
Current Advances in Basic and Translational Research of Cholangiocarcinoma
by
 ,
,
Keisaku Sato
1,* ,
,
Leonardo Baiocchi
2,
Lindsey Kennedy
1,3,
Wenjun Zhang
4,
Burcin Ekser
4,
Shannon Glaser
5,
Heather Francis
1,3 and
Gianfranco Alpini
1,3 1
Division of Gastroenterology and Hepatology, Department of Medicine, Indiana University School of Medicine, Indianapolis, IN 46202, USA
2
Hepatology Unit, Department of Medicine, University of Tor Vergata, 00133 Rome, Italy
3
Department of Research, Richard L. Roudebush VA Medical Center, Indianapolis, IN 46202, USA
4
Division of Transplant Surgery, Department of Surgery, Indiana University School of Medicine, Indianapolis, IN 46202, USA
5
Department of Medical Physiology, Texas A&M University College of Medicine, Bryan, TX 77807, USA
*
Author to whom correspondence should be addressed.
Cancers 2021, 13(13), 3307; https://doi.org/10.3390/cancers13133307
Submission received: 31 May 2021
/
Revised: 25 June 2021
/
Accepted: 26 June 2021
/
Published: 1 July 2021
(This article belongs to the Special Issue Theranostic Advances in Hepatobiliary Tumors)
Abstract
:Simple Summary
Cholangiocarcinoma (CCA) is highly malignant biliary tract cancer, which is characterized by limited treatment options and poor prognosis. Basic science studies to seek therapies for CCA are also limited due to lack of gold-standard experimental models and heterogeneity of CCA resulting in various genetic alterations and origins of tumor cells. Recent studies have developed new experimental models and techniques that may facilitate CCA studies leading to the development of novel treatments. This review summarizes the update in current basic studies of CCA.
Abstract
Cholangiocarcinoma (CCA) is a type of biliary tract cancer emerging from the biliary tree. CCA is the second most common primary liver cancer after hepatocellular carcinoma and is highly aggressive resulting in poor prognosis and patient survival. Treatment options for CCA patients are limited since early diagnosis is challenging, and the efficacy of chemotherapy or radiotherapy is also limited because CCA is a heterogeneous malignancy. Basic research is important for CCA to establish novel diagnostic testing and more effective therapies. Previous studies have introduced new techniques and methodologies for animal models, in vitro models, and biomarkers. Recent experimental strategies include patient-derived xenograft, syngeneic mouse models, and CCA organoids to mimic heterogeneous CCA characteristics of each patient or three-dimensional cellular architecture in vitro. Recent studies have identified various novel CCA biomarkers, especially non-coding RNAs that were associated with poor prognosis or metastases in CCA patients. This review summarizes current advances and limitations in basic and translational studies of CCA.
1. Introduction
Cholangiocarcinoma (CCA) emerges from the biliary epithelium and is the second most common primary liver cancer after hepatocellular carcinoma (HCC) [1]. CCA is a relatively rare cancer, but worldwide incidence has been increasing in past years [2]. CCA is an aggressive malignancy, and early diagnosis is typically challenging due to asymptomatic characteristics at the early stage, resulting in poor prognosis with five-year survival under 20% [1,3]. Thus, treatment options are limited for diagnosed patients, who, in the majority of the cases, are already at advanced stages. Although liver transplantation or curative surgical resection is the sole effective procedures, CCA patients often have recurrence after surgery. A meta-analysis for 428 patients with unresectable CCA showed high recurrence rates after three years of orthotopic liver transplantation for patients with (24.1%) or without (51.7%) neoadjuvant chemoradiation [4]. Therefore, there is a critical need to develop novel diagnostic testing and therapeutic strategies for CCA [5].
Unique characteristics of CCA provide difficulties in basic and translational studies and prevent research progress in the field. The major barriers in CCA studies come from its heterogeneity. CCA is categorized into three types depending on the anatomical location of the tumor: intrahepatic (iCCA), perihilar (pCCA), and distal (dCCA) [5,6]. Since CCA tumor cells have biliary phenotypes, such as cytokeratin (CK)-7 and CK-19 expression [7], cholangiocytes have been established as the primary source of CCA tumors. Primary sclerosing cholangitis (PSC) is a bile duct disorder characterized by ductular reaction, biliary damage and inflammation, and liver fibrosis, and patients with PSC are at increased risk for CCA development, indicating carcinogenesis in cholangiocytes [8]; however, CCA can emerge from hepatic progenitor cells (HPCs) localized in the Canals of Hering, and this type of CCA is referred to as cholangiolocellular carcinoma [9,10]. Animal studies have shown that hepatocytes can initiate iCCA tumors with biliary phenotypes including CK-19 expression [11,12]. Furthermore, previous studies have reported combined hepatocellular cholangiocarcinoma (cHCC-CCA), which is liver cancer showing pathological features of both HCC and CCA [13,14,15]. Although the origin of cHCC-CCA may be HPCs, the pathophysiology of cHCC-CCA is largely undefined [16]. During liver injury, hepatocytes can transdifferentiate into cholangiocytes and vice versa, and HPCs can differentiate into both hepatocytes and cholangiocytes [17]. It is challenging to distinguish and identify each type of CCA and its origin. Pathophysiological characteristics of CCA tumors may differ depending on the type or origin of the tumor. This plasticity of hepatic cells and multiple potential origins of tumors contribute to the high heterogeneity of CCA and add to the complexity and difficulties in CCA research [18]. Recent studies have introduced novel biomarkers for diagnosis, experimental models, and advanced techniques to facilitate CCA studies. This review summarizes current advances and limitations in basic and translational research of CCA.
2. Current Advances in Basic CCA Research
2.1. Whole-Genome Screening
It is known that CCA tumors often have genetic mutations or aberrations. Previous studies identified various genetic mutations, and commonly mutated genes include TP53, KRAS, IDH1/2, and FGFR fusion [19,20]. Genomic profiling for 803 patients with biliary tract cancer found the most frequently altered genes, such as TP53 (53%) and KRAS (26%), which were associated with poor prognosis in this cohort [21]. Based on these findings, clinical trials for IDH or FGFR inhibitors are ongoing, although results are often disappointing for CCA patients [5,22]. Since CCA is highly heterogeneous, only a limited percentage of patients have mutations or aberrations on targeted genes (under 10% for IDH1/2 mutations and 3–50% for FGFR2 fusion [22]), meaning the limited efficacy of drugs targeting mutated genes. Genome-wide screening to identify novel candidate genes that are altered in CCA tumors is ongoing research. A previous study analyzed 412 samples of biliary tract cancer including iCCA, pCCA, and dCCA by whole-exome sequencing and whole-genome sequencing [23]. This study identified commonly mutated genes, such as TP53, KRAS, and SMAD4, and found a novel deletion of MUC17 on 7q22.1 [23]. Patients with MUC17 deletion showed poor survival rates [23]. The landscape of genome and transcriptome in CCA may facilitate identification of target genes for novel therapies. Whole-exome and transcriptome sequencing for tumor and corresponding peritumor tissue samples of 9 iCCA patients identified an average of 378 somatic single nucleotide variants and 2366 differentially expressed genes in tumor tissues [24]. Interaction networks have shown that somatic mutations are highly correlated with altered gene expression, and mutations in key genes, such as TP53, may alter expression of numbers of genes [24]. Whole-exome sequencing for 318 iCCA patients identified 32 mutated genes associated with poor survival rates, which included TP53 and KRAS [25]. Genome-wide transcriptomic profiling for iCCA tumors developed a transcriptomic panel with 8 genes, BIRC5, CDC20, CDH2, CENPW, JPH1, MAD2L1, NEIL3, and POC1A, which were robustly identified in patients with recurrence [26]. This study has demonstrated that the combination of this transcriptomic panel and clinical features, such as tumor size, could be useful to predict the risk of CCA recurrence [26]. A previous study categorized 133 cHCC-CCA samples into three subtypes according to the localization of HCC and CCA tumors: separate, combined, and mixed [27]. Genomic and transcriptomic sequencing when comparing HCC and iCCA found that genomic landscape and transcriptomic profiles of the combined type of cHCC-CCA was similar to those of iCCA, and the mixed type of cHCC-CCA was similar to HCC, indicating that therapies for cHCC-CCA may be adopted depending on subtypes [27]. This study showed that a high percentage of cHCC-CCA tumors expressed Nestin compared to HCC or iCCA, and positivity of Nestin expression was associated with poor survival rates, suggesting that Nestin could be useful as a novel biomarker for diagnosis testing of cHCC-CCA [27].
DNA methylation regulates gene expression, and hypermethylation on specific promoter CpG islands have been identified in CCA tumors, which may play a key role in the pathophysiology of CCA [20]. Genome-wide analysis for promoter methylation identified hypermethylation on promoter regions of genes associated with Wnt signaling, such as SFRP2, which expression levels were downregulated in CCA tumors [28]. Analysis of gene expression and gene methylation for CCA samples using datasets obtained from Gene Expression Omnibus (GEO) database found 98 hypermethylated, downregulated genes and 93 hypomethylated, upregulated genes [29]. Another analysis using GEO datasets showed that CCA patients with widespread hypermethylation of promoter-related CpG sites suffered unfavorable prognosis with poor survival rates [30]. Genome-wide profiling of DNA methylation and gene/microRNA (miRNA) expression using data obtained from The Cancer Genome Atlas (TCGA) have shown altered DNA methylation on 12,259 CpGs and altered expression patterns in 3,305 genes and 101 miRNAs [31]. This study identified candidate genes and miRNAs, such as MDK, DEPDC1, miR-22, and miR-551b, which could be useful as prognostic biomarkers of CCA [31]. These studies show that whole-genome screening and landscape of mutations, genome, transcriptome, and methylation may lead to identification of novel biomarkers and therapeutic targets of CCA; however, a genome-wide association study for CCA is still not available to date, which may provide novel understandings of genetic variations associated with CCA.
2.2. Animal Models
Availability of experimental models is a key factor to facilitate basic and translational research; however, no gold standard CCA models are available to date regardless of various animal models introduced in previous studies [32,33,34]. Xenograft models generated by subcutaneous transplantation of human CCA cell lines into flanks of nude mice are still one of the most commonly used models in CCA studies, but this model does not mimic CCA tumorigenesis and microenvironment in addition to the species mismatched conditions [33]. Administration of toxins, such as thioacetamide (TAA) and diethylnitrosamine (DEN), was used in previous studies to generate CCA tumors in rodents; however, ~22-week TAA administration for rats [35] and combination of DEN treatments with bile duct ligation (BDL)-induced biliary damage in mice [36] do not mimic pathological conditions of CCA patients. CCA tumors often have mutations in specific genes, such as TP53 and KRAS, and studies in past years developed various genetically engineered mouse models targeting these genes [32,33,34]. Mice with a mutation in a target gene, such as KrasG12D mice, which have Alb-Cre-mediated somatic KRAS activation, showed iCCA-like tumor in vivo, although tumors showed low penetrance and long latency [37]. KrasG12D;p53L/L mice, which have liver-specific KRAS activation and p53 deletion, showed higher penetrance and iCCA tumors in a short time [37]. The chance of CCA development in a practically reasonable time frame is limited in mice with a mutation or deletion in a single target gene; therefore, to increase penetrance and shorten the period of CCA tumor development, two or more genetic mutations or deletions are increasingly introduced in current CCA mouse models [32,33,34]. However, CCA is heterogeneous and only a limited percentage of CCA patients share mutations in specific genes. Conditions in these genetically engineered models (e.g., Alb-Cre-mediated Smad4Co/CoPtenCo/Co mice [38]) are rare in CCA patients. In addition, some models, such as Alb-Cre-mediated Ptenf/fGrp94f/f mice [39], show both HCC and CCA. It is unclear if these mice mimic conditions of human patients with CCA or cHCC-CCA. Since Alb-Cre mice have Cre expression in hepatocytes and hepatic stellate cells [40], not cholangiocytes, these mice may have high possibilities to have a mixture of HCC and CCA. Searching for ideal CCA models still continues in current studies. Injection of three oncogenic plasmids, myristoylated AKT1, mutated YAP, and pCMV-Sleeping Beauty, into mouse livers generated iCCA-like peripheral tumors and pCCA-like perihilar tumors after 10 weeks [41]. Gene and protein expression levels, such as Fgfr2 and αSMA, were different between peripheral and perihilar tumors, showing that characteristics of tumors generated by the same technique can differ depending on the tumor location [41]. A previous study has demonstrated that Opn-CreER triggers recombination in 99.9% of cholangiocytes while Ck19-CreER has only 32% recombination efficiency [42]. A subsequent study generated Opn-Cre-mediated cholangiocyte-specific KrasG12D mice and fed them with 3,5-diethoxycarbonyl-1,4-dihydrocollidine (DDC) diet [43]. DDC diet induces bile duct damage and ductular reaction in mice [17]. DDC-fed KrasG12D mice showed iCCA tumors after 21 weeks [43]. This study has demonstrated that iCCA tumors in this model, as well as human patients overexpress Tensin-4, which drives CCA cell proliferation [43]. Alb-Cre-mediated KrasG12D/CDH1ΔL mice showed tumors after 8 months of age, but most of tumors were HCC, and CCA was only 10% in the established tumors [44]. Feeding with high-fat diet generated liver tumors in KrasG12D/CDH1ΔL mice at 5 months of age, and 34% of tumors was CCA, showing that high-fat diet facilitates tumorigenesis in the liver and increases the possibility of CCA development [45]. These studies indicate that the combination of genetic alterations and feeding diets may significantly affect penetrance and the time period of cancer development in the liver as well as phenotypes of cancer (HCC, CCA, or mix). Table 1 lists selected genetically engineered CCA mouse models used in current CCA studies. Limitations of these models include the lack of tumorigenic conditions in patients with cholangiopathies. As mentioned, PSC is a common risk factor for CCA development; however, the most common PSC model mice, Mdr2−/− mice show only HCC, not CCA, at 7–12 months of age regardless of their genetic backgrounds [46]. Detailed mechanisms of biliary tumorigenesis in cholangiopathies are undefined, and appropriate animal models resembling conditions in patients are still unavailable.
Although it is still common to generate human CCA tumors in mice by transplantation of CCA cell lines, such as Mz-ChA-1 cells [33], recent studies have introduced techniques to generate CCA tumors using tissues obtained from patients. A previous study subcutaneously transplanted fresh iCCA tumors excised from patients into NOD/SCID mice [47]. Although only one tumor out of 17 (5.8%) was successfully engrafted after 4 months, generated tumors could be explanted and implanted into new mice [47]. This patient-derived xenograft (PDX) mouse model maintained histological tumor features as well as genetic alterations, such as KRAS G12D mutation, with the original tumor [47]. Another study generated PDX mice highly successfully (75%) by transplantation of frozen CCA tissues into Balb/c Rag-2−/−/Jak3−/− mice [48]. This study established four novel CCA cell lines from generated PDX mice, and those CCA cell lines showed high transplantation efficiency (up to 100%), which may be useful for generation of xenograft CCA models [48]. A large scale study used tumor samples created from surgical resection specimens or radiographic biopsies of 87 patients with biliary tract cancers including iCCA, pCCA, and dCCA [49]. Out of 87 patient specimens, 47 PDX models were successfully generated in NOD/SCID mice [49]. Generated PDX mice maintained histological and genetic characteristics compared to original tumors in patients [49]. This study has demonstrated that patients whose specimen are successful for engraftment suffer poor survival rates compared to patients with tumors that do not generate PDX mice [49]. PDX mouse models resemble unique and heterogeneous characteristics of CCA tumors in the individual patient. These models may be useful for biomarker detection, mutation screening, and drug testing, which may lead to customized treatments for each patient.
2.3. In Vitro Models
Various human CCA cell lines, such as HuCCT1 and TFK-1, have been used for in vitro studies, and genomic and transcriptomic analysis showed that these cell lines shared similar mutational signatures and transcriptomic profiles compared to primary tumors [50]. Due to the heterogeneity of CCA, multiple cell lines are usually used to confirm results, and cell line authentication by detecting short tandem repeats is required to determine genetic stability and contamination. Novel human CCA cell lines have been established in recent studies from PDX mice [48] or iCCA and pCCA tumor tissues [51]. Recent studies have established the methodology to generate and characterize organoids, three-dimensional (3D) cultured mini-organs, using primary liver tissues of patients with cholangiopathies or CCA [52,53]. A previous study established CCA organoids with 50% success rates using resected tissue specimens of iCCA patients and maintained them over one year [54]. Established organoids showed similar histopathological features to primary tumors [54]. Organoids mimic 3D cellular architecture and interaction more appropriately compared to two-dimensional (2D) monolayer cultures and may be suitable for in vitro models in CCA studies [52]. A previous study cultured patient-derived iCCA cells in 2D or 3D culture systems and compared cellular functions and expressions [55]. CCA organoids showed significantly higher liver function indexes, such as expression levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST), as well as elevated fibrosis indexes including matrix metalloproteinase (MMP) expression compared to same CCA cells cultured as 2D monolayers, indicating that cell functions and characteristics may differ between 2D and 3D culture systems, and CCA cells may be more functional in 3D cultures [55]. CCA organoids are useful for drug testing or genetic screening. A previous study cultured CCA organoids established from iCCA tissues in glucose-free media and found that CCA organoids showed reduced proliferation but elevated gemcitabine resistance in glucose-free media [56]. Patient-derived CCA organoids showed enlarged mitochondria compared to organoids generated from normal liver tissues, and knockdown of genes that are associated with mitochondrial fusion process, OPA1 and MFN1, inhibited mitochondrial fusion and cell viability in populating cells of CCA organoids [57]. A study using these CCA and normal liver organoids found that CCA organoids expressed elevated lysyl-tRNA synthetase (KARS), and treatment of KARS inhibitor cladosporin decreased cell viability of CCA organoid cells, showing potential anti-cancer effects of cladosporin for CCA [58].
A recent study isolated cholangiocytes from bile ducts of wild-type (WT) or Ink4a/Arf−/− mice and transfected them with retrovirus vectors for KRASG12V to generate malignant murine cholangiocytes [59]. Although KRASG12V expression inhibited proliferation of WT cholangiocytes, transplantation of KRASG12V-expressing Ink4a/Arf−/− cholangiocytes generated CCA tumors in WT mice, and CCA organoids were established from tumor tissues of these syngeneic CCA mice [59]. This study has demonstrated that CCA organoids have cancer stem cell-like characteristics, such as high expression of Cd24, Cd44, and Sca1, as well as the ability to generate secondary tumors when transplanted in mice; however, the same cells cultured in 2D monolayers have decreased successful rates to generate CCA tumors after transplantation into syngeneic mice, indicating that organoids may be a better technique to retain CCA cell functions in vitro compared to classic monolayer cultures [59].
Another study established hepatic organoids using liver tissues explanted from WT or TP53−/− mice, and FGFR2 fusion proteins identified in CCA patients, such as FGFR2-TACC3 and FGFR2-BICC1, were expressed in established organoids by retroviruses [60]. Expression of FGFR2 fusion proteins inhibited WT organoid growth, but transplantation of TP53−/− organoids expressing fusion proteins generated CCA tumors in NOD/SCID mice while TP53−/− organoids with no fusion protein expression did not [60]. Although FGFR inhibitors may be therapeutic for CCA patients with FGFR2 fusion proteins, some patients show resistance and the efficacy of FGFR inhibitors may be limited [61]. This study has demonstrated that combination of FGFR kinase inhibitor BGJ398 and MEK1/2 inhibitor trametinib significantly inhibited tumor growth compared to treatments with BGJ398 alone using syngeneic CCA mouse models driven by FGFR2-BICC1-expressing organoids [60]. These studies showed that robust KRAS activation and FGFR2 fusion proteins inhibit proliferation and growth in normal hepatobiliary cells, and the combination with suppression/depletion of tumor-suppressor genes, such as Ink4a/Arf or TP53, is required for CCA tumorigenesis. Although syngeneic CCA mouse models were generated using murine organoids, patient-derived human organoids can also be the source of CCA tumors in xenograft mouse models. Transplantation of organoids established from human iCCA tumor tissues generated xenograft tumors in NOD/SCID mice, which show similar histological features compared to the original patient’s tumor [62]. Figure 1 summarizes recent experimental CCA models including PDX and organoid models, and Table 2 shows pros and cons for mentioned CCA animal models.
2.4. Biomarkers
Since early diagnosis is still challenging, searching for novel biomarkers that could be used for diagnostic testing or prediction of prognosis continues in current CCA studies. Various candidate biomarkers have been identified in previous studies, and the majority of current biomarkers includes genetic mutations, such as TP53 and KRAS mutations, proteins including cytokines, and non-coding RNAs [63,64]. This review focuses on recent advances in biomarker search, and for more information of other currently known CCA biomarkers, see previous reviews [63,64]. Samples to be analyzed in previous studies are usually tissue samples or fluid samples, such as serum, bile, or urine. For example, analysis of RNA-sequencing (RNA-seq) data obtained from TCGA, which include 36 CCA tissues and 9 adjacent normal tissues, identified bloom syndrome helicase (BLM) as a potential CCA biomarker [65]. BLM is upregulated in CCA tumors and cell lines, and high BLM expression is associated with poor survival rates [65]. Receiver operating characteristic (ROC) curve analysis showed that BLM could be useful for diagnostic testing [65]. Another study analyzing RNA-seq data obtained from TCGA identified multiple candidate biomarkers, such as cyclin-dependent kinase 1 (CDK1), which is significantly upregulated in CCA tumors and associated with poor survival rates [66]. Immunohistochemistry for liver tissues identified overexpression of cadherin 17 (CDH17 or CA17) in CCA, which was associated with poor survival rates [67]. Serum levels of carbohydrate antigen 19-9 (CA 19-9) is clinically useful as a biomarker of CCA [63], and a previous study has demonstrated that the combination of serum levels of CA 19-9 and dickkopf-related protein 1 (DKK-1) could provide better diagnostic and prognostic performance compared to CA 19-9 alone in iCCA patients [68].
Extracellular vesicles (EVs) are small particles secreted from cells and contain cargo proteins, DNAs, and RNAs, which regulate cellular functions in recipient cells [69]. EV-mediated intercellular communication may play a vital role in the pathophysiology of cholangiopathies as well as CCA [70,71,72]; therefore, EVs secreted from CCA tumors may contain unique cargo mediators that could be used as early diagnostic biomarkers. EVs isolated from serum samples of patients with CCA contained elevated levels of proteins, including pantetheinase and C-reactive protein, compared to EVs isolated from healthy individuals or patients with PSC or HCC [73]. Analysis of miRNAs contained in serum EVs of 36 CCA patients and 12 healthy individuals identified miR-200 family enriched in CCA EVs, and these miRNAs showed higher area under the ROC curve (AUC) than CA 19-9 [74]. Proteomic analysis for EVs isolated from human bile samples of 10 CCA and 10 choledocholithiasis (bile duct stone) patients identified 166 proteins as CCA-specific [75]. This study has demonstrated that CCA EVs contain significantly higher levels of claudin-3, which could be a useful biomarker to distinguish CCA and bile duct stones [75]. Transcriptomic analysis for isolated EVs from serum or urine samples found that EVs from CCA patients contain robust levels of various mRNAs, such as CMIP for serum EVs and UBE2C for urine EVs, compared to EVs isolated from healthy individuals or patients with PSC or ulcerative colitis [76]. These studies show the potentials of EVs as biomarker carriers and important samples for diagnostic testing.
Recent advances in CCA biomarkers include identification of non-coding RNAs, such as miRNAs, long non-coding RNAs (lncRNAs), circular RNAs (circRNAs), and P-element-induced wimpy testis (PIWI)-interacting RNAs (piRNAs). Numbers of candidate non-coding RNAs have been identified as potential biomarkers from CCA tissues or cell lines. For example, whole-transcriptome sequencing for 8 CCA tumor and adjacent normal tissues detected 2,895 mRNAs, 56 miRNAs, 151 lncRNAs, and 110 circRNAs that were differentially expressed in CCA tumors [77]. It is known that miR-21 and miR-122 are associated with CCA, and combined validation of plasma levels of miR-21, miR-122, and CA 19-9 showed better AUC compared to CA 19-9, showing the potentials of non-coding RNAs as diagnostic biomarkers to distinguish iCCA and control individuals [78]. This review introduces recently identified non-coding RNAs, which are associated with poor prognosis or the pathophysiology of CCA. For more information of other non-coding RNAs, see previous review articles [79,80,81,82]. For miRNAs, a previous study analyzed TCGA dataset of CCA and found that expression levels of miR-3913 were elevated in CCA tumors [83]. High expression of miR-3913 was associated with poor survival rates in CCA patients, indicating the potential as a prognostic biomarker [83]. A study using 30 CCA tumor tissues and 20 adjacent normal tissues showed that miR-29b was significantly downregulated in CCA tumors, and low miR-29b expression was associated with poor survival rates [84]. This study has demonstrated that miR-29b targets and regulates expression of DNA methyltransferase 3 beta (DNMT3B), which is upregulated in CCA tumors and induces CCA growth, indicating the potential of miR-29b/DNMT3B as a therapeutic target [84]. Other recent studies have identified miR-150, miR-144, miR-451a, miR-1182, and let-7a as being associated with CCA and could be useful as biomarkers [85,86,87] (Table 3).
Numerous studies have identified various lncRNAs, which are associated with CCA [81]. lncRNAs have binding sites for target miRNAs and inhibit their functions by sponging. As a result, expression levels of genes, which are targeted by sponged miRNAs, will be upregulated due to lncRNAs. This function of lncRNAs as competing endogenous RNAs (ceRNAs) may be vital in the pathophysiology of CCA. Data analysis for TCGA dataset of 36 CCA tissues and 9 control tissues constructed ceRNA network associated with CCA [88]. CCA tumors may express unique lncRNAs regulating downstream miRNAs and gene expression, which could be useful as CCA biomarkers. Analysis of TCGA dataset identified five lncRNAs, which expression levels were altered in CCA tumors, and combination of these lncRNAs showed better AUC compared to single lncRNA and the potentials as a biomarker to predict poor prognosis and recurrence [89]. A previous study using CCA tumor and adjacent normal tissues obtained from 57 patients has demonstrated that lncRNA FOXD2-AS1 is upregulated in CCA tumors as well as CCA cell lines, and patients with high FOXD2-AS1 expression showed worse prognosis [90]. This study showed that FOXD2-AS1 inhibited miR-760 functions by sponging, resulting in upregulated expression of target gene of miR-760, oncogene E2 transcription factor 3 (E2F3) in CCA [90]. Inhibition of FOXD2-AS1 or E2F3 decreased CCA cell proliferation, but inhibition of miR-760 promoted proliferation in vitro [90]. Table 4 lists other lncRNAs identified in recent CCA studies [91,92,93,94,95,96,97].
circRNAs are a type of non-coding RNA that have a closed loop structure. Recent CCA studies revealed the association of circRNAs with CCA and that circRNAs could function as ceRNAs similar to lncRNAs, inhibiting target miRNA functions and inducing downstream gene expression. A previous study has demonstrated that the crcRNA circ-LAMP1 is upregulated in CCA tumor tissues and cell lines, and high expression of circ-LAMP1 is associated with poor survival rates [98]. This study showed that circ-LAMP had binding sites for miR-556 and miR-567, and both miRNAs regulated expression of YY1 [98]. Inhibition of circ-LAMP1 decreased cell proliferation and invasion of CCA cell lines in vitro, as well as CCA tumor growth in xenograft mouse models, indicating the potentials of circ-LAMP1 as a biomarker and therapeutic target of CCA [98]. The authors isolated EVs from bile samples of CCA patients and performed circRNA profiling and found that circ-CCAC1 was enriched in CCA bile EVs and was highly expressed in CCA tumors and cell lines, which was associated with poor prognosis in patients [99]. circ-CCAC1 sponges miR-514a, which regulates YY1 expression, and high levels of circ-CCAC1 in circulating EVs promoted CCA growth and metastases in xenograft mouse models [99]. A study using 35 paired CCA tissues and adjacent normal tissues identified elevated expression of circ-DNM3OS associated with TNM stage and lymph node invasion in CCA patients [100]. circ-DNM3OS induces MORC family CW-type zinc finger 2 (MORC2) expression via sponging miR-145, and inhibition of circ-DNM3OS decreased tumor growth by upregulation of miR-145 and downregulation of MORC2 in xenograft mice [100]. These studies demonstrated the pathophysiological roles of circRNAs as ceRNAs in CCA. Other circRNAs identified in recent CCA studies are listed in Table 5 [101,102,103,104,105].
piRNA is a class of small non-coding RNA, which makes an RNA-protein complex with PIWI proteins. Functions of piRNAs include post-transcriptional regulation of gene expression, which is similar to miRNAs, and piRNAs may be involved in cancer development and have the potentials as novel cancer biomarkers [106]. A previous study isolated EVs from serum samples of patients with gastric cancer, and found that cancer-derived serum EVs contained elevated levels of piRNAs, such as piR-019308, compared to EVs isolated from healthy individuals, and ROC curve analysis showed the diagnostic potentials of these piRNAs (for piR-019308, AUC = 0.82) [107]. Although studies are limited for CCA and functional roles of piRNAs in CCA are largely undefined, a previous study performed piRNA profiling for EVs isolated from plasma samples of patients with CCA or gallbladder carcinoma [108]. This study identified various piRNAs upregulated in CCA-derived EVs compared to normal EVs, such as piR-10506469, as well as downregulated piRNAs, such as miR-17802142 [108]. Cargo levels of piR-10506469 in plasma EVs of CCA patients were significantly decreased after one week of surgery, indicating the correlation of EV piR-10506469 levels with CCA tumors [108]. Future studies may identify novel piRNAs associated with CCA development leading to novel diagnostic testing.
3. Emerging Roles of Gut Microbiota in CCA
One of the emerging fields in CCA studies includes the functional roles of gut microbiota. As mentioned earlier, PSC is a common risk factor for CCA development [8]. Previous studies showed that PSC patients had different gut bacteria profiles with decreased diversity, which is associated with cholestatic liver injury [109]; therefore, gut microbiota may be associated with biliary carcinogenesis and CCA. A previous study analyzed gut microbiota using stool samples from patients with HCC, iCCA, or liver cirrhosis and healthy individuals and found that iCCA patients had abundance of Lactobacillus, Actinomyces, Peptostreptococcaceae, and Alloscardovia genera compared to other patient groups [110]. Fluke infection with Opisthorchis viverrini or Clonorchis sinensis is a common risk factor for CCA, and O. viverrini infection in hamsters is an animal model to mimic CCA development [5]. Analysis of colorectal feces of hamsters demonstrated that O. viverrini infection altered gut microbiota, increasing Lachnospiraceae, Ruminococcaceae, and Lactobacillaceae, and decreasing Porphyromonadaceae, Erysipelotrichaceae, and Eubacteriaceae [111]. Myeloid-derived suppressor cells (MDSCs) are immunosuppressive cells with an ability to inactivate T cells, and high MDSC population is associated with CCA [112,113]. A previous study has demonstrated that cholestatic liver injury induces bacterial leakage of gut bacteria via elevated intestinal permeability, which promotes MDSC accumulation in the liver leading to CCA development in mouse models in vivo [114]. Although these studies indicate the association of gut bacteria with CCA, it is still undefined which genera/species of bacteria are involved in the pathophysiology of CCA. Further studies are required to elucidate the detailed mechanisms of CCA development induced by gut bacteria.
4. Conclusions and Future Perspectives
Although CCA studies are still limited and further studies are required, current studies have developed novel methodologies and techniques to facilitate CCA research. Important advances in CCA studies include new CCA models. PDX mouse models maintain genetic and histological features of primary tumors found in the patient and are suitable for genetic screening, drug testing, or even design of customized treatments for the patient. Syngeneic CCA mouse models have overcome the mismatch of species between CCA tumors and the host organisms. Xenograft mouse models established by transplantation of human CCA cell lines usually do not generate the tumor microenvironment, which is often observed in CCA patients [33]. Syngeneic CCA mouse models established by murine malignant cholangiocytes showed the tumor microenvironment-like conditions in vivo [59]. The tumor microenvironment is a dense stroma, which may play a vital role in the pathophysiology of CCA and could be a promising therapeutic target [115], although it is challenging to resemble the tumor microenvironment in current CCA animal models. In a previous study, rat CCA models established using malignant rat cholangiocyte lines, BDEneu cells, showed liver conditions like CCA tumor microenvironment [116]. Syngeneic CCA models may be suitable as animal models for studies of CCA tumor microenvironment. Although PDX mouse models maintain characteristics of patients’ primary CCA tumors, it is undefined if PDX models also resemble CCA microenvironment that is found in the patient. Since both syngeneic rat and mouse models showed CCA tumor microenvironment-like stroma, matched species between CCA tumors and the host animals may be required to generate CCA microenvironment in animals. Syngeneic animal models established using genetically modified cancerous cholangiocytes or organoids still do not mimic CCA development associated with biliary damage, such as PSC. Combination of biliary damage with other factors may be required to form CCA tumors. For example, only KRAS activation in cholangiocytes inhibited cell proliferation and did not generate CCA tumors, and knockout of Ink4a/Arf was also required [59]. DDC diet generated iCCA tumors in Opn-Cre-mediated cholangiocyte-specific KrasG12D mice after 21 weeks but control feeding did not [43]. High-fat diet feeding increased the chance of CCA development (34%) in KrasG12D/CDH1ΔL mice compared to control diet (10%) [44]. These studies indicate that CCA development may be mediated by multiple factors, damage, or mutations in hepatobiliary cells. Future studies may establish CCA development models from PSC (e.g., Mdr2−/− mice with other factors) to mimic transitional conditions from biliary inflammation and damage to CCA. Establishment of CCA organoids is another important advance in CCA research. Compared to 2D cell culture, 3D organoid system can retain CCA tumor functions, such as the ability to generate secondary tumors in mice after excision and re-transplantation [59]. Data using 3D organoids may be required for future CCA studies to confirm results and functions of CCA tumor cells. Searching for genetic mutations, aberrations, methylations, or biomarkers is still ongoing in current basic studies, but accumulation of these studies may lead to the development of novel diagnostic testing or therapies. According to PubMed, papers or entries found in a search with “cholangiocarcinoma” have been increasing in every year (2010: 607, 2015: 1092, and 2020: 1682 entries). CCA studies may develop better models and identify novel therapeutic approach to overcome this rare heterogeneous cancer soon.
Author Contributions
Writing—original draft preparation, K.S.; writing—review and editing, L.B., L.K., W.Z., B.E., S.G., H.F. and G.A.; funding acquisition, S.G., H.F. and G.A.; supervision and finalization, K.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Hickam Endowed Chair, Gastroenterology, Medicine, Indiana University, the Indiana University Health—Indiana University School of Medicine Strategic Research Initiative, the VA Merit awards to G.A. (5I01BX000574) and H.F. (1I01BX003031) from the United States Department of Veteran’s Affairs, Biomedical Laboratory Research and Development Service and NIH grants DK108959 and DK119421 (H.F.), DK054811, DK115184, DK076898, DK107310, DK110035, DK062975 and AA028711 (G.A. and S.G.) and the PSC Partners Seeking a Cure (G.A.). Portions of these studies were supported by resources at the Central Texas Veterans Health Care System, Temple, TX, Richard L. Roudebush VA Medical Center, Indianapolis, IN, and Medical Physiology, Medical Research Building, Temple, TX. The views expressed in this article are those of the authors and do not necessarily represent the views of the Department of Veterans Affairs.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviation
2D = two-dimensional; 3D = three-dimensional; ALT = alanine aminotransferase; AST = aspartate aminotransferase; AUC = area under the ROC curve; BLM = bloom syndrome helicase; BDL = bile duct ligation; CA 19-9 = carbohydrate antigen 19-9; CCA = cholangiocarcinoma; CDK1 = cyclin-dependent kinase 1; ceRNAs = competing endogenous RNAs; cHCC-CCA = combined hepatocellular cholangiocarcinoma; circRNAs = circular RNAs; CK = cytokeratin; dCCA = distal cholangiocarcinoma; DDC = 3,5-diethoxycarbonyl-1,4-dihydrocollidine; DEN = diethylnitrosamine; DKK-1 = dickkopf-related protein 1; DNMT3B = DNA methyltransferase 3 beta; E2F3 = E2 transcription factor 3; EVs = extracellular vesicles; GEO = Gene Expression Omnibus; HCC = hepatocellular carcinoma; HPCs = hepatic progenitor cells; iCCA = intrahepatic cholangiocarcinoma; KARS = lysyl-tRNA synthetase; lncRNAs = long non-coding RNAs; MDSCs = myeloid-derived suppressor cells; miRNAs = microRNAs; MMP = matrix metalloproteinase; MORC2 = MORC family CW-type zinc finger 2; pCCA = perihilar cholangiocarcinoma; PDX = patient-derived xenograft; piRNAs = PIWI-interacting RNAs; PIWI = P-element-induced wimpy testis; PSC = primary sclerosing cholangitis; RNA-seq = RNA-sequencing; ROC = receiver operating characteristic; TAA = thioacetamide; TCGA = The Cancer Genome Atlas; WT = wild-type.
References
- Banales, J.M.; Marin, J.J.G.; Lamarca, A.; Rodrigues, P.M.; Khan, S.A.; Roberts, L.R.; Cardinale, V.; Carpino, G.; Andersen, J.B.; Braconi, C.; et al. Cholangiocarcinoma 2020: The next horizon in mechanisms and management. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 577–588. [Google Scholar] [CrossRef]
- Florio, A.A.; Ferlay, J.; Znaor, A.; Ruggieri, D.; Alvarez, C.S.; Laversanne, M.; Bray, F.; McGlynn, K.A.; Petrick, J.L. Global trends in intrahepatic and extrahepatic cholangiocarcinoma incidence from 1993 to 2012. Cancer 2020, 126, 2666–2678. [Google Scholar] [CrossRef]
- Banales, J.M.; Cardinale, V.; Carpino, G.; Marzioni, M.; Andersen, J.B.; Invernizzi, P.; Lind, G.E.; Folseraas, T.; Forbes, S.J.; Fouassier, L.; et al. Cholangiocarcinoma: Current knowledge and future perspectives consensus statement from the European Network for the Study of Cholangiocarcinoma (ENS-CCA). Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 261–280. [Google Scholar] [CrossRef]
- Cambridge, W.A.; Fairfield, C.; Powell, J.J.; Harrison, E.M.; Søreide, K.; Wigmore, S.J.; Guest, R.V. Meta-analysis and Meta-regression of Survival After Liver Transplantation for Unresectable Perihilar Cholangiocarcinoma. Ann. Surg. 2021, 273, 240–250. [Google Scholar] [CrossRef]
- Sato, K.; Glaser, S.; Alvaro, D.; Meng, F.; Francis, H.; Alpini, G. Cholangiocarcinoma: Novel therapeutic targets. Expert Opin. Ther. Targets 2020, 24, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Alsaleh, M.; Leftley, Z.; Barbera, T.A.; Sithithaworn, P.; Khuntikeo, N.; Loilome, W.; Yongvanit, P.; Cox, I.J.; Chamodol, N.; Syms, R.R.; et al. Cholangiocarcinoma: A guide for the nonspecialist. Int. J. Gen. Med. 2018, 12, 13–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.-Z.; Yang, L.-X.; Zheng, B.; Dong, P.-P.; Liu, X.-Y.; Wang, Z.-C.; Zhou, J.; Fan, J.; Wang, X.-Y.; Gao, Q. CK7/CK19 index: A potential prognostic factor for postoperative intrahepatic cholangiocarcinoma patients. J. Surg. Oncol. 2018, 117, 1531–1539. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.K.; Karlsen, T.H.; Folseraas, T. Cholangiocytes in the pathogenesis of primary sclerosing cholangitis and development of cholangiocarcinoma. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2018, 1864, 1390–1400. [Google Scholar] [CrossRef]
- Yamane, H.; Abe, T.; Amano, H.; Kobayashi, T.; Hanada, K.; Yonehara, S.; Ohdan, H.; Nakahara, M.; Noriyuki, T. A case of cholangiolocellular carcinoma featuring intratumoral hepatic artery penetration: A case report. Int. J. Surg. Case Rep. 2017, 35, 77–81. [Google Scholar] [CrossRef]
- Balitzer, D.; Joseph, N.M.; Ferrell, L.; Shafizadeh, N.; Jain, D.; Zhang, X.; Yeh, M.; Di Tommaso, L.; Kakar, S. Immunohistochemical and molecular features of cholangiolocellular carcinoma are similar to well-differentiated intrahepatic cholangiocarcinoma. Mod. Pathol. 2019, 32, 1486–1494. [Google Scholar] [CrossRef]
- Fan, B.; Malato, Y.; Calvisi, D.F.; Naqvi, S.; Razumilava, N.; Ribback, S.; Gores, G.J.; Dombrowski, F.; Evert, M.; Chen, X.; et al. Cholangiocarcinomas can originate from hepatocytes in mice. J. Clin. Investig. 2012, 122, 2911–2915. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Dong, M.; Xu, Z.; Song, X.; Zhang, S.; Qiao, Y.; Che, L.; Gordan, J.; Hu, K.; Liu, Y.; et al. Notch2 controls hepatocyte-derived cholangiocarcinoma formation in mice. Oncogene 2018, 37, 3229–3242. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, X.-P.; Zhang, W.; Dong, H.-H.; Xiang, S.; Zhang, B.-X. Combined hepatocellular cholangiocarcinoma originating from hepatic progenitor cells: Immunohistochemical and double-fluorescence immunostaining evidence. Histopathology 2007, 52, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Brunt, E.; Aishima, S.; Clavien, P.-A.; Fowler, K.; Goodman, Z.; Gores, G.; Gouw, A.; Kagen, A.; Klimstra, D.; Komuta, M.; et al. cHCC-CCA: Consensus terminology for primary liver carcinomas with both hepatocytic and cholangiocytic differentation. Hepatology 2018, 68, 113–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuo, J.-Y.; Lu, D.; Tan, W.-Y.; Zheng, S.-S.; Shen, Y.-Q.; Xu, X. CK19-positive Hepatocellular Carcinoma is a Characteristic Subtype. J. Cancer 2020, 11, 5069–5077. [Google Scholar] [CrossRef]
- Beaufrère, A.; Calderaro, J.; Paradis, V. Combined hepatocellular-cholangiocarcinoma: An update. J. Hepatol. 2021, 74, 1212–1224. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Marzioni, M.; Meng, F.; Francis, H.; Glaser, S.; Alpini, G. Ductular Reaction in Liver Diseases: Pathological Mechanisms and Translational Significances. Hepatology 2019, 69, 420–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moeini, A.; Haber, P.K.; Sia, D. Cell of origin in biliary tract cancers and clinical implications. JHEP Rep. 2021, 3, 100226. [Google Scholar] [CrossRef]
- Zou, S.; Li, J.; Zhou, H.; Frech, C.; Jiang, X.; Chu, J.S.C.; Zhao, X.; Li, Y.; Li, Q.; Wang, H.; et al. Mutational landscape of intrahepatic cholangiocarcinoma. Nat. Commun. 2014, 5, 5696. [Google Scholar] [CrossRef]
- Jusakul, A.; Cutcutache, I.; Yong, C.H.; Lim, J.Q.; Ni Huang, M.; Padmanabhan, N.; Nellore, V.; Kongpetch, S.; Ng, A.W.T.; Ng, L.M.; et al. Whole-Genome and Epigenomic Landscapes of Etiologically Distinct Subtypes of Cholangiocarcinoma. Cancer Discov. 2017, 7, 1116–1135. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Cao, Y.; Yang, X.; Li, G.; Shi, Y.; Wang, D.; Long, J.; Song, Y.; Mao, J.; Xie, F.; et al. Mutational spectrum and precision oncology for biliary tract carcinoma. Theranostics 2021, 11, 4585–4598. [Google Scholar] [CrossRef] [PubMed]
- Simile, M.M.; Bagella, P.; Vidili, G.; Spanu, A.; Manetti, R.; Seddaiu, M.A.; Babudieri, S.; Madeddu, G.; Serra, P.A.; Altana, M.; et al. Targeted Therapies in Cholangiocarcinoma: Emerging Evidence from Clinical Trials. Medicina 2019, 55, 42. [Google Scholar] [CrossRef] [Green Version]
- Wardell, C.P.; Fujita, M.; Yamada, T.; Simbolo, M.; Fassan, M.; Karlic, R.; Polak, P.; Kim, J.; Hatanaka, Y.; Maejima, K.; et al. Genomic characterization of biliary tract cancers identifies driver genes and predisposing mutations. J. Hepatol. 2018, 68, 959–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Cai, Z.; Dong, X.; Zhao, J.; Lin, S.; Hu, X.; Liu, F.-E.; Liu, X.; Zhang, H. Genomic and Transcriptomic Landscape of Tumor Clonal Evolution in Cholangiocarcinoma. Front. Genet. 2020, 11. [Google Scholar] [CrossRef]
- Zhang, R.; Li, Q.; Fu, J.; Jin, Z.; Su, J.; Zhang, J.; Chen, C.; Geng, Z.; Zhang, D. Comprehensive analysis of genomic mutation signature and tumor mutation burden for prognosis of intrahepatic cholangiocarcinoma. BMC Cancer 2021, 21, 112. [Google Scholar] [CrossRef]
- Wada, Y.; Shimada, M.; Yamamura, K.; Toshima, T.; Banwait, J.K.; Morine, Y.; Ikemoto, T.; Saito, Y.; Baba, H.; Mori, M.; et al. A Transcriptomic Signature for Risk-Stratification and Recurrence Prediction in Intrahepatic Cholangiocarcinoma. Hepatology 2021. [Google Scholar] [CrossRef]
- Xue, R.; Chen, L.; Zhang, C.; Fujita, M.; Li, R.; Yan, S.-M.; Ong, C.K.; Liao, X.; Gao, Q.; Sasagawa, S.; et al. Genomic and Transcriptomic Profiling of Combined Hepatocellular and Intrahepatic Cholangiocarcinoma Reveals Distinct Molecular Subtypes. Cancer Cell 2019, 35, 932–947.e8. [Google Scholar] [CrossRef]
- Goeppert, B.; Konermann, C.; Schmidt, C.R.; Bogatyrova, O.; Geiselhart, L.; Ernst, C.; Gu, L.; Becker, N.; Zucknick, M.; Mehrabi, A.; et al. Global alterations of DNA methylation in cholangiocarcinoma target the Wnt signaling pathway. Hepatology 2014, 59, 544–554. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, B.; Meng, D.; Ge, C. Comprehensive analysis of DNA methylation and gene expression profiles in cholangiocarcinoma. Cancer Cell Int. 2019, 19, 352. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Liu, T.; Liao, J.; Zhang, L.; Zhou, M.; Xu, W.; He, Y.; Cai, G.; Jin, G.; Song, J.; et al. Development and validation of a CpG island methylator phenotype-related prognostic signature for cholangiocarcinoma. J. Cell. Physiol. 2021, 236, 3143–3156. [Google Scholar] [CrossRef]
- Mishra, N.K.; Niu, M.; Southekal, S.; Bajpai, P.; Elkholy, A.; Manne, U.; Guda, C. Identification of Prognostic Markers in Cholangiocarcinoma Using Altered DNA Methylation and Gene Expression Profiles. Front. Genet. 2020, 11, 522125. [Google Scholar] [CrossRef] [PubMed]
- Cadamuro, M.; Brivio, S.; Stecca, T.; Kaffe, E.; Mariotti, V.; Milani, C.; Fiorotto, R.; Spirli, C.; Strazzabosco, M.; Fabris, L. Animal models of cholangiocarcinoma: What they teach us about the human disease. Clin. Res. Hepatol. Gastroenterol. 2018, 42, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Loeuillard, E.; Fischbach, S.R.; Gores, G.J.; Rizvi, S. Animal models of cholangiocarcinoma. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2019, 1865, 982–992. [Google Scholar] [CrossRef]
- Mohr, R.; Özdirik, B.; Knorr, J.; Wree, A.; Demir, M.; Tacke, F.; Roderburg, C. In Vivo Models for Cholangiocarcinoma—What Can We Learn for Human Disease? Int. J. Mol. Sci. 2020, 21, 4993. [Google Scholar] [CrossRef]
- Yeh, C.-N.; Maitra, A.; Lee, K.-F.; Jan, Y.-Y.; Chen, M.-F. Thioacetamide-induced intestinal-type cholangiocarcinoma in rat: An animal model recapitulating the multi-stage progression of human cholangiocarcinoma. Carcinogenesis 2003, 25, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, T.W.; Peng, J.; Tang, X.; Ko, K.S.; Xia, M.; Aller, M. Ángeles A Mouse Model of Cholestasis-Associated Cholangiocarcinoma and Transcription Factors Involved in Progression. Gastroenterology 2011, 141, 378–388.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Dell, M.R.; Huang, J.L.; Whitney-Miller, C.L.; Deshpande, V.; Rothberg, P.G.; Grose, V.; Rossi, R.M.; Zhu, A.X.; Land, H.; Bardeesy, N.; et al. KrasG12D and p53 Mutation Cause Primary Intrahepatic Cholangiocarcinoma. Cancer Res. 2012, 72, 1557–1567. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Kobayashi, S.; Qiao, W.; Li, C.; Xiao, C.; Radaeva, S.; Stiles, B.; Wang, R.-H.; Ohara, N.; Yoshino, T.; et al. Induction of intrahepatic cholangiocellular carcinoma by liver-specific disruption ofSmad4 andPten in mice. J. Clin. Investig. 2006, 116, 1843–1852. [Google Scholar] [CrossRef]
- Chen, W.-T.; Tseng, C.-C.; Pfaffenbach, K.; Kanel, G.; Luo, B.; Stiles, B.L.; Lee, A.S. Liver-specific knockout of GRP94 in mice disrupts cell adhesion, activates liver progenitor cells, and accelerates liver tumorigenesis. Hepatology 2013, 59, 947–957. [Google Scholar] [CrossRef] [Green Version]
- Newberry, E.P.; Xie, Y.; Lodeiro, C.; Solis, R.; Moritz, W.; Kennedy, S.; Barron, L.; Onufer, E.; Alpini, G.; Zhou, T.; et al. Hepatocyte and stellate cell deletion of liver fatty acid binding protein reveals distinct roles in fibrogenic injury. FASEB J. 2018, 33, 4610–4625. [Google Scholar] [CrossRef]
- Adachi, T.; Adachi, T.; Nakagaki, T.; Ono, S.; Hidaka, M.; Ito, S.; Kanetaka, K.; Takatsuki, M.; Nishida, N.; Eguchi, S. Difference in driver gene expression patterns between perihilar and peripheral intrahepatic cholangiocarcinoma in an experimental mouse model. J. Hepato-Biliary-Pancreat. Sci. 2020, 27, 477–486. [Google Scholar] [CrossRef]
- Lesaffer, B.; Verboven, E.; Van Huffel, L.; Moya, I.M.; Van Grunsven, L.A.; Leclercq, I.A.; Lemaigre, F.P.; Halder, G. Comparison of the Opn-CreER and Ck19-CreER Drivers in Bile Ducts of Normal and Injured Mouse Livers. Cells 2019, 8, 380. [Google Scholar] [CrossRef] [Green Version]
- Di-Luoffo, M.; Pirenne, S.; Saandi, T.; Loriot, A.; Gérard, C.; Dauguet, N.; Manzano-Núñez, F.; Carvalhais, N.A.S.; Lamoline, F.; Cordi, S.; et al. A novel mouse model of cholangiocarcinoma uncovers a role for Tensin-4 in tumor progression. Hepatology 2021. [Google Scholar] [CrossRef]
- Nakagawa, H.; Hikiba, Y.; Hirata, Y.; Font-Burgada, J.; Sakamoto, K.; Hayakawa, Y.; Taniguchi, K.; Umemura, A.; Kinoshita, H.; Sakitani, K.; et al. Loss of liver E-cadherin induces sclerosing cholangitis and promotes carcinogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, 1090–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, S.; Hikiba, Y.; Fujiwara, H.; Ikenoue, T.; Sue, S.; Sugimori, M.; Matsubayashi, M.; Kaneko, H.; Irie, K.; Sasaki, T.; et al. NAFLD exacerbates cholangitis and promotes cholangiocellular carcinoma in mice. Cancer Sci. 2021, 112, 1471–1480. [Google Scholar] [CrossRef] [PubMed]
- Ikenaga, N.; Liu, S.B.; Sverdlov, D.Y.; Yoshida, S.; Nasser, I.; Ke, Q.; Kang, P.M.; Popov, Y. A new Mdr2-/- mouse model of sclerosing cholangitis with rapid fibrosis progression, early-onset portal hypertension, and liver cancer. Am. J. Pathol. 2015, 185, 325–334. [Google Scholar] [CrossRef]
- Cavalloni, G.; Peraldo-Neia, C.; Sassi, F.; Chiorino, G.; Sarotto, I.; Aglietta, M.; Leone, F. Establishment of a patient-derived intrahepatic cholangiocarcinoma xenograft model with KRAS mutation. BMC Cancer 2016, 16, 90. [Google Scholar] [CrossRef] [Green Version]
- Vaeteewoottacharn, K.; Pairojkul, C.; Kariya, R.; Muisuk, K.; Imtawil, K.; Chamgramol, Y.; Bhudhisawasdi, V.; Khuntikeo, N.; Pugkhem, A.; Saeseow, O.-T.; et al. Establishment of Highly Transplantable Cholangiocarcinoma Cell Lines from a Patient-Derived Xenograft Mouse Model. Cells 2019, 8, 496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiting, J.L.; Murphy, S.J.; Bergquist, J.R.; Hernandez, M.C.; Ivanics, T.; Abdelrahman, A.M.; Yang, L.; Lynch, I.; Smadbeck, J.B.; Cleary, S.P.; et al. Biliary tract cancer patient-derived xenografts: Surgeon impact on individualized medicine. JHEP Rep. 2020, 2, 100068. [Google Scholar] [CrossRef] [Green Version]
- Lau, D.K.; Mouradov, D.; Wasenang, W.; Luk, I.Y.; Scott, C.M.; Williams, D.S.; Yeung, Y.H.; Limpaiboon, T.; Iatropoulos, G.F.; Jenkins, L.J.; et al. Genomic Profiling of Biliary Tract Cancer Cell Lines Reveals Molecular Subtypes and Actionable Drug Targets. iScience 2019, 21, 624–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Luo, J.; Dong, X.; Yang, F.; Zhang, M.; Zhao, J.; Wang, Q.; Zhou, F.; Sun, J.; Yang, X. Establishment and Characterization of Two Novel Cholangiocarcinoma Cell Lines. Ann. Surg. Oncol. 2019, 26, 4134–4147. [Google Scholar] [CrossRef]
- Sato, K.; Zhang, W.; Safarikia, S.; Isidan, A.; Chen, A.M.; Li, P.; Francis, H.; Kennedy, L.; Baiocchi, L.; Alvaro, D.; et al. Organoids and spheroids as novel models for studying cholestatic liver injury and cholangiocarcinoma. Hepatology 2020. [Google Scholar] [CrossRef]
- Hof, L.; Moreth, T.; Koch, M.; Liebisch, T.; Kurtz, M.; Tarnick, J.; Lissek, S.M.; Verstegen, M.M.A.; van der Laan, L.J.W.; Huch, M.; et al. Long-term live imaging and multiscale analysis identify heterogeneity and core principles of epithelial organoid morphogenesis. BMC Biol. 2021, 19, 37. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Muramatsu, T.; Kanai, Y.; Ojima, H.; Sukeda, A.; Hiraoka, N.; Arai, E.; Sugiyama, Y.; Matsuzaki, J.; Uchida, R.; et al. Establishment of Patient-Derived Organoids and Drug Screening for Biliary Tract Carcinoma. Cell Rep. 2019, 27, 1265–1276.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, S.; He, J.; Zhao, Y.; Liu, T.; Xie, F.; Yang, H.; Mao, Y.; Pang, Y.; Sun, W. Bioprinting of patient-derived in vitro intrahepatic cholangiocarcinoma tumor model: Establishment, evaluation and anti-cancer drug testing. Biofabrication 2020, 12, 045014. [Google Scholar] [CrossRef]
- Yoshikawa, N.; Saito, Y.; Manabe, H.; Nakaoka, T.; Uchida, R.; Furukawa, R.; Muramatsu, T.; Sugiyama, Y.; Kimura, M.; Saito, H. Glucose Depletion Enhances the Stem Cell Phenotype and Gemcitabine Resistance of Cholangiocarcinoma Organoids through AKT Phosphorylation and Reactive Oxygen Species. Cancers 2019, 11, 1993. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wang, L.; Wang, Y.; Zhang, S.; Zhou, G.; Lieshout, R.; Ma, B.; Liu, J.; Qu, C.; Verstegen, M.M.A.; et al. Mitochondrial Fusion Via OPA1 and MFN1 Supports Liver Tumor Cell Metabolism and Growth. Cells 2020, 9, 121. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Noordam, L.; Ou, X.; Ma, B.; Li, Y.; Das, P.; Shi, S.; Liu, J.; Wang, L.; Li, P.; et al. The biological process of lysine-tRNA charging is therapeutically targetable in liver cancer. Liver Int. 2021, 41, 206–219. [Google Scholar] [CrossRef] [PubMed]
- Kasuga, A.; Semba, T.; Sato, R.; Nobusue, H.; Sugihara, E.; Takaishi, H.; Kanai, T.; Saya, H.; Arima, Y. Oncogenic KRAS–expressing organoids with biliary epithelial stem cell properties give rise to biliary tract cancer in mice. Cancer Sci. 2021, 112, 1822–1838. [Google Scholar] [CrossRef]
- Cristinziano, G.; Porru, M.; Lamberti, D.; Buglioni, S.; Rollo, F.; Amoreo, C.A.; Manni, I.; Giannarelli, D.; Cristofoletti, C.; Russo, G.; et al. FGFR2 fusion proteins drive oncogenic transformation of mouse liver organoids towards cholangiocarcinoma. J. Hepatol. 2021. [Google Scholar] [CrossRef]
- Goyal, L.; Shi, L.; Liu, L.Y.; De La Cruz, F.F.; Lennerz, J.K.; Raghavan, S.; Leschiner, I.; Elagina, L.; Siravegna, G.; Ng, R.W.; et al. TAS-120 Overcomes Resistance to ATP-Competitive FGFR Inhibitors in Patients with FGFR2 Fusion–Positive Intrahepatic Cholangiocarcinoma. Cancer Discov. 2019, 9, 1064–1079. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, H.; Takahara, N.; Tateishi, K.; Tanaka, M.; Kanai, S.; Kato, H.; Nakatsuka, T.; Yamamoto, K.; Kogure, H.; Arita, J.; et al. 5-Aminolevulinic acid-mediated photodynamic activity in patient-derived cholangiocarcinoma organoids. Surg. Oncol. 2020, 35, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Macias, R.I.; Banales, J.; Sangro, B.; Muntané, J.; Avila, M.; Lozano, E.; Perugorria, M.J.; Padillo, F.J.; Bujanda, L.; Marin, J.J. The search for novel diagnostic and prognostic biomarkers in cholangiocarcinoma. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2018, 1864, 1468–1477. [Google Scholar] [CrossRef] [PubMed]
- Macias, R.I.R.; Kornek, M.; Rodrigues, P.M.; Paiva, N.A.; Castro, R.E.; Urban, S.; Pereira, S.P.; Cadamuro, M.; Rupp, C.; Loosen, S.H.; et al. Diagnostic and prognostic biomarkers in cholangiocarcinoma. Liver Int. 2019, 39, 108–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, X.; Zhang, C.; Yin, C.; Wang, W.; Yan, X.; Xie, D.; Zheng, X.; Zheng, Q.; Li, M.; Song, Z. High BLM Expression Predicts Poor Clinical Outcome and Contributes to Malignant Progression in Human Cholangiocarcinoma. Front. Oncol. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Long, J.; Xie, F.; Kang, K.; Shi, Y.; Xu, W.; Wu, X.; Lin, J.; Xu, H.; Du, S.; et al. Transcriptomic analysis and identification of prognostic biomarkers in cholangiocarcinoma. Oncol. Rep. 2019, 42, 1833–1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, B.; Shen, S.; Wong, K.; Gong, Z.; Sun, W.; Ni, X.; Wang, J.; Hu, M.; Liu, H.; Liu, H.; et al. Clinical correlation of cadherin-17 marker with advanced tumor stages and poor prognosis of cholangiocarcinoma. J. Surg. Oncol. 2021, 123, 1253–1262. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Lee, H.-S.; Bang, S.-M.; Han, D.-H.; Hwang, H.-K.; Choi, G.-H.; Chung, M.-J.; Kim, S.-U. Serum Dickkopf-1 in Combined with CA 19-9 as a Biomarker of Intrahepatic Cholangiocarcinoma. Cancers 2021, 13, 1828. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Meng, F.; Glaser, S.; Alpini, G. Exosomes in liver pathology. J. Hepatol. 2016, 65, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Li, X.; Zhu, W.; Wang, Y.; Zhao, D.; Wang, X.; Gurley, E.C.; Liang, G.; Chen, W.; Lai, G.; et al. Cholangiocyte-Derived Exosomal Long Noncoding RNA H19 Promotes Hepatic Stellate Cell Activation and Cholestatic Liver Fibrosis. Hepatology 2019, 70, 1317–1335. [Google Scholar] [CrossRef]
- Arunsan, P.; Chaidee, A.; Cochran, C.J.; Mann, V.H.; Tanno, T.; Kumkhaek, C.; Smout, M.; Karinshak, S.E.; Rodpai, R.; Sotillo, J.; et al. Liver fluke granulin promotes extracellular vesicle-mediated crosstalk and cellular microenvironment conducive to cholangiocarcinoma. Neoplasia 2020, 22, 203–216. [Google Scholar] [CrossRef]
- Sato, K.; Meng, F.; Venter, J.; Giang, T.; Glaser, S.; Alpini, G. The role of the secretin/secretin receptor axis in inflammatory cholangiocyte communication via extracellular vesicles. Sci. Rep. 2017, 7, 11183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arbelaiz, A.; Azkargorta, M.; Krawczyk, M.; Santos-Laso, A.; Lapitz, A.; Perugorria, M.J.; Erice, O.; Gonzalez, E.; Jimenez-Agüero, R.; La Casta, A.; et al. Serum extracellular vesicles contain protein biomarkers for primary sclerosing cholangitis and cholangiocarcinoma. Hepatology 2017, 66, 1125–1143. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Chen, G.; Xia, Q.; Shao, S.; Fang, H. Exosomal miR-200 family as serum biomarkers for early detection and prognostic prediction of cholangiocarcinoma. Int. J. Clin. Exp. Pathol 2019, 12, 3870–3876. [Google Scholar]
- Ikeda, C.; Haga, H.; Makino, N.; Inuzuka, T.; Kurimoto, A.; Ueda, T.; Matsuda, A.; Kakizaki, Y.; Ishizawa, T.; Kobayashi, T.; et al. Utility of Claudin-3 in extracellular vesicles from human bile as biomarkers of cholangiocarcinoma. Sci. Rep. 2021, 11, 1195. [Google Scholar] [CrossRef]
- Lapitz, A.; Arbelaiz, A.; O’Rourke, C.J.; Lavin, J.L.; La Casta, A.; Ibarra, C.; Jimeno, J.P.; Santos-Laso, A.; Izquierdo-Sanchez, L.; Krawczyk, M.; et al. Patients with Cholangiocarcinoma Present Specific RNA Profiles in Serum and Urine Extracellular Vesicles Mirroring the Tumor Expression: Novel Liquid Biopsy Biomarkers for Disease Diagnosis. Cells 2020, 9, 721. [Google Scholar] [CrossRef] [Green Version]
- Chu, K.-J.; Ma, Y.-S.; Jiang, X.-H.; Wu, T.-M.; Wu, Z.-J.; Li, Z.-Z.; Wang, J.-H.; Gao, Q.-X.; Yi, B.; Shi, Y.; et al. Whole-Transcriptome Sequencing Identifies Key Differentially Expressed mRNAs, miRNAs, lncRNAs, and circRNAs Associated with CHOL. Mol. Ther. Nucleic Acids 2020, 21, 592–603. [Google Scholar] [CrossRef]
- Hu, J.; Wang, Y.-N.; Song, D.-J.; Tan, J.-P.; Cao, Y.; Fan, J.; Wang, Z.; Zhou, J. A High-Accuracy Model Based on Plasma miRNAs Diagnoses Intrahepatic Cholangiocarcinoma: A Single Center with 1001 Samples. Diagnostics 2021, 11, 610. [Google Scholar] [CrossRef] [PubMed]
- Limb, C.; Liu, D.; Veno, M.; Rees, E.; Krell, J.; Bagwan, I.; Giovannetti, E.; Pandha, H.; Strobel, O.; Rockall, T.; et al. The Role of Circular RNAs in Pancreatic Ductal Adenocarcinoma and Biliary-Tract Cancers. Cancers 2020, 12, 3250. [Google Scholar] [CrossRef]
- Laschos, K.; Lampropoulou, D.I.; Aravantinos, G.; Piperis, M.; Filippou, D.; Theodoropoulos, G.; Gazouli, M. Exosomal noncoding RNAs in cholangiocarcinoma: Laboratory noise or hope? World J. Gastrointest. Surg. 2020, 12, 407–424. [Google Scholar] [CrossRef]
- Sato, K.; Glaser, S.; Francis, H.; Alpini, G. Concise Review: Functional Roles and Therapeutic Potentials of Long Non-coding RNAs in Cholangiopathies. Front. Med. 2020, 7. [Google Scholar] [CrossRef]
- Salati, M.; Braconi, C. Noncoding RNA in Cholangiocarcinoma. Semin. Liver Dis. 2019, 39, 013–025. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Jiao, D.; Liu, Z.; Chen, J.; Zhou, X.; Li, Z.; Li, J.; Han, X. Novel miRNA Predicts Survival and Prognosis of Cholangiocarcinoma Based on RNA-seq Data and In Vitro Experiments. BioMed Res. Int. 2020, 2020, 5976127. [Google Scholar] [CrossRef]
- Cao, K.; Li, B.; Zhang, Y.-W.; Song, H.; Chen, Y.-G.; Gong, Y.-J.; Li, H.-Y.; Zuo, S. miR-29b restrains cholangiocarcinoma progression by relieving DNMT3B-mediated repression of CDKN2B expression. Aging 2021, 13, 6055–6065. [Google Scholar] [CrossRef]
- Salem, P.E.S.; Ghazala, R.A.; El Gendi, A.M.; Emara, D.M.; Ahmed, N.M. The association between circulating MicroRNA-150 level and cholangiocarcinoma. J. Clin. Lab. Anal. 2020, 34, e23397. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Yu, G.; Liang, J.; Fan, P.; Dong, K.; Zhang, B.; Chen, X.; Zhu, H.; Chu, L. miR-144-5p and miR-451a Inhibit the Growth of Cholangiocarcinoma Cells Through Decreasing the Expression of ST8SIA. Front. Oncol. 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Wang, G.; Wang, B. MicroRNA-1182 and let-7a exert synergistic inhibition on invasion, migration and autophagy of cholangiocarcinoma cells through down-regulation of NUAK. Cancer Cell Int. 2021, 21, 161. [Google Scholar] [CrossRef]
- Li, G.; Liu, T.; Zhang, B.; Chen, W.; Ding, Z. Genome-wide identification of a competing endogenous RNA network in cholangiocarcinoma. J. Cell. Biochem. 2019, 120, 18995–19003. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Wang, Y.; Zhang, S.; Li, J.; Yu, Z.; Ding, X.; Ye, L.; Gong, P.; Zhu, Q.; Li, J.; et al. A novel five-lncRNA signature panel improves high-risk survival prediction in patients with cholangiocarcinoma. Aging 2021, 13, 2959–2981. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Huang, L.; Wang, W.; Guan, C.; Zhao, Y.; Liu, L.; Jiang, X. Long Non-coding RNA FOXD2-AS1 Promotes Proliferation, Migration, and Invasion in Cholangiocarcinoma Through Regulating miR-760/E2F3 Axis. Dig. Dis. Sci. 2021, 1–13. [Google Scholar] [CrossRef]
- Li, Q.; Fu, L.; Han, L.; Li, S.; Zhang, Y.; Wang, J. Long Noncoding RNA GAS5 Accelerates Cholangiocarcinoma Progression by Regulating hsa-miR-1297. Cancer Manag. Res. 2021, 13, 2745–2753. [Google Scholar] [CrossRef]
- Zhu, H.; Zhai, B.; He, C.; Li, Z.; Gao, H.; Niu, Z.; Jiang, X.; Lu, J.; Sun, X. LncRNA TTN-AS1 promotes the progression of cholangiocarcinoma via the miR-320a/neuropilin-1 axis. Cell Death Dis. 2020, 11, 637. [Google Scholar] [CrossRef]
- Xia, L.; Chen, X.; Yang, J.; Zhu, S.; Zhang, L.; Yin, Q.; Hong, Y.; Chen, H.; Chen, G.; Li, H. Long Non-Coding RNA-PAICC Promotes the Tumorigenesis of Human Intrahepatic Cholangiocarcinoma by Increasing YAP1 Transcription. Front. Oncol. 2021, 10. [Google Scholar] [CrossRef]
- Wu, T.; Lei, M.-S.; Gao, X.-Z.; Xiong, T.-G.; Yang, K.; Gong, Q.; Tang, R.; Tian, Y.-P.; Fu, X.-H. lncRNA SNHG16 Mediates Cell Proliferation and Apoptosis in Cholangiocarcinoma by Directly Targeting miR-146a-5p/GATA6 Axis. Biochem. Genet. 2021, 1–15. [Google Scholar] [CrossRef]
- Zhao, W.; Zhao, J.; Guo, X.; Feng, Y.; Zhang, B.; Tian, L. LncRNA MT1JP plays a protective role in intrahepatic cholangiocarcinoma by regulating miR-18a-5p/FBP1 axis. BMC Cancer 2021, 21, 142. [Google Scholar] [CrossRef]
- Peng, L.; Liu, Y.-H.; Nie, S.; Gao, M. LncRNA CASC2 inhibits cell proliferation, metastasis and EMT through miR-18a/SOCS5 axis in cholangiocarcinoma. Eur. Rev. Med. Pharm. Sci. 2021, 25, 1767. [Google Scholar]
- Lu, M.; Qin, X.; Zhou, Y.; Li, G.; Liu, Z.; Yue, H.; Geng, X. LncRNA HOTAIR suppresses cell apoptosis, autophagy and induces cell proliferation in cholangiocarcinoma by modulating the miR-204-5p/HMGB1 axis. Biomed. Pharmacother. 2020, 130, 110566. [Google Scholar] [CrossRef]
- Xu, Y.; Gao, P.; Wang, Z.; Su, Z.; Liao, G.; Han, Y.; Cui, Y.; Yao, Y.; Zhong, X. Circ-LAMP1 contributes to the growth and metastasis of cholangiocarcinoma via miR-556-5p and miR-567 mediated YY1 activation. J. Cell. Mol. Med. 2021, 25, 3226–3238. [Google Scholar] [CrossRef]
- Xu, Y.; Leng, K.; Yao, Y.; Kang, P.; Liao, G.; Han, Y.; Shi, G.; Ji, D.; Huang, P.; Zheng, W.; et al. A Circular RNA, Cholangiocarcinoma-Associated Circular RNA 1, Contributes to Cholangiocarcinoma Progression, Induces Angiogenesis, and Disrupts Vascular Endothelial Barriers. Hepatology 2021, 73, 1419–1435. [Google Scholar] [CrossRef]
- Su, Y.; Yu, T.; Wang, Y.; Huang, X.; Wei, X. Circular RNA circDNM3OS Functions as a miR-145-5p Sponge to Accelerate Cholangiocarcinoma Growth and Glutamine Metabolism by Upregulating MORC. OncoTargets Ther. 2021, 14, 1117–1129. [Google Scholar] [CrossRef]
- You, J.; Wang, X. Circ_HIPK3 Knockdown Inhibits Cell Proliferation, Migration and Invasion of Cholangiocarcinoma Partly via Mediating the miR-148a-3p/ULK1 Pathway. Cancer Manag. Res. 2021, 13, 3827–3839. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hu, Y.; Lv, X.; Li, B.; Gu, D.; Li, Y.; Sun, Y.; Su, Y. Circ-0000284 arouses malignant phenotype of cholangiocarcinoma cells and regulates the biological functions of peripheral cells through cellular communication. Clin. Sci. 2019, 133, 1935–1953. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yao, Y.; Liu, Y.; Wang, Z.; Hu, Z.; Su, Z.; Li, C.; Wang, H.; Jiang, X.; Kang, P.; et al. Elevation of circular RNA circ_0005230 facilitates cell growth and metastasis via sponging miR-1238 and miR-1299 in cholangiocarcinoma. Aging 2019, 11, 1907–1917. [Google Scholar] [CrossRef]
- Lu, Q.; Fang, T. Circular RNA SMARCA5 correlates with favorable clinical tumor features and prognosis, and increases chemotherapy sensitivity in intrahepatic cholangiocarcinoma. J. Clin. Lab. Anal. 2020, 34, e23138. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, X.; Zhang, Z.; Liu, Z.; Zhu, J.; Lyu, S.; Li, L.; Lang, R.; He, Q. Comprehensive circular RNA expression profiling constructs a ceRNA network and identifies hsa_circ_0000673 as a novel oncogene in distal cholangiocarcinoma. Aging 2020, 12, 23251–23274. [Google Scholar] [CrossRef]
- Weng, W.; Li, H.; Goel, A. Piwi-interacting RNAs (piRNAs) and cancer: Emerging biological concepts and potential clinical implications. Biochim. Biophys. Acta (BBA) Bioenerg. 2019, 1871, 160–169. [Google Scholar] [CrossRef]
- Ge, L.; Zhang, N.; Li, D.; Wu, Y.; Wang, H.; Wang, J. Circulating exosomal small RNAs are promising non-invasive diagnostic biomarkers for gastric cancer. J. Cell. Mol. Med. 2020, 24, 14502–14513. [Google Scholar] [CrossRef]
- Gu, X.; Wang, C.; Deng, H.; Qing, C.; Liu, R.; Liu, S.; Xue, X. Exosomal piRNA profiling revealed unique circulating piRNA signatures of cholangiocarcinoma and gallbladder carcinoma. Acta Biochim. Biophys. Sin. 2020, 52, 475–484. [Google Scholar] [CrossRef]
- Sato, K.; Meng, F.; Fava, G.; Glaser, S.; Alpini, G. Functional roles of gut bacteria imbalance in cholangiopathies. Liver Res. 2019, 3, 40–45. [Google Scholar] [CrossRef]
- Jia, X.; Lu, S.; Zeng, Z.; Liu, Q.; Dong, Z.; Chen, Y.; Zhu, Z.; Hong, Z.; Zhang, T.; Du, G.; et al. Characterization of Gut Microbiota, Bile Acid Metabolism, and Cytokines in Intrahepatic Cholangiocarcinoma. Hepatology 2020, 71, 893–906. [Google Scholar] [CrossRef]
- Plieskatt, J.L.; Deenonpoe, R.; Mulvenna, J.; Krause, L.; Sripa, B.; Bethony, J.M.; Brindley, P.J. Infection with the carcinogenic liver fluke Opisthorchis viverrini modifies intestinal and biliary microbiome. FASEB J. 2013, 27, 4572–4584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, S.; Cheng, X.; Zhang, L.; Lu, X.; Chaudhary, S.; Teng, R.; Frederickson, C.; Champion, M.; Zhao, R.; Cheng, L.; et al. Myeloid-derived suppressor cells inhibit T cell activation through nitrating LCK in mouse cancers. Proc. Natl. Acad. Sci. USA 2018, 115, 10094–10099. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.-D.; Hu, J.; Wang, M.; Peng, F.; Tian, R.; Guo, X.-J.; Xie, Y.; Qin, R.-Y. Circulating myeloid-derived suppressor cells in patients with pancreatic cancer. Hepatobiliary Pancreat. Dis. Int. 2016, 15, 099–105. [Google Scholar] [CrossRef]
- Zhang, Q.; Ma, C.; Duan, Y.; Heinrich, B.; Rosato, U.; Diggs, L.P.; Ma, L.; Roy, S.; Fu, Q.; Brown, Z.J.; et al. Gut Microbiome Directs Hepatocytes to Recruit MDSCs and Promote Cholangiocarcinoma. Cancer Discov. 2021, 11, 1248–1267. [Google Scholar] [CrossRef]
- Fabris, L.; Sato, K.; Alpini, G.; Strazzabosco, M. The Tumor Microenvironment in Cholangiocarcinoma Progression. Hepatology 2021, 73, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Sirica, A.E.; Zhang, Z.; Lai, G.-H.; Asano, T.; Shen, X.-N.; Ward, D.J.; Mahatme, A.; DeWitt, J.L. A novel “patient-like” model of cholangiocarcinoma progression based on bile duct inoculation of tumorigenic rat cholangiocyte cell lines. Hepatology 2007, 47, 1178–1190. [Google Scholar] [CrossRef]
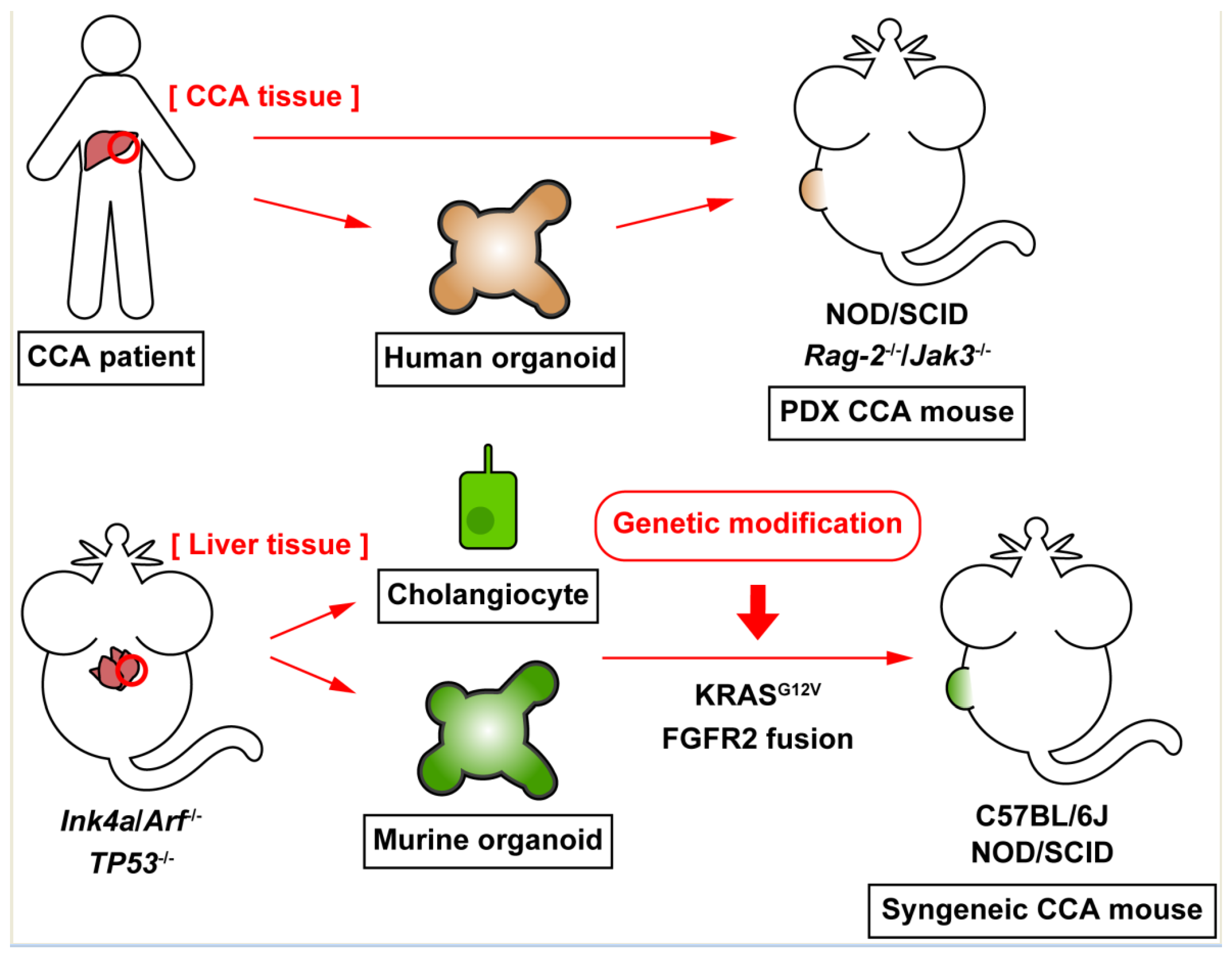
Figure 1.
Current advances in experimental models of cholangiocarcinoma (CCA). CCA tumor tissues excised from patients can be engrafted in immunodeficient mice, such as NOD/SCID mice, to generate patient-derived xenograft (PDX) mouse models. CCA organoids, which are established from CCA tumor tissues, resemble 3D cellular architecture, and maintain functions of CCA tumor cells. Human CCA organoids can be transplanted into mice to generate xenograft animals. Murine cholangiocytes or hepatic organoids, which are derived from knockout mice for tumor-suppressor genes become malignant by genetic modification, such as KRAS activation or FGFR2 fusion protein expression, and these malignant cholangiocytes or organoids establish CCA tumors in WT or immunodeficient mice. These syngeneic CCA mouse models overcome the mismatch of species between CCA tumors and the host animals.
Figure 1.
Current advances in experimental models of cholangiocarcinoma (CCA). CCA tumor tissues excised from patients can be engrafted in immunodeficient mice, such as NOD/SCID mice, to generate patient-derived xenograft (PDX) mouse models. CCA organoids, which are established from CCA tumor tissues, resemble 3D cellular architecture, and maintain functions of CCA tumor cells. Human CCA organoids can be transplanted into mice to generate xenograft animals. Murine cholangiocytes or hepatic organoids, which are derived from knockout mice for tumor-suppressor genes become malignant by genetic modification, such as KRAS activation or FGFR2 fusion protein expression, and these malignant cholangiocytes or organoids establish CCA tumors in WT or immunodeficient mice. These syngeneic CCA mouse models overcome the mismatch of species between CCA tumors and the host animals.


{kind=link}
Table 1.
Selected genetically engineered CCA mouse models.
| Mouse Model | Recombination | Induced Alterations |
|---|---|---|
| KrasG12D mice [37] | Alb-Cre | KRAS activation |
| KrasG12D;p53L/L mice [37] | Alb-Cre | KRAS activation and p53 deletion |
| Smad4Co/CoPtenCo/Co mice [38] | Alb-Cre | Deletion of SMAD4 and PTEN |
| Ptenf/fGrp94f/f mice [39] | Alb-Cre | Deletion of PTEN and GRP94 |
| AKT/YAP Sleeping Beauty [41] | Sleeping Beauty transposon | Activation of AKT and YAP |
| KrasG12D mice with DDC diet [43] | Opn-Cre | KRAS activation and biliary damage |
| KrasG12D/CDH1ΔL mice with high fat diet [45] | Alb-Cre | KRAS activation, deletion of E-cadherin, and non-alcoholic fatty liver disease |
Table 2.
Brief characteristics of current CCA animal models.
| Model | Pros | Cons |
|---|---|---|
| Carcinogen administration (TAA, DEN) | Established, reproducible, and easy procedures Allows to compare early stage and late stage | Long time administration to generate tumors Procedures established mainly for rats Does not mimic human conditions |
| Genetically engineered mouse | Mimics common genetic aberrations found in humans Allows to compare early stage and late stage | Double or triple knockout required to generate tumors Does not mimic CCA development associated with biliary damage and inflammation |
| Combination of genetically engineered mouse and special feeding | Only single or double knockout or mutation required Mimics biliary damage and inflammation by feeding | Limited previous studies Need to evaluate established tumors as CCA Relatively long period required to establish tumors |
| Xenograft mouse | Relatively easy procedures Established methodologies for mice Fast tumor formation | Mismatch species Lack of tumor microenvironment Cannot compare early stage and late stage |
| Patient-derived xenograft mouse | Maintains individual CCA characteristics Allows drug testing or genetic screening personalized for the donor patient | Mismatch species Need to maintain mice with tumors for each donor patient |
| Syngeneic CCA mouse | Matched species Mimics tumor microenvironment | Relatively challenging procedures Does not mimic CCA development associated with biliary damage and inflammation Cannot compare early stage and late stage |
Table 3.
Selected miRNAs identified in recent CCA studies.
| miRNAs | Samples Analyzed | Expression in CCA | Targets | Association with Poor Survival |
|---|---|---|---|---|
| miR-22 [31] | CCA tumor | Downregulated | N/A | High expression |
| miR-551b [31] | CCA tumor | Downregulated | N/A | Low expression |
| miR-200 family [74] | Serum EV | Upregulated | N/A | High levels |
| miR-3913 [83] | CCA tumor | Upregulated | N/A | High expression |
| miR-29b [84] | CCA tumor | Downregulated | DNMT3B | Low expression |
| miR-150 [85] | Serum | Downregulated | N/A | N/A |
| miR-144 [86] | CCA tumor | Downregulated | ST8SIA4 | N/A |
| miR-451a [86] | CCA tumor | Downregulated | ST8SIA4 | N/A |
| miR-1182 [87] | CCA tumor | Downregulated | NUAK1 | N/A |
| let-7a [87] | CCA tumor | Downregulated | NUAK1 | N/A |
Table 4.
Selected lncRNAs identified in recent CCA studies.
| lncRNAs | Samples Analyzed | Expression in CCA | Primary Targets | Secondary Targets |
|---|---|---|---|---|
| FOXD2-AS1 [90] | CCA tissues and cells | Upregulated | miR-760 | E2F3 |
| GAS5 [91] | CCA tissues and cells | Upregulated | miR-1297 | N/A |
| TTN-AS1 [92] | CCA tissues and cells | Upregulated | miR-320a | NRP-1 |
| PAICC [93] | CCA tissues and cells | Upregulated | miR-141-3p, miR-27a-3p | YAP1 |
| SNHG16 [94] | CCA tissues and cells | Upregulated | miR-146a | GATA6 |
| MT1JP [95] | CCA tissues and cells | Downregulated | miR-18a | FBP1 |
| CASC2 [96] | CCA tissues and cells | Downregulated | miR-18a | SOCS5 |
| HOTAIR [97] | CCA tissues and cells | Upregulated | miR-204 | HMGB1 |
Table 5.
Selected circRNAs identified in recent CCA studies.
| circRNA | Samples Analyzed | Expression in CCA | Primary Targets | Secondary Targets |
|---|---|---|---|---|
| circ-LAMP1 [98] | CCA tissues and cells | Upregulated | miR-556, miR-567 | YY1 |
| circ-CCAC1 [99] | CCA tissues and cells, bile EVs | Upregulated | miR-514a | YY1 |
| circ-DNM3OS [100] | CCA tissues and cells | Upregulated | miR-145 | MORC2 |
| circ-HIPK3 [101] | CCA tissues and cells | Upregulated | miR-148a-3p | ULK1 |
| circ-0000284 [102] | CCA tissues and cells | Upregulated | miR-637 | LY6E |
| circ-0005230 [103] | CCA tissues and cells | Upregulated | miR-1238, miR-1299 | N/A |
| circ-SMARCA5 [104] | CCA tissues | Downregulated | N/A | N/A |
| circ-0000673 [105] | CCA tissues | Upregulated | miR-548b-3p | Various genes predicted |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sato, K.; Baiocchi, L.; Kennedy, L.; Zhang, W.; Ekser, B.; Glaser, S.; Francis, H.; Alpini, G. Current Advances in Basic and Translational Research of Cholangiocarcinoma. Cancers 2021, 13, 3307. https://doi.org/10.3390/cancers13133307
AMA Style
Sato K, Baiocchi L, Kennedy L, Zhang W, Ekser B, Glaser S, Francis H, Alpini G. Current Advances in Basic and Translational Research of Cholangiocarcinoma. Cancers. 2021; 13(13):3307. https://doi.org/10.3390/cancers13133307
Chicago/Turabian StyleSato, Keisaku, Leonardo Baiocchi, Lindsey Kennedy, Wenjun Zhang, Burcin Ekser, Shannon Glaser, Heather Francis, and Gianfranco Alpini. 2021. "Current Advances in Basic and Translational Research of Cholangiocarcinoma" Cancers 13, no. 13: 3307. https://doi.org/10.3390/cancers13133307
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.