
The Cardiac Glycoside Deslanoside Exerts Anticancer Activity in Prostate Cancer Cells by Modulating Multiple Signaling Pathways

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Deslanoside and Cell Lines
2.2. Cell Viability Analysis
2.3. Colony Formation Assay
2.4. EdU (5-ethynyl-2-deoxyuridine) Cell Proliferation Assay
2.5. Cell Cycle Analysis
2.6. Apoptosis Assay
2.7. Transwell Assay
2.8. Western Blotting
2.9. Animal Experiments
2.10. Histology and Immunohistochemistry
2.11. RNA-Seq and Bioinformatic Analyses
2.12. Survival Analysis
2.13. Statistical Analysis
3. Results
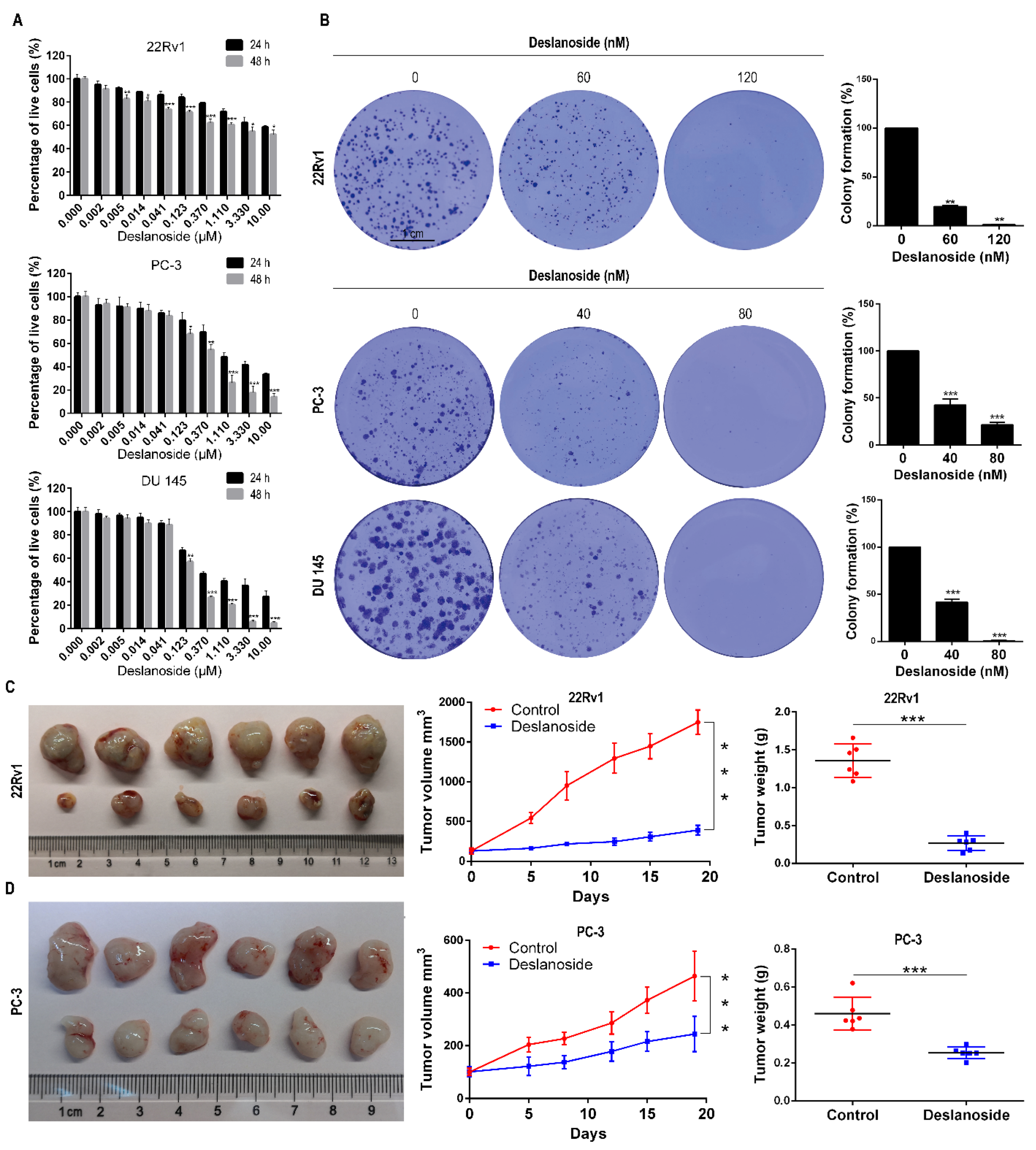
3.1. Deslanoside Exerts an Anticancer Effect in PCa Cells In Vitro and In Vivo
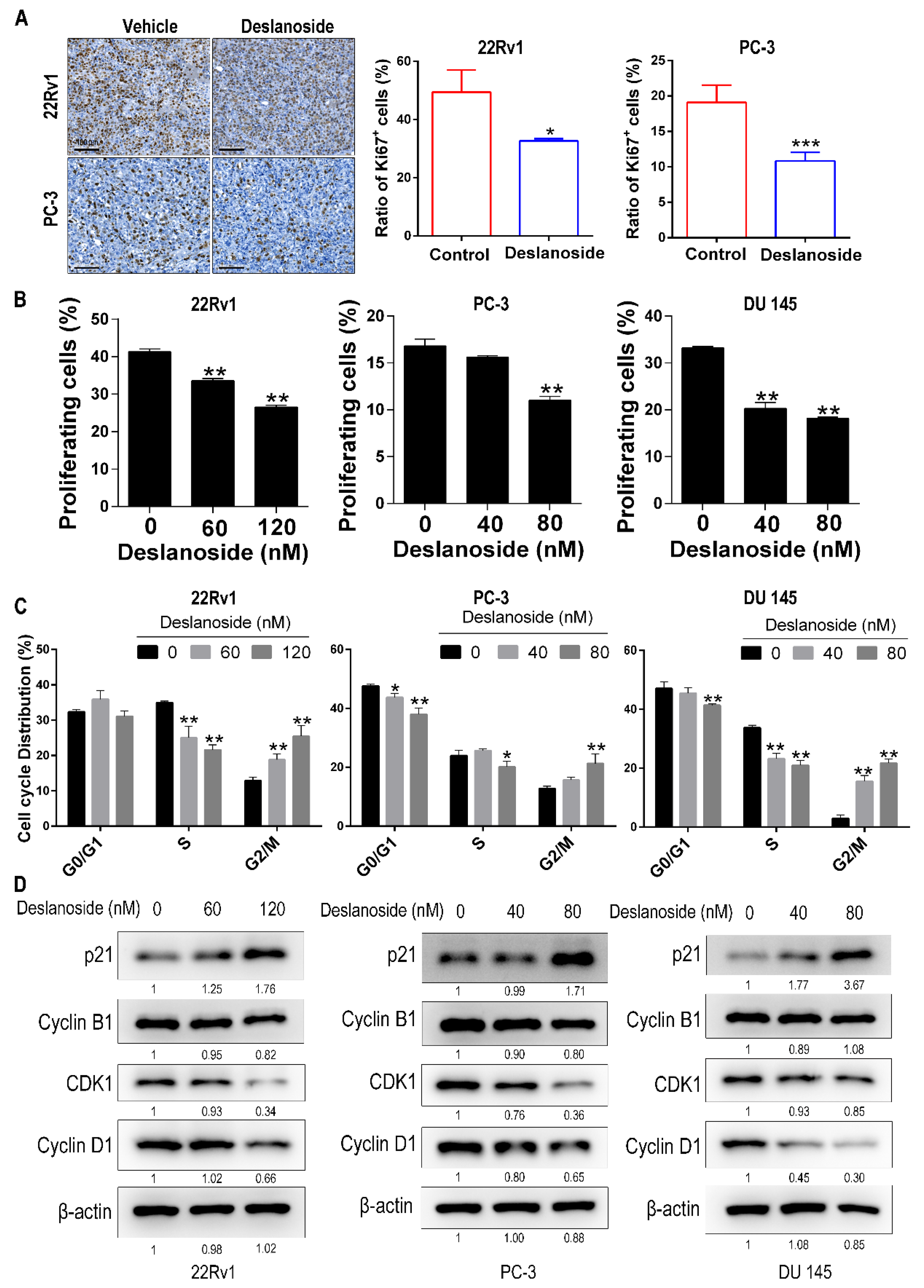
3.2. Deslanoside’s Inhibitory Effect on Tumor Growth Involves Cell Cycle Arrest at G2/M
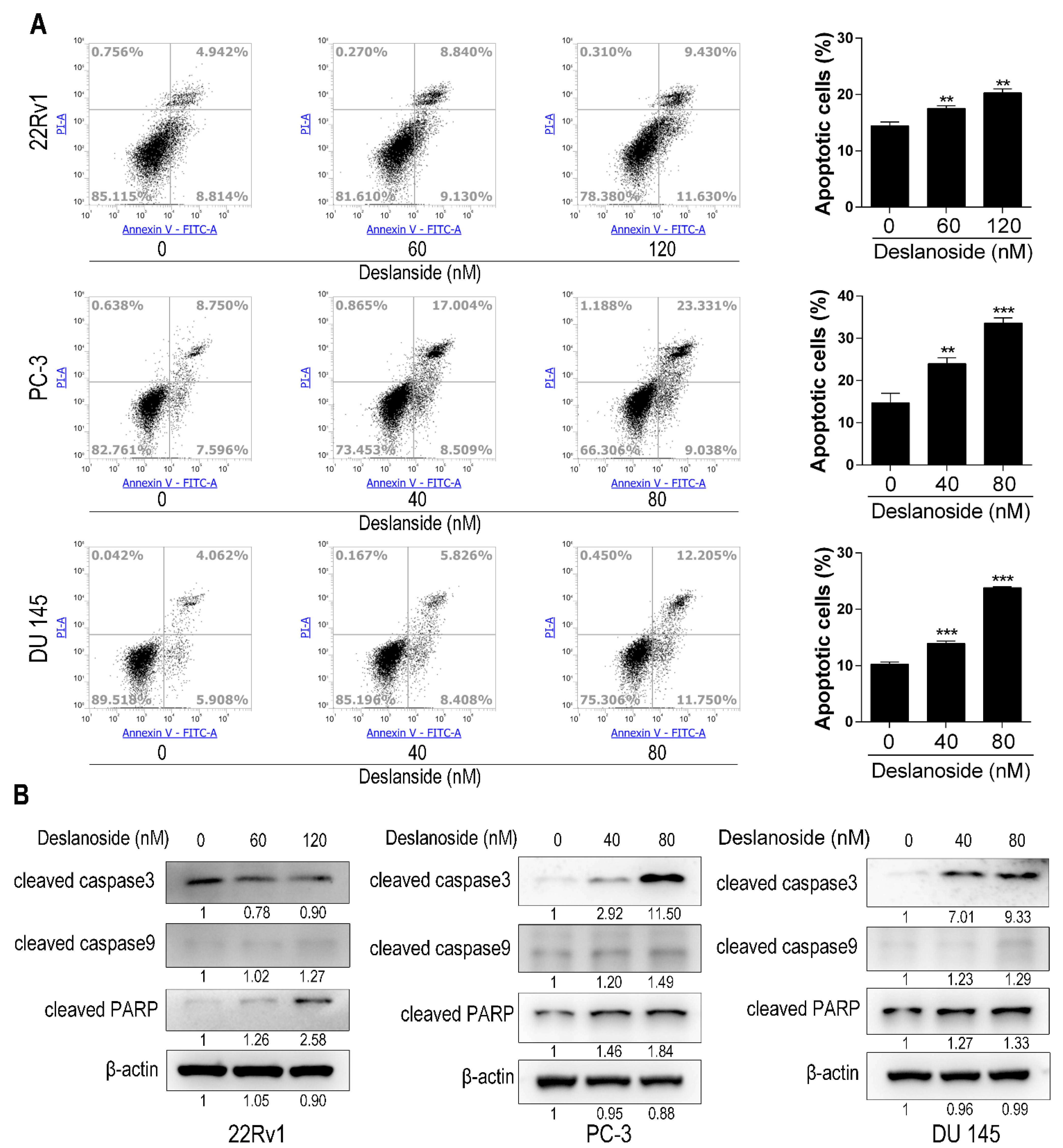
3.3. Deslanoside Also Causes Apoptosis in PCa Cells
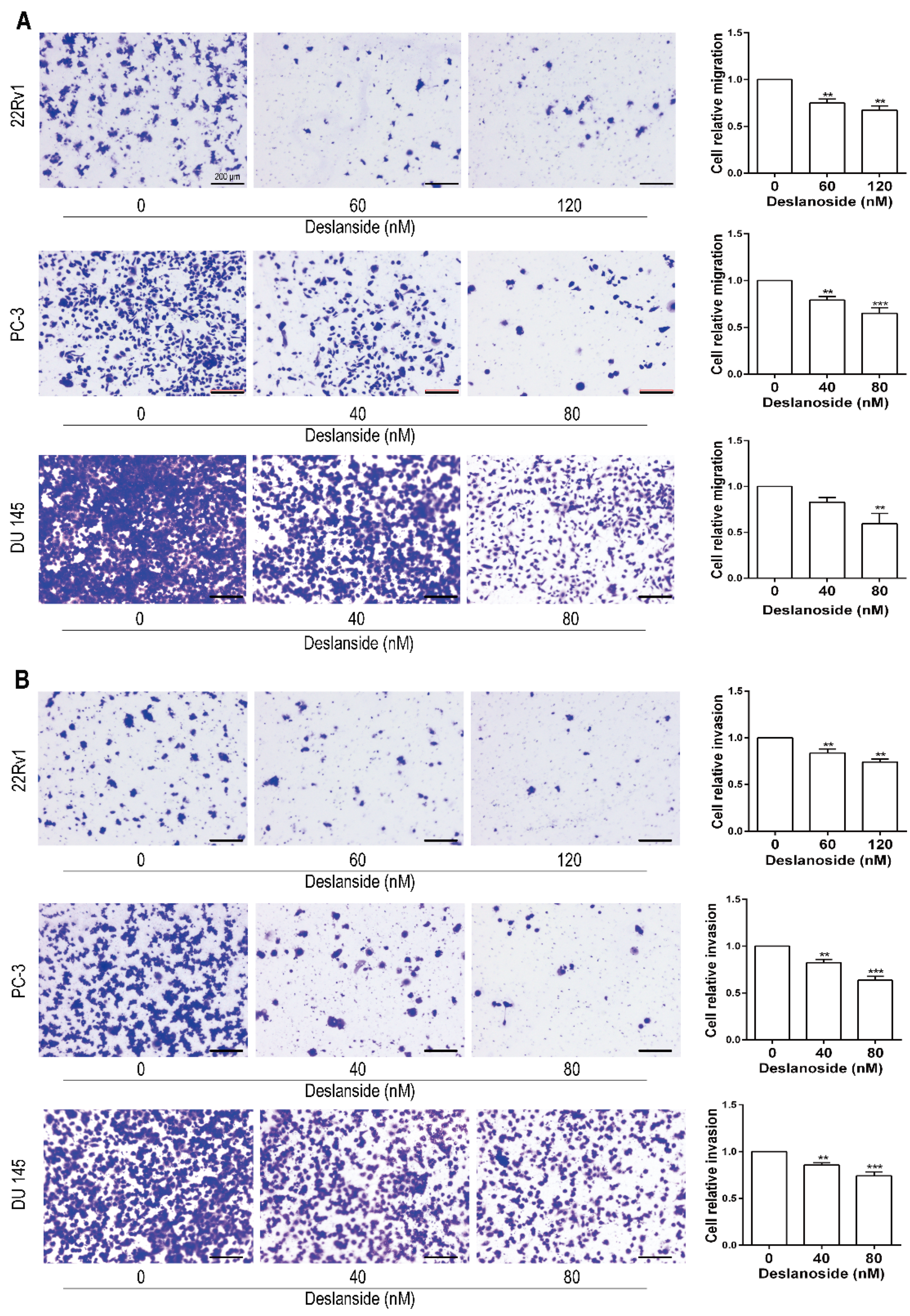
3.4. Deslanoside Suppresses the Migration and Invasion Abilities of Prostate Cancer Cells
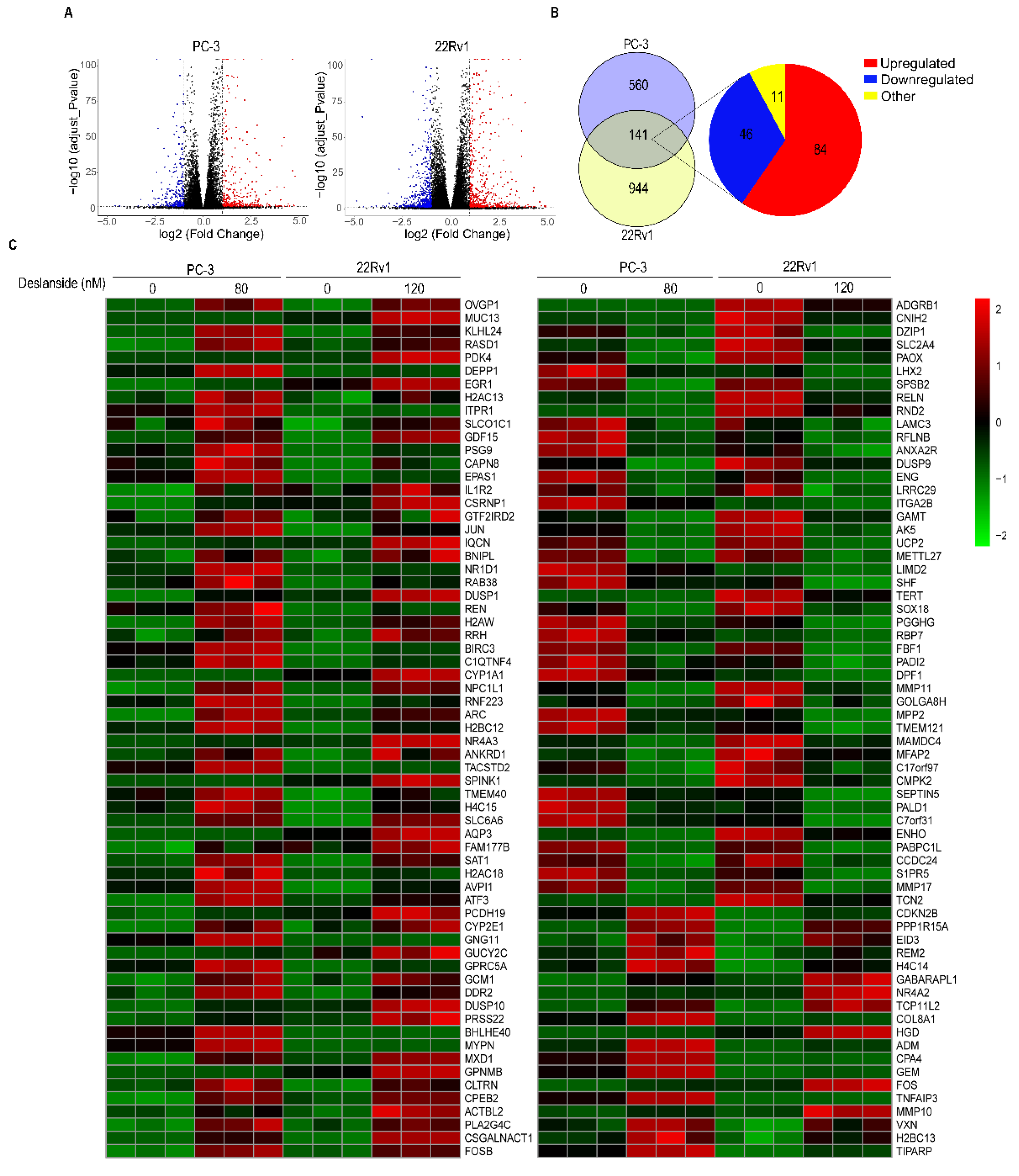
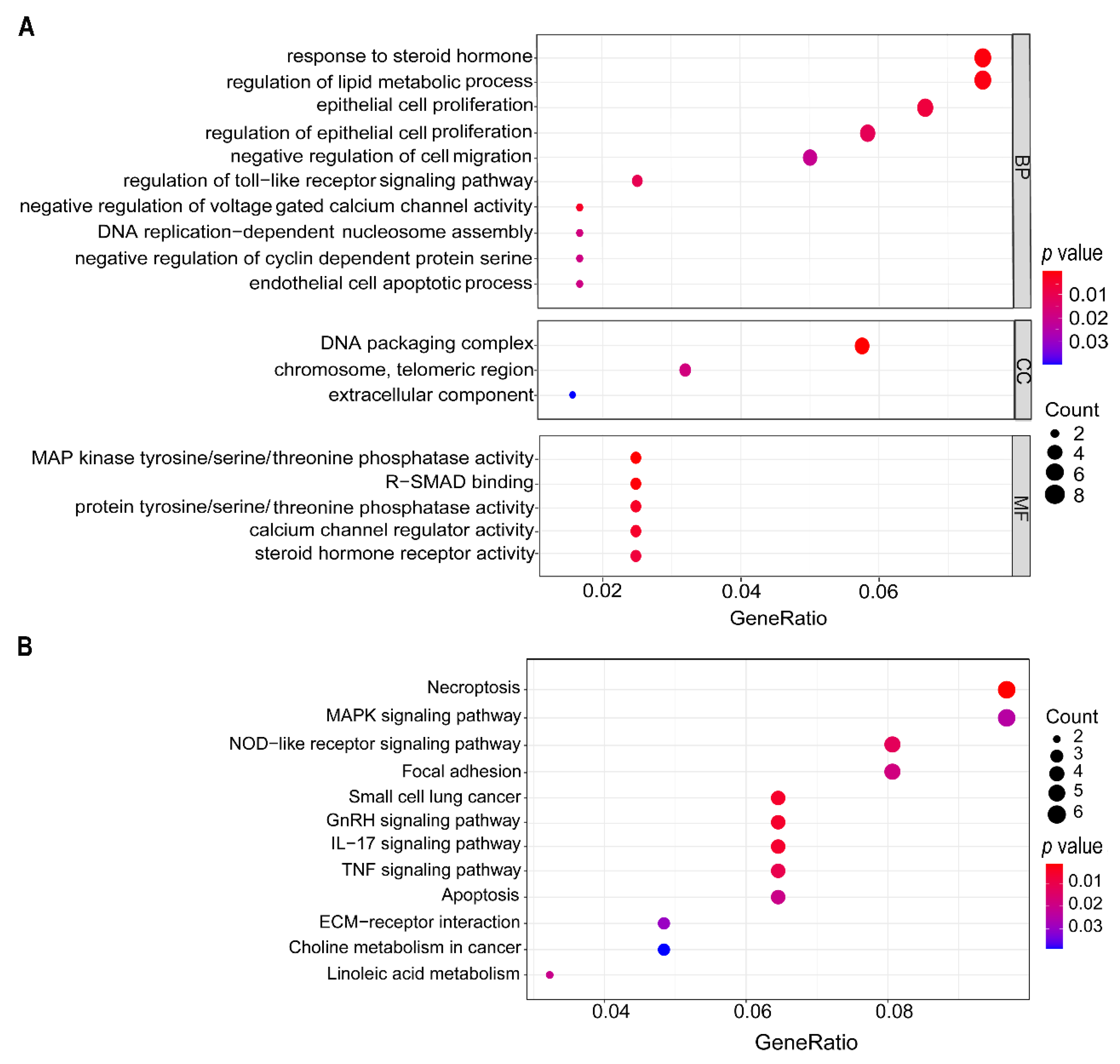
3.5. Deslanoside Alters Multiple Biological Processes and Signaling Pathways
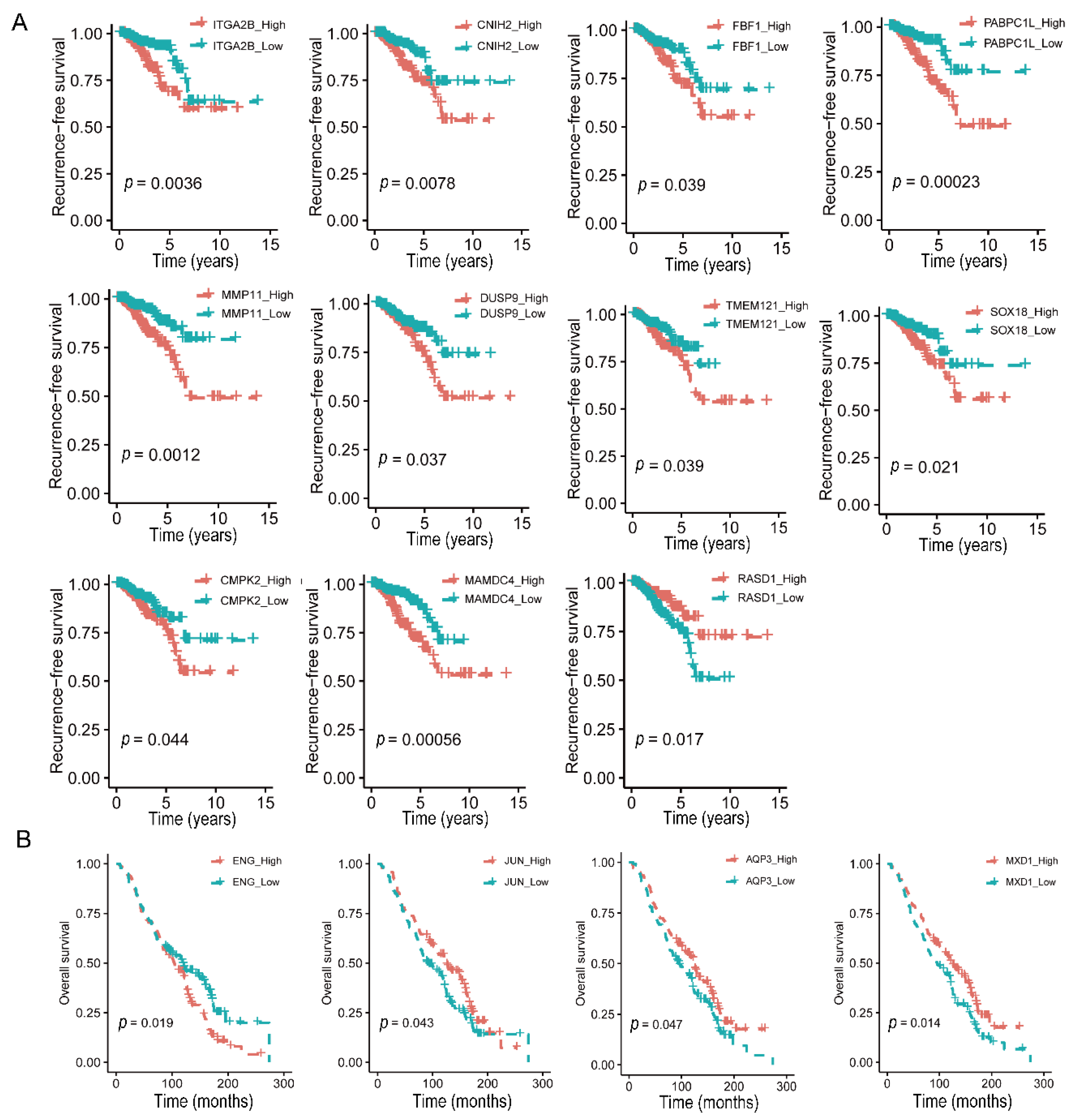
3.6. Expression Levels of 15 Deslanoside-Modulated Genes Are Associated with Patient Survival in Prostate Cancer
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Teo, M.Y.; Rathkopf, D.E.; Kantoff, P. Treatment of Advanced Prostate Cancer. Annu. Rev. Med. 2019, 70, 479–499. [Google Scholar] [CrossRef] [PubMed]
- Nevedomskaya, E.; Baumgart, S.; Haendler, B. Recent Advances in Prostate Cancer Treatment and Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roviello, G.; Sigala, S.; Sandhu, S.; Bonetta, A.; Cappelletti, M.; Zanotti, L.; Bottini, A.; Sternberg, C.; Fox, S.; Generali, D. Role of the novel generation of androgen receptor pathway targeted agents in the management of castration-resistant prostate cancer: A literature based meta-analysis of randomized trials. Eur. J. Cancer 2016, 61, 111–121. [Google Scholar] [CrossRef]
- Lingrel, J. The physiological significance of the cardiotonic steroid/ouabain-binding site of the Na, K-ATPase. Annu. Rev. Physiol. 2010, 72, 395–412. [Google Scholar] [CrossRef] [Green Version]
- Altamirano, J.; Li, Y.; DeSantiago, J.; Piacentino, V.; Houser, S.; Bers, D. The inotropic effect of cardioactive glycosides in ventricular myocytes requires Na+-Ca2+ exchanger function. J. Physiol. 2006, 575, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Ayogu, J.I.; Odoh, A.S. Prospects and Therapeutic Applications of Cardiac Glycosides in Cancer Remediation. ACS Comb. Sci. 2020, 22, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Babula, P.; Masarik, M.; Adam, V.; Provaznik, I.; Kizek, R. From Na+/K+-ATPase and cardiac glycosides to cytotoxicity and cancer treatment. Anti-Cancer Agents Med. Chem. 2013, 13, 1069–1087. [Google Scholar] [CrossRef]
- Botelho, A.F.M.; Pierezan, F.; Soto-Blanco, B.; Melo, M.M. A review of cardiac glycosides: Structure, toxicokinetics, clinical signs, diagnosis and antineoplastic potential. Toxicon 2019, 158, 63–68. [Google Scholar] [CrossRef]
- Guerrero, A.; Herranz, N.; Sun, B.; Wagner, V.; Gallage, S.; Guiho, R.; Wolter, K.; Pombo, J.; Irvine, E.; Innes, A.; et al. Cardiac glycosides are broad-spectrum senolytics. Nat. Metab. 2019, 1, 1074–1088. [Google Scholar] [CrossRef]
- Gkountela, S.; Castro-Giner, F.; Szczerba, B.; Vetter, M.; Landin, J.; Scherrer, R.; Krol, I.; Scheidmann, M.; Beisel, C.; Stirnimann, C.; et al. Circulating Tumor Cell Clustering Shapes DNA Methylation to Enable Metastasis Seeding. Cell 2019, 176, 98–112.e114. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Yu, K.; Wang, G.; Zhang, D.; Shi, C.; Ding, Y.; Hong, D.; Zhang, D.; He, H.; Sun, L.; et al. Lanatoside C inhibits cell proliferation and induces apoptosis through attenuating Wnt/β-catenin/c-Myc signaling pathway in human gastric cancer cell. Biochem. Pharmacol. 2018, 150, 280–292. [Google Scholar] [CrossRef]
- Reddy, D.; Kumavath, R.; Ghosh, P.; Barh, D. Lanatoside C Induces G2/M Cell Cycle Arrest and Suppresses Cancer Cell Growth by Attenuating MAPK, Wnt, JAK-STAT, and PI3K/AKT/mTOR Signaling Pathways. Biomolecules 2019, 9, 792. [Google Scholar] [CrossRef] [Green Version]
- Durmaz, I.; Guven, E.; Ersahin, T.; Ozturk, M.; Calis, I.; Cetin-Atalay, R. Liver cancer cells are sensitive to Lanatoside C induced cell death independent of their PTEN status. Phytomedicine 2016, 23, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Crommentuijn, M.; Maguire, C.; Niers, J.; Vandertop, W.; Badr, C.; Würdinger, T.; Tannous, B. Intracranial AAV-sTRAIL combined with lanatoside C prolongs survival in an orthotopic xenograft mouse model of invasive glioblastoma. Mol. Oncol. 2016, 10, 625–634. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Juang, J.L.; Wang, P.S. Involvement of Cdk5/p25 in digoxin-triggered prostate cancer cell apoptosis. J. Biol. Chem. 2004, 279, 29302–29307. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Khan, M.; Yang, J.; Yao, M.; Yu, S.; Gao, H. Proscillaridin A induces apoptosis, inhibits STAT3 activation and augments doxorubicin toxicity in prostate cancer cells. Int. J. Med. Sci. 2018, 15, 832–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gayed, B.A.; O’Malley, K.J.; Pilch, J.; Wang, Z. Digoxin inhibits blood vessel density and HIF-1a expression in castration-resistant C4-2 xenograft prostate tumors. Clin. Transl. Sci. 2012, 5, 39–42. [Google Scholar] [CrossRef]
- Klys, M.; Gross, A. Determination of deslanoside in antemortem and postmortem specimens. Forensic Sci. Int. 1990, 45, 231–238. [Google Scholar] [CrossRef]
- Li, F.; Lu, T.; Liu, D.; Zhang, C.; Zhang, Y.; Dong, F. Upregulated PPARG2 facilitates interaction with demethylated AKAP12 gene promoter and suppresses proliferation in prostate cancer. Cell Death Dis. 2021, 12, 528. [Google Scholar] [CrossRef]
- Schindelin, J.; Rueden, C.T.; Hiner, M.C.; Eliceiri, K.W. The ImageJ ecosystem: An open platform for biomedical image analysis. Mol. Reprod. Dev. 2015, 82, 518–529. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerella, C.; Dicato, M.; Diederich, M. Assembling the puzzle of anticancer mechanisms triggered by cardiac glycosides. Mitochondrion 2013, 13, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Zhang, Y.; Zhao, W.; Zhou, X.; Wang, C.; Deng, F. The cardiac glycoside oleandrin induces apoptosis in human colon cancer cells via the mitochondrial pathway. Cancer Chemother. Pharmacol. 2017, 80, 91–100. [Google Scholar] [CrossRef]
- Weng, J.R.; Lin, W.Y.; Bai, L.Y.; Hu, J.L.; Feng, C.H. Antitumor activity of the cardiac glycoside αlDiginoside by modulating Mcl-1 in human oral squamous cell carcinoma cells. Int. J. Mol. Sci. 2020, 21, 7947. [Google Scholar] [CrossRef]
- Chang, Y.M.; Shih, Y.L.; Chen, C.P.; Liu, K.L.; Lee, M.H.; Lee, M.Z.; Hou, H.T.; Huang, H.C.; Lu, H.F.; Peng, S.F.; et al. ouabain induces apoptotic cell death in human prostate DU 145 cancer cells through DNA damage and TRAIL pathways. Environ. Toxicol. 2019, 34, 1329–1339. [Google Scholar] [CrossRef]
- Frese, S.; Frese-Schaper, M.; Andres, A.C.; Miescher, D.; Zumkehr, B.; Schmid, R.A. Cardiac glycosides initiate Apo2L/TRAIL-induced apoptosis in non-small cell lung cancer cells by up-regulation of death receptors 4 and 5. Cancer Res. 2006, 66, 5867–5874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossan, M.S.; Chan, Z.Y.; Collins, H.M.; Shipton, F.N.; Butler, M.S.; Rahmatullah, M.; Lee, J.B.; Gershkovich, P.; Kagan, L.; Khoo, T.J.; et al. Cardiac glycoside cerberin exerts anticancer activity through PI3K/AKT/mTOR signal transduction inhibition. Cancer Lett. 2019, 453, 57–73. [Google Scholar] [CrossRef]
- Ark, M.; Ozdemir, A.; Polat, B. Ouabain-induced apoptosis and Rho kinase: A novel caspase-2 cleavage site and fragment of Rock-2. Apoptosis 2010, 15, 1494–1506. [Google Scholar] [CrossRef]
- Rasheduzzaman, M.; Yin, H.; Park, S.Y. Cardiac glycoside sensitized hepatocellular carcinoma cells to TRAIL via ROS generation, p38MAPK, mitochondrial transition, and autophagy mediation. Mol. Carcinog. 2019, 58, 2040–2051. [Google Scholar] [CrossRef]
- Smith, J.A.; Madden, T.; Vijjeswarapu, M.; Newman, R.A. Inhibition of export of fibroblast growth factor-2 (FGF-2) from the prostate cancer cell lines PC3 and DU145 by Anvirzel and its cardiac glycoside component, oleandrin. Biochem. Pharmacol. 2001, 62, 469–472. [Google Scholar] [CrossRef]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of Matrix Metalloproteinases in Photoaging and Photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallas, N.A.; Samuel, S.; Xia, L.; Fan, F.; Gray, M.J.; Lim, S.J.; Ellis, L.M. Endoglin (CD105): A marker of tumor vasculature and potential target for therapy. Clin. Cancer Res. 2008, 14, 1931–1937. [Google Scholar] [CrossRef] [Green Version]
- Kasprzak, A.; Adamek, A. Role of endoglin (CD105) in the progression of hepatocellular carcinoma and anti-angiogenic therapy. Int. J. Mol. Sci. 2018, 19, 3887. [Google Scholar] [CrossRef] [Green Version]
- Nassiri, F.; Cusimano, M.D.; Scheithauer, B.W.; Rotondo, F.; Fazio, A.; Yousef, G.M.; Syro, L.V.; Kovacs, K.; Lloyd, R.V. Endoglin (CD105): A review of its role in angiogenesis and tumor diagnosis, progression and therapy. Anticancer Res. 2011, 31, 2283–2290. [Google Scholar] [PubMed]
- Duff, S.E.; Li, C.; Garland, J.M.; Kumar, S. CD105 is important for angiogenesis: Evidence and potential applications. FASEB J. 2003, 17, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Fonsatti, E.; Nicolay, H.J.; Altomonte, M.; Covre, A.; Maio, M. Targeting cancer vasculature via endoglin/CD105: A novel antibody-based diagnostic and therapeutic strategy in solid tumours. Cardiovasc. Res. 2010, 86, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Jeng, K.S.; Sheen, I.S.; Lin, S.S.; Leu, C.M.; Chang, C.F. The role of endoglin in hepatocellular carcinoma. Int. J. Mol. Sci. 2021, 22, 3208. [Google Scholar] [CrossRef]
- Eferl, R.; Wagner, E.F. AP-1: A double-edged sword in tumorigenesis. Nat. Rev. Cancer 2003, 3, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Zhang, X.; Li, B.; Meng, X. MAGI2-AS3 suppresses MYC signaling to inhibit cell proliferation and migration in ovarian cancer through targeting miR-525-5p/MXD1 axis. Cancer Med. 2020, 9, 6377–6386. [Google Scholar] [CrossRef] [PubMed]
- Salehi-Tabar, R.; Nguyen-Yamamoto, L.; Tavera-Mendoza, L.E.; Quail, T.; Dimitrov, V.; An, B.S.; Glass, L.; Goltzman, D.; White, J.H. Vitamin D receptor as a master regulator of the c-MYC/MXD1 network. Proc. Natl. Acad. Sci. USA 2012, 109, 18827–18832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cascón, A.; Robledo, M. MAX and MYC: A heritable breakup. Cancer Res. 2012, 72, 3119–3124. [Google Scholar] [CrossRef] [Green Version]
- Grimm, D.; Bauer, J.; Wise, P.; Krüger, M.; Simonsen, U.; Wehland, M.; Infanger, M.; Corydon, T.J. The role of SOX family members in solid tumours and metastasis. Semin. Cancer Biol. 2020, 67, 122–153. [Google Scholar] [CrossRef]
- Higashijima, Y.; Kanki, Y. Molecular mechanistic insights: The emerging role of SOXF transcription factors in tumorigenesis and development. Semin. Cancer Biol. 2020, 67, 39–48. [Google Scholar] [CrossRef]
- Olbromski, M.; Podhorska-Okołów, M.; Dzięgiel, P. Role of SOX Protein Groups F and H in Lung Cancer Progression. Cancers 2020, 12, 3235. [Google Scholar] [CrossRef]
- Vaidyanathan, G.; Cismowski, M.J.; Wang, G.; Vincent, T.S.; Brown, K.D.; Lanier, S.M. The Ras-related protein AGS1/RASD1 suppresses cell growth. Oncogene 2004, 23, 5858–5863. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Duan, Y.X.; Bei, C.Y.; Chen, J. Calycosin induces apoptosis by upregulation of RASD1 in human breast cancer cells MCF-7. Horm. Metab. Res. 2013, 45, 593–598. [Google Scholar] [CrossRef]
- Liu, X.J.; Li, Y.Q.; Chen, Q.Y.; Xiao, S.J.; Zeng, S.E. Up-regulating of RASD1 and apoptosis of DU-145 human prostate cancer cells induced by formononetin in vitro. Asian Pac. J. Cancer Prev. 2014, 15, 2835–2839. [Google Scholar] [CrossRef] [Green Version]
- Zellinger, B.; Bodenhofer, U.; Engländer, I.A.; Kronberger, C.; Strasser, P.; Grambozov, B.; Fastner, G.; Stana, M.; Reitsamer, R.; Sotlar, K.; et al. Hsa-miR-375/RASD1 signaling may predict local control in early breast cancer. Genes 2020, 11, 1404. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Jin, L.; Liu, G.; Wang, P.; Sun, Z.; Cao, Y.; Shi, H.; Liu, X.; Shi, Q.; Zhou, X.; et al. Overexpression of RASD1 inhibits glioma cell migration/invasion and inactivates the AKT/mTOR signaling pathway. Sci. Rep. 2017, 7, 3202. [Google Scholar] [CrossRef] [Green Version]
- Marlar, S.; Jensen, H.H.; Login, F.H.; Nejsum, L.N. Aquaporin-3 in Cancer. Int. J. Mol. Sci. 2017, 18, 2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Wan, F.; Zhang, H.; Shi, G.; Ye, D. ITGA2B and ITGA8 are predictive of prognosis in clear cell renal cell carcinoma patients. Tumour Biol. 2016, 37, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Y.; Hua, Q.; Jiang, Y.; Liu, P.; Zhang, W.; Xiang, R. Novel prognostic model based on immune signature for head and neck squamous cell carcinoma. Biomed. Res. Int. 2020, 2020, 4725314. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Meng, J.; Zhang, Y.; Gu, J.; Han, Z.; Wang, X.; Gao, S. Development and validation of a six-RNA binding proteins prognostic signature and candidate drugs for prostate cancer. Genomics 2020, 112, 4980–4992. [Google Scholar] [CrossRef] [PubMed]
- Hua, X.; Ge, S.; Chen, J.; Zhang, L.; Tai, S.; Liang, C. Effects of RNA binding proteins on the prognosis and malignant progression in prostate cancer. Front. Genet. 2020, 11, 591667. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Q.; Ju, C.L.; Wang, B.J.; Wang, R.G. PABPC1L depletion inhibits proliferation and migration via blockage of AKT pathway in human colorectal cancer cells. Oncol. Lett. 2019, 17, 3439–3445. [Google Scholar] [CrossRef]
- Gao, Q.; Zhou, R.; Meng, Y.; Duan, R.; Wu, L.; Li, R.; Deng, F.; Lin, C.; Zhao, L. Long noncoding RNA CMPK2 promotes colorectal cancer progression by activating the FUBP3-c-Myc axis. Oncogene 2020, 39, 3926–3938. [Google Scholar] [CrossRef]
- Chang, J.; Huang, L.; Cao, Q.; Liu, F. Identification of colorectal cancer-restricted microRNAs and their target genes based on high-throughput sequencing data. Onco. Targets Ther. 2016, 9, 1787–1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Gorgen, A.; Ding, A.; Du, L.; Jiang, K.; Ding, Y.; Sapisochin, G.; Ghanekar, A. Dual-specificity phosphatase 9 regulates cellular proliferation and predicts recurrence after surgery in hepatocellular carcinoma. Hepatol. Commun. 2021, 5, 1310–1328. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, T.; Barrios, A.; Tucker, A.; Collazo, J.; Arias, N.; Fazel, S.; Baker, M.; Halim, M.; Huynh, T.; Singh, R.; et al. DUSP9-mediated reduction of pERK1/2 supports cancer stem cell-like traits and promotes triple negative breast cancer. Am. J. Cancer Res. 2020, 10, 3487–3506. [Google Scholar] [PubMed]
- Qiu, Z.; Liang, N.; Huang, Q.; Sun, T.; Xue, H.; Xie, T.; Wang, X.; Wang, Q. Downregulation of DUSP9 promotes tumor progression and contributes to poor prognosis in human colorectal cancer. Front. Oncol. 2020, 10, 547011. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Lv, T.; Chen, G.; Ye, H.; Wu, W.; Li, G.; Zhi, F.C. Epigenetic silencing of DUSP9 induces the proliferation of human gastric cancer by activating JNK signaling. Oncol. Rep. 2015, 34, 121–128. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Survival | Gene Symbol | Gene ID | HR | p Value | Des (Up or Down) |
|---|---|---|---|---|---|
| Recurrence-free survival (TCGA) | ITGA2B | 3674 | 0.438 | 0.005 | ↓ |
| FBF1 | 85,302 | 0.562 | 0.042 | ↓ | |
| TMEM121 | 80,757 | 0.559 | 0.042 | ↓ | |
| CMPK2 | 129,607 | 0.567 | 0.047 | ↓ | |
| RASD1 | 51,655 | 1.969 | 0.019 | ↑ | |
| C1QTNF4 | 114,900 | 0.559 | 0.042 | ↑ | |
| PABPC1L | 80,336 | 0.341 | <0.0001 | ↓ | |
| COL8A1 | 1295 | 0.408 | 0.004 | ↑ | |
| GPNMB | 10,457 | 0.481 | 0.015 | ↑ | |
| REM2 | 161,253 | 0.516 | 0.022 | ↑ | |
| CDKN2B | 1030 | 0.548 | 0.042 | ↑ | |
| MMP11 | 4320 | 0.384 | 0.002 | ↓ | |
| SOX18 | 54,345 | 0.51 | 0.024 | ↓ | |
| CYP2E1 | 1571 | 0.371 | 0.001 | ↑ | |
| RBP7 | 116,362 | 2.221 | 0.008 | ↓ | |
| CNIH2 | 254,263 | 0.463 | 0.009 | ↓ | |
| DUSP9 | 1852 | 0.556 | 0.04 | ↓ | |
| SLC6A6 | 6533 | 0.521 | 0.027 | ↑ | |
| MAMDC4 | 158,056 | 0.374 | 0.001 | ↓ | |
| Overall survival (GSE16560) | ENG | 2022 | 1.389 | 0.02 | ↓ |
| GEM | 2669 | 1.36 | 0.03 | ↑ | |
| JUN | 3725 | 0.754 | 0.044 | ↑ | |
| AQP3 | 360 | 0.758 | 0.048 | ↑ | |
| MXD1 | 4084 | 0.709 | 0.015 | ↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Huang, Q.; A, J.; Li, L.; Li, X.; Zhang, Z.; Dong, J.-T. The Cardiac Glycoside Deslanoside Exerts Anticancer Activity in Prostate Cancer Cells by Modulating Multiple Signaling Pathways. Cancers 2021, 13, 5809. https://doi.org/10.3390/cancers13225809
Liu M, Huang Q, A J, Li L, Li X, Zhang Z, Dong J-T. The Cardiac Glycoside Deslanoside Exerts Anticancer Activity in Prostate Cancer Cells by Modulating Multiple Signaling Pathways. Cancers. 2021; 13(22):5809. https://doi.org/10.3390/cancers13225809
Chicago/Turabian StyleLiu, Mingcheng, Qingqing Huang, Jun A, Linyue Li, Xiawei Li, Zhiqian Zhang, and Jin-Tang Dong. 2021. "The Cardiac Glycoside Deslanoside Exerts Anticancer Activity in Prostate Cancer Cells by Modulating Multiple Signaling Pathways" Cancers 13, no. 22: 5809. https://doi.org/10.3390/cancers13225809