
Causes and Consequences of HPV Integration in Head and Neck Squamous Cell Carcinomas: State of the Art

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Involvement of APOBEC Mediated Anti-Viral Defense in HPV Integration
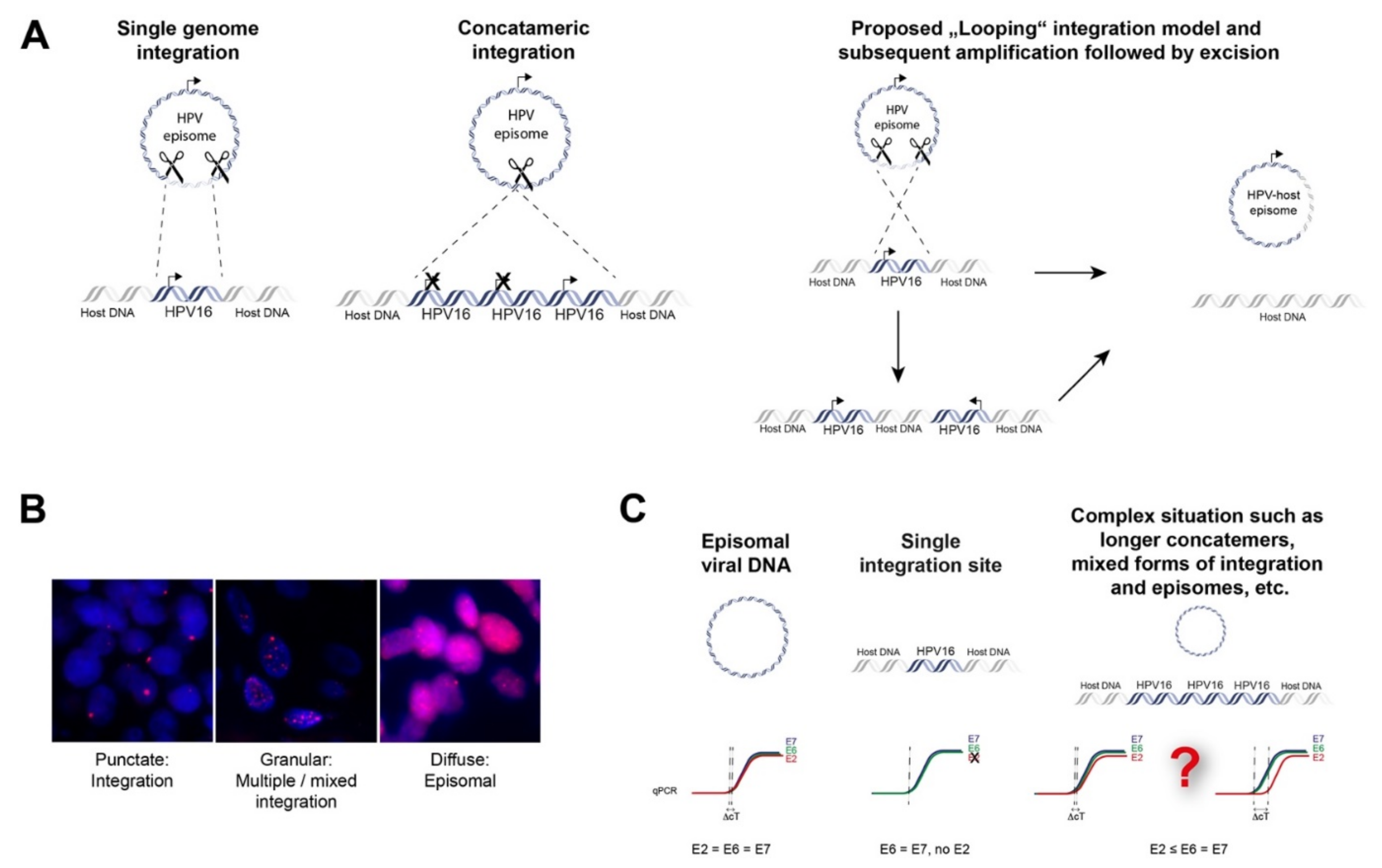
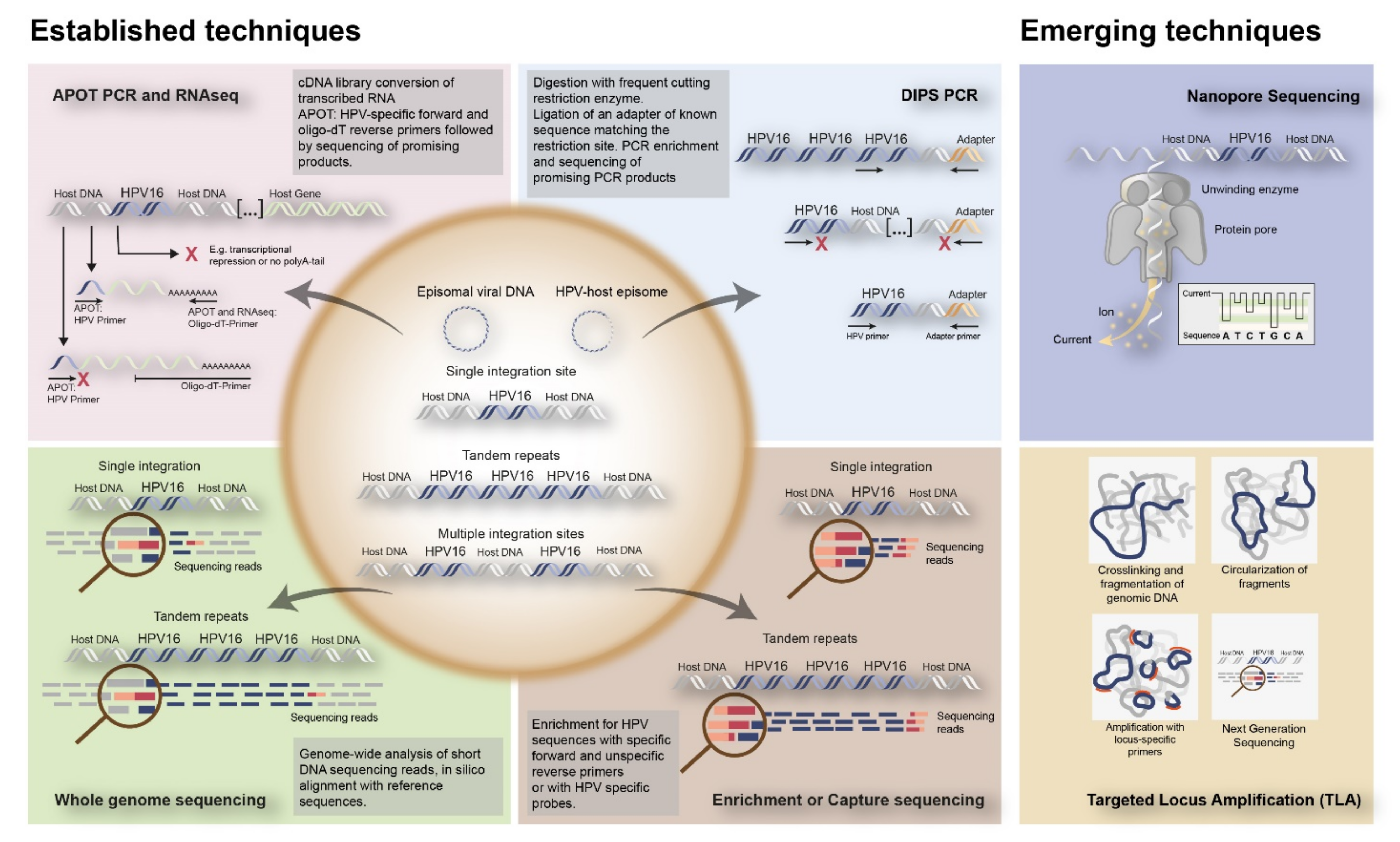
3.2. Approaches to Detect HPV Integration in Tumor Tissue
3.3. Prevalence of HPV Integration
3.4. Low HPV Copy Numbers Are Associated with Integration in Liquid Biopsy
3.5. Loci of HPV Integration in the Human Genome
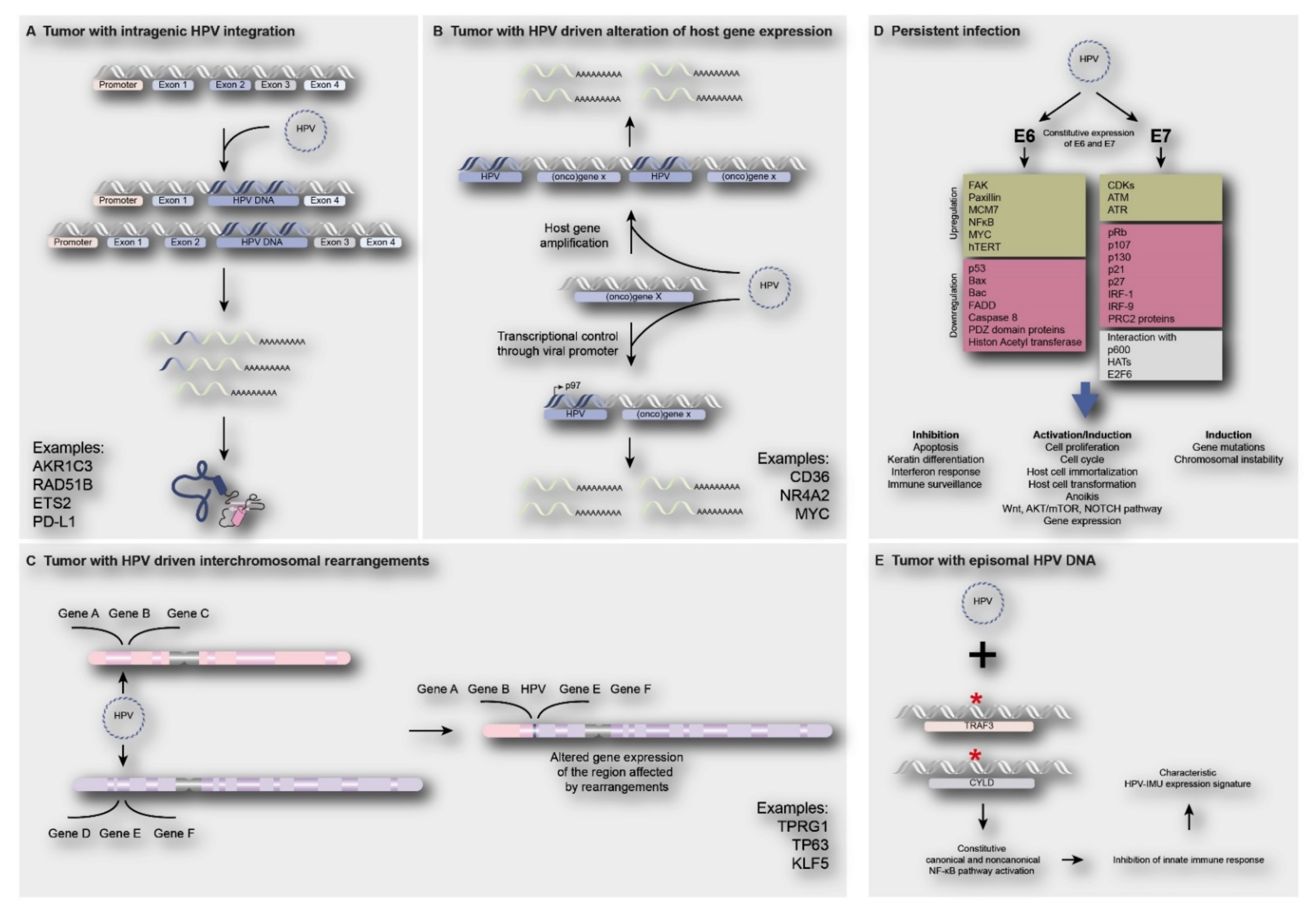
3.6. Consequences of Viral Integration
3.6.1. Deregulated Viral Gene Expression
3.6.2. Deregulated Human Gene Expression
3.6.3. Deregulated Expression of the Targeted Gene by HPV Integration
3.6.4. Deregulated Expression of Human Genes by HPV Integration
3.7. Subgroups of HPV-Positive Tumors Associated with Viral Integration Status
3.8. HPV Integration in Relation to Prognosis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A. Search Terms Used for Systematic PubMed Search
- ((Head[Tiab] OR neck[Tiab] OR “head and neck” [Tiab] OR “head-neck” OR “head-and-neck” [Tiab] OR oral[Tiab] OR pharyn*[Tiab] OR OR laryn*[Tiab] OR oropharyn*[Tiab] OR nasopharyn*[Tiab] OR hypopharyn*[Tiab] OR throat[Tiab] OR glotti*[Tiab] OR mouth[Tiab] OR palate[Tiab] OR gingiva*[Tiab] OR lip[Tiab] OR cheek[Tiab] OR bucc*[Tiab] OR gum*[Tiab] OR tonsil*[Tiab] OR tongue[Tiab] OR nasal[Tiab] OR paranasal[Tiab] OR sinus[Tiab] OR saliv*[Tiab] OR ent[Tiab] OR aerodigestive[Tiab] OR “aero digestive” [Tiab] OR aero-digestive[Tiab])
- AND (cancer* OR carcinoma* OR neoplas* OR tumor* OR tumour* OR malignan* OR SCC OR “Neoplasms”[Mesh])) OR (hnscc[Tiab] OR scchn[Tiab] OR “Head and Neck Neoplasms”[Mesh])
- AND
- (“Human papilloma virus” [Tiab] OR “Human papilloma viruses” [Tiab] OR “Papillomavirus, Human” [Tiab] OR “Human papillomavirus” [Tiab] OR HPV [Tiab] OR HR-HPV [Tiab] OR “High-risk HPV” [Tiab] OR “HPV infection*” [Tiab] OR “Papillomavirus Infections/pathology” [Mesh])
- AND
- (integration [Tiab] OR “virus integration*” [Tiab] OR “virus integration” [Mesh] OR “Viral integration*” [Tiab] OR “human papillomavirus integration” [Tiab] OR “HPV integration” [Tiab] OR “genome integration” [Tiab] OR “viral DNA integration” [Tiab] OR “virus DNA integration” [Tiab] OR “HPV DNA integration” [Tiab] OR “HPV insertion*” [Tiab] OR “Human papillomavirus insertion*” [Tiab])
References
- Vigneswaran, N.; Williams, M.D. Epidemiologic trends in head and neck cancer and aids in diagnosis. Oral Maxillofac. Surg. Clin. N. Am. 2014, 26, 123–141. [Google Scholar]
- Leemans, C.R.; Braakhuis, B.J.M.; Brakenhoff, R.H. The molecular biology of head and neck cancer. Nat. Rev. Cancer 2010, 11, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Marur, S.; Forastiere, A.A. Head and Neck Squamous Cell Carcinoma: Update on Epidemiology, Diagnosis, and Treatment. Mayo Clin. Proc. 2016, 91, 386–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsahafi, E.; Begg, K.; Amelio, I.; Raulf, N.; Lucarelli, P.; Sauter, T.; Tavassoli, M. Clinical update on head and neck cancer: Molecular biology and ongoing challenges. Cell Death Dis. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Seiwert, T.Y.; Burtness, B.; Mehra, R.; Weiss, J.; Berger, R.; Eder, J.P.; Heath, K.; McClanahan, T.; Lunceford, J.; Gause, C.; et al. Safety and clinical activity of pembrolizumab for treatment of recurrent or metastatic squamous cell carcinoma of the head and neck (KEYNOTE-012): An open-label, multicentre, phase 1b trial. Lancet Oncol. 2016, 17, 956–965. [Google Scholar] [CrossRef]
- Vermorken, J.B.; Trigo, J.; Hitt, R.; Koralewski, P.; Diaz-Rubio, E.; Rolland, F.; Knecht, R.; Amellal, N.; Schueler, A.; Baselga, J. Open-label, uncontrolled, multicenter phase II study to evaluate the efficacy and toxicity of cetuximab as a single agent in pa-tients with recurrent and/or metastatic squamous cell carcinoma of the head and neck who failed to respond to platinum-based therapy. J. Clin. Oncol. 2007, 25, 2171–2177. [Google Scholar]
- Rietbergen, M.M.; Leemans, C.R.; Bloemena, E.; Heideman, D.A.; Braakhuis, B.J.; Hesselink, A.T.; Witte, B.I.; Baatenburg de Jong, R.J.; Meijer, C.J.; Snijders, P.J.; et al. Increasing prevalence rates of HPV attributable oropharyngeal squamous cell car-cinomas in the Netherlands as assessed by a validated test algorithm. Int. J. Cancer 2013, 132, 1565–1571. [Google Scholar] [CrossRef]
- Olthof, N.C.; Huebbers, C.U.; Kolligs, J.; Henfling, M.; Ramaekers, F.C.; Cornet, I.; Van Lent-Albrechts, J.A.; Stegmann, A.P.; Silling, S.; Wieland, U.; et al. Viral load, gene expression and mapping of viral integration sites in HPV16-associated HNSCC cell lines. Int. J. Cancer 2014, 136, E207–E218. [Google Scholar] [CrossRef] [Green Version]
- Faraji, F.; Zaidi, M.; Fakhry, C.; Gaykalova, D.A. Molecular mechanisms of human papillomavirus-related carcinogenesis in head and neck cancer. Microbes Infect. 2017, 19, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Pinatti, L.; Walline, H.; Carey, T. Human Papillomavirus Genome Integration and Head and Neck Cancer. J. Dent. Res. 2017, 97, 691–700. [Google Scholar] [CrossRef]
- Olthof, N.C.; Straetmans, J.M.; Snoeck, R.; Ramaekers, F.C.; Kremer, B.; Speel, E.J. Next-generation treatment strategies for human papillomavirus-related head and neck squamous cell carcinoma: Where do we go? Rev. Med. Virol. 2012, 22, 88–105. [Google Scholar] [CrossRef] [Green Version]
- Elrefaey, S.; Massaro, M.; Chiocca, S.; Chiesa, F.; Ansarin, M. HPV in oropharyngeal cancer: The basics to know in clinical practice. Acta Otorhinolaryngol. Ital. 2014, 34, 299–309. [Google Scholar]
- Woodman, C.B.J.; Collins, S.I.; Young, L. The natural history of cervical HPV infection: Unresolved issues. Nat. Rev. Cancer 2007, 7, 11–22. [Google Scholar] [CrossRef]
- Ozbun, M.A. Extracellular events impacting human papillomavirus infections: Epithelial wounding to cell signaling involved in virus entry. Papillomavirus Res. 2019, 7, 188–192. [Google Scholar] [CrossRef]
- Speel, E.J.M. HPV Integration in Head and Neck Squamous Cell Carcinomas: Cause and Consequence. HPV Infect. Head Neck Cancer 2016, 206, 57–72. [Google Scholar] [CrossRef]
- Williams, V.; Filippova, M.; Soto, U.; Duerksen-Hughes, P.J. HPV-DNA integration and carcinogenesis: Putative roles for inflammation and oxidative stress. Futur. Virol. 2011, 6, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Lace, M.J.; Anson, J.R.; Haugen, T.; Dierdorff, J.M.; Turek, L.P. Interferon treatment of human keratinocytes harboring extrachromosomal, persistent HPV-16 plasmid genomes induces de novo viral integration. Carcinogenesis 2014, 36, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visalli, G.; Riso, R.; Facciolà, A.; Mondello, P.; Caruso, C.; Picerno, I.; Di Pietro, A.; Spataro, P.; Bertuccio, M.P. Higher levels of oxidative DNA damage in cervical cells are correlated with the grade of dysplasia and HPV infection. J. Med. Virol. 2015, 88, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Gravitt, P.E.; Song, H.; Maldonado, A.M.; Ozbun, M.A. Nitric Oxide Induces Early Viral Transcription Coincident with Increased DNA Damage and Mutation Rates in Human Papillomavirus–Infected Cells. Cancer Res. 2009, 69, 4878–4884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leeman, J.E.; Li, Y.; Bell, A.; Hussain, S.; Majumdar, R.; Rong-Mullins, X.; Blecua, P.; Damerla, R.; Narang, H.; Ravindran, P.; et al. Human papillomavirus 16 promotes microhomology-mediated end-joining. Proc. Natl. Acad. Sci. 2019, 116, 21573–21579. [Google Scholar] [CrossRef] [Green Version]
- Groves, I.J.; Coleman, N. Human papillomavirus genome integration in squamous carcinogenesis: What have next-generation sequencing studies taught us? J. Pathol. 2018, 245, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akagi, K.; Li, J.; Broutian, T.R.; Padilla-Nash, H.; Xiao, W.; Jiang, B.; Rocco, J.W.; Teknos, T.N.; Kumar, B.; Wangsa, D.; et al. Genome-wide analysis of HPV integration in human cancers reveals recurrent, focal genomic instability. Genome Res. 2013, 24, 185–199. [Google Scholar] [CrossRef] [Green Version]
- Rusan, M.; Li, Y.Y.; Hammerman, P.S. Genomic Landscape of Human Papillomavirus–Associated Cancers. Clin. Cancer Res. 2015, 21, 2009–2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, S.; Wakae, K.; Wakisaka, N.; Nakanishi, Y.; Ishikawa, K.; Komori, T.; Moriyama-Kita, M.; Endo, K.; Murono, S.; Wang, Z.; et al. APOBEC3A associates with human papillomavirus genome integration in oropharyngeal cancers. Oncogene 2016, 36, 1687–1697. [Google Scholar] [CrossRef]
- Zapatka, M.; Pathogens, P.; Borozan, I.; Brewer, D.S.; Iskar, M.; Grundhoff, A.; Alawi, M.; Desai, N.; Sültmann, H.; Moch, H.; et al. The landscape of viral associations in human cancers. Nat. Genet. 2020, 52, 320–330. [Google Scholar] [CrossRef] [Green Version]
- Kukurba, K.R.; Montgomery, S. RNA Sequencing and Analysis. Cold Spring Harb. Protoc. 2015, 2015, 951–969. [Google Scholar] [CrossRef] [Green Version]
- Nouri-Aria, K.T. Allergy methods and protocols. In Situ Hybridization; Jones, M.G., Lympany, P., Eds.; Humana: Totowa, NJ, USA, 2008; Volume 138. [Google Scholar]
- Abreu, A.L.P.; Souza, R.P.; Gimenes, F.; Consolaro, M.E.L. A review of methods for detect human Papillomavirus infection. Virol. J. 2012, 9, 262. [Google Scholar] [CrossRef] [Green Version]
- Morgan, I.M.; DiNardo, L.J.; Windle, B. Integration of Human Papillomavirus Genomes in Head and Neck Cancer: Is It Time to Consider a Paradigm Shift? Viruses 2017, 9, 208. [Google Scholar] [CrossRef]
- Huebbers, C.U.; Verhees, F.; Poluschkin, L.; Olthof, N.C.; Kolligs, J.; Siefer, O.G.; Henfling, M.; Ramaekers, F.C.; Preuss, S.F.; Beutner, D.; et al. Upregulation of AKR1C1 and AKR1C3 expression in OPSCC with integrated HPV16 and HPV-negative tumors is an indicator of poor prognosis. Int. J. Cancer 2019, 144, 2465–2477. [Google Scholar] [CrossRef]
- Luft, F.; Klaes, R.; Nees, M.; Durst, M.; Heilmann, V.; Melsheimer, P.; von Knebel Doeberitz, M. Detection of integrated pap-illomavirus sequences by ligation-mediated PCR (DIPS-PCR) and molecular characterization in cervical cancer cells. Int. J. Cancer 2001, 92, 9–17. [Google Scholar] [CrossRef]
- Olthof, N.C.; Speel, E.J.; Kolligs, J.; Haesevoets, A.; Henfling, M.; Ramaekers, F.C.; Preuss, S.F.; Drebber, U.; Wieland, U.; Silling, S.; et al. Comprehensive analysis of HPV16 integration in OSCC reveals no significant impact of physical status on viral on-cogene and virally disrupted human gene expression. PLoS ONE 2014, 9, e88718. [Google Scholar] [CrossRef]
- Ziegert, C.; Wentzensen, N.; Vinokurova, S.; Kisseljov, F.; Einenkel, J.; Hoeckel, M.; Doeberitz, M.V.K. A comprehensive analysis of HPV integration loci in anogenital lesions combining transcript and genome-based amplification techniques. Oncogene 2003, 22, 3977–3984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gradíssimo, A.; Burk, R.D. Molecular tests potentially improving HPV screening and genotyping for cervical cancer prevention. Expert Rev. Mol. Diagn. 2017, 17, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Petersen, B.-S.; Fredrich, B.; Hoeppner, M.P.; Ellinghaus, D.; Franke, A. Opportunities and challenges of whole-genome and -exome sequencing. BMC Genet. 2017, 18, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harlé, A.; Guillet, J.; Thomas, J.; Demange, J.; Dolivet, G.; Peiffert, D.; Leroux, A.; Sastre-Garau, X. HPV insertional pattern as a personalized tumor marker for the optimized tumor diagnosis and follow-up of patients with HPV-associated carcinomas: A case report. BMC Cancer 2019, 19, 277. [Google Scholar] [CrossRef] [PubMed]
- Kono, N.; Arakawa, K. Nanopore sequencing: Review of potential applications in functional genomics. Dev. Growth Differ. 2019, 61, 316–326. [Google Scholar] [CrossRef] [Green Version]
- De Vree, P.J.P.; de Wit, E.; Yilmaz, M.; Van De Heijning, M.; Klous, P.; Verstegen, M.J.A.M.; Wan, Y.; Teunissen, H.; Krijger, P.; Geeven, G.; et al. Targeted sequencing by proximity ligation for comprehensive variant detection and local haplotyping. Nat. Biotechnol. 2014, 32, 1019–1025. [Google Scholar] [CrossRef]
- Chen, Y.; Yao, H.; Thompson, E.J.; Tannir, N.M.; Weinstein, J.N.; Su, X. VirusSeq: Software to identify viruses and their inte-gration sites using next-generation sequencing of human cancer tissue. Bioinformatics 2013, 29, 266–267. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Jia, P.; Zhao, Z. VirusFinder: Software for Efficient and Accurate Detection of Viruses and Their Integration Sites in Host Genomes through Next Generation Sequencing Data. PLoS ONE 2013, 8, e64465. [Google Scholar] [CrossRef]
- Rajaby, R.; Zhou, Y.; Meng, Y.; Zeng, X.; Li, G.; Wu, P.; Sung, W.-K. SurVirus: A repeat-aware virus integration caller. Nucleic Acids Res. 2021, 49, e33. [Google Scholar] [CrossRef]
- Khan, A.; Liu, Q.; Chen, X.; Stucky, A.; Sedghizadeh, P.; Adelpour, D.; Zhang, X.; Wang, K.; Zhong, J.F. Detection of human papillomavirus in cases of head and neck squamous cell carcinoma by RNA-seq and VirTect. Mol. Oncol. 2018, 13, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Tian, S.; Wang, P.; Zang, Y.; Chen, X.; Yao, Y.; Li, W. The characteristics of HPV integration in cervical intraepithelial cells. J. Cancer 2019, 10, 2783–2787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baheti, S.; Tang, X.; O’Brien, D.R.; Chia, N.; Roberts, L.R.; Nelson, H.; Boughey, J.C.; Wang, L.; Goetz, M.P.; Kocher, J.-P.A.; et al. HGT-ID: An efficient and sensitive workflow to detect human-viral insertion sites using next-generation sequencing data. BMC Bioinform. 2018, 19, 271. [Google Scholar] [CrossRef]
- Li, J.W.; Wan, R.; Yu, A.C.-S.; Na Co, N.; Wong, N.; Chan, T.-F. ViralFusionSeq: Accurately discover viral integration events and reconstruct fusion transcripts at single-base resolution. Bioinformatics 2013, 29, 649–651. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, T.B.; Dantzer, J.; Peters, B.; Dinauer, M.; Mockaitis, K.; Mooney, S.; Cornetta, K. Identifying viral integration sites using SeqMap 2.0. Bioinformatics 2011, 27, 720–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, D.W.; Sze, K.M.; Ng, I.O. Virus-Clip: A fast and memory-efficient viral integration site detection tool at single-base reso-lution with annotation capability. Oncotarget 2015, 6, 20959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tennakoon, C.; Sung, W.K. BATVI: Fast, sensitive and accurate detection of virus integrations. BMC Bioinform. 2017, 18, 71–111. [Google Scholar] [CrossRef] [Green Version]
- Forster, M.; Szymczak, S.; Ellinghaus, D.; Hemmrich, G.; Rühlemann, M.; Kraemer, L.; Mucha, S.; Wienbrandt, L.; Stanulla, M.; Franke, A. Vy-PER: Eliminating false positive detection of virus integration events in next generation sequencing data. Sci. Rep. 2015, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Zhu, D.; Wang, W.; Li, W.; Jia, W.; Zeng, X.; Ding, W.; Yu, L.; Wang, X.; Ma, D.; et al. Genome-wide profiling of HPV integration in cervical cancer identifies clustered genomic hot spots and a potential microhomology-mediated integration mechanism. Nat. Genet. 2015, 47, 158–163. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, C.; Gao, W.; Wang, L.; Pan, Y.; Gao, Y.; Lu, Z.; Ke, Y. Genome-wide profiling of the human papillomavirus DNA integration in cervical intraepithelial neoplasia and normal cervical epithelium by HPV capture technology. Sci. Rep. 2016, 6, 35427. [Google Scholar] [CrossRef] [PubMed]
- Dyer, N.; Young, L.; Ott, S. Artifacts in the data of Hu et al. Nat. Genet. 2015, 48, 2–3. [Google Scholar] [CrossRef]
- Nguyen, N.-P.; Deshpande, V.; Luebeck, J.; Mischel, P.S.; Bafna, V. ViFi: Accurate detection of viral integration and mRNA fusion reveals indiscriminate and unregulated transcription in proximal genomic regions in cervical cancer. Nucleic Acids Res. 2018, 46, 3309–3325. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Jia, P.; Zhao, Z. VERSE: A novel approach to detect virus integration in host genomes through reference genome customization. Genome Med. 2015, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bs, L.M.P.; Gu, W.; Wang, Y.; Bs, A.E.; Ms, A.D.B.; Ba, C.V.B.; Carey, T.E.; Mills, R.E.; Brenner, J.C. SearcHPV: A novel approach to identify and assemble human papillomavirus–host genomic integration events in cancer. Cancer 2021. [Google Scholar] [CrossRef]
- Cameron, D.L.; Jacobs, N.; Roepman, P.; Priestley, P.; Cuppen, E.; Papenfuss, A.T. VIRUSBreakend: Viral Integration Recognition Using Single Breakends. Bioinformatics 2021. [Google Scholar] [CrossRef]
- Castellsagué, X.; Alemany, L.; Quer, M.; Halec, G.; Quirós, B.; Tous, S.; Clavero, O.; Alòs, L.; Biegner, T.; Szafarowski, T.; et al. HPV Involvement in Head and Neck Cancers: Comprehensive Assessment of Biomarkers in 3680 Patients. J. Natl. Cancer Inst. 2016, 108, djv403. [Google Scholar] [CrossRef] [PubMed]
- Hafkamp, H.C.; Speel, E.J.; Haesevoets, A.; Bot, F.J.; Dinjens, W.N.; Ramaekers, F.C.; Hopman, A.H.; Manni, J.J. A subset of head and neck squamous cell carcinomas exhibits integration of HPV 16/18 DNA and overexpression of p16INK4A and p53 in the absence of mutations in p53 exons 5-8. Int. J. Cancer 2003, 107, 394–400. [Google Scholar] [CrossRef]
- Tang, K.D.; Baeten, K.; Kenny, L.; Frazer, I.H.; Scheper, G.; Punyadeera, C. Unlocking the Potential of Saliva-Based Test to Detect HPV-16-Driven Oropharyngeal Cancer. Cancers 2019, 11, 473. [Google Scholar] [CrossRef] [Green Version]
- Parfenov, M.; Pedamallu, C.S.; Gehlenborg, N.; Freeman, S.; Danilova, L.; Bristow, C.A.; Lee, S.; Hadjipanayis, A.G.; Ivanova, E.V.; Wilkerson, M.D.; et al. Characterization of HPV and host genome interactions in primary head and neck cancers. Proc. Natl. Acad. Sci. 2014, 111, 15544–15549. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; Hasegawa, M.; Kiyuna, A.; Matayoshi, S.; Uehara, T.; Agena, S.; Yamashita, Y.; Ogawa, K.; Maeda, H.; Suzuki, M. Viral load, physical status, and E6/E7 mRNA expression of human papillomavirus in head and neck squamous cell carcinoma. Head Neck 2013, 35, 800–808. [Google Scholar] [CrossRef]
- Gao, G.; Wang, J.; Kasperbauer, J.L.; Tombers, N.M.; Teng, F.; Gou, H.; Zhao, Y.; Bao, Z.; Smith, D.I. Whole genome sequencing reveals complexity in both HPV sequences present and HPV integrations in HPV-positive oropharyngeal squamous cell car-cinomas. BMC Cancer 2019, 19, 352. [Google Scholar] [CrossRef]
- Wuerdemann, N.; Jain, R.; Adams, A.; Speel, E.-J.M.; Wagner, S.; Joosse, S.A.; Klussmann, J.P. Cell-Free HPV-DNA as a Biomarker for Oropharyngeal Squamous Cell Carcinoma—A Step Towards Personalized Medicine? Cancers 2020, 12, 2997. [Google Scholar] [CrossRef]
- Chera, B.S.; Kumar, S.; Beaty, B.T.; Marron, D.; Jefferys, S.; Green, R.; Goldman, E.C.; Amdur, R.; Sheets, N.; Dagan, R.; et al. Rapid Clearance Profile of Plasma Circulating Tumor HPV Type 16 DNA during Chemoradiotherapy Correlates with Disease Control in HPV-Associated Oropharyngeal Cancer. Clin. Cancer Res. 2019, 25, 4682–4690. [Google Scholar] [CrossRef]
- Bodelon, C.; Untereiner, M.E.; Machiela, M.J.; Vinokurova, S.; Wentzensen, N. Genomic characterization of viral integration sites in HPV-related cancers. Int. J. Cancer 2016, 139, 2001–2011. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.Z.; Flam, E.L.; Izumchenko, E.; Danilova, L.V.; Wulf, H.A.; Guo, T.; Singman, D.A.; Afsari, B.; Skaist, A.M.; Considine, M.; et al. Integrated Analysis of Whole-Genome ChIP-Seq and RNA-Seq Data of Primary Head and Neck Tumor Samples Associates HPV Integration Sites with Open Chromatin Marks. Cancer Res. 2017, 77, 6538–6550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walline, H.M.; Komarck, C.M.; McHugh, J.B.; Bellile, E.L.; Brenner, J.C.; Prince, M.E.; McKean, E.L.; Chepeha, D.B.; Wolf, G.T.; Worden, F.P.; et al. Genomic Integration of High-Risk HPV Alters Gene Expression in Oropharyngeal Squamous Cell Carci-noma. Mol. Cancer Res. 2016, 14, 941–952. [Google Scholar] [CrossRef] [Green Version]
- Pinatti, L.M.; Walline, H.M.; Carey, T.E.; Klussmann, J.P.; Huebbers, C.U. Viral Integration Analysis Reveals Likely Common Clonal Origin of Bilateral HPV16-Positive, p16-Positive Tonsil Tumors. Arch. Clin. Med. Case Rep. 2020, 4, 680–696. [Google Scholar] [CrossRef]
- Reuschenbach, M.; Huebbers, C.; Prigge, E.-S.; Bermejo, J.L.; Kalteis, M.S.; Preuss, S.F.; Seuthe, I.M.C.; Kolligs, J.; Speel, E.-J.M.; Olthof, N.; et al. Methylation status of HPV16 E2-binding sites classifies subtypes of HPV-associated oropharyngeal cancers. Cancer 2015, 121, 1966–1976. [Google Scholar] [CrossRef]
- Khanal, S.; Shumway, B.S.; Zahin, M.; Redman, R.A.; Strickley, J.D.; Trainor, P.J.; Rai, S.N.; Ghim, S.-J.; Jenson, A.B.; Joh, J. Viral DNA integration and methylation of human papillomavirus type 16 in high-grade oral epithelial dysplasia and head and neck squamous cell carcinoma. Oncotarget 2018, 9, 30419–30433. [Google Scholar] [CrossRef]
- Hatano, T.; Sano, D.; Takahashi, H.; Hyakusoku, H.; Isono, Y.; Shimada, S.; Sawakuma, K.; Takada, K.; Oikawa, R.; Watanabe, Y.; et al. Identification of human papillomavirus (HPV) 16 DNA integration and the ensuing patterns of methylation in HPV-associated head and neck squamous cell carcinoma cell lines. Int. J. Cancer 2016, 140, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Hausen, H.Z. Papillomaviruses and cancer: From basic studies to clinical application. Nat. Rev. Cancer 2002, 2, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Pim, D.; Banks, L. Interaction of viral oncoproteins with cellular target molecules: Infection with high-risk vs low-risk human papillomaviruses. APMIS 2010, 118, 471–493. [Google Scholar] [CrossRef]
- Arenz, R.N.A.; Ziemann, F.; Mayer, C.; Wittig, A.; Dreffke, K.; Preising, S.; Wagner, S.; Klussmann, J.-P.; Engenhart-Cabillic, R.; Wittekindt, C. Increased radiosensitivity of HPV-positive head and neck cancer cell lines due to cell cycle dysregulation and induction of apoptosis. Strahlenther. Onkol. 2014, 190, 839–846. [Google Scholar] [CrossRef]
- Rieckmann, T.; Tribius, S.; Grob, T.J.; Meyer, F.; Busch, C.-J.; Petersen, C.; Dikomey, E.; Kriegs, M. HNSCC cell lines positive for HPV and p16 possess higher cellular radiosensitivity due to an impaired DSB repair capacity. Radiother. Oncol. 2013, 107, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Penning, T.M. Aldo-Keto Reductase Regulation by the Nrf2 System: Implications for Stress Response, Chemotherapy Drug Resistance, and Carcinogenesis. Chem. Res. Toxicol. 2016, 30, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Groves, I.J.; Coleman, N. Pathogenesis of human papillomavirus-associated mucosal disease. J. Pathol. 2015, 235, 527–538. [Google Scholar] [CrossRef] [Green Version]
- Hajek, M.; Sewell, A.; Kaech, S.; Burtness, B.; Yarbrough, W.G.; Issaeva, N. TRAF3/CYLDmutations identify a distinct subset of human papillomavirus-associated head and neck squamous cell carcinoma. Cancer 2017, 123, 1778–1790. [Google Scholar] [CrossRef] [Green Version]
- Hassounah, N.B.; Malladi, V.; Huang, Y.; Freeman, S.; Beauchamp, E.M.; Koyama, S.; Souders, N.; Martin, S.; Dranoff, G.; Wong, K.-K.; et al. Identification and characterization of an alternative cancer-derived PD-L1 splice variant. Cancer Immunol. Immunother. 2018, 68, 407–420. [Google Scholar] [CrossRef]
- Koneva, L.A.; Zhang, Y.; Virani, S.; Hall, P.B.; McHugh, J.B.; Chepeha, D.; Wolf, G.T.; Carey, T.E.; Rozek, L.S.; Sartor, M.A. HPV Integration in HNSCC Correlates with Survival Outcomes, Immune Response Signatures, and Candidate Drivers. Mol. Cancer Res. 2017, 16, 90–102. [Google Scholar] [CrossRef] [Green Version]
- Broutian, T.R.; Jiang, B.; Li, J.; Akagi, K.; Gui, S.; Zhou, Z.; Xiao, W.; Symer, D.E.; Gillison, M.L. Human papillomavirus inser-tions identify the PIM family of serine/threonine kinases as targetable driver genes in head and neck squamous cell carcinoma. Cancer Lett. 2020, 476, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Beier, U.H.; Weise, J.B.; Laudien, M.; Sauerwein, H.; Görögh, T. Overexpression of Pim-1 in head and neck squamous cell carcinomas. Int. J. Oncol. 2007, 30, 1381–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peltola, K.; Hollmen, M.; Maula, S.-M.; Rainio, E.; Ristamäki, R.; Luukkaa, M.; Sandholm, J.; Sundvall, M.; Elenius, K.; Koskinen, P.J.; et al. Pim-1 Kinase Expression Predicts Radiation Response in Squamocellular Carcinoma of Head and Neck and Is under the Control of Epidermal Growth Factor Receptor. Neoplasia 2009, 11, 629-IN1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, W.-F.; Yen, C.-Y.; Lin, C.-N.; Liaw, G.-A.; Chiu, C.-T.; Hsia, Y.-J.; Liu, S.-Y. Up-regulation of a serine–threonine kinase proto-oncogene Pim-1 in oral squamous cell carcinoma. Int. J. Oral Maxillofac. Surg. 2006, 35, 740–745. [Google Scholar] [CrossRef]
- Huebbers, C.U.; Preuss, S.F.; Kolligs, J.; Vent, J.; Stenner, M.; Wieland, U.; Silling, S.; Drebber, U.; Speel, E.-J.M.; Klussmann, J.P. Integration of HPV6 and downregulation of AKR1C3 expression mark malignant transformation in a patient with juve-nile-onset laryngeal papillomatosis. PLoS ONE 2013, 8, e57207. [Google Scholar] [CrossRef] [PubMed]
- Wanichwatanadecha, P.; Sirisrimangkorn, S.; Kaewprag, J.; Ponglikitmongkol, M. Transactivation activity of human papil-lomavirus type 16 E6*I on aldo-keto reductase genes enhances chemoresistance in cervical cancer cells. J. Gen. Virol. 2012, 93, 1081–1092. [Google Scholar] [CrossRef]
- Ruiz, F.X.; Porté, S.; Parés, X.; Farrés, J. Biological Role of Aldo–Keto Reductases in Retinoic Acid Biosynthesis and Signaling. Front. Pharmacol. 2012, 3, 58. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Koneva, L.A.; Virani, S.; Arthur, A.E.; Virani, A.; Hall, P.B.; Warden, C.D.; Carey, T.E.; Chepeha, D.; Prince, M.E.; et al. Subtypes of HPV-Positive Head and Neck Cancers Are Associated with HPV Characteristics, Copy Number Alterations, PIK3CA Mutation, and Pathway Signatures. Clin. Cancer Res. 2016, 22, 4735–4745. [Google Scholar] [CrossRef] [Green Version]
- Smeets, S.J.; Hesselink, A.T.; Speel, E.-J.M.; Haesevoets, A.; Snijders, P.J.; Pawlita, M.; Meijer, C.J.; Braakhuis, B.J.; Leemans, C.R.; Brakenhoff, R.H. A novel algorithm for reliable detection of human papillomavirus in paraffin embedded head and neck cancer specimen. Int. J. Cancer 2007, 121, 2465–2472. [Google Scholar] [CrossRef]
- Paget-Bailly, P.; Meznad, K.; Bruyère, D.; Perrard, J.; Herfs, M.; Jung, A.; Mougin, C.; Prétet, J.-L.; Baguet, A. Comparative RNA sequencing reveals that HPV16 E6 abrogates the effect of E6*I on ROS metabolism. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Qin, T.; Koneva, L.A.; Liu, Y.; Zhang, Y.; Arthur, A.E.; Zarins, K.R.; Carey, T.E.; Chepeha, D.; Wolf, G.T.; Rozek, L.S. Significant association between host transcriptome-derived HPV oncogene E6* influence score and carcinogenic pathways, tumor size, and survival in head and neck cancer. Head Neck 2020, 42, 2375–2389. [Google Scholar] [CrossRef]
- Pannone, G.; Bufo, P.; Pace, M.; Lepore, S.; Russo, G.M.; Rubini, C.; Franco, R.; Aquino, G.; Santoro, A.; Campisi, G.; et al. TLR4 down-regulation identifies high risk HPV infection and integration in head and neck squamous cell carcinomas. Front. Biosci. 2016, 8. [Google Scholar]
- Yang, W.; Liu, Y.; Dong, R.; Liu, J.; Lang, J.; Yang, J.; Wang, W.; Li, J.; Meng, B.; Tian, G. Accurate Detection of HPV Integration Sites in Cervical Cancer Samples Using the Nanopore MinION Sequencer Without Error Correction. Front. Genet. 2020, 11, 660. [Google Scholar] [CrossRef]
- Cancer Genome Atlas, N. Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature 2015, 517, 576–582. [Google Scholar] [CrossRef] [Green Version]
- Häcker, H.; Tseng, P.-H.; Karin, M. Expanding TRAF function: TRAF3 as a tri-faced immune regulator. Nat. Rev. Immunol. 2011, 11, 457–468. [Google Scholar] [CrossRef]
- Harhaj, E.W.; Dixit, V.M. Regulation of NF-kappaB by deubiquitinases. Immunol. Rev. 2012, 246, 107–124. [Google Scholar] [CrossRef] [Green Version]
- Keck, M.K.; Zuo, Z.; Khattri, A.; Stricker, T.P.; Brown, C.D.; Imanguli, M.; Rieke, D.; Endhardt, K.; Fang, P.; Brägelmann, J.; et al. Integrative Analysis of Head and Neck Cancer Identifies Two Biologically Distinct HPV and Three Non-HPV Subtypes. Clin. Cancer Res. 2014, 21, 870–881. [Google Scholar] [CrossRef] [Green Version]
- Locati, L.D.; Serafini, M.S.; Ianno’, M.F.; Carenzo, A.; Orlandi, E.; Resteghini, C.; Cavalieri, S.; Bossi, P.; Canevari, S.; Licitra, L.; et al. Mining of Self-Organizing Map Gene-Expression Portraits Reveals Prognostic Stratification of HPV-Positive Head and Neck Squamous Cell Carcinoma. Cancers 2019, 11, 1057. [Google Scholar] [CrossRef] [Green Version]
- Nulton, T.J.; Kim, N.-K.; DiNardo, L.J.; Morgan, I.M.; Windle, B. Patients with integrated HPV16 in head and neck cancer show poor survival. Oral Oncol. 2018, 80, 52–55. [Google Scholar] [CrossRef] [PubMed]
- Veitía, D.; Liuzzi, J.; Ávila, M.; Rodriguez, I.; Toro, F.; Correnti, M. Association of viral load and physical status of HPV-16 with survival of patients with head and neck cancer. ecancermedicalscience 2020, 14. [Google Scholar] [CrossRef] [PubMed]
- Niya, M.H.K.; Keyvani, H.; Tameshkel, F.S.; Salehi-Vaziri, M.; Teaghinezhad-S, S.; Salim, F.B.; Monavari, S.H.R.; Javanmard, D. Human Papillomavirus Type 16 Integration Analysis by Real-time PCR Assay in Associated Cancers. Transl. Oncol. 2018, 11, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, A.; Rautava, J.; Kero, K.; Syrjänen, K.; Longatto-Filho, A.; Grenman, S.; Syrjänen, S. Physical state and copy numbers of HPV16 in oral asymptomatic infections that persisted or cleared during the 6-year follow-up. J. Gen. Virol. 2017, 98, 681–689. [Google Scholar] [CrossRef]
- Walline, H.M.; Bs, C.M.G.; McHugh, J.B.; Tang, A.L.; Owen, J.H.; Teh, B.T.; Mckean, E.; Glover, T.W.; Bs, M.P.G.; Prince, M.E.; et al. Integration of high-risk human papillomavirus into cellular cancer-related genes in head and neck cancer cell lines. Head Neck 2017, 39, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Vojtěchová, Z.; Sabol, I.; Saláková, M.; Turek, L.; Grega, M.; Šmahelová, J.; Vencalek, O.; Lukesova, E.; Klozar, J.; Tachezy, R. Analysis of the integration of human papillomaviruses in head and neck tumours in relation to patients’ prognosis. Int. J. Cancer 2015, 138, 386–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, M.Y.; Dahlstrom, K.R.; Sturgis, E.M.; Li, G. Human papillomavirus integration pattern and demographic, clinical, and survival characteristics of patients with oropharyngeal squamous cell carcinoma. Head Neck 2016, 38, 1139–1144. [Google Scholar] [CrossRef] [PubMed]
- Bs, L.M.P.; Sinha, H.N.; Ba, C.V.B.; Bs, C.M.G.; Bs, T.J.G.; Wilson, G.D.; Akervall, J.A.; Brenner, C.J.; Walline, H.M.; Carey, T.E. Association of human papillomavirus integration with better patient outcomes in oropharyngeal squamous cell carcinoma. Head Neck 2020, 43, 544–557. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Technique | Advantages | Disadvantages | Ref | |
|---|---|---|---|---|
| In-situ hybridization (ISH) | (Fluorescence) in-situ hybridization ((F) ISH) | Highly sensitive Suitable for morphologically preserved isolated cells, histological tissue sections or chromosome preparations Relatively fast results within one day Relatively expensive with respect to PCR; relatively cheap with respect to sequencing Able to identify number of integration sites per nucleus Able to determine if integration site produces active transcripts (RNAse and DNAse pre-treatment) | Requires prior knowledge about sequence of interest, e.g., in case of human–virus colocalization Requires probe mixture to allow high-risk HPV detection, typing needs additional ISH experiment Cannot determine site of integration if only virus probe is used Cross-hybridization can occur when analyzing highly similar sequences (e.g., HPV6 and HPV11) | [26,27] |
| Polymerase Chain Reaction (PCR) | Quantitative or Real-Time PCR (qPCR, RT-PCR) | Highly specific Extremely sensitive Suitable for fresh frozen material Relatively cheap with respect to sequencing Able to detect viral load based on fluorescence timing | Less suitable for FFPE 1 material Cannot determine site of integration Cannot indicate physical status Cut-off for E2:E6/7-ratio is either less or strong discriminating Integration can occur in different genes: E2 is not always deleted, E1 can also be deleted | [28,29] |
| Detection of Integrated Papillomavirus Sequences PCR (DIPS-PCR) | Suitable for fresh frozen material Relatively cheap with respect to sequencing Able to indicate physical status Able to determine site of integration. | Less suitable for FFPE material Aimed only at fractures in E2 Restriction enzyme is a limiting factor, since the site of integration into the human genome is unknown Digested fragment needs to be at correct length: too long fragments make it difficult to be accurately detected by PCR, too short fragments ensures that integration site remains unknown | [30,31,32,33] | |
| Amplification of Papillomavirus Oncogene Transcripts PCR (APOT-PCR) | Suitable for fresh frozen material Relatively cheap with respect to sequencing Able to indicate physical status Able to determine site of integration if integration occurred in a gene Able to determine if integration site produces active transcripts Highly accurate Highly sensitive, even with large number of samples Able to determine site of integration and viral copy number Able to identify both 5′ and 3′ end breakpoints through hybrid reads Little to no bias due to nature of technique | Less suitable for FFPE material Requires stable RNA of good quality Requires expression of active transcripts Cannot determine site of integration if integration occurred in an intergenic region or an intron due to alternative splicing | [30,31,32,33] | |
| Next-Generation Sequencing (NGS) | RNASeq | Suitable for RNA from blood, fresh-frozen biopsy, FFPE, fine needle aspirates, core needle biopsies and single cells Able to deep profile the transcriptome Able to determine if integration site produces active transcripts Requires lower depth to find 3′ HPV breakpoints with respect to DNA-based NGS due to level of virus transcripts Unbiased approach to view entire RNA population | Cannot find 5′ ends of HPV breakpoints Cannot find HPV integrants that are transcriptionally repressed Can produce false 3′ calls with splice reads Depth may be reduced because of breadth of coverage | [21,26] |
| Whole Genome Sequencing (WGS) | Suitable for genomic DNA (gDNA) from blood and fresh-frozen biopsy. Highly accurate Highly sensitive, even with large number of samples Able to determine site of integration and viral copy number Able to identify both 5′ and 3′ end breakpoints through hybrid reads Little to no bias due to nature of technique | Requires high read depth, deep sequencing and good coverage to find absolute integrant breakpoints Relatively expensive with respect to PCR and (F)ISH Relatively time consuming Cannot determine if HPV integrants are transcriptionally active | [21,34,35] | |
| Whole Exome Sequencing (WES) | Suitable for genomic DNA (gDNA) from blood, fresh-frozen biopsy Highly accurate Extremely sensitive, even with large number of samples Relatively cheap with respect to WGS due to limited target Able to obtain higher depth with respect to WGS due to limited target Able to determine site of integration and viral copy number Able to identify both 5′ and 3′ end breakpoints through hybrid reads Little to no bias due to nature of technique | Less suitable for FFPE material Requires high read depth, deep sequencing and good coverage to find absolute integrant breakpoints. Cannot identify integration sites in non-coding regions. Cannot determine if HPV integrants are transcriptionally active | [21,34,35] | |
| Capture-based assay | Suitable for genomic DNA (gDNA) and/or RNA from blood, fresh-frozen biopsy, DNA and RNA from FFPE, fine needle aspirates, and core needle biopsies. Able to determine site of integration and viral copy number Able to identify both 5′ and 3′ end breakpoints through hybrid reads Increases chance of finding HPV integration sites due to sequence capture Little to no bias due to nature of technique Can be adapted for additional methods, such as chromosome conformation studies | Requires high read depth, deep sequencing and good coverage to find absolute integrant breakpoints Requires individual probes for each HPV type Cannot determine if HPV integrants are transcriptionally active Excludes majority of host sequence | [21,36] | |
| Emerging Techniques | Nanopore Sequencing | Imaging equipment is not required; hence the system can be scaled down to portable level On comparison to other massively parallel sequencers, the device is of much lower cost The captured DNA can be sequenced rapidly Long reads of DNA can be sequenced Able to sequence long repetitive DNA sequences and structural variants | Less suitable for FFPE 1 material Not suitable for single nucleotide variation detection Extremely high molecular weight DNA needed for library preparation The sequencer has the drawback of having high error rate ranging from 5% to 20%, based on the sort of molecules and methods of library preparation | [37] |
| Targeted Locus Amplification | Suitable for purified gDNA from fresh-frozen tissues, fresh tissues and FFPE material Does not require detailed knowledge on locus sequence information Able to determine site of integration and viral copy number Able to identify both 5′ and 3′ end breakpoints through hybrid reads Increases chance of finding HPV integration sites due to sequence capture Relatively long reads of DNA can be sequenced (1 kb in FFPE up to 50–100 kb in fresh cells) surrounding a known/specific sequence/captured target enabling more robust analysis with respect to traditional/standard DNA-based NGS. | Requires high read depth, deep sequencing and good coverage to find absolute integrant breakpoints Complex and extensive integration profile may be challenging to map out completely. Integration sites could be missed in case of a large number of episomal HPV Requires individual probes for each HPV type Cannot determine if HPV integrants are transcriptionally active Excludes majority of host sequence | [38] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balaji, H.; Demers, I.; Wuerdemann, N.; Schrijnder, J.; Kremer, B.; Klussmann, J.P.; Huebbers, C.U.; Speel, E.-J.M. Causes and Consequences of HPV Integration in Head and Neck Squamous Cell Carcinomas: State of the Art. Cancers 2021, 13, 4089. https://doi.org/10.3390/cancers13164089
Balaji H, Demers I, Wuerdemann N, Schrijnder J, Kremer B, Klussmann JP, Huebbers CU, Speel E-JM. Causes and Consequences of HPV Integration in Head and Neck Squamous Cell Carcinomas: State of the Art. Cancers. 2021; 13(16):4089. https://doi.org/10.3390/cancers13164089
Chicago/Turabian StyleBalaji, Harini, Imke Demers, Nora Wuerdemann, Julia Schrijnder, Bernd Kremer, Jens Peter Klussmann, Christian Ulrich Huebbers, and Ernst-Jan Maria Speel. 2021. "Causes and Consequences of HPV Integration in Head and Neck Squamous Cell Carcinomas: State of the Art" Cancers 13, no. 16: 4089. https://doi.org/10.3390/cancers13164089