR-433/PAK4 axis. BMC Cancer 2020, 20, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Xiong, H.G.; Xiao, Y.; Yang, Q.C.; Yang, S.C.; Tang, H.C.; Zhang, W.F.; Sun, Z.J. Long Non-coding RNA LINC02195 as a Regulator of MHC I Molecules and Favorable Prognostic Marker for Head and Neck Squamous Cell Carcinoma. Front. Oncol. 2020, 10, 615. [Google Scholar] [CrossRef]
- Liu, M.; Liu, Q.; Fan, S.; Su, F.; Jiang, C.; Cai, G.; Wang, Y.; Liao, G.; Lei, X.; Chen, W.; et al. LncRNA LTSCCAT promotes tongue squamous cell carcinoma metastasis via targeting the miR-103a-2-5p/SMYD3/TWIST1 axis. Cell Death Dis. 2021, 12, 144. [Google Scholar] [CrossRef]
- Fang, Z.; Zhang, S.; Wang, Y.; Shen, S.; Wang, F.; Hao, Y.; Li, Y.; Zhang, B.; Zhou, Y.; Yang, H. Long non-coding RNA MALAT-1 modulates metastatic potential of tongue squamous cell carcinomas partially through the regulation of small proline rich proteins. BMC Cancer 2016, 16, 706. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Liu, S.; Cai, G.; Kong, L.; Zhang, T.; Ren, Y.; Wu, Y.; Mei, M.; Zhang, L.; Wang, X. Long non coding RNA MALAT1 promotes tumor growth and metastasis by inducing epithelial-mesenchymal transition in oral squamous cell carcinoma. Sci. Rep. 2015, 5, 15972. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.; Hu, W. Long non-coding RNA MALAT1 promotes oral squamous cell carcinoma development via microRNA-125b/STAT3 axis. J. Cell. Physiol. 2018, 233, 3384–3396. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, C.; Xie, N.; Wang, P. Long non-coding RNA MEG3 inhibits the proliferation and metastasis of oral squamous cell carcinoma by regulating the WNT/beta-catenin signaling pathway. Oncol. Lett. 2017, 14, 4053–4058. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Xiang, L.; Xu, G. LncRNA MEG3 suppresses migration and promotes apoptosis by sponging miR-548d-3p to modulate JAK-STAT pathway in oral squamous cell carcinoma. IUBMB Life 2019, 71, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Shih, J.W.; Chiang, W.F.; Wu, A.T.H.; Wu, M.H.; Wang, L.Y.; Yu, Y.L.; Hung, Y.W.; Wang, W.C.; Chu, C.Y.; Hung, C.L.; et al. Long noncoding RNA LncHIFCAR/MIR31HG is a HIF-1alpha co-activator driving oral cancer progression. Nat. Commun. 2017, 8, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.; Hung, W.; Chou, C.; Tu, H.; Chang, S.; Liu, Y.; Liu, C.; Lin, S. MIR31HGLncRNA drives oncogenicity by inhibiting the limb-bud and heart development gene (LBH) during oral carcinoma. Int. J. Mol. Sci. 2021, 22, 8383. [Google Scholar] [CrossRef]
- Huang, W.; Cui, X.; Chen, J.; Feng, Y.; Song, E.; Li, J.; Liu, Y. Long non-coding RNA NKILA inhibits migration and invasion of tongue squamous cell carcinoma cells via suppressing epithelial-mesenchymal transition. Oncotarget 2016, 7, 62520–62532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Ning, J.; Li, Z.; Fei, Q.; Zhao, C.; Ge, Y.; Wang, L. Long noncoding RNA OIP5-AS1 promotes the progression of oral squamous cell carcinoma via regulating miR-338-3p/NRP1 axis. Biomed. Pharmacother. 2019, 118, 109259. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Wang, Z.; Hu, Q.; Wu, J.; Li, Y.; Ren, X.; Wu, T.; Tao, X.; Chen, X.; et al. LncRNA-p23154 promotes the invasion-metastasis potential of oral squamous cell carcinoma by regulating Glut1-mediated glycolysis. Cancer Lett. 2018, 434, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Zhao, T.; Fu, L.; Tian, R.; Li, D.; Da, Y. Knockdown of long non-coding RNA prostate cancer-associated transcript 1 inhibits the proliferation and metastasis of tongue squamous cell carcinoma cells by upregulating p21. Oncol. Lett. 2020, 19, 2839–2845. [Google Scholar] [CrossRef] [PubMed]
- Sur, S.; Nakanishi, H.; Steele, R.; Ray, R.B. Depletion of PCAT-1 in head and neck cancer cells inhibits tumor growth and induces apoptosis by modulating c-Myc-AKT1-p38 MAPK signalling pathways. BMC Cancer 2019, 19, 354. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Qi, S.; Zhang, X.; Wu, J.; Yang, X.; Wang, R. lncRNA PLAC2 activated by H3K27 acetylation promotes cell proliferation and invasion via the activation of Wnt/betacatenin pathway in oral squamous cell carcinoma. Int. J. Oncol. 2019, 54, 1183–1194. [Google Scholar] [CrossRef]
- Wang, F.; Ji, X.; Wang, J.; Ma, X.; Yang, Y.; Zuo, J.; Cui, J. LncRNA PVT1 enhances proliferation and cisplatin resistance via regulating miR-194-5p/HIF1a axis in oral squamous cell carcinoma. OncoTargets Ther. 2020, 13, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Ren, H. Long noncoding RNA PVT1 promotes tumor cell proliferation, invasion, migration and inhibits apoptosis in oral squamous cell carcinoma by regulating miR1505p/GLUT1. Oncol. Rep. 2020, 44, 1524–1538. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Jiang, Y.; Zhou, W.; Zhang, B.; Li, Y.; Xie, F.; Zhang, J.; Wang, X.; Yan, M.; Xu, Q.; et al. Long noncoding RNA RC3H2 facilitates cell proliferation and invasion by targeting MicroRNA-101-3p/EZH2 axis in OSCC. Mol. Ther. Nucleic. Acids 2020, 20, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Zhang, X.; Lai, W.; Wang, J. Long non-coding RNA SLC16A1-AS1: Its multiple tumorigenesis features and regulatory role in cell cycle in oral squamous cell carcinoma. Cell Cycle 2020, 19, 1641–1653. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, S.; Chen, J. c-Myc induced upregulation of long non-coding RNA SNHG16 enhances progression and carcinogenesis in oral squamous cell carcinoma. Cancer Gene Ther. 2019, 26, 400–410. [Google Scholar] [CrossRef]
- Wang, Q.; Han, J.; Xu, P.; Jian, X.; Huang, X.; Liu, D. Silencing of LncRNA SNHG16 downregulates cyclin D1 (CCND1) to abrogate malignant phenotypes in oral squamous cell carcinoma (OSCC) through upregulating miR-17-5p. Cancer Manag. Res. 2021, 13, 1831–1841. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Yin, Y.; Yao, Y.; Zhang, P. SNHG3/miR-2682-5p/HOXB8 promotes cell proliferation and migration in oral squamous cell carcinoma. Oral Dis. 2021, 27, 1161–1170. [Google Scholar] [CrossRef]
- Jin, N.; Jin, N.; Bu, W.; Li, X.; Liu, L.; Wang, Z.; Tong, J.; Li, D. Long non-coding RNA TIRY promotes tumor metastasis by enhancing epithelial-to-mesenchymal transition in oral cancer. Exp. Biol. Med. 2020, 245, 585–596. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, K.X.; Zou, R.; Liang, J.; Bai, Z.B.; Li, Z.Q.; Zhao, J.J. TUC338 overexpression leads to enhanced proliferation and reduced apoptosis in tongue squamous cell carcinoma cells in vitro. J. Oral Maxillofac. Surg. 2017, 75, 423–428. [Google Scholar] [CrossRef]
- Liang, S.; Zhang, S.; Wang, P.; Yang, C.; Shang, C.; Yang, J.; Wang, J. LncRNA, TUG1 regulates the oral squamous cell carcinoma progression possibly via interacting with Wnt/beta-catenin signaling. Gene 2017, 608, 49–57. [Google Scholar] [CrossRef]
- Fang, Z.; Wu, L.; Wang, L.; Yang, Y.; Meng, Y.; Yang, H. Increased expression of the long non-coding RNA UCA1 in tongue squamous cell carcinomas: A possible correlation with cancer metastasis. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2014, 117, 89–95. [Google Scholar] [CrossRef]
- Fang, Z.; Zhao, J.; Xie, W.; Sun, Q.; Wang, H.; Qiao, B. LncRNA UCA1 promotes proliferation and cisplatin resistance of oral squamous cell carcinoma by sunppressing miR-184 expression. Cancer Med. 2017, 6, 2897–2908. [Google Scholar] [CrossRef]
- Yang, Y.T.; Wang, Y.F.; Lai, J.Y.; Shen, S.Y.; Wang, F.; Kong, J.; Zhang, W.; Yang, H.Y. Long non-coding RNA UCA1 contributes to the progression of oral squamous cell carcinoma by regulating the WNT/beta-catenin signaling pathway. Cancer Sci. 2016, 107, 1581–1589. [Google Scholar] [CrossRef]
- Li, J.; Meng, H.; Bai, Y.; Wang, K. Regulation of lncRNA and its role in cancer metastasis. Oncol. Res. 2016, 23, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Choudhari, R.; Sedano, M.; Harrison, A.; Subramani, R.; Lin, K.; Ramos, E.; Lakshmanaswamy, R.; Gadad, S. Long noncoding RNAs in cancer: From discovery to therapeutic targets. Adv. Clin. Chem. 2020, 95, 105–147. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Sun, L.; Liu, Q.; Gong, C.; Yao, Y.; Lv, X.; Lin, L.; Yao, H.; Su, F.; Li, D.; et al. A cytoplasmic NF-kappaB interacting long noncoding RNA blocks IkappaB phosphorylation and suppresses breast cancer metastasis. Cancer Cell 2015, 27, 370–381. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Lou, Q.Y.; Yang, W.Y.; Wang, Y.R.; Chen, R.; Wang, L.; Xu, T.; Zhang, L. The role of non-coding RNAs in drug resistance of oral squamous cell carcinoma and therapeutic potential. Cancer Commun. 2021, 41, 981–1006. [Google Scholar] [CrossRef] [PubMed]
- Rafiee, A.; Riazi-Rad, F.; Havaskary, M.; Nuri, F. Long noncoding RNAs: Regulation, function and cancer. Biotechnol. Genet. Eng. Rev. 2018, 34, 153–180. [Google Scholar] [CrossRef]
- Bach, D.H.; Lee, S.K. Long noncoding RNAs in cancer cells. Cancer Lett. 2018, 419, 152–166. [Google Scholar] [CrossRef]
- Saxena, A.; Carninci, P. Long non-coding RNA modifies chromatin: Epigenetic silencing by long non-coding RNAs. Bioessays 2011, 33, 830–839. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Li, S.; Fang, J.; Zhu, Y.; Yang, J. Functional long non-coding RNAs in hepatocellular carcinoma. Cancer Lett. 2021, 500, 281–291. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, H.; Chen, S. Mechanisms of long non-coding RNAs in cancers and their dynamic regulations. Cancers 2020, 12, 1245. [Google Scholar] [CrossRef] [PubMed]
- Pisignano, G.; Pavlaki, I.; Murrell, A. Being in a loop: How long non-coding RNAs organise genome architecture. Essays Biochem. 2019, 63, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Zhang, D.; Wu, N.; Xiao, J.; Wang, X.; Ma, W. ceRNA in cancer: Possible functions and clinical implications. J. Med. Genet. 2015, 52, 710–718. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P. A ceRNA hypothesis: The rosetta stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Karreth, F.A.; Pandolfi, P.P. ceRNA cross-talk in cancer: When ce-bling rivalries go awry. Cancer Discov. 2013, 3, 1113–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef]
- Kuo, T.C.; Kung, H.J.; Shih, J.W. Signaling in and out: Long-noncoding RNAs in tumor hypoxia. J. Biomed. Sci. 2020, 27, 59. [Google Scholar] [CrossRef]
- Yang, J.; Shi, X.; Yang, M.; Luo, J.; Gao, Q.; Wang, X.; Wu, Y.; Tian, Y.; Wu, F.; Zhou, H. Glycolysis reprogramming in cancer-associated fibroblasts promotes the growth of oral cancer through the lncRNA H19/miR-675-5p/PFKFB3 signaling pathway. Int. J. Oral Sci. 2021, 13, 12. [Google Scholar] [CrossRef]
- Ahn, Y.H.; Kim, J.S. Long non-coding RNAs as regulators of interactions between cancer-associated fibroblasts and cancer cells in the tumor microenvironment. Int. J. Mol. Sci. 2020, 21, 7484. [Google Scholar] [CrossRef]
- Del Vecchio, F.; Lee, G.H.; Hawezi, J.; Bhome, R.; Pugh, S.; Sayan, E.; Thomas, G.; Packham, G.; Primrose, J.; Pichler, M.; et al. Long non-coding RNAs within the tumour microenvironment and their role in tumour-stroma cross-talk. Cancer Lett. 2018, 421, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, Q.; Liao, Q. Long noncoding RNA: A dazzling dancer in tumor immune microenvironment. J. Exp. Clin. Cancer Res. 2020, 39, 231. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Pan, S.; Chen, X.; Wang, Z.W.; Zhu, X. The role of lncRNAs and circRNAs in the PD-1/PD-L1 pathway in cancer immunotherapy. Mol. Cancer 2021, 20, 116. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Tang, C.; Li, B.; Fu, Z. Effect of LncTUG1 on NK cell killing sensitivity in oral squamous cell carcinoma cells by targeting miR-212-3p. Shanghai Kou Qiang Yi Xue 2019, 28, 567–571. [Google Scholar] [PubMed]
- Luo, Y.; Yang, J.; Yu, J.; Liu, X.; Yu, C.; Hu, J.; Shi, H.; Ma, X. Long non-coding RNAs: Emerging roles in the immunosuppressive tumor microenvironment. Front. Oncol. 2020, 10, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, K.; Zhao, Z.; Liu, K.; Zhang, J.; Li, G.; Wang, L. Long noncoding RNA lnc-sox5 modulates CRC tumorigenesis by unbalancing tumor microenvironment. Cell Cycle 2017, 16, 1295–1301. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, R.; Adam, M.; Hammond, J.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar] [CrossRef]
- Ringuette Goulet, C.; Bernard, G.; Tremblay, S.; Chabaud, S.; Bolduc, S.; Pouliot, F. exosomes induce fibroblast differentiation into cancer-associated fibroblasts through TGFbeta signaling. Mol. Cancer Res. 2018, 16, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- Naderi-Meshkin, H.; Lai, X.; Amirkhah, R.; Vera, J.; Rasko, J.E.J.; Schmitz, U. Exosomal lncRNAs and cancer: Connecting the missing links. Bioinformatics 2019, 35, 352–360. [Google Scholar] [CrossRef]
- Wang, M.; Zhou, L.; Yu, F.; Zhang, Y.; Li, P.; Wang, K. The functional roles of exosomal long non-coding RNAs in cancer. Cell Mol. Life Sci. 2019, 76, 2059–2076. [Google Scholar] [CrossRef]
- Zhang, W.L.; Liu, Y.; Jiang, J.; Tang, Y.J.; Tang, Y.L.; Liang, X.H. Extracellular vesicle long non-coding RNA-mediated crosstalk in the tumor microenvironment: Tiny molecules, huge roles. Cancer Sci. 2020, 111, 2726–2735. [Google Scholar] [CrossRef]
- Sun, Z.; Yang, S.; Zhou, Q.; Wang, G.; Song, J.; Li, Z.; Zhang, Z.; Xu, J.; Xia, K.; Chang, Y.; et al. Emerging role of exosome-derived long non-coding RNAs in tumor microenvironment. Mol. Cancer 2018, 17, 82. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tang, T.; Yang, X.; Qin, P.; Wang, P.; Zhang, H.; Bai, M.; Wu, R.; Li, F. Tumor-derived exosomal long noncoding RNA LINC01133, regulated by Periostin, contributes to pancreatic ductal adenocarcinoma epithelial-mesenchymal transition through the Wnt/beta-catenin pathway by silencing AXIN2. Oncogene 2021, 40, 3164–3179. [Google Scholar] [CrossRef]
- Yang, H.; Qu, H.; Huang, H.; Mu, Z.; Mao, M.; Xie, Q.; Wang, K.; Hu, B. Exosomes-mediated transfer of long noncoding RNA LINC01133 represses bladder cancer progression via regulating the Wnt signaling pathway. Cell Biol. Int. 2021, 45, 1510–1522. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Qi, Y.; Luo, Y.; Chen, X.; Liang, H. Exosomal long non-coding RNA: Interaction between cancer cells and non-cancer cells. Front. Oncol. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bolha, L.; Ravnik-Glavač, M.; Glavač, D.J. Long noncoding RNAs as biomarkers in cancer. Dis. Markers 2017, 2017, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bhan, A.; Soleimani, M.; Mandal, S. Long noncoding RNA and cancer: A new paradigm. Cancer Res. 2017, 77, 3965–3981. [Google Scholar] [CrossRef] [Green Version]
- Hessels, D.; Klein Gunnewiek, J.; van Oort, I.; Karthaus, H.; van Leenders, G.; van Balken, B.; Kiemeney, L.; Witjes, J.; Schalken, J. DD3(PCA3)-based molecular urine analysis for the diagnosis of prostate cancer. Eur. Urol. 2003, 44, 8–16. [Google Scholar] [CrossRef]
- Bussemakers, M.; van Bokhoven, A.; Verhaegh, G.; Smit, F.; Karthaus, H.; Schalken, J.; Debruyne, F.; Ru, N.; Isaacs, W. DD3: A new prostate-specific gene, highly overexpressed in prostate cancer. Cancer Res. 1999, 59, 5975–5979. [Google Scholar]
- de Kok, J.; Verhaegh, G.; Roelofs, R.; Hessels, D.; Kiemeney, L.; Aalders, T.; Swinkels, D.; Schalken, J. DD3(PCA3), a very sensitive and specific marker to detect prostate tumors. Cancer Res. 2002, 62, 2695–2698. [Google Scholar]
- Jia, H.; Wang, X.; Sun, Z. Screening and validation of plasma long non-coding RNAs as biomarkers for the early diagnosis and staging of oral squamous cell carcinoma. Oncol. Lett. 2021, 21, 172. [Google Scholar] [CrossRef]
- Shao, T.; Huang, J.; Zheng, Z.; Wu, Q.; Liu, T.; Lv, X. SCCA, TSGF, and the long non-coding RNA AC007271.3 are effective biomarkers for diagnosing oral squamous cell carcinoma. Cell. Physiol. Biochem. 2018, 47, 26–38. [Google Scholar] [CrossRef]
- Liu, S.J.; Dang, H.X.; Lim, D.A.; Feng, F.Y.; Maher, C.A. Long noncoding RNAs in cancer metastasis. Nat. Rev. Cancer 2021, 21, 446–460. [Google Scholar] [CrossRef] [PubMed]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA therapeutics—Challenges and potential solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.; Kurzrock, R.; Kim, Y.; Woessner, R.; Younes, A.; Nemunaitis, J.; Fowler, N.; Zhou, T.; Schmidt, J.; Jo, M.; et al. AZD9150, a next-generation antisense oligonucleotide inhibitor of STAT3 with early evidence of clinical activity in lymphoma and lung cancer. Sci. Transl. Med. 2015, 7, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Meng, X.; Pan, J.; Jiang, N.; Zhou, C.; Wu, Z.; Gong, Z. CRISPR/Cas9-mediated noncoding RNA editing in human cancers. RNA Biol. 2018, 15, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, B.; Wang, D.; Yang, S.; Liu, Y.; Wu, H.; Li, Z.; Chang, L.; Yang, Z.; Liu, W. Cucurbitacin B inhibits cell proliferation by regulating x-inactive specific transcript expression in tongue cancer. Front. Oncol. 2021, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Saâda-Bouzid, E.; Defaucheux, C.; Karabajakian, A.; Coloma, V.; Servois, V.; Paoletti, X.; Even, C.; Fayette, J.; Guigay, J.; Loirat, D.; et al. Hyperprogression during anti-PD-1/PD-L1 therapy in patients with recurrent and/or metastatic head and neck squamous cell carcinoma. Ann. Oncol. 2017, 28, 1605–1611. [Google Scholar] [CrossRef] [PubMed]
- Nishino, M.; Ramaiya, N.; Hatabu, H.; Hodi, F. Monitoring immune-checkpoint blockade: Response evaluation and biomarker development. Nat. Rev. Clin. Oncol. 2017, 14, 655–668. [Google Scholar] [CrossRef]
- Zhang, C.; Jiang, F.; Su, C.; Xie, P.; Xu, L. Upregulation of long noncoding RNA SNHG20 promotes cell growth and metastasis in esophageal squamous cell carcinoma via modulating ATM-JAK-PD-L1 pathway. J. Cell. Biochem. 2019, 120, 11642–11650. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, N.; Song, P.; Fu, Y.; Ren, Y.; Li, Z.; Wang, J. LncRNA GATA3-AS1 facilitates tumour progression and immune escape in triple-negative breast cancer through destabilization of GATA3 but stabilization of PD-L1. Cell Proliferat 2020, 53, e12855. [Google Scholar] [CrossRef]
- Zhen, S.; Lu, J.; Chen, W.; Zhao, L.; Li, X. Synergistic antitumor effect on bladder cancer by rational combination of programmed cell death 1 blockade and CRISPR-Cas9-Mediated long non-coding RNA urothelial carcinoma associated 1 knockout. Hum. Gene Ther. 2018, 29, 1352–1363. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Long non-coding RNA in OSCC beginning from identification to characterization of their functions and relevant mechanisms. The first step is to acquire samples of primary tumor and normal tissues from OSCC patients, and then RNA sequencing is applied to screen dysregulated lncRNAs. Furthermore, a series oncogenic and tumor-suppressor functions in tumor biological behaviors such as cell cycle, metastasis, angiogenesis, apoptosis, and drug resistance were identified in vitro and in vivo. Mechanistically, lncRNAs regulate OSCC biological behaviors via the following aspects: (a) Epigenetic regulation. For example, FALEC can recruit polycomb complex EZH2 to specific genomic loci, where they methylate H3K27me3 to induce chromatin compaction and affect transcriptional activity. (b) Transcription regulation. For example, NKILA interacts with transcription factors NF-κB to block the action of transcription factors, thus repressing Twist expression. (c) Signaling pathway regulation. LEF1-AS1 interacts with LATS1 protein to inhibit the Hippo signaling pathway, resulting in attenuation of YAP1 phosphorylation (d) ceRNA mechanism/miRNA sponge. For example, H19, acting as a molecular sponge, competitively binds to miR-138, thereby upregulating the level of miR-138 target gene. (e) mRNA stability. For example, CEBPA-AS1 binding to CEBPA mRNA enhances its stability, leading to a gain of target transcripts expression.
Figure 1.
Long non-coding RNA in OSCC beginning from identification to characterization of their functions and relevant mechanisms. The first step is to acquire samples of primary tumor and normal tissues from OSCC patients, and then RNA sequencing is applied to screen dysregulated lncRNAs. Furthermore, a series oncogenic and tumor-suppressor functions in tumor biological behaviors such as cell cycle, metastasis, angiogenesis, apoptosis, and drug resistance were identified in vitro and in vivo. Mechanistically, lncRNAs regulate OSCC biological behaviors via the following aspects: (a) Epigenetic regulation. For example, FALEC can recruit polycomb complex EZH2 to specific genomic loci, where they methylate H3K27me3 to induce chromatin compaction and affect transcriptional activity. (b) Transcription regulation. For example, NKILA interacts with transcription factors NF-κB to block the action of transcription factors, thus repressing Twist expression. (c) Signaling pathway regulation. LEF1-AS1 interacts with LATS1 protein to inhibit the Hippo signaling pathway, resulting in attenuation of YAP1 phosphorylation (d) ceRNA mechanism/miRNA sponge. For example, H19, acting as a molecular sponge, competitively binds to miR-138, thereby upregulating the level of miR-138 target gene. (e) mRNA stability. For example, CEBPA-AS1 binding to CEBPA mRNA enhances its stability, leading to a gain of target transcripts expression.

Figure 2.
Roles of lncRNA in OSCC tumor microenvironment. (A) Hypoxic TME. HAS2-AS1 activates NF-κB in response to HIF-1α induced by microenvironment hypoxia, leading to HAS2 accumulation in CD44, RHAMM depend-way, which promotes EMT and invasion. (B) Metabolic reprogramming. lncRNA-p23154 promotes GLUT1 expression to enhance glycolysis via binding with the promoter region of miR-378a-3p, leading to increased metastatic potential. (C) CAF transformation. loc100506114 promotes functional transformation of NFs to the phenotype of CAFs by forming a feedback loop with EZH2 to activate CAFs secrete GDF10. (D) Immune regulation. Linc02195 regulates MHC I protein to affect immunosurveillance by being closely associated with high expression of HLA I gene, and it showed a positive correlation between increasing number of infiltrating CD8+T and CD4+T cells. (E) Extracellular vesicles. Exosomal-LBX1-AS1 from RBPJ overexpressed macrophages inhibited the proliferation and invasion of OSCC cells by the miR-182-5p/FOXO3 pathway.
Figure 2.
Roles of lncRNA in OSCC tumor microenvironment. (A) Hypoxic TME. HAS2-AS1 activates NF-κB in response to HIF-1α induced by microenvironment hypoxia, leading to HAS2 accumulation in CD44, RHAMM depend-way, which promotes EMT and invasion. (B) Metabolic reprogramming. lncRNA-p23154 promotes GLUT1 expression to enhance glycolysis via binding with the promoter region of miR-378a-3p, leading to increased metastatic potential. (C) CAF transformation. loc100506114 promotes functional transformation of NFs to the phenotype of CAFs by forming a feedback loop with EZH2 to activate CAFs secrete GDF10. (D) Immune regulation. Linc02195 regulates MHC I protein to affect immunosurveillance by being closely associated with high expression of HLA I gene, and it showed a positive correlation between increasing number of infiltrating CD8+T and CD4+T cells. (E) Extracellular vesicles. Exosomal-LBX1-AS1 from RBPJ overexpressed macrophages inhibited the proliferation and invasion of OSCC cells by the miR-182-5p/FOXO3 pathway.

Figure 3.
Potential clinical application of lncRNAs in OSCC. (A). lncRNAs as diagnostic and prognostic biomarkers; (B) lncRNAs as therapeutic targets based on four main lncRNA-based techniques. (a) MALAT1 silenced by siRNA in cytoplasm of Tscca and Tca8113 cells cause the inhibition of invasion and migration. (b) ASO. ASO targets FOXD2-AS1 in the nucleus of CAL27cells, leading to inhibition of tumor growth. (c) shRNA. shRNA targeting lnc-p23154 and transfected into HSC-3 cells led to the inhibition of tumor metastasis. (d) CRISPR/Cas9 system. Guided by the sgRNA, Cas9 can specifically knock out the sequence of the XIST gene in SCC9 cells, leading to the tumor suppressor function.
Figure 3.
Potential clinical application of lncRNAs in OSCC. (A). lncRNAs as diagnostic and prognostic biomarkers; (B) lncRNAs as therapeutic targets based on four main lncRNA-based techniques. (a) MALAT1 silenced by siRNA in cytoplasm of Tscca and Tca8113 cells cause the inhibition of invasion and migration. (b) ASO. ASO targets FOXD2-AS1 in the nucleus of CAL27cells, leading to inhibition of tumor growth. (c) shRNA. shRNA targeting lnc-p23154 and transfected into HSC-3 cells led to the inhibition of tumor metastasis. (d) CRISPR/Cas9 system. Guided by the sgRNA, Cas9 can specifically knock out the sequence of the XIST gene in SCC9 cells, leading to the tumor suppressor function.


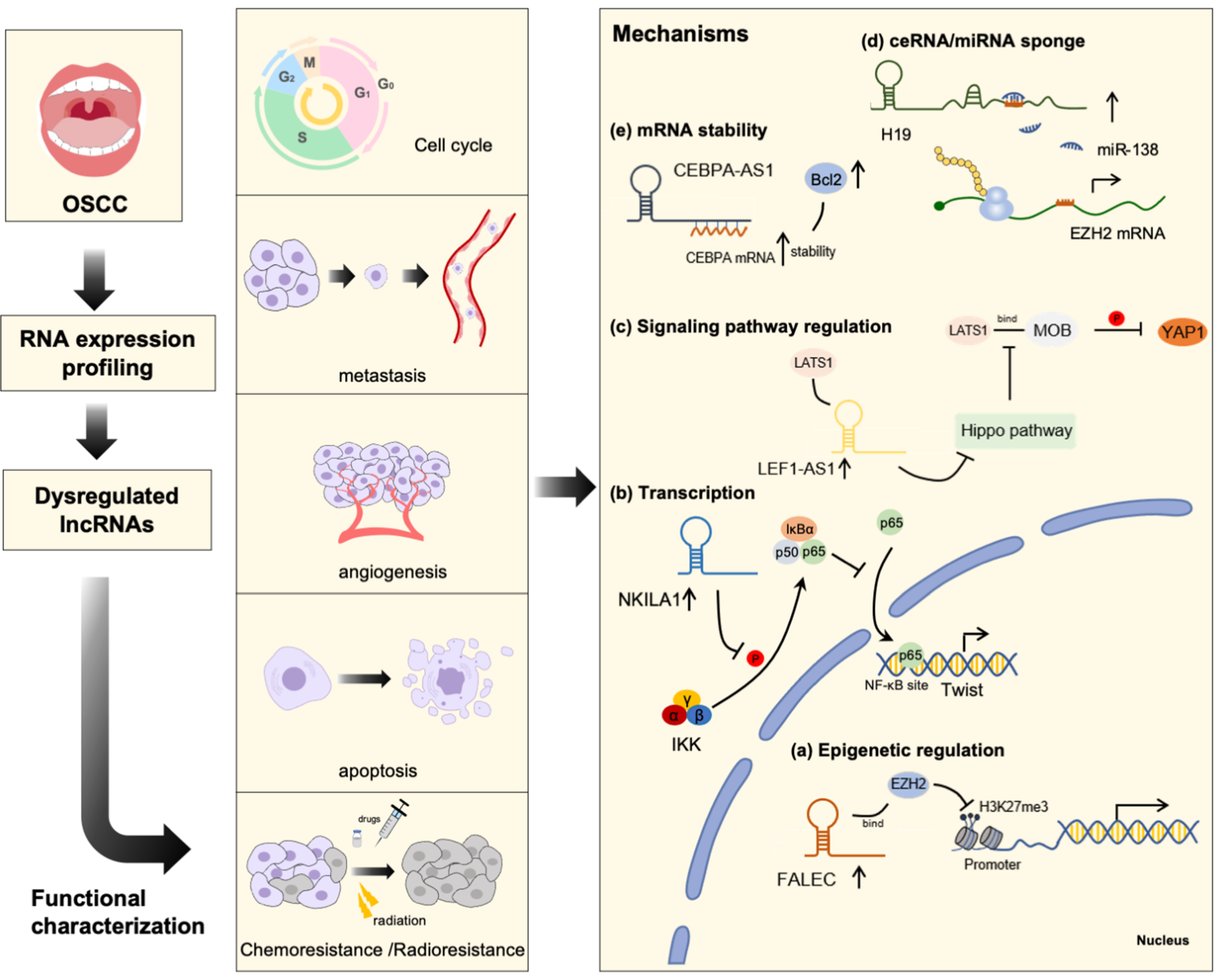{kind=link}
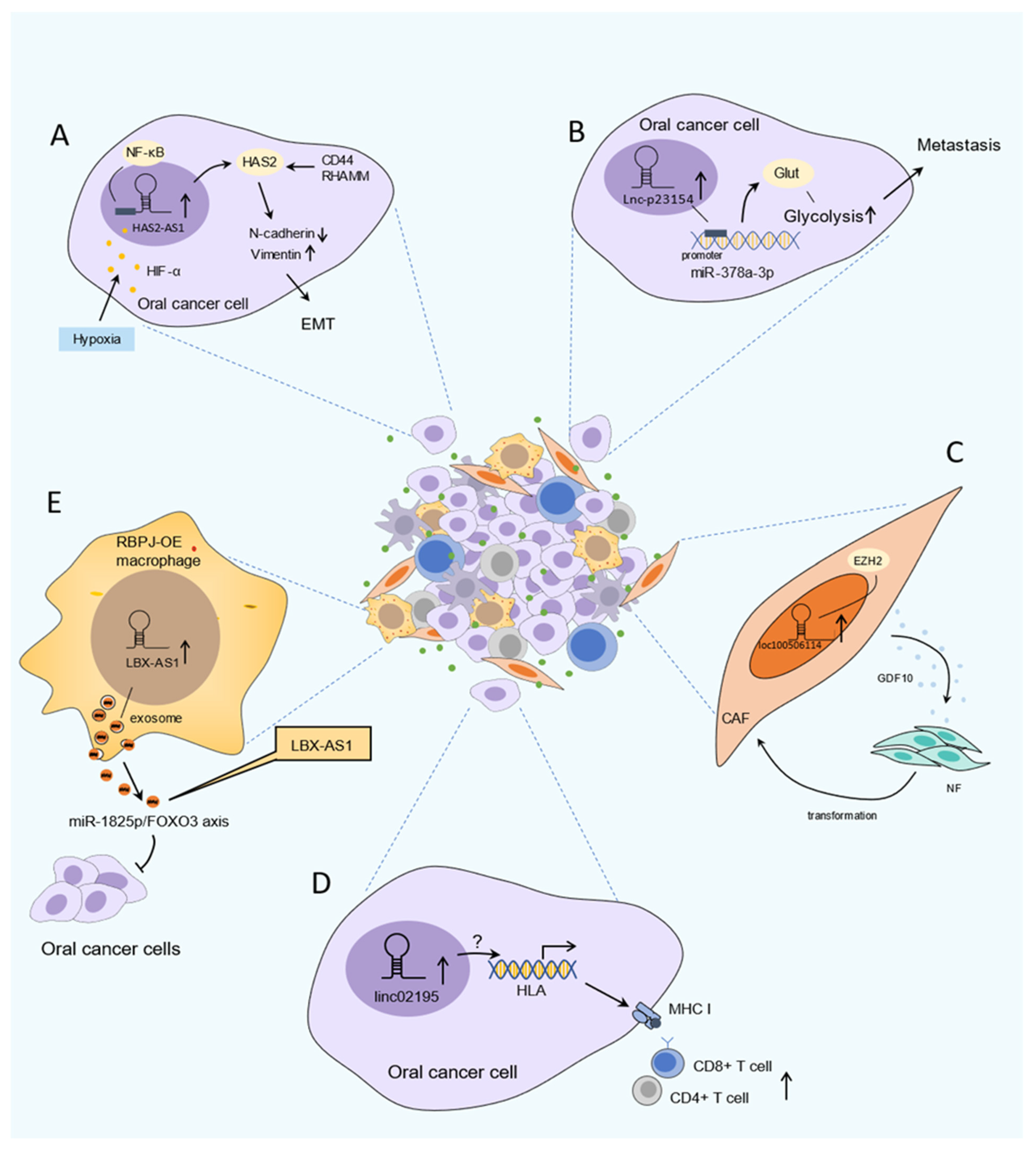{kind=link}
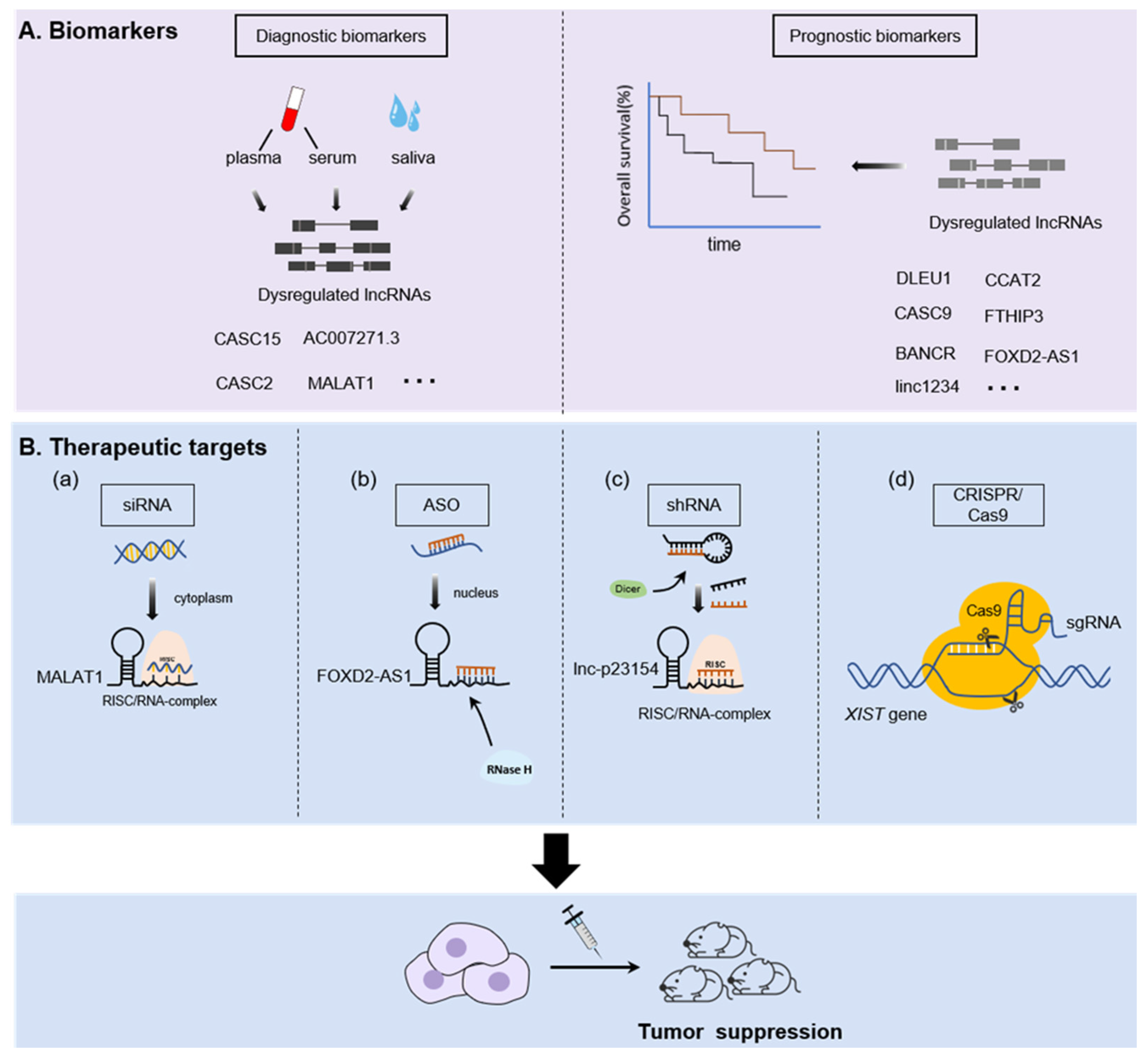{kind=link}
Table 1.
Summary of dysregulated lncRNAs and related targets in OSCC.
| lncRNA | Expression | Functions | Targets | References |
|---|---|---|---|---|
| AC104041.1 | ↑ | Promotes growth and metastasis | Wnt2B/β-catenin signaling | [19] |
| ADAMTS9-AS2 | ↑ | Promotes proliferation, migration, EMT | miR-600/EZH2 | [20] |
| AFAP1-AS1 | ↑ | Promotes proliferation, migration, invasion | miR-145/HOXA1 | [21] |
| ANRIL | ↑ | Promotes proliferation, drug resistance | Midkine/anti-apoptotic protein Bcl-2 | [22] |
| BANCR | ↑ | Promotes proliferation, migration, inhibits apoptosis | MAPK signaling | [23] |
| BLACAT1 | ↑ | Promotes proliferation, radioresistance | PSEN1 | [24] |
| CASC-15 | ↑ | Promotes proliferation, invasion, regulate cell cycle | miR-124, miR-33a-5p | [25,26,27] |
| CASC2 | ↑ | Inhibits proliferation and tumor recurrence | miR-21 | [28] |
| CASC-9 | ↑ | Promotes proliferation, invasion inhibits apoptosis and autophagy | AKT/mTOR pathway, miR-423-5p/SOX12 | [29,30] |
| CCAT-1 | ↑ | Promotes proliferation, invasion, migration, inhibits apoptosis, regulate cell cycle | miR-181a/Wnt/β-catenin DDR2/ERK/AKT, miR155-5p, let7b-5p | [31,32,33] |
| CCAT-2 | ↑ | Promotes proliferation, invasion, inhibits apoptosis | Wnt/β-catenin, GSK-3β | [34] |
| CEBPA-AS1 | ↑ | Promotes tumorigenesis | CEBPA/Bcl2 | [35] |
| CILA1 | ↑ | Promotes EMT, drug resistance | Wnt/β-catenin signaling | [36] |
| DLEU1 | ↑ | Promotes proliferation, invasion, migration and inhabits apoptosis | miR-149-5p/CDK6 HA-CD44 signaling | [37,38] |
| DNM3OS | ↑ | Promotes proliferation, migration, invasion | miR-204-5p/HIP1 | [14] |
| ELF3-AS1 | ↑ | Promotes proliferation | Glucose metabolism | [39] |
| FALEC | ↓ | Inhibit proliferation and migration | ECM1/EZH2 | [40] |
| FER1L4 | ↑ | Enhances growth, migration, invasion | miR-133a-5p/Prx1 | [41] |
| FLJ22447 | ↑ | CAFs activation | IL-33 | [42] |
| FOXCUT | ↑ | Promotes proliferation, migration, and angiogenesis | FOXC1 | [43] |
| FOXD2-AS1 | ↑ | Promotes cell proliferation, migration, immunity inhibition | E2F/G2/M checkpoint | [44] |
| FTH1P3 | ↑ | Enhances growth | mi-224-5p/fizzled 5 | [45] |
| GAS5 | ↓ | Inhibit proliferation, migration, invasion, EMT Promotes radioresistance | miR21/PTEN/PI3K/Akt | [46] |
| H19 | ↑ | Promotes proliferation, invasion | miR-138/EZH2/β-catenin/GSK-3β, H19/miR-675-5p/PFKFB3 | [47,48] |
| HAS2-AS1 | ↑ | Induces EMT, invasion | HF-1α, NF-κB signaling | [49] |
| HCP5 | ↑ | Promotes proliferation, invasion, EMT, regulate cell cycle | miR-140-5p/SOX4 | [50] |
| HOTAIR | ↑ | Promotes proliferation, invasion, EMT, drug resistance, inhibits apoptosis | EZH2/H3K27me3 | [9,51] |
| HOTTIP | ↑ | Promotes proliferation, invasion, migration | miR-124-3p/HMGA2/Wnt/β-Catenin | [52] |
| HOXA11-AS | ↑ | Enhances growth, proliferation, drug resistance, inhibits apoptosis | miR-214-3p, PIM1 | [53] |
| LNC-SOX5 | ↑ | Enhances growth, invasion, migration, inhibits apoptosis | HuR | [54] |
| JPX | ↑ | Promotes proliferation, invasion, migration | miR-944/CDH2 | [55] |
| KCNQ1OT1 | ↑ | Enhances proliferation, drug resistance | miR-211-5p, Ezrin/Fak/Src miR-124-3p/TRIM14 | [56,57] |
| LBX1-AS1 | ↑ | Inhibit proliferation and invasion | miR-182-5p/FOXO3 | [58] |
| LEF1-AS1 | ↑ | Promotes proliferation, invasion, inhibits apoptosis, regulates cell cycle | LATS1/YAP1, Hippo signaling | [59] |
| LHFPL3-AS1 | ↑ | Promotes proliferation, invasion, migration, chemo-resistance | miR-362-5p/CHSY1 | [60] |
| LINC00152 | ↑ | Enhances growth, proliferation, invasion, migration | miR-139-5p | [8] |
| LINC00284 | ↑ | Promotes proliferation, invasion | miR-211-3p/MAFG, FUS/KAZN | [61] |
| LINC00319 | ↑ | Promotes proliferation, angiogenesis | CCL18, miR-199-5P/FZD4 | [62] |
| LINC00460 | ↑ | Promotes proliferation, invasion, migration, EMT | miR-320b/IGF2BP3 | [63,64] |
| LINC00473 | ↑ | Inhibit apoptosis | Wnt/β-catenin signaling | [65] |
| LINC00511 | ↑ | Promotes proliferation, invasion | miR-765/LAMC2 | [66] |
| LINC00668 | ↑ | Enhances growth, proliferation | miR-297/VEGFA | [67] |
| LINC00673 | ↑ | Promotes metastasis | unclear | [68] |
| LINC00941 | ↑ | Promotes proliferation and tumor formation | CAPRIN2/Wnt/β-catenin signaling | [69] |
| LINC00958 | ↑ | Enhances growth, proliferation, regulate cell cycle | miR-211-5p/CENPK, JAK/STAT3 signaling | [70] |
| LINC01133 | ↓ | Inhibited metastasis | GDF15 | [71] |
| LINC01137 | ↑ | Promotes proliferation, invasion, migration | miR-22-3p | [72] |
| LINC01234 | ↑ | Enhances proliferation, invasion, Inhibits apoptosis | miR-637/NUPR, miR-433/PAK4 | [73,74] |
| LINC02195 | ↑ | Immune regulation | MHC I | [75] |
| LINC-ROR | ↑ | Promotes proliferation | miR-145-5p, c-Myc, Klf4, Oct4, Sox2 | [15] |
| LTSCCAT | ↑ | Promotes EMT, migration | miR-103a-2-5p/SMYD3/Twist1 | [76] |
| MALAT1 | ↑ | Promotes proliferation, invasion, EMT | SPRR, miR-125b/STAT3 NF-κB | [77,78,79] |
| MEG3 | ↓ | Inhibits proliferation, invasion, migration, promotes apoptosis | Wnt/β-catenin signaling miR-548d-3p/JAK–STAT | [80,81] |
| MIR31HG | ↑ | Promotes proliferation, invasion, migration and metabolic regulation | HIF1α/p300, MMP1, BMP2, LBH | [82,83] |
| NKILA | ↓ | inhibit EMT, invasion, migration | NF-κB/Twist signaling | [84] |
| OIP5-AS1 | ↑ | Enhances growth, proliferation, migration, invasion | miR-338-3p/NRP1 | [85] |
| p23154 | ↑ | Promote metastasis | miR-378a-3p/ GLUT1 | [86] |
| PCAT-1 | ↑ | Enhances growth, proliferation, invasion, migration, inhibits apoptosis | p21, c-MycAKT1-p38 MAPK signaling | [87,88] |
| PLAC2 | ↑ | Promotes proliferation, invasion | H3K27 acetylation, Wnt/β-catenin signaling | [89] |
| PVT-1 | ↑ | Promotes proliferation, invasion, migration, drug resistance, inhabits apoptosis | miR-150-5p/GLUT-1 miR-194-5p/HIF1a | [90,91] |
| RC3H2 | ↑ | Promotes proliferation and invasion | miR-101-3p/EZH2 | [92] |
| SLC16A1-AS1 | ↑ | Promotes proliferation regulate cell cycle | Cyclin D1 | [93] |
| SNHG16 | ↑ | Promotes proliferation, invasion, inhibits apoptosis | c-Myc, miR-17-5p/CCND1 | [94,95] |
| SNHG3 | ↑ | Enhances proliferation, migration | miR-2682-5p/HOXB8 | [96] |
| TIRY | ↑ | Induces CAFs EMT, promotes invasion and migration | miR-14/Wnt/β-catenin signaling | [97] |
| TUC338 | ↑ | Enhances proliferation, regulate cell cycle, inhabits apoptosis | Unclear | [98] |
| TUG1 | ↑ | Promotes proliferation, invasion, inhibits apoptosis | Wnt/β-catenin signaling | [99] |
| UCA1 | ↑ | Enhances growth, proliferation, invasion, migration drug resistance, inhibits apoptosis | P27, Wnt/β-catenin, miR-184/miR-184/SF1 | [100,101,102] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tang, J.; Fang, X.; Chen, J.; Zhang, H.; Tang, Z. Long Non-Coding RNA (lncRNA) in Oral Squamous Cell Carcinoma: Biological Function and Clinical Application. Cancers 2021, 13, 5944. https://doi.org/10.3390/cancers13235944
AMA Style
Tang J, Fang X, Chen J, Zhang H, Tang Z. Long Non-Coding RNA (lncRNA) in Oral Squamous Cell Carcinoma: Biological Function and Clinical Application. Cancers. 2021; 13(23):5944. https://doi.org/10.3390/cancers13235944
Chicago/Turabian StyleTang, Jianfei, Xiaodan Fang, Juan Chen, Haixia Zhang, and Zhangui Tang. 2021. "Long Non-Coding RNA (lncRNA) in Oral Squamous Cell Carcinoma: Biological Function and Clinical Application" Cancers 13, no. 23: 5944. https://doi.org/10.3390/cancers13235944
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.