
Cancer-Testis Antigens in Triple-Negative Breast Cancer: Role and Potential Utility in Clinical Practice


Abstract
:Simple Summary
Abstract
1. Introduction
2. Expression of CTAs in Triple-Negative Breast Cancer
2.1. CTAs Associated with Worse Outcome in TNBC
2.2. CTAs Associated with Improved Outcome in TNBC
2.3. CTAs with Oncogenic Potential
2.4. CTAs with Increased Expression in TNBC but with Unclear Implications
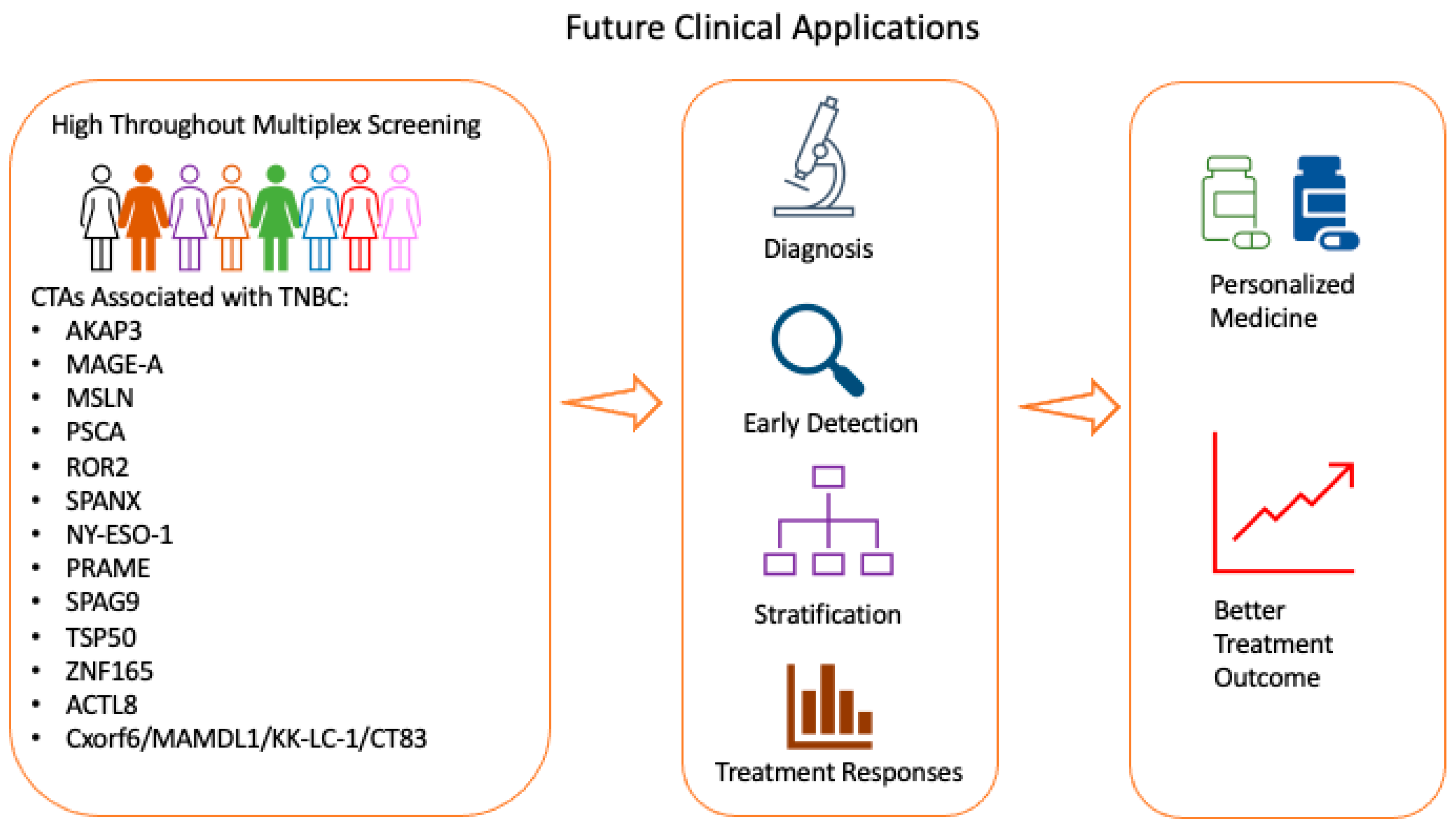
3. Future Potential Application of CTAs Clinically
3.1. Future Potential Application of CTAs in Screening Workflow in Clinical Practice
3.2. Therapeutic Application of CTAs
3.3. Future Research Development
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Thike, A.A.; Cheok, P.Y.; Jara-Lazaro, A.R.; Tan, B.; Tan, P.; Tan, P.H. Triple-negative breast cancer: Clinicopathological characteristics and relationship with basal-like breast cancer. Mod. Pathol. 2010, 23, 123–133. [Google Scholar] [CrossRef]
- Badve, S.; Dabbs, D.J.; Schnitt, S.J.; Baehner, F.L.; Decker, T.; Eusebi, V.; Fox, S.B.; Ichihara, S.; Jacquemier, J.; Lakhani, S.R.; et al. Basal-like and triple-negative breast cancers: A critical review with an emphasis on the implications for pathologists and oncologists. Mod. Pathol. 2011, 24, 157–167. [Google Scholar] [CrossRef]
- Teng, Y.H.; Thike, A.A.; Wong, N.S.; Tan, P.H. Therapeutic targets in triple negative breast cancer—Where are we now? Recent Pat. Anti Cancer Drug Discov. 2011, 6, 196–209. [Google Scholar] [CrossRef]
- Iqbal, J.; Ginsburg, O.; Rochon, P.A.; Sun, P.; Narod, S.A. Differences in breast cancer stage at diagnosis and cancer-specific survival by race and ethnicity in the United States. JAMA 2015, 313, 165–173. [Google Scholar] [CrossRef]
- Foulkes, W.D.; Smith, I.E.; Reis-Filho, J.S. Triple-negative breast cancer. N. Engl. J. Med. 2010, 363, 1938–1948. [Google Scholar] [CrossRef]
- Li, Y.; Li, J.; Wang, Y.; Zhang, Y.; Chu, J.; Sun, C.; Fu, Z.; Huang, Y.; Zhang, H.; Yuan, H.; et al. Roles of cancer/testis antigens (CTAs) in breast cancer. Cancer Lett. 2017, 399, 64–73. [Google Scholar] [CrossRef]
- Schwentner, L.; Wockel, A.; Konig, J.; Janni, W.; Ebner, F.; Blettner, M.; Kreienberg, R.; Van Ewijk, R.; Brenda Study Group. Adherence to treatment guidelines and survival in triple-negative breast cancer: A retrospective multi-center cohort study with 9156 patients. BMC Cancer 2013, 13, 487. [Google Scholar] [CrossRef]
- Ernst, B.; Anderson, K.S. Immunotherapy for the treatment of breast cancer. Curr. Oncol. Rep. 2015, 17, 5. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Thike, A.A.; Ong, C.C.H.; Lim, J.X.; Md Nasir, N.D.; Li, H.; Koh, V.C.Y.; Chen, X.Y.; Yeong, J.P.S.; Sasano, H.; et al. Characteristics, behaviour and role of biomarkers in metastatic triple-negative breast cancer. J. Clin. Pathol. 2020, 73, 147–153. [Google Scholar] [CrossRef]
- Gole, L.; Yeong, J.; Lim, J.C.T.; Ong, K.H.; Han, H.; Thike, A.A.; Poh, Y.C.; Yee, S.; Iqbal, J.; Hong, W.; et al. Quantitative stain-free imaging and digital profiling of collagen structure reveal diverse survival of triple negative breast cancer patients. Breast Cancer Res. 2020, 22, 42. [Google Scholar] [CrossRef]
- Seow, D.Y.B.; Yeong, J.P.S.; Lim, J.X.; Chia, N.; Lim, J.C.T.; Ong, C.C.H.; Tan, P.H.; Iqbal, J. Tertiary lymphoid structures and associated plasma cells play an important role in the biology of triple-negative breast cancers. Breast Cancer Res. Treat. 2020, 180, 369–377. [Google Scholar] [CrossRef]
- Yeong, J.; Lim, J.C.T.; Lee, B.; Li, H.; Ong, C.C.H.; Thike, A.A.; Yeap, W.H.; Yang, Y.; Lim, A.Y.H.; Tay, T.K.Y.; et al. Prognostic value of CD8 + PD-1+ immune infiltrates and PDCD1 gene expression in triple negative breast cancer. J. Immunother. Cancer 2019, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Yeong, J.; Lim, J.C.T.; Lee, B.; Li, H.; Chia, N.; Ong, C.C.H.; Lye, W.K.; Putti, T.C.; Dent, R.; Lim, E.; et al. High Densities of Tumor-Associated Plasma Cells Predict Improved Prognosis in Triple Negative Breast Cancer. Front. Immunol. 2018, 9, 1209. [Google Scholar] [CrossRef]
- Iqbal, J.; Thike, A.A.; Cheok, P.Y.; Tse, G.M.; Tan, P.H. Insulin growth factor receptor-1 expression and loss of PTEN protein predict early recurrence in triple-negative breast cancer. Histopathology 2012, 61, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Thike, A.A.; Yong-Zheng Chong, L.; Cheok, P.Y.; Li, H.H.; Wai-Cheong Yip, G.; Huat Bay, B.; Tse, G.M.; Iqbal, J.; Tan, P.H. Loss of androgen receptor expression predicts early recurrence in triple-negative and basal-like breast cancer. Mod. Pathol. 2014, 27, 352–360. [Google Scholar] [CrossRef]
- Thike, A.A.; Tan, P.H.; Ikeda, M.; Iqbal, J. Increased ID4 expression, accompanied by mutant p53 accumulation and loss of BRCA1/2 proteins in triple-negative breast cancer, adversely affects survival. Histopathology 2016, 68, 702–712. [Google Scholar] [CrossRef]
- Cheung, S.Y.; Boey, Y.J.; Koh, V.C.; Thike, A.A.; Lim, J.C.; Iqbal, J.; Tan, P.H. Role of epithelial-mesenchymal transition markers in triple-negative breast cancer. Breast Cancer Res. Treat. 2015, 152, 489–498. [Google Scholar] [CrossRef]
- Scanlan, M.J.; Gordon, C.M.; Williamson, B.; Lee, S.Y.; Chen, Y.T.; Stockert, E.; Jungbluth, A.; Ritter, G.; Jager, D.; Jager, E.; et al. Identification of cancer/testis genes by database mining and mRNA expression analysis. Int. J. Cancer 2002, 98, 485–492. [Google Scholar] [CrossRef]
- Simpson, A.J.; Caballero, O.L.; Jungbluth, A.; Chen, Y.T.; Old, L.J. Cancer/testis antigens, gametogenesis and cancer. Nat. Rev. Cancer 2005, 5, 615–625. [Google Scholar] [CrossRef]
- Van der Bruggen, P.; Traversari, C.; Chomez, P.; Lurquin, C.; De Plaen, E.; Van den Eynde, B.; Knuth, A.; Boon, T. A gene encoding an antigen recognized by cytolytic T lymphocytes on a human melanoma. Science 1991, 254, 1643–1647. [Google Scholar] [CrossRef]
- Thomas, R.; Al-Khadairi, G.; Roelands, J.; Hendrickx, W.; Dermime, S.; Bedognetti, D.; Decock, J. NY-ESO-1 Based Immunotherapy of Cancer: Current Perspectives. Front. Immunol. 2018, 9, 947. [Google Scholar] [CrossRef]
- D’Angelo, S.P.; Melchiori, L.; Merchant, M.S.; Bernstein, D.; Glod, J.; Kaplan, R.; Grupp, S.; Tap, W.D.; Chagin, K.; Binder, G.K.; et al. Antitumor Activity Associated with Prolonged Persistence of Adoptively Transferred NY-ESO-1 (c259)T Cells in Synovial Sarcoma. Cancer Discov. 2018, 8, 944–957. [Google Scholar] [CrossRef]
- Almeida, L.G.; Sakabe, N.J.; deOliveira, A.R.; Silva, M.C.; Mundstein, A.S.; Cohen, T.; Chen, Y.T.; Chua, R.; Gurung, S.; Gnjatic, S.; et al. CTdatabase: A knowledge-base of high-throughput and curated data on cancer-testis antigens. Nucleic Acids Res. 2009, 37, D816–D819. [Google Scholar] [CrossRef]
- Wang, C.; Gu, Y.; Zhang, K.; Xie, K.; Zhu, M.; Dai, N.; Jiang, Y.; Guo, X.; Liu, M.; Dai, J.; et al. Systematic identification of genes with a cancer-testis expression pattern in 19 cancer types. Nat. Commun. 2016, 7, 10499. [Google Scholar] [CrossRef]
- Da Silva, V.L.; Fonseca, A.F.; Fonseca, M.; da Silva, T.E.; Coelho, A.C.; Kroll, J.E.; de Souza, J.E.S.; Stransky, B.; de Souza, G.A.; de Souza, S.J. Genome-wide identification of cancer/testis genes and their association with prognosis in a pan-cancer analysis. Oncotarget 2017, 8, 92966–92977. [Google Scholar] [CrossRef]
- Rousseaux, S.; Debernardi, A.; Jacquiau, B.; Vitte, A.L.; Vesin, A.; Nagy-Mignotte, H.; Moro-Sibilot, D.; Brichon, P.Y.; Lantuejoul, S.; Hainaut, P.; et al. Ectopic activation of germline and placental genes identifies aggressive metastasis-prone lung cancers. Sci. Transl. Med. 2013, 5, 186ra66. [Google Scholar] [CrossRef]
- Maxfield, K.E.; Taus, P.J.; Corcoran, K.; Wooten, J.; Macion, J.; Zhou, Y.; Borromeo, M.; Kollipara, R.K.; Yan, J.; Xie, Y.; et al. Comprehensive functional characterization of cancer-testis antigens defines obligate participation in multiple hallmarks of cancer. Nat. Commun. 2015, 6, 8840. [Google Scholar] [CrossRef]
- Pineda, C.T.; Ramanathan, S.; Fon Tacer, K.; Weon, J.L.; Potts, M.B.; Ou, Y.H.; White, M.A.; Potts, P.R. Degradation of AMPK by a cancer-specific ubiquitin ligase. Cell 2015, 160, 715–728. [Google Scholar] [CrossRef]
- Gibbs, Z.A.; Whitehurst, A.W. Emerging Contributions of Cancer/Testis Antigens to Neoplastic Behaviors. Trends Cancer 2018, 4, 701–712. [Google Scholar] [CrossRef]
- Esmaeili, R.; Majidzadeh, A.K.; Farahmand, L.; Ghasemi, M.; Salehi, M.; Khoshdel, A.R. AKAP3 correlates with triple negative status and disease free survival in breast cancer. BMC Cancer 2015, 15, 681. [Google Scholar] [CrossRef]
- Sharma, S.; Qian, F.; Keitz, B.; Driscoll, D.; Scanlan, M.J.; Skipper, J.; Rodabaugh, K.; Lele, S.; Old, L.J.; Odunsi, K. A-kinase anchoring protein 3 messenger RNA expression correlates with poor prognosis in epithelial ovarian cancer. Gynecol. Oncol. 2005, 99, 183–188. [Google Scholar] [CrossRef]
- Hamai, A.; Memeo, L.; Colarossi, C.; Canzonieri, V.; Perin, T.; Ayyoub, M.; Valmori, D. Expression of MAGE-A antigens is frequent in triple-negative breast cancers but does not correlate with that of basal-like markers. Ann. Oncol. 2011, 22, 986–987. [Google Scholar] [CrossRef]
- Curigliano, G.; Viale, G.; Ghioni, M.; Jungbluth, A.A.; Bagnardi, V.; Spagnoli, G.C.; Neville, A.M.; Nole, F.; Rotmensz, N.; Goldhirsch, A. Cancer-testis antigen expression in triple-negative breast cancer. Ann. Oncol. 2011, 22, 98–103. [Google Scholar] [CrossRef]
- Grigoriadis, A.; Caballero, O.L.; Hoek, K.S.; da Silva, L.; Chen, Y.T.; Shin, S.J.; Jungbluth, A.A.; Miller, L.D.; Clouston, D.; Cebon, J.; et al. CT-X antigen expression in human breast cancer. Proc. Natl. Acad. Sci. USA 2009, 106, 13493–13498. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Ross, D.S.; Chiu, R.; Zhou, X.K.; Chen, Y.Y.; Lee, P.; Hoda, S.A.; Simpson, A.J.; Old, L.J.; Caballero, O.; et al. Multiple cancer/testis antigens are preferentially expressed in hormone-receptor negative and high-grade breast cancers. PLoS ONE 2011, 6, e17876. [Google Scholar] [CrossRef]
- Karn, T.; Pusztai, L.; Ruckhaberle, E.; Liedtke, C.; Muller, V.; Schmidt, M.; Metzler, D.; Wang, J.; Coombes, K.R.; Gatje, R.; et al. Melanoma antigen family A identified by the bimodality index defines a subset of triple negative breast cancers as candidates for immune response augmentation. Eur. J. Cancer 2012, 48, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Ayyoub, M.; Scarlata, C.M.; Hamai, A.; Pignon, P.; Valmori, D. Expression of MAGE-A3/6 in primary breast cancer is associated with hormone receptor negative status, high histologic grade, and poor survival. J. Immunother. 2014, 37, 73–76. [Google Scholar] [CrossRef]
- Yang, F.; Zhou, X.; Miao, X.; Zhang, T.; Hang, X.; Tie, R.; Liu, N.; Tian, F.; Wang, F.; Yuan, J. MAGEC2, an epithelial-mesenchymal transition inducer, is associated with breast cancer metastasis. Breast Cancer Res. Treat. 2014, 145, 23–32. [Google Scholar] [CrossRef]
- Badovinac Crnjevic, T.; Spagnoli, G.; Juretic, A.; Jakic-Razumovic, J.; Podolski, P.; Saric, N. High expression of MAGE-A10 cancer-testis antigen in triple-negative breast cancer. Med. Oncol. 2012, 29, 1586–1591. [Google Scholar] [CrossRef]
- Raghavendra, A.; Kalita-de Croft, P.; Vargas, A.C.; Smart, C.E.; Simpson, P.T.; Saunus, J.M.; Lakhani, S.R. Expression of MAGE-A and NY-ESO-1 cancer/testis antigens is enriched in triple-negative invasive breast cancers. Histopathology 2018, 73, 68–80. [Google Scholar] [CrossRef]
- Zhao, Q.; Xu, W.T.; Shalieer, T. Pilot Study on MAGE-C2 as a Potential Biomarker for Triple-Negative Breast Cancer. Dis. Markers 2016, 2016, 2325987. [Google Scholar] [CrossRef]
- Wang, H.; Sang, M.; Geng, C.; Liu, F.; Gu, L.; Shan, B. MAGE-A is frequently expressed in triple negative breast cancer and associated with epithelial-mesenchymal transition. Neoplasma 2016, 63, 44–56. [Google Scholar] [CrossRef]
- Tessari, A.; Pilla, L.; Silvia, D.; Duca, M.; Paolini, B.; Carcangiu, M.L.; Mariani, L.; de Braud, F.G.; Cresta, S. Expression of NY-ESO-1, MAGE-A3, PRAME and WT1 in different subgroups of breast cancer: An indication to immunotherapy? Breast 2018, 42, 68–73. [Google Scholar] [CrossRef]
- Mrklic, I.; Spagnoli, G.C.; Juretic, A.; Pogorelic, Z.; Tomic, S. Co-expression of cancer testis antigens and topoisomerase 2-alpha in triple negative breast carcinomas. Acta Histochem. 2014, 116, 740–746. [Google Scholar] [CrossRef]
- Cabezon, T.; Gromova, I.; Gromov, P.; Serizawa, R.; Timmermans Wielenga, V.; Kroman, N.; Celis, J.E.; Moreira, J.M. Proteomic profiling of triple-negative breast carcinomas in combination with a three-tier orthogonal technology approach identifies Mage-A4 as potential therapeutic target in estrogen receptor negative breast cancer. Mol. Cell. Proteom. 2013, 12, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Tchou, J.; Wang, L.C.; Selven, B.; Zhang, H.; Conejo-Garcia, J.; Borghaei, H.; Kalos, M.; Vondeheide, R.H.; Albelda, S.M.; June, C.H.; et al. Mesothelin, a novel immunotherapy target for triple negative breast cancer. Breast Cancer Res. Treat. 2012, 133, 799–804. [Google Scholar] [CrossRef]
- Tozbikian, G.; Brogi, E.; Kadota, K.; Catalano, J.; Akram, M.; Patil, S.; Ho, A.Y.; Reis-Filho, J.S.; Weigelt, B.; Norton, L.; et al. Mesothelin expression in triple negative breast carcinomas correlates significantly with basal-like phenotype, distant metastases and decreased survival. PLoS ONE 2014, 9, e114900. [Google Scholar] [CrossRef]
- Parinyanitikul, N.; Blumenschein, G.R.; Wu, Y.; Lei, X.; Chavez-Macgregor, M.; Smart, M.; Gonzalez-Angulo, A.M. Mesothelin expression and survival outcomes in triple receptor negative breast cancer. Clin. Breast Cancer 2013, 13, 378–384. [Google Scholar] [CrossRef]
- Link, T.; Kuithan, F.; Ehninger, A.; Kuhlmann, J.D.; Kramer, M.; Werner, A.; Gatzweiler, A.; Richter, B.; Ehninger, G.; Baretton, G.; et al. Exploratory investigation of PSCA-protein expression in primary breast cancer patients reveals a link to HER2/neu overexpression. Oncotarget 2017, 8, 54592–54603. [Google Scholar] [CrossRef]
- Meng, F.; Liu, B.; Xie, G.; Song, Y.; Zheng, X.; Qian, X.; Li, S.; Jia, H.; Zhang, X.; Zhang, L.; et al. Amplification and overexpression of PSCA at 8q24 in invasive micropapillary carcinoma of breast. Breast Cancer Res. Treat. 2017, 166, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Henry, C.; Quadir, A.; Hawkins, N.J.; Jary, E.; Llamosas, E.; Kumar, D.; Daniels, B.; Ward, R.L.; Ford, C.E. Expression of the novel Wnt receptor ROR2 is increased in breast cancer and may regulate both beta-catenin dependent and independent Wnt signalling. J. Cancer Res. Clin. Oncol. 2015, 141, 243–254. [Google Scholar] [CrossRef]
- Kannan, A.; Philley, J.V.; Hertweck, K.L.; Ndetan, H.; Singh, K.P.; Sivakumar, S.; Wells, R.B.; Vadlamudi, R.K.; Dasgupta, S. Cancer Testis Antigen Promotes Triple Negative Breast Cancer Metastasis and is Traceable in the Circulating Extracellular Vesicles. Sci. Rep. 2019, 9, 11632. [Google Scholar] [CrossRef]
- Maine, E.A.; Westcott, J.M.; Prechtl, A.M.; Dang, T.T.; Whitehurst, A.W.; Pearson, G.W. The cancer-testis antigens SPANX-A/C/D and CTAG2 promote breast cancer invasion. Oncotarget 2016, 7, 14708–14726. [Google Scholar] [CrossRef]
- Ademuyiwa, F.O.; Bshara, W.; Attwood, K.; Morrison, C.; Edge, S.B.; Karpf, A.R.; James, S.A.; Ambrosone, C.B.; O’Connor, T.L.; Levine, E.G.; et al. NY-ESO-1 cancer testis antigen demonstrates high immunogenicity in triple negative breast cancer. PLoS ONE 2012, 7, e38783. [Google Scholar] [CrossRef]
- Bandic, D.; Juretic, A.; Sarcevic, B.; Separovic, V.; Kujundzic-Tiljak, M.; Hudolin, T.; Spagnoli, G.C.; Covic, D.; Samija, M. Expression and possible prognostic role of MAGE-A4, NY-ESO-1, and HER-2 antigens in women with relapsing invasive ductal breast cancer: Retrospective immunohistochemical study. Croat. Med. J. 2006, 47, 32–41. [Google Scholar]
- Lee, H.J.; Kim, J.Y.; Song, I.H.; Park, I.A.; Yu, J.H.; Gong, G. Expression of NY-ESO-1 in Triple-Negative Breast Cancer Is Associated with Tumor-Infiltrating Lymphocytes and a Good Prognosis. Oncology 2015, 89, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Curigliano, G.; Bagnardi, V.; Ghioni, M.; Louahed, J.; Brichard, V.; Lehmann, F.F.; Marra, A.; Trapani, D.; Criscitiello, C.; Viale, G. Expression of tumor-associated antigens in breast cancer subtypes. Breast 2020, 49, 202–209. [Google Scholar] [CrossRef]
- Epping, M.T.; Hart, A.A.; Glas, A.M.; Krijgsman, O.; Bernards, R. PRAME expression and clinical outcome of breast cancer. Br. J. Cancer 2008, 99, 398–403. [Google Scholar] [CrossRef]
- Yao, J.; Caballero, O.L.; Yung, W.K.; Weinstein, J.N.; Riggins, G.J.; Strausberg, R.L.; Zhao, Q. Tumor subtype-specific cancer-testis antigens as potential biomarkers and immunotherapeutic targets for cancers. Cancer Immunol. Res. 2014, 2, 371–379. [Google Scholar] [CrossRef]
- Al-Khadairi, G.; Naik, A.; Thomas, R.; Al-Sulaiti, B.; Rizly, S.; Decock, J. PRAME promotes epithelial-to-mesenchymal transition in triple negative breast cancer. J. Transl. Med. 2019, 17, 9. [Google Scholar] [CrossRef]
- Kanojia, D.; Garg, M.; Gupta, S.; Gupta, A.; Suri, A. Sperm-associated antigen 9, a novel biomarker for early detection of breast cancer. Cancer Epidemiol. Biomark. Prev. 2009, 18, 630–639. [Google Scholar] [CrossRef]
- Sinha, A.; Agarwal, S.; Parashar, D.; Verma, A.; Saini, S.; Jagadish, N.; Ansari, A.S.; Lohiya, N.K.; Suri, A. Down regulation of SPAG9 reduces growth and invasive potential of triple-negative breast cancer cells: Possible implications in targeted therapy. J. Exp. Clin. Cancer Res. 2013, 32, 69. [Google Scholar] [CrossRef]
- Song, Z.B.; Ni, J.S.; Wu, P.; Bao, Y.L.; Liu, T.; Li, M.; Fan, C.; Zhang, W.J.; Sun, L.G.; Huang, Y.X.; et al. Testes-specific protease 50 promotes cell invasion and metastasis by increasing NF-kappaB-dependent matrix metalloproteinase-9 expression. Cell Death Dis. 2015, 6, e1703. [Google Scholar] [CrossRef]
- Wang, M.; Bao, Y.L.; Wu, Y.; Yu, C.L.; Meng, X.Y.; Huang, Y.X.; Sun, Y.; Zheng, L.H.; Li, Y.X. Basic FGF downregulates TSP50 expression via the ERK/Sp1 pathway. J. Cell. Biochem. 2010, 111, 75–81. [Google Scholar] [CrossRef]
- Gibbs, Z.A.; Reza, L.C.; Cheng, C.C.; Westcott, J.M.; McGlynn, K.; Whitehurst, A.W. The testis protein ZNF165 is a SMAD3 cofactor that coordinates oncogenic TGFbeta signaling in triple-negative breast cancer. eLife 2020, 9. [Google Scholar] [CrossRef]
- Kaufmann, J.; Wentzensen, N.; Brinker, T.J.; Grabe, N. Large-scale in-silico identification of a tumor-specific antigen pool for targeted immunotherapy in triple-negative breast cancer. Oncotarget 2019, 10, 2515–2529. [Google Scholar] [CrossRef]
- Paret, C.; Simon, P.; Vormbrock, K.; Bender, C.; Kolsch, A.; Breitkreuz, A.; Yildiz, O.; Omokoko, T.; Hubich-Rau, S.; Hartmann, C.; et al. CXorf61 is a target for T cell based immunotherapy of triple-negative breast cancer. Oncotarget 2015, 6, 25356–25367. [Google Scholar] [CrossRef]
- Kondo, Y.; Fukuyama, T.; Yamamura, R.; Futawatari, N.; Ichiki, Y.; Tanaka, Y.; Nishi, Y.; Takahashi, Y.; Yamazaki, H.; Kobayashi, N.; et al. Detection of KK-LC-1 Protein, a Cancer/Testis Antigen, in Patients with Breast Cancer. Anticancer. Res. 2018, 38, 5923–5928. [Google Scholar] [CrossRef]
- Mirandola, L.; Pedretti, E.; Figueroa, J.A.; Chiaramonte, R.; Colombo, M.; Chapman, C.; Grizzi, F.; Patrinicola, F.; Kast, W.M.; Nguyen, D.D.; et al. Cancer testis antigen Sperm Protein 17 as a new target for triple negative breast cancer immunotherapy. Oncotarget 2017, 8, 74378–74390. [Google Scholar] [CrossRef]
- Newman, J.A.; Cooper, C.D.; Roos, A.K.; Aitkenhead, H.; Oppermann, U.C.; Cho, H.J.; Osman, R.; Gileadi, O. Structures of Two Melanoma-Associated Antigens Suggest Allosteric Regulation of Effector Binding. PLoS ONE 2016, 11, e0148762. [Google Scholar] [CrossRef]
- Poojary, M.; Jishnu, P.V.; Kabekkodu, S.P. Prognostic Value of Melanoma-Associated Antigen-A (MAGE-A) Gene Expression in Various Human Cancers: A Systematic Review and Meta-analysis of 7428 Patients and 44 Studies. Mol. Diagn. Ther. 2020. [Google Scholar] [CrossRef]
- Weon, J.L.; Potts, P.R. The MAGE protein family and cancer. Curr. Opin. Cell Biol. 2015, 37, 1–8. [Google Scholar] [CrossRef]
- Zajac, P.; Schultz-Thater, E.; Tornillo, L.; Sadowski, C.; Trella, E.; Mengus, C.; Iezzi, G.; Spagnoli, G.C. MAGE-A Antigens and Cancer Immunotherapy. Front. Med. (Lausanne) 2017, 4, 18. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Li, P. Mesothelin as a biomarker for targeted therapy. Biomark. Res. 2019, 7, 18. [Google Scholar] [CrossRef]
- Morello, A.; Sadelain, M.; Adusumilli, P.S. Mesothelin-Targeted CARs: Driving T Cells to Solid Tumors. Cancer Discov. 2016, 6, 133–146. [Google Scholar] [CrossRef]
- Masiakowski, P.; Carroll, R.D. A novel family of cell surface receptors with tyrosine kinase-like domain. J. Biol. Chem. 1992, 267, 26181–26190. [Google Scholar] [CrossRef]
- Westbrook, V.A.; Diekman, A.B.; Klotz, K.L.; Khole, V.V.; von Kap-Herr, C.; Golden, W.L.; Eddy, R.L.; Shows, T.B.; Stoler, M.H.; Lee, C.Y.; et al. Spermatid-specific expression of the novel X-linked gene product SPAN-X localized to the nucleus of human spermatozoa. Biol. Reprod. 2000, 63, 469–481. [Google Scholar] [CrossRef]
- Carnegie, G.K.; Means, C.K.; Scott, J.D. A-kinase anchoring proteins: From protein complexes to physiology and disease. IUBMB Life 2009, 61, 394–406. [Google Scholar] [CrossRef]
- Sugita, Y.; Wada, H.; Fujita, S.; Nakata, T.; Sato, S.; Noguchi, Y.; Jungbluth, A.A.; Yamaguchi, M.; Chen, Y.T.; Stockert, E.; et al. NY-ESO-1 expression and immunogenicity in malignant and benign breast tumors. Cancer Res. 2004, 64, 2199–2204. [Google Scholar] [CrossRef]
- Gjerstorff, M.F.; Andersen, M.H.; Ditzel, H.J. Oncogenic cancer/testis antigens: Prime candidates for immunotherapy. Oncotarget 2015, 6, 15772–15787. [Google Scholar] [CrossRef]
- Doyle, J.M.; Gao, J.; Wang, J.; Yang, M.; Potts, P.R. MAGE-RING protein complexes comprise a family of E3 ubiquitin ligases. Mol. Cell 2010, 39, 963–974. [Google Scholar] [CrossRef]
- Xu, Y.; Zou, R.; Wang, J.; Wang, Z.W.; Zhu, X. The role of the cancer testis antigen PRAME in tumorigenesis and immunotherapy in human cancer. Cell Prolif. 2020, 53, e12770. [Google Scholar] [CrossRef]
- Fukami, M.; Wada, Y.; Miyabayashi, K.; Nishino, I.; Hasegawa, T.; Nordenskjold, A.; Camerino, G.; Kretz, C.; Buj-Bello, A.; Laporte, J.; et al. CXorf6 is a causative gene for hypospadias. Nat. Genet. 2006, 38, 1369–1371. [Google Scholar] [CrossRef]
- Ogata, T.; Laporte, J.; Fukami, M. MAMLD1 (CXorf6): A new gene involved in hypospadias. Horm. Res. 2009, 71, 245–252. [Google Scholar] [CrossRef]
- Grizzi, F.; Gaetani, P.; Franceschini, B.; Di Ieva, A.; Colombo, P.; Ceva-Grimaldi, G.; Bollati, A.; Frezza, E.E.; Cobos, E.; Rodriguez y Baena, R.; et al. Sperm protein 17 is expressed in human nervous system tumours. BMC Cancer 2006, 6, 23. [Google Scholar] [CrossRef]
- Gupta, G.; Sharma, R.; Chattopadhyay, T.K.; Gupta, S.D.; Ralhan, R. Clinical significance of sperm protein 17 expression and immunogenicity in esophageal cancer. Int. J. Cancer 2007, 120, 1739–1747. [Google Scholar] [CrossRef]
- Lim, S.H.; Wang, Z.; Chiriva-Internati, M.; Xue, Y. Sperm protein 17 is a novel cancer-testis antigen in multiple myeloma. Blood 2001, 97, 1508–1510. [Google Scholar] [CrossRef] [PubMed]
- Nakazato, T.; Kanuma, T.; Tamura, T.; Faried, L.S.; Aoki, H.; Minegishi, T. Sperm protein 17 influences the tissue-specific malignancy of clear cell adenocarcinoma in human epithelial ovarian cancer. Int. J. Gynecol. Cancer 2007, 17, 426–432. [Google Scholar] [CrossRef]
- Bumm, K.; Grizzi, F.; Franceschini, B.; Koch, M.; Iro, H.; Wurm, J.; Ceva-Grimaldi, G.; Dimmler, A.; Cobos, E.; Dioguardi, N.; et al. Sperm protein 17 expression defines 2 subsets of primary esthesioneuroblastoma. Hum. Pathol. 2005, 36, 1289–1293. [Google Scholar] [CrossRef]
- Chiriva-Internati, M. Sperm protein 17: Clinical relevance of a cancer/testis antigen, from contraception to cancer immunotherapy, and beyond. Int. Rev. Immunol. 2011, 30, 138–149. [Google Scholar] [CrossRef]
- Lacy, H.M.; Sanderson, R.D. Sperm protein 17 is expressed on normal and malignant lymphocytes and promotes heparan sulfate-mediated cell-cell adhesion. Blood 2001, 98, 2160–2165. [Google Scholar] [CrossRef]
- Li, F.Q.; Han, Y.L.; Liu, Q.; Wu, B.; Huang, W.B.; Zeng, S.Y. Overexpression of human sperm protein 17 increases migration and decreases the chemosensitivity of human epithelial ovarian cancer cells. BMC Cancer 2009, 9, 323. [Google Scholar] [CrossRef]
- Call, K.M.; Glaser, T.; Ito, C.Y.; Buckler, A.J.; Pelletier, J.; Haber, D.A.; Rose, E.A.; Kral, A.; Yeger, H.; Lewis, W.H.; et al. Isolation and characterization of a zinc finger polypeptide gene at the human chromosome 11 Wilms’ tumor locus. Cell 1990, 60, 509–520. [Google Scholar] [CrossRef]
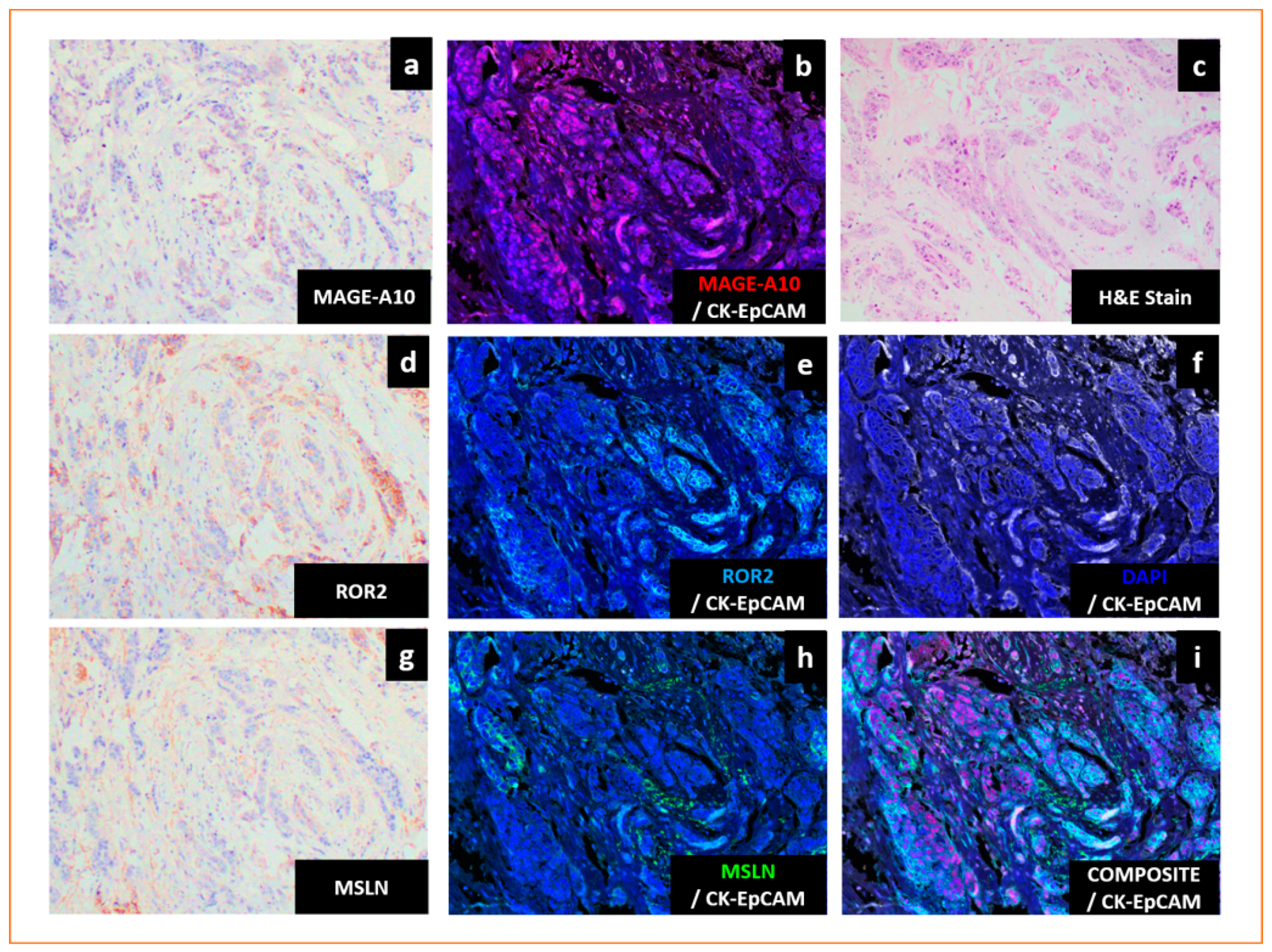
- Tien, T.Z.; Lee, J.; Lim, J.C.T.; Chen, X.Y.; Thike, A.A.; Tan, P.H.; Yeong, J.P.S. Delineating the breast cancer immune microenvironment in the era of multiplex immunohistochemistry/immunofluorescence. Histopathology 2021, 79, 139–159. [Google Scholar] [CrossRef]
- Tan, W.C.C.; Nerurkar, S.N.; Cai, H.Y.; Ng, H.H.M.; Wu, D.; Wee, Y.T.F.; Lim, J.C.T.; Yeong, J.; Lim, T.K.H. Overview of multiplex immunohistochemistry/immunofluorescence techniques in the era of cancer immunotherapy. Cancer Commun. (Lond.) 2020, 40, 135–153. [Google Scholar] [CrossRef]
- Dixon, A.R.; Bathany, C.; Tsuei, M.; White, J.; Barald, K.F.; Takayama, S. Recent developments in multiplexing techniques for immunohistochemistry. Expert Rev. Mol. Diagn. 2015, 15, 1171–1186. [Google Scholar] [CrossRef]
- Lu, S.; Stein, J.E.; Rimm, D.L.; Wang, D.W.; Bell, J.M.; Johnson, D.B.; Sosman, J.A.; Schalper, K.A.; Anders, R.A.; Wang, H.; et al. Comparison of Biomarker Modalities for Predicting Response to PD-1/PD-L1 Checkpoint Blockade: A Systematic Review and Meta-analysis. JAMA Oncol. 2019, 5, 1195–1204. [Google Scholar] [CrossRef]
- Yeong, J.; Tan, T.; Chow, Z.L.; Cheng, Q.; Lee, B.; Seet, A.; Lim, J.X.; Lim, J.C.T.; Ong, C.C.H.; Thike, A.A.; et al. Multiplex immunohistochemistry/immunofluorescence (mIHC/IF) for PD-L1 testing in triple-negative breast cancer: A translational assay compared with conventional IHC. J. Clin. Pathol. 2020, 73, 557–562. [Google Scholar] [CrossRef]
- Taube, J.M.; Roman, K.; Engle, E.L.; Wang, C.; Ballesteros-Merino, C.; Jensen, S.M.; McGuire, J.; Jiang, M.; Coltharp, C.; Remeniuk, B.; et al. Multi-institutional TSA-amplified Multiplexed Immunofluorescence Reproducibility Evaluation (MITRE) Study. J. Immunother. Cancer 2021, 9. [Google Scholar] [CrossRef]
- Cho, C.H.; Cho, M.; Park, J.K. Biomarker barcodes: Multiplexed microfluidic immunohistochemistry enables high-throughput analysis of tissue microarray. Lab Chip 2021. [Google Scholar] [CrossRef]
- Marotti, J.D.; de Abreu, F.B.; Wells, W.A.; Tsongalis, G.J. Triple-Negative Breast Cancer: Next-Generation Sequencing for Target Identification. Am. J. Pathol. 2017, 187, 2133–2138. [Google Scholar] [CrossRef]
- Dillon, J.L.; Mockus, S.M.; Ananda, G.; Spotlow, V.; Wells, W.A.; Tsongalis, G.J.; Marotti, J.D. Somatic gene mutation analysis of triple negative breast cancers. Breast 2016, 29, 202–207. [Google Scholar] [CrossRef]
- Staaf, J.; Glodzik, D.; Bosch, A.; Vallon-Christersson, J.; Reutersward, C.; Hakkinen, J.; Degasperi, A.; Amarante, T.D.; Saal, L.H.; Hegardt, C.; et al. Whole-genome sequencing of triple-negative breast cancers in a population-based clinical study. Nat. Med. 2019, 25, 1526–1533. [Google Scholar] [CrossRef]
- Pinto, C.; Biffoni, M.; Popoli, P.; Marchetti, A.; Marchetti, P.; Martini, N.; Normanno, N. Molecular Tests and Target Therapies in Oncology: Recommendations from the Italian workshop. Future Oncol. 2021. [Google Scholar] [CrossRef]
- Li, X.F.; Ren, P.; Shen, W.Z.; Jin, X.; Zhang, J. The expression, modulation and use of cancer-testis antigens as potential biomarkers for cancer immunotherapy. Am. J. Transl. Res. 2020, 12, 7002–7019. [Google Scholar]
- Somaiah, N.; Block, M.S.; Kim, J.W.; Shapiro, G.I.; Do, K.T.; Hwu, P.; Eder, J.P.; Jones, R.L.; Lu, H.; Ter Meulen, J.H.; et al. First-in-Class, First-in-Human Study Evaluating LV305, a Dendritic-Cell Tropic Lentiviral Vector, in Sarcoma and Other Solid Tumors Expressing NY-ESO-1. Clin. Cancer Res. 2019, 25, 5808–5817. [Google Scholar] [CrossRef]
- Mahipal, A.; Ejadi, S.; Gnjatic, S.; Kim-Schulze, S.; Lu, H.; Ter Meulen, J.H.; Kenney, R.; Odunsi, K. First-in-human phase 1 dose-escalating trial of G305 in patients with advanced solid tumors expressing NY-ESO-1. Cancer Immunol. Immunother. 2019, 68, 1211–1222. [Google Scholar] [CrossRef]
- Pollack, S.M. The potential of the CMB305 vaccine regimen to target NY-ESO-1 and improve outcomes for synovial sarcoma and myxoid/round cell liposarcoma patients. Expert Rev. Vaccines 2018, 17, 107–114. [Google Scholar] [CrossRef]
- Baumgaertner, P.; Costa Nunes, C.; Cachot, A.; Maby-El Hajjami, H.; Cagnon, L.; Braun, M.; Derre, L.; Rivals, J.P.; Rimoldi, D.; Gnjatic, S.; et al. Vaccination of stage III/IV melanoma patients with long NY-ESO-1 peptide and CpG-B elicits robust CD8(+) and CD4(+) T-cell responses with multiple specificities including a novel DR7-restricted epitope. Oncoimmunology 2016, 5, e1216290. [Google Scholar] [CrossRef]
- Lu, Y.C.; Parker, L.L.; Lu, T.; Zheng, Z.; Toomey, M.A.; White, D.E.; Yao, X.; Li, Y.F.; Robbins, P.F.; Feldman, S.A.; et al. Treatment of Patients with Metastatic Cancer Using a Major Histocompatibility Complex Class II-Restricted T-Cell Receptor Targeting the Cancer Germline Antigen MAGE-A3. J. Clin. Oncol. 2017, 35, 3322–3329. [Google Scholar] [CrossRef]
- Hong, D.S.; Tine, B.A.V.; Olszanski, A.J.; Johnson, M.L.; Liebner, D.A.; Trivedi, T.; Lin, Q.; Elefant, E.; Dryer-Minnerly, R.; Navenot, J.-M.; et al. Phase I dose escalation and expansion trial to assess the safety and efficacy of ADP-A2M4 SPEAR T cells in advanced solid tumors. J. Clin. Oncol. 2020, 38, 102. [Google Scholar] [CrossRef]
- Jin, S.; Cao, S.; Li, J.; Meng, Q.; Wang, C.; Yao, L.; Lang, Y.; Cao, J.; Shen, J.; Pan, B.; et al. Cancer/testis antigens (CTAs) expression in resected lung cancer. Onco Targets Ther. 2018, 11, 4491–4499. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| CTAs | Cellular Function | Institute | Cohort | Prevalence of CTAs in TNBC | Type of Assay | Antibodies | Role in TNBC | Ref. |
|---|---|---|---|---|---|---|---|---|
| CTAs associated with worse prognosis in TNBC | ||||||||
| A-kinase anchoring proteins (AKAP3) | Sperm function | Breast Cancer Research Centre (Tehran, Iran) [30] | Asian | 20% (n = 25) | Real-Time Polymerase Chain Reaction (RT-PCR) | Loss of expression in TNBC. Breast cancer patients who were positive for AKAP3 had better 5-year disease-free survival. | [30,31] | |
| Melanoma antigen gene (MAGE) | Not known. May promote tumourigenesis and metastasis. | Italian National Cancer Institute [32] | Caucasian | MAGE-A: 23% (n = 44) | IHC | MAGE-A Antibody (6C1) | Frequently overexpressed in TNBC. Higher expression of MAGE-A was reported to define a very aggressive subtype of TNBC and correlated with poor prognosis of patients. MAGE-A3, -A6 and -C2 expression in breast cancers was significantly associated with negative ER or negative PR status, higher-grade tumours and correlated with worse outcomes. MAGE-A10 expression was associated with ER-negative, PR-negative and HER2-negative status. | [32,33,34,35,36,37,38,39,40,41,42,43] |
| Royal Brisbane Women’s Hospital [40] | Caucasian | MAGE-A: 47% (n = 65) | IHC | MAGE-A Antibody (6C1), Santa Cruz Biotechnology(USA) | ||||
| Affiliated Tumour Hospital of Xinjiang Medical University [41] | Asian | MAGE-C: 38.2% (n = 110) | IHC | Rabbit polyclonal MAGE-C2 Antibody, Sigma-Aldrich (USA) | ||||
| Centre of Breast Cancer of The Fourth Hospital of Hebei Medical University (Shijiazhuang Hebei) [42] | Asian | MAGE-A: 76.5% (n = 17) | IHC | MAGE-A Antibody (6C1), Santa Cruz Biotechnology(USA) | ||||
| University Hospital Center Zagreb [39] | Caucasian | MAGE-A: 85.7% (n = 49) | IHC | 3DA3 Monoclonal Antibody | ||||
| Split University Hospital Centre, Croatia [44] | Caucasian | MAGE-A1 Specific: 69.2% (n = 81) | IHC | Monoclonal Antibody 77B | ||||
| Multi-MAGE: 58% (n = 81) | IHC | Monoclonal Antibody 57B | ||||||
| MAGE-A10: 16% (n = 81) | IHC | Monoclonal Antibody 3GA11 | ||||||
| European Institute of Oncology (Milan, Italy) [33] | Caucasian | MAGE-A: 32% (n = 50) | IHC | Antibody cocktail of monoclonal antibodies 6C1, MA454, M3H67 and 57B | ||||
| Copenhagen University Hospital [45] | Caucasian | MAGE-A: 33% (n = 78) | IHC | Rabbit polyclonal anti-peptide antibody EP101638 (rab Ab 1982) raised against Mage-4, Eurogentec (Belgium) | ||||
| National Cancer Institute (Milan, Italy) [43] | Caucasian | MAGE-A: 85.7–93% (n = 21) | IHC | MAGE-A3 (Clone 60054-1-Ig) Monoclonal Antibody, Proteinthec (USA) | ||||
| Mesothelin (MSLN) | GPI-anchored membrane protein | Perelman School of Medicine, University of Pennsylvania [46] | Caucasian | 67% (n = 99) | IHC | Mesothelin Monoclonal Antibody (clone 5B2), Thermo Scientific (USA) | MSLN is significantly expressed in TNBC compared to non-TNBC and is an independent prognostic marker associated with distant metastasis and worse survival. | [46,47,48] |
| University of Texas MD Anderson Cancer Center [48] | Caucasian | 34% (n = 109) | IHC | Mesothelin Monoclonal Antibody (clone 5B2), Novocastra (USA) | ||||
| Prostate stem cell antigen (PSCA) | GPI-anchored membrane protein | University Hospital of Dresden, Germany [49] | Caucasian | 17% (n = 90) | IHC | PSCA antibody MB1 | Distribution of PSCA expression among TNBC was comparable to the total population. Patients with PSCA-positive invasive micropapillary carcinoma (IMPC) of the breast had decreased disease-free survival. | [49,50] |
| Receptor tyrosine kinase-like orphan receptor 2 (ROR2) | Tyrosine kinase receptor family | University of New South Wales [51] | Caucasian | 87% (n = 295, breast cancer including triple- negative) | IHC | Human ROR2 polyclonal antibody, Sigma-Aldrich (Australia) | Breast cancer patients including TNBC expressing ROR2 had significantly worse prognoses with shorter overall survival compared to those lacking ROR2. | [51] |
| Sperm protein associated with the nucleus X-linked (SPANX) | Sperm function | University of Texas Health Science Center [52] | Caucasian | 73% (n = 15) | IHC | SPANXB1 (#H00728695), Abnova (Taiwan) | SPANXB1 was frequently overexpressed in human primary and metastatic TNBC. In ER-negative patients, elevated SPANX-A/C/D was correlated with shorter distant metastasis-free survival time. | [52,53] |
| CTAs associated with better prognosis in TNBC | ||||||||
| New York oesophageal squamous cell carcinoma-1 (NY-ESO-1) | Unknown; might be involved in cell cycle progression and growth | New York Presbyterian Hospital-Weill Cornell Medical Center and UCSF Medical Center [35] | Caucasian | 19.2% (n = 50) | IHC | NY-ESO-1 Monoclonal Antibody(E978) produced in author’s laboratory | Higher expression of NY-ESO-1 was detected in TNBC. NY-ESO-1 expression was correlated with tumour-infiltrating lymphocytes and associated with good prognosis. | [33,35,39,40,43,54,55,56,57] |
| University Hospital Center Zagreb [39] | Caucasian | 10% (n = 50) | IHC | NY-ESO-1 Monoclonal Antibody (B9.8.1.1) | ||||
| Roswell Park Cancer Institute [54] | Caucasian | 16% (n = 168) | IHC | NY-ESO-1 Mouse Monoclonal, Zymed/Invitrogen (USA) | ||||
| Asan Medical Centre, Korea [56] | Asian | 9.3% (n = 172) | IHC | NY-ESO-1 Monoclonal Antibody (E978), Invitrogen (USA) | ||||
| Royal Brisbane Women’s hospital [40] | Caucasian | ~20% (n = 65) | IHC | NY-ESO-1 Antibody (E978), Santa Cruz Biotechnology(USA) | ||||
| National Cancer Institute (Milan, Italy) [43] | Caucasian | 28.6% (n = 21) | IHC | NY-ESO-1 Monoclonal Antibody (E978), Invitrogen (USA) | ||||
| European Institute of Oncology (Milan, Italy) [57] | Caucasian | 16% (n = 50) | IHC | NY-ESO-1 Monoclonal antibody (E978) provided by Ludwig Institute for Cancer Research | ||||
| CTAs with oncogenic potential | ||||||||
| Melanoma antigen gene (MAGE) | Not known. May promote tumourigenesis and metastasis. | See Above | Promote tumourigenesis and metastasis via various mechanisms such as acting as master regulator of E3 RING ubiquitin ligase, inhibiting p53 tumour suppressor or by enhancing cell motility. | [33,34,35,36,37,38,39] | ||||
| New York oesophageal squamous cell carcinoma-1 (NY-ESO-1) | Unknown; might be involved in cell cycle progression and growth | See Above | Might be involved in cellular proliferation and growth. | [21] | ||||
| Preferentially expressed antigen of melanoma (PRAME) | Membrane-bound protein | National Cancer Institute (Milan, Italy) [43] | Caucasian | 85.7–96.6% (n = 21) | IHC | PRAME Polyclonal Antibody (Clone NBP1-85418), Novus Boilogicals (USA) | Role in EMT reprogramming. Expression of PRAME was associated with negative ER status. | [58,59,60] |
| Sperm-associated antigen 9 (SPAG9) | Sperm function | National Institute of Immunology, Aruna Asaf Ali Marg, (New Delhi, India) [61] | Asian | NA | IHC | Polyclonal antibody to SPAG9 was prepared in authors’ laboratory | Analysis of 100 breast cancer tissues (94 infiltrating ductal carcinomas [IDC], 2 ductal carcinomas in situ [DCIS] and 4 invasive lobular carcinomas [ILC]) revealed that 88% of samples stained positive for SPAG9. Role in invasiveness of breast cancer. Downregulation could reduce invasive potential of TNBC. | [61,62] |
| Sperm protein associated with the nucleus X-linked (SPANX) | Sperm function | See Above | Required for metastasis. Interacts with lamin A/C at the inner nuclear membrane and involved in the formation of actin-rich cellular protrusions that reorganise the extracellular matrix. | [52,53] | ||||
| Testes-specific protease 50 (TSP50) | Oncogene | Northeast Normal University (Changchun, China) | Caucasian | NA | IHC | TSP50 Monoclonal Antibody was prepared in authors’ laboratory | Analysis of 88 clinical breast cancer tissue microarrays (BR955 and BR 1101 from US Biomax, Rockville, MD, USA) revealed that 90.9% of specimens stained positive for TSP50 compared to 10% of adjacent normal tissues. Role in cell growth. Knockdown of TSP50 in breast cancer cells significantly inhibits cellular proliferation. TSP50-positive tumours were associated with negative ER expression and higher grade. | [63,64] |
| Zinc-finger protein 165 (ZNF165) | Gene regulation | Simmons Comprehensive Cancer Center, UT-Southwestern Medical Center, Dallas [27] | Caucasian | 90% (n = 10) | IHC | ZNF165 (H00007718), Novus Biologicals (USA) | Enhances growth and survival of human TNBC cells both in vitro and in vivo by regulating TGF-β signalling. Frequently overexpressed in TNBC. | [27,65] |
| Tripartite motif containing 27 (TRIM27) | Gene regulation | Simmons Comprehensive Cancer Center, UT-Southwestern Medical Center, Dallas [27] | Caucasian | NA | TCGA | TRIM27 expression was significantly elevated in TNBC compared to normal breast tissue based on TCGA data. Displayed difference in cellular localisation, as it was mainly cytoplasmic in normal breast epithelia and more nuclear in TNBC tissues. Regulates TGFβ-dependent transcription in complex with ZNF165, ZNF446 and SMAD in TNBC. | [27,65] | |
| Other CTAs with increased expression in TNBC | ||||||||
| Actin like 8 (ACTL8) | Cellular architecture | National Centre for Tumour Diseases (Heidelberg, Germany) [66] | Caucasian | 57% (n = 98, TCGA) | TCGA | Frequently expressed in TNBC based on in silico analysis. | [66] | |
| Chromosome X open reading frame 6/ mastermind-like domain containing 1/Kita-Kyu-Shu lung cancer antigen-1 (CXorf6/MAMDL1/KK-LC-1/CT83) | Development of male genitalia Not known | Johannes Gutenberg-University (Mainz, Germany) [67] | Caucasian | 64.7% (n = 17, from commercial vendor) | IHC | Anti-CXorf61-A polyclonal antibody | Frequently expressed in TNBC. | [67] |
| Kitasato University Medical Center (Japan) [68] | Asian | 100% (n = 8) | IHC | Mouse monoclonal antibody was prepared by CLEA Japan (Japan) | Frequently expressed in TNBC based on in silico analysis. Frequently overexpressed in TNBC and tumours without ER expression. | [66,68] | ||
| Sperm protein 17 (SP17) | Sperm function | University of Texas MD Anderson Cancer [69] | Caucasian | 47.2% (n = 36) | IHC | Antibody against SP17 | SP17 is frequently expressed in primary breast tumours and in TNBC. | [69] |
| Wilms tumour-1 (WT-1) | Transcription factor | European Institute of Oncology (Milan, Italy) [57] | Caucasian | 54% (n = 27) | IHC | WT1 Monoclonal Antibody (Clone WT49), Monosan (Netherlands) | Highest expression in TNBC compared to other breast cancer subtypes. | [57] |
| Target | Clinical Trials.Gov Identifier | Type | Drug Details | Phase | Recruitment Status | Breast Cancer Subtypes/Other Cancers |
|---|---|---|---|---|---|---|
| NY-ESO-1 | NCT03093350 | T-cell immunotherapy | Tumour-associated antigen (TAA)-specific cytotoxic T- lymphocytes targeting NY-ESO-1, MAGEA4, PRAME, survivin and SSX2 | Phase II | Active, not recruiting | Metastatic or locally recurrent unresectable breast cancer |
| NCT02015416 | Cancer vaccine | IDC-G305: immunotherapy consisting of recombinant NY-ESO-1 antigen and the adjuvant GLA-SE | Phase I | Completed | Breast cancer, melanoma, ovarian cancer, sarcoma or NSCLC | |
| NCT01522820 | Cancer vaccine | DEC-205/NY-ESO-1 fusion protein CDX-1401 with and without sirolimus | Phase I | Completed | Breast cancer, other solid tumours | |
| NCT00291473 | Cancer vaccine | Cholesterol-bearing hydrophobized pullulan HER2 protein 146 (CHP-HER2) and NY-ESO-1 protein (CHP-NY-ESO-1) in combination with OK-432 | Phase I | Completed | HER2- and/or NY-ESO-1-expressing cancers | |
| NCT01967823 | T-cell immunotherapy | Anti-ESO mTCR-engineered peripheral blood lymphocytes with high-dose aldesleukin | Phase II | Completed | Metastatic cancer, including melanoma whose tumours express the ESO antigen | |
| NCT02661100 | Cancer vaccine | CDX-1401: human monoclonal antibody specific for DEC-205, fused to full-length tumour antigen NY-ESO-1 in combination with poly-ICLC and pembrolizumab | Phase I/II | Withdrawn (Drug unavailable) | Advanced TNBC, NSCLC, small-cell lung cancer, urothelial cancer, mesothelioma, malignant melanoma | |
| NCT02457650 | T-cell immunotherapy | Anti-NY-ESO-1 TCR transduced T cells | Phase I | Unknown | Breast cancer, other solid tumours | |
| NCT00623831 | Cancer vaccine | mixed bacteria vaccine in patients with tumours expressing NY-ESO-1 antigen | Phase I | Completed | Breast cancer, other solid tumours | |
| NCT03159585 | T-cell immunotherapy | TAEST16001: NY-ESO-1-specific TCR affinity enhancing specific T-cell therapy | Phase I | Completed | Breast cancer stage IV, other advanced solid tumours | |
| NCT01234012 | Cancer vaccine | MF-001: CHP-NY-ESO-1 complex consisting of recombinant NY-ESO-1 protein and cholesteryl hydrophobized pullulan (CHP) | Phase I | Completed | Metastatic or refractory breast cancer, other solid tumours | |
| NCT00948961 | Cancer vaccine | CDX-1401 with immune stimulants such as resiquimod and poly-ICLC (Hiltonol) | Phase I/II | Completed | Advanced malignancies expressing NY-ESO-1 | |
| MAGE | NCT04639245 | T-cell immunotherapy | Genetically engineered cells (MAGE-A1-specific T-cell receptor-transduced autologous T cells) and atezolizumab | Phase I/II | Not yet recruiting | Metastatic TNBC, urothelial cancer or NSCLC |
| NCT02153905 | T-cell immunotherapy | Autologous T cells transduced with an anti-MAGE-A3 HLA-A*01-restricted TCR (MAGE-A3-01) TCR and aldesleukin | Phase I/II | Terminated | Breast cancer, cervical cancer, renal cancer, melanoma, bladder cancer | |
| NCT02111850 | T-cell immunotherapy | HLA-DP0401/0402 restricted anti-MAGE-A3 TCR-gene engineered lymphocytes and aldesleukin | Phase I/II | Active, not recruiting | Breast cancer, cervical cancer, renal cancer, urothelial cancer, melanoma | |
| NCT00020267 | Cancer vaccine | MAGE-12 peptide vaccine | Phase I | Completed | Refractory metastatic cancer expressing MAGE-12 antigen: Breast cancer, other solid tumours | |
| NCT03093350 | T-cell immunotherapy | Tumour-associated antigen (TAA)-specific cytotoxic T- lymphocytes targeting NY-ESO-1, MAGEA4, PRAME, survivin and SSX2 | Phase II | Active, not recruiting | Any breast cancer patient with metastatic or locally recurrent unresectable breast cancer | |
| PRAME | NCT03093350 | T-cell immunotherapy | Tumour-associated antigen (TAA)-specific cytotoxic T-lymphocytes targeting NY-ESO-1, MAGEA4, PRAME, survivin and SSX2 | Phase II | Active, not recruiting | Any breast cancer patient with metastatic or locally recurrent unresectable breast cancer |
| NCT00423254 | Cancer vaccine | DNA vector pPRA-PSM with synthetic peptides, E-PRA and E-PSM | Phase I | Completed | Advanced solid malignancies: Breast cancer, other solid tumours | |
| MSLN | NCT02792114 | T-cell immunotherapy | Mesothelin-specific chimeric antigen receptor-positive T cells | Phase I | Recruiting | Breast cancer, metastatic HER2-negative breast cancer |
| NCT02414269 | T-cell immunotherapy | Mesothelin-targeted T cells | Phase I/II | Recruiting | Breast cancer, malignant pleural disease, mesothelioma, metastases, lung cancer | |
| NCT02580747 | T-cell immunotherapy | Anti-meso-CAR vector transduced T cells | Phase I | Unknown | TNBC, other mesothelin-positive tumours | |
| NCT03102320 | Antibody-drug conjugate | Anetumab–ravtansine: mesothelin-targeting antibody-drug conjugate | Phase Ib | Active, not recruiting | TNBC, cholangiocarcinoma, adenocarcinoma of the pancreas, NSCLC, gastric adenocarcinoma | |
| NCT02485119 | Antibody-drug conjugate | BAY94-9343: anetumab– ravtansine | Phase I | Completed | Advanced malignancies | |
| PSCA | NCT03927573 | Antibody | GEM3PSCA: PSCA-targeted bispecific antibody engaging T cells | Phase I | Recruiting | PSCA-positive cancer: urogenital tract (renal, transitional cell, prostate), NSCLC, breast and pancreatic cancer refractory to standard treatments |
| ROR2 | NCT03504488 | Antibody-drug conjugate | Conditionally active biologic (CAB) ROR2-targeted antibody-drug conjugate (CAB-ROR2-ADC) | Phase I/II | Recruiting | TNBC, locally advanced unresectable or metastatic solid tumours that have failed all available standard therapies, NSCLC, soft tissue sarcoma |
| WT1 | NCT01220128 | Cancer vaccine | GSK2302024A: recombinant WT1 antigen-specific cancer immunotherapeutic (ASCI) | Phase II | Terminated (negative phase III of another study product from same technology platform) | WT1-positive stage II or III breast cancer |
| NCT02018458 | Cancer vaccine | Cyclin B1/WT-1/CEF (antigen)-loaded dendritic cell vaccination with preoperative chemotherapy | Phase I/II | Completed | TNBC, ER+/HER2-breast cancer | |
| NCT03761914 | Cancer vaccine | Galinpepimut-S: WT1 analogue peptide vaccine | Phase I/II | Recruiting | TNBC, acute myelogenous leukaemia, ovarian cancer, colorectal cancer, small-cell lung cancer | |
| NCT01291420 | Cancer vaccine | Autologous WT1 mRNA-transfected dendritic cell vaccine | Phase I/II | Unknown | Breast cancers, glioblastoma, renal cell carcinoma, sarcomas, malignant mesothelioma, colorectal tumours |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lam, R.A.; Tien, T.Z.; Joseph, C.R.; Lim, J.X.; Thike, A.A.; Iqbal, J.; Tan, P.H.; Yeong, J.P.S. Cancer-Testis Antigens in Triple-Negative Breast Cancer: Role and Potential Utility in Clinical Practice. Cancers 2021, 13, 3875. https://doi.org/10.3390/cancers13153875
Lam RA, Tien TZ, Joseph CR, Lim JX, Thike AA, Iqbal J, Tan PH, Yeong JPS. Cancer-Testis Antigens in Triple-Negative Breast Cancer: Role and Potential Utility in Clinical Practice. Cancers. 2021; 13(15):3875. https://doi.org/10.3390/cancers13153875
Chicago/Turabian StyleLam, Runyi Adeline, Tracy Zhijun Tien, Craig Ryan Joseph, Johnathan Xiande Lim, Aye Aye Thike, Jabed Iqbal, Puay Hoon Tan, and Joe Poh Sheng Yeong. 2021. "Cancer-Testis Antigens in Triple-Negative Breast Cancer: Role and Potential Utility in Clinical Practice" Cancers 13, no. 15: 3875. https://doi.org/10.3390/cancers13153875