
Simultaneous Expression of Long Non-Coding RNA FAL1 and Extracellular Matrix Protein 1 Defines Tumour Behaviour in Young Patients with Papillary Thyroid Cancer

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Analysis of lncRNA and mRNA Expression Using Public Databases
2.2. Analysis of Copy Number Alteration
2.3. Prediction and Validation of Putative Transcription Factors
2.4. Statistical Analysis
3. Results
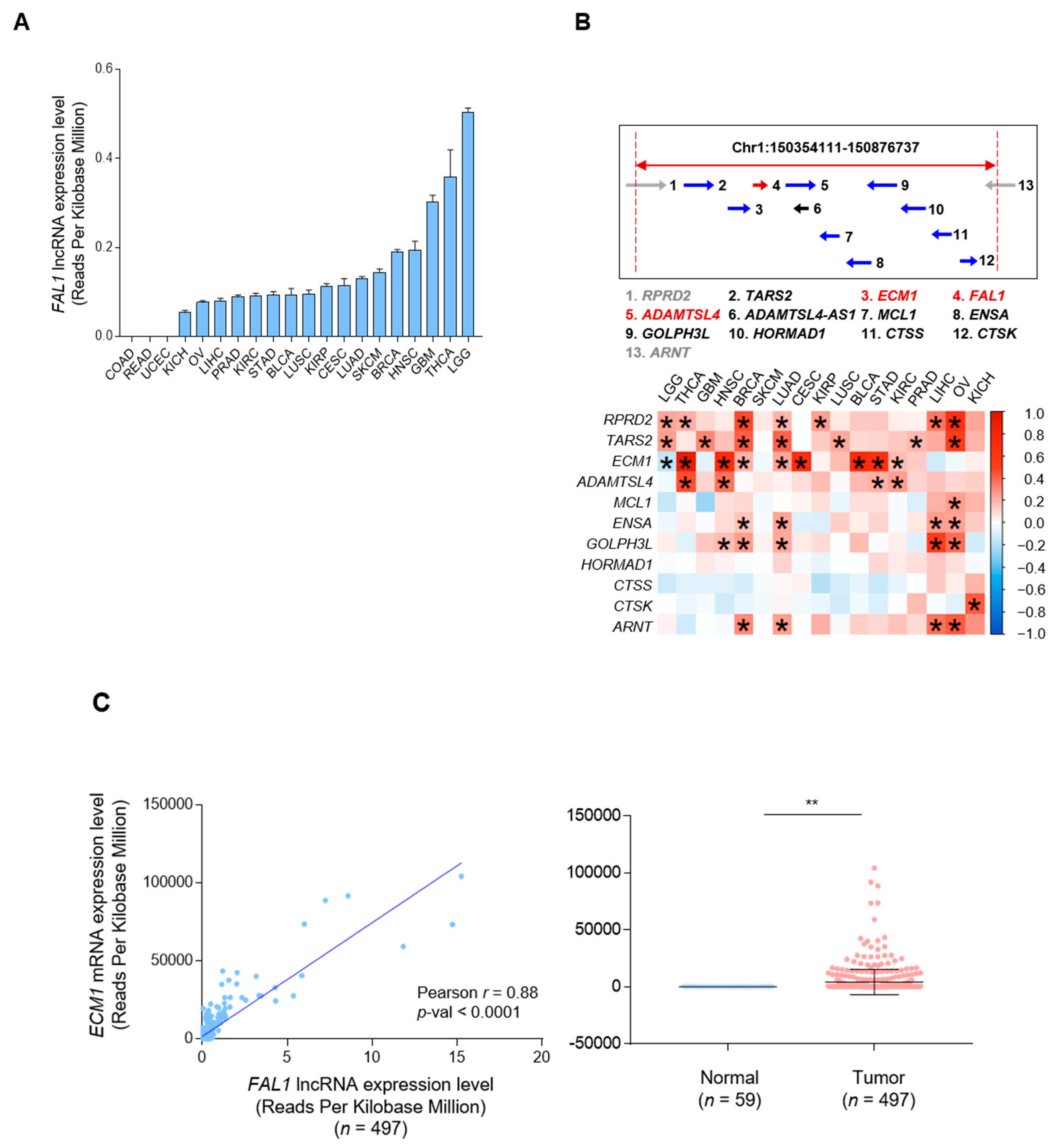
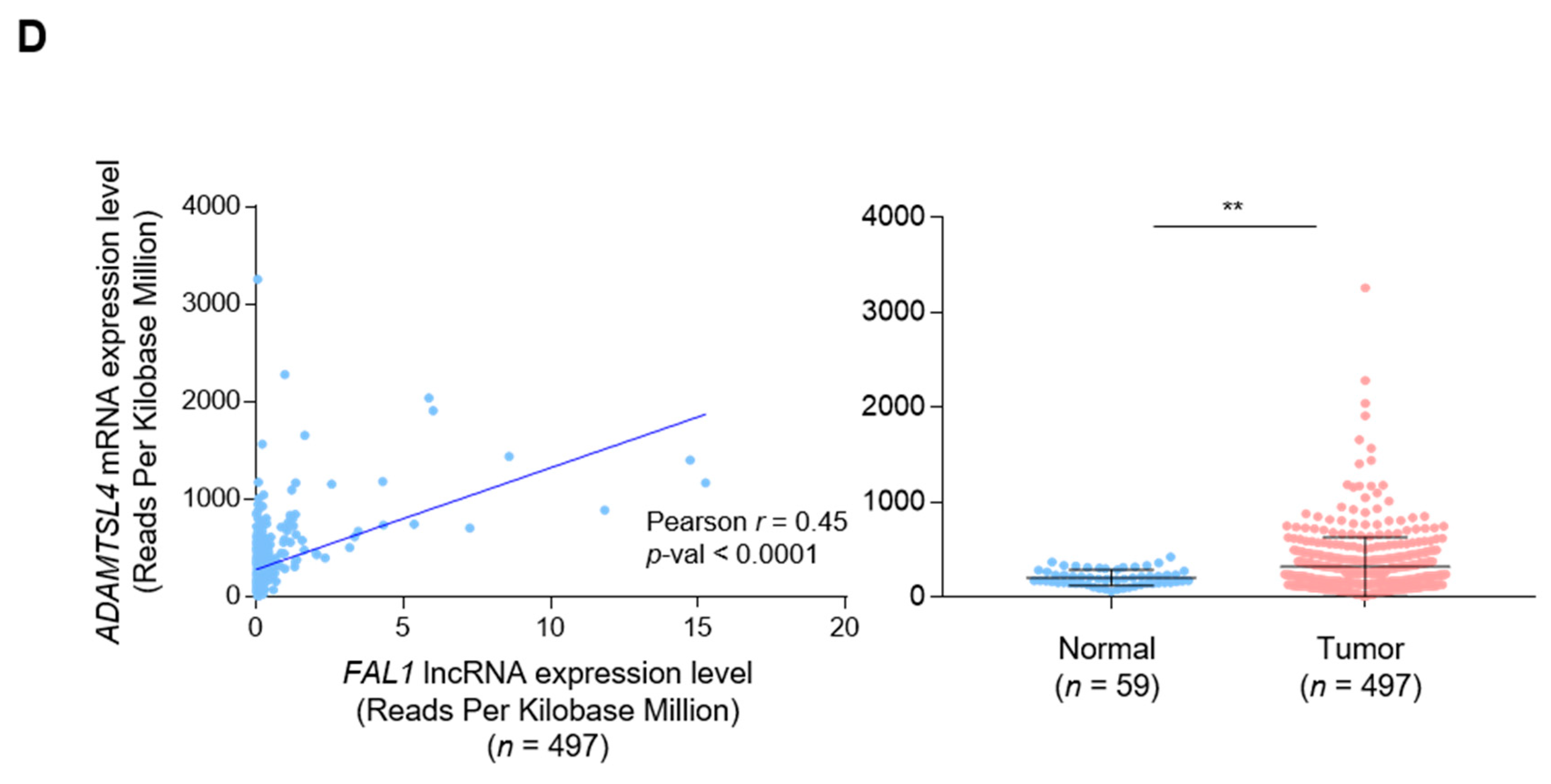
3.1. Positive Correlation between FAL1 and ECM1 Expression in Human Cancers
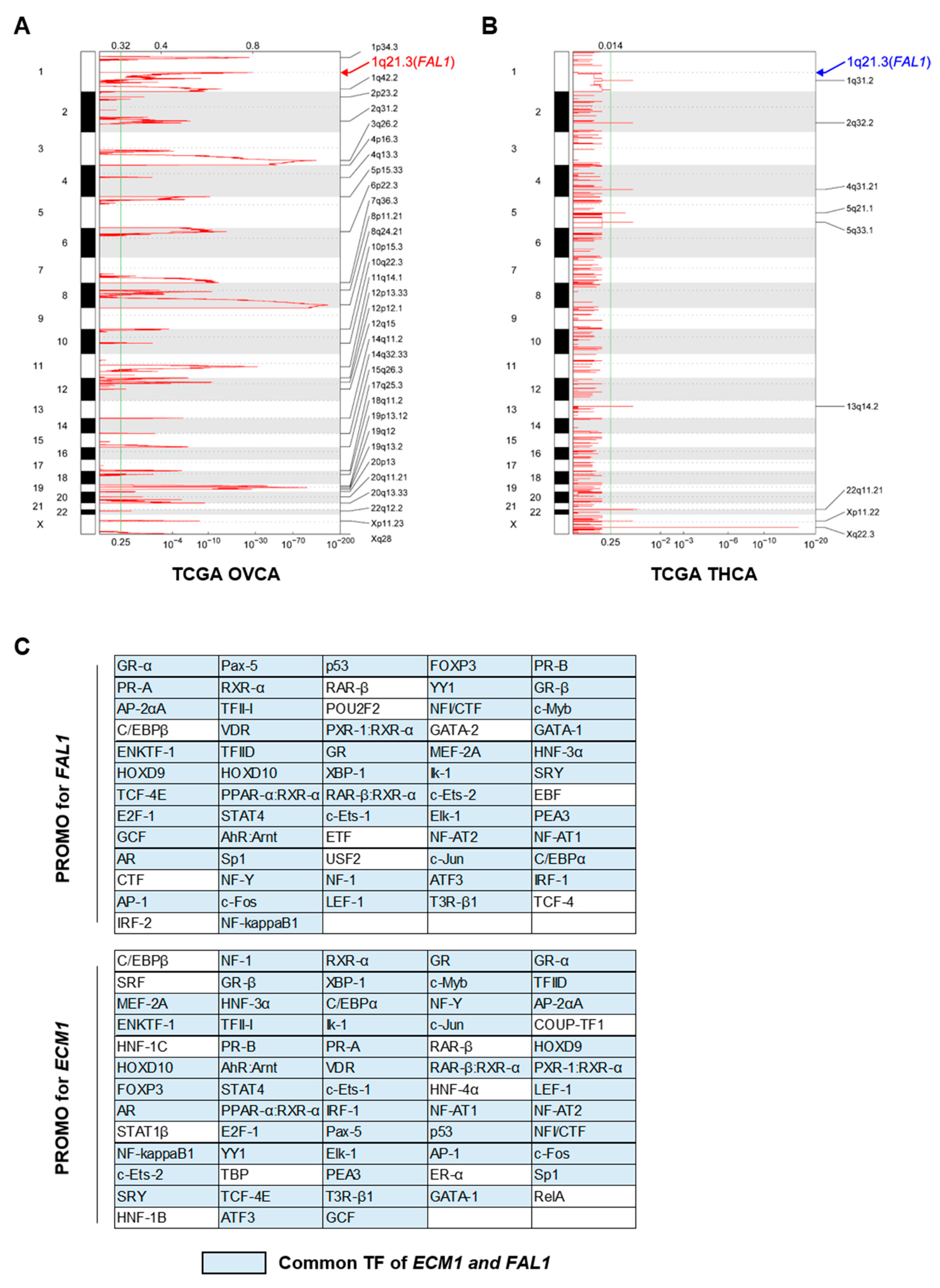
3.2. Focal Amplification of the FAL1 Gene in Ovarian Cancer but Not In PTC
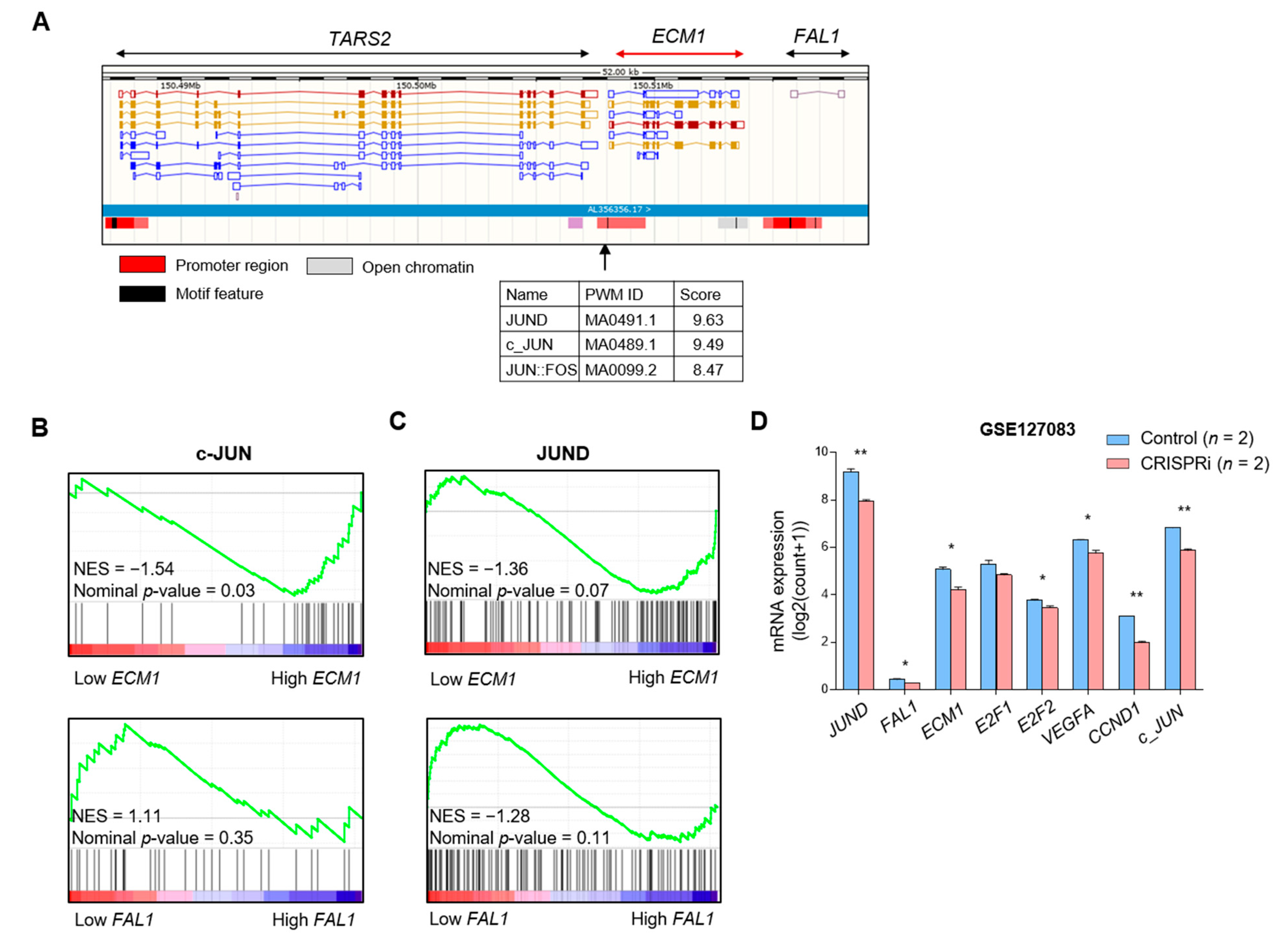
3.3. JUND as a Candidate for Common Transcription Factor for FAL1 and ECM1
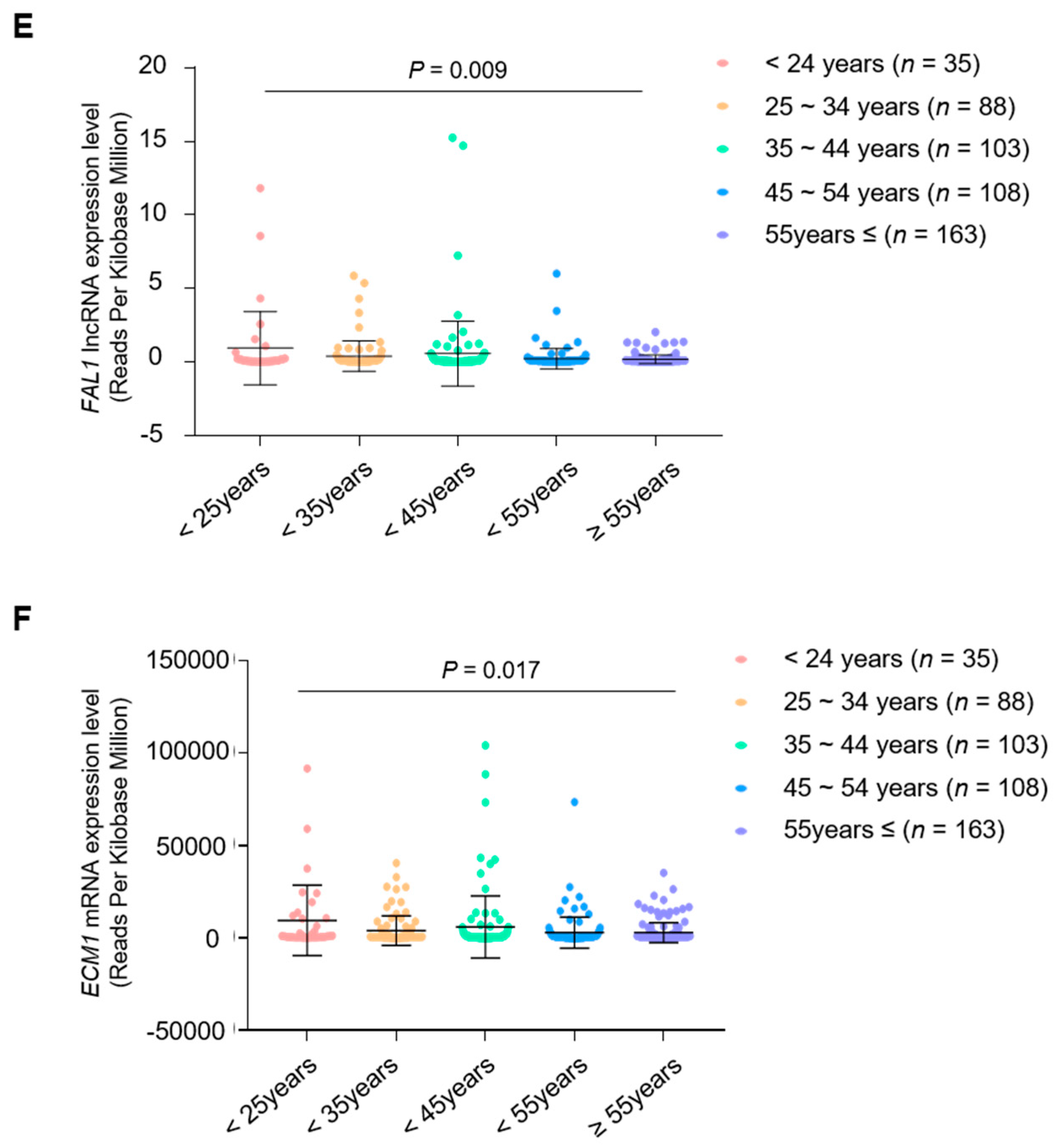
3.4. Clinical Relevance of the Simultaneous Expression of FAL1 and ECM1
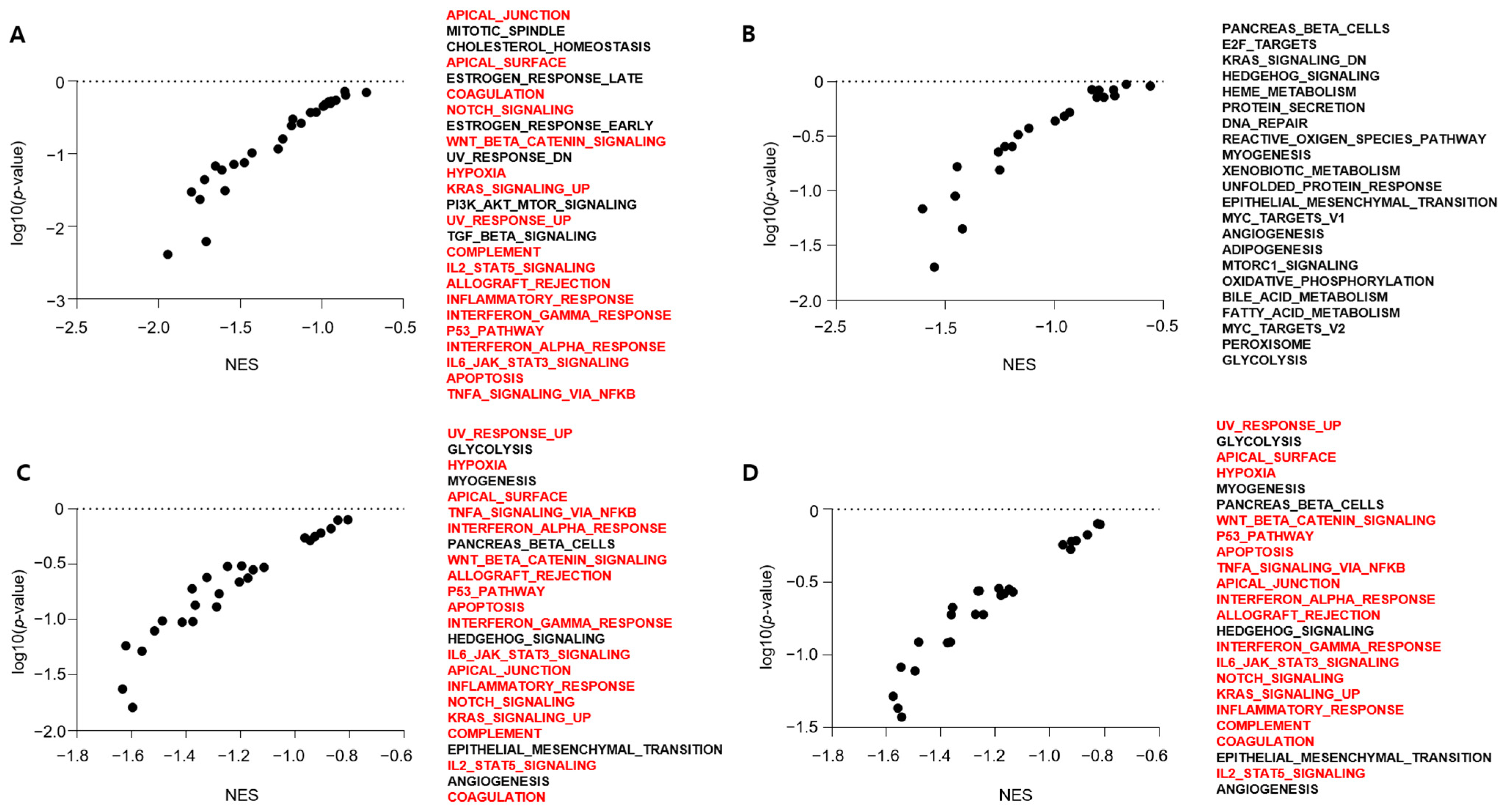
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davies, L.; Welch, H.G. Increasing incidence of thyroid cancer in the United States, 1973–2002. Jama 2006, 295, 2164–2167. [Google Scholar] [CrossRef] [Green Version]
- Jung, C.K.; Little, M.P.; Lubin, J.H.; Brenner, A.V.; Wells, S.A., Jr.; Sigurdson, A.J.; Nikiforov, Y.E. The increase in thyroid cancer incidence during the last four decades is accompanied by a high frequency of BRAF mutations and a sharp increase in RAS mutations. J. Clin. Endocrinol. Metab. 2014, 99, E276–E285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, J.; Kim, E.; Han, M.; Jung, I.; Lee, J.; Jo, Y.S. Impact of Dyslipidemia on the Risk of Second Cancer in Thyroid Cancer Patients: A Korean National Cohort Study. Ann. Surg. Oncol. 2021. [Google Scholar] [CrossRef]
- Lee, W.K.; Lee, J.; Kim, H.; Lee, S.G.; Choi, S.H.; Jeong, S.; Kwon, H.J.; Jung, S.G.; Jo, Y.S. Peripheral location and infiltrative margin predict invasive features of papillary thyroid microcarcinoma. Eur. J. Endocrinol. 2019, 181, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jeong, S.; Park, J.H.; Lee, C.R.; Ku, C.R.; Kang, S.W.; Jeong, J.J.; Nam, K.H.; Shin, D.Y.; Lee, E.J.; et al. Aberrant expression of COT is related to recurrence of papillary thyroid cancer. Medicine 2015, 94, e548. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Seol, M.Y.; Jeong, S.; Lee, C.R.; Ku, C.R.; Kang, S.W.; Jeong, J.J.; Shin, D.Y.; Nam, K.H.; Lee, E.J.; et al. A metabolic phenotype based on mitochondrial ribosomal protein expression as a predictor of lymph node metastasis in papillary thyroid carcinoma. Medicine 2015, 94, e380. [Google Scholar] [CrossRef]
- Zhang, L.; Wei, W.J.; Ji, Q.H.; Zhu, Y.X.; Wang, Z.Y.; Wang, Y.; Huang, C.P.; Shen, Q.; Li, D.S.; Wu, Y. Risk factors for neck nodal metastasis in papillary thyroid microcarcinoma: A study of 1066 patients. J. Clin. Endocrinol. Metab. 2012, 97, 1250–1257. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Kim, C.H.; Min, I.K.; Jeong, S.; Kim, H.; Choi, M.J.; Kwon, H.J.; Jung, S.G.; Jo, Y.S. Detailed characterization of metastatic lymph nodes improves the prediction accuracy of currently used risk stratification systems in N1 stage papillary thyroid cancer. Eur. J. Endocrinol. 2020, 183, 83–93. [Google Scholar] [CrossRef]
- Vasko, V.; Espinosa, A.V.; Scouten, W.; He, H.; Auer, H.; Liyanarachchi, S.; Larin, A.; Savchenko, V.; Francis, G.L.; de la Chapelle, A.; et al. Gene expression and functional evidence of epithelial-to-mesenchymal transition in papillary thyroid carcinoma invasion. Proc. Natl. Acad. Sci. USA 2007, 104, 2803–2808. [Google Scholar] [CrossRef] [Green Version]
- Higgs, P.G.; Lehman, N. The RNA World: Molecular cooperation at the origins of life. Nat. Rev. Genet. 2015, 16, 7–17. [Google Scholar] [CrossRef]
- Morris, K.V.; Mattick, J.S. The rise of regulatory RNA. Nat. Rev. Genet. 2014, 15, 423–437. [Google Scholar] [CrossRef] [Green Version]
- Esteller, M. Non-coding RNAs in human disease. Nat. Reviews. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Li, W.; Liyanarachchi, S.; Jendrzejewski, J.; Srinivas, M.; Davuluri, R.V.; Nagy, R.; de la Chapelle, A. Genetic predisposition to papillary thyroid carcinoma: Involvement of FOXE1, TSHR, and a novel lincRNA gene, PTCSC2. J. Clin. Endocrinol. Metab. 2015, 100, E164–E172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jendrzejewski, J.; He, H.; Radomska, H.S.; Li, W.; Tomsic, J.; Liyanarachchi, S.; Davuluri, R.V.; Nagy, R.; de la Chapelle, A. The polymorphism rs944289 predisposes to papillary thyroid carcinoma through a large intergenic noncoding RNA gene of tumor suppressor type. Proc. Natl. Acad. Sci. USA 2012, 109, 8646–8651. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Guo, Q.; Zhao, Y.; Chen, J.; Wang, S.; Hu, J.; Sun, Y. BRAF-activated long non-coding RNA contributes to cell proliferation and activates autophagy in papillary thyroid carcinoma. Oncol. Lett. 2014, 8, 1947–1952. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.; Lee, J.; Kim, D.; Seol, M.Y.; Lee, W.K.; Jeong, J.J.; Nam, K.H.; Jung, S.G.; Shin, D.Y.; Lee, E.J.; et al. Relationship of Focally Amplified Long Noncoding on Chromosome 1 (FAL1) lncRNA with E2F Transcription Factors in Thyroid Cancer. Medicine 2016, 95, e2592. [Google Scholar] [CrossRef]
- Li, J.; Han, L.; Roebuck, P.; Diao, L.; Liu, L.; Yuan, Y.; Weinstein, J.N.; Liang, H. TANRIC: An Interactive Open Platform to Explore the Function of lncRNAs in Cancer. Cancer Res. 2015, 75, 3728–3737. [Google Scholar] [CrossRef] [Green Version]
- Mermel, C.H.; Schumacher, S.E.; Hill, B.; Meyerson, M.L.; Beroukhim, R.; Getz, G. GISTIC2.0 facilitates sensitive and confident localization of the targets of focal somatic copy-number alteration in human cancers. Genome Biol. 2011, 12, R41. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Feng, Y.; Zhang, D.; Zhao, S.D.; Hu, Z.; Greshock, J.; Zhang, Y.; Yang, L.; Zhong, X.; Wang, L.P.; et al. A functional genomic approach identifies FAL1 as an oncogenic long noncoding RNA that associates with BMI1 and represses p21 expression in cancer. Cancer Cell 2014, 26, 344–357. [Google Scholar] [CrossRef] [Green Version]
- Messeguer, X.; Escudero, R.; Farre, D.; Nunez, O.; Martinez, J.; Alba, M.M. PROMO: Detection of known transcription regulatory elements using species-tailored searches. Bioinformatics 2002, 18, 333–334. [Google Scholar] [CrossRef] [PubMed]
- Farre, D.; Roset, R.; Huerta, M.; Adsuara, J.E.; Rosello, L.; Alba, M.M.; Messeguer, X. Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res. 2003, 31, 3651–3653. [Google Scholar] [CrossRef] [Green Version]
- Guigo, R.; Flicek, P.; Abril, J.F.; Reymond, A.; Lagarde, J.; Denoeud, F.; Antonarakis, S.; Ashburner, M.; Bajic, V.B.; Birney, E.; et al. EGASP: The human ENCODE Genome Annotation Assessment Project. Genome Biol. 2006, 7 (Suppl. 1), 1–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S.; Kim, I.K.; Kim, H.; Choi, M.J.; Lee, J.; Jo, Y.S. Liver X Receptor beta Related to Tumor Progression and Ribosome Gene Expression in Papillary Thyroid Cancer. Endocrinol. Metab. 2020, 35, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.B.; Lee, W.K.; Lee, S.G.; Ryu, H.; Lee, C.R.; Kang, S.W.; Jeong, J.J.; Nam, K.H.; Lee, E.J.; Chung, W.Y.; et al. Long-term oncologic outcomes of papillary thyroid microcarcinoma according to the presence of clinically apparent lymph node metastasis: A large retrospective analysis of 5,348 patients. Cancer Manag. Res. 2018, 10, 2883–2891. [Google Scholar] [CrossRef] [Green Version]
- Sui, F.; Ji, M.; Hou, P. Long non-coding RNAs in thyroid cancer: Biological functions and clinical significance. Mol. Cell Endocrinol. 2018, 469, 11–22. [Google Scholar] [CrossRef]
- Mahmoudian-Sani, M.R.; Jalali, A.; Jamshidi, M.; Moridi, H.; Alghasi, A.; Shojaeian, A.; Mobini, G.R. Long Non-Coding RNAs in Thyroid Cancer: Implications for Pathogenesis, Diagnosis, and Therapy. Oncol. Res. Treat. 2019, 42, 136–142. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Mohammad-Rahimi, H.; Taheri, M. The role of long non-coding RNAs in the pathogenesis of thyroid cancer. Exp. Mol. Pathol. 2020, 112, 104332. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, K.; Ma, L.; Xu, J.; Chang, W. The Role of Long Non-Coding RNAs in Thyroid Cancer. Front. Oncol. 2020, 10, 941. [Google Scholar] [CrossRef] [PubMed]
- Samimi, H.; Sajjadi-Jazi, S.M.; Seifirad, S.; Atlasi, R.; Mahmoodzadeh, H.; Faghihi, M.A.; Haghpanah, V. Molecular mechanisms of long non-coding RNAs in anaplastic thyroid cancer: A systematic review. Cancer Cell Int. 2020, 20, 352. [Google Scholar] [CrossRef]
- Kim, D.; Lee, W.K.; Jeong, S.; Seol, M.Y.; Kim, H.; Kim, K.S.; Lee, E.J.; Lee, J.; Jo, Y.S. Upregulation of long noncoding RNA LOC100507661 promotes tumor aggressiveness in thyroid cancer. Mol. Cell Endocrinol. 2016, 431, 36–45. [Google Scholar] [CrossRef]
- Lee, J.; Jeong, S.; Lee, C.R.; Ku, C.R.; Kang, S.W.; Jeong, J.J.; Nam, K.H.; Shin, D.Y.; Chung, W.Y.; Lee, E.J.; et al. GLI1 Transcription Factor Affects Tumor Aggressiveness in Patients With Papillary Thyroid Cancers. Medicine 2015, 94, e998. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Seol, M.Y.; Jeong, S.; Kwon, H.J.; Lee, C.R.; Ku, C.R.; Kang, S.W.; Jeong, J.J.; Shin, D.Y.; Nam, K.H.; et al. KSR1 is coordinately regulated with Notch signaling and oxidative phosphorylation in thyroid cancer. J. Mol. Endocrinol. 2015, 54, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.K.; Lee, S.G.; Yim, S.H.; Kim, D.; Kim, H.; Jeong, S.; Jung, S.G.; Jo, Y.S.; Lee, J. Whole Exome Sequencing Identifies a Novel Hedgehog-Interacting Protein G516R Mutation in Locally Advanced Papillary Thyroid Cancer. Int. J. Mol. Sci. 2018, 19, 2867. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular Feature | FAL1 | p Value | |
|---|---|---|---|
| Low Expression n = 124 (%) | High Expression n = 124 (%) | ||
| mRNA cluster number | |||
| 1 | 12 (9.8) | 59 (48.8) | <0.0001 † |
| 2 | 13 (10.7) | 22 (18.2) | |
| 3 | 32 (26.2) | 5 (4.1) | |
| 4 | 39 (32.0) | 8 (6.6) | |
| 5 | 26 (21.3) | 27 (22.3) | |
| miRNA cluster number | |||
| 1 | 2 (1.6) | 2 (1.6) | <0.0001 † |
| 2 | 48 (39.0) | 19 (15.4) | |
| 3 | 26 (21.1) | 7 (5.7) | |
| 4 | 16 (13.0) | 58 (47.2) | |
| 5 | 16 (13.0) | 19 (15.4) | |
| 6 | 15 (12.2) | 18 (14.6) | |
| RAS mutation | |||
| Absent | 122 (98.4) | 89 (72.4) | <0.0001 † |
| Present | 2 (1.6) | 34 (27.6) | |
| BRAF mutation | |||
| Absent | 31 (25.0) | 97 (78.9) | <0.0001 † |
| Present | 93 (75.0) | 26 (21.1) | |
| TERT promoter mutation | |||
| Absent | 88 (87.1) | 81 (89.0) | 0.688 † |
| Present | 13 (12.9) | 10 (11.0) | |
| RAS/RAF score | −0.67 ± 0.49 | 0.21 ± 0.67 | <0.0001 * |
| ERK score | 11.37 ± 16.27 | 2.14 ± 20.26 | 0.001 * |
| Differentiation score | −0.49 ± 0.99 | 0.54 ± 1.08 | <0.0001 * |
| Akt pT308 | −0.02 ± 0.64 | 0.06 ± 0.47 | 0.275 * |
| Akt pS473 | −0.02 ± 0.47 | 0.09 ± 0.34 | 0.047 * |
| Molecular Feature | ECM1 | p Value | |
|---|---|---|---|
| Low Expression n = 124 (%) | High Expression n = 124 (%) | ||
| mRNA cluster number | |||
| 1 | 58 (47.9) | 51 (43.2) | 0.190 † |
| 2 | 7 (5.8) | 21 (17.8) | |
| 3 | 36 (29.8) | 5 (4.2) | |
| 4 | 5 (4.1) | 14 (11.9) | |
| 5 | 15 (12.4) | 27 (22.9) | |
| miRNA cluster number | |||
| 1 | 3 (2.5) | 3 (2.4) | 0.796 † |
| 2 | 16 (13.1) | 22 (17.9) | |
| 3 | 24 (19.7) | 11 (8.9) | |
| 4 | 46 (37.7) | 51 (41.5) | |
| 5 | 19 (15.6) | 21 (17.1) | |
| 6 | 14 (11.5) | 15 (12.2) | |
| RAS mutation | |||
| Absent | 117 (95.1) | 90 (73.2) | <0.0001 † |
| Present | 6 (4.9) | 33 (26.8) | |
| BRAF mutation | |||
| Absent | 71 (57.7) | 92 (74.8) | 0.005 † |
| Present | 52 (42.3) | 31 (25.2) | |
| TERT promoter mutation | |||
| Absent | 91 (95.8) | 78 (84.8) | 0.011 † |
| Present | 4 (4.2) | 14 (15.2) | |
| RAS/RAF score | 0.01 ± 0.81 | 0.07 ± 0.68 | 0.533 * |
| ERK score | −5.25 ± 23.26 | 4.92 ± 19.35 | 0.001 * |
| Differentiation score | 0.32 ± 1.37 | 0.39 ± 1.00 | 0.674 * |
| Akt pT308 | 0.09 ± 0.54 | 0.06 ± 0.35 | 0.691 * |
| Akt pS473 | 0.14 ± 0.66 | 0.01 ± 0.46 | 0.122 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, S.; Lee, S.-G.; Kim, H.; Lee, G.; Park, S.; Kim, I.-K.; Lee, J.; Jo, Y.-S. Simultaneous Expression of Long Non-Coding RNA FAL1 and Extracellular Matrix Protein 1 Defines Tumour Behaviour in Young Patients with Papillary Thyroid Cancer. Cancers 2021, 13, 3223. https://doi.org/10.3390/cancers13133223
Jeong S, Lee S-G, Kim H, Lee G, Park S, Kim I-K, Lee J, Jo Y-S. Simultaneous Expression of Long Non-Coding RNA FAL1 and Extracellular Matrix Protein 1 Defines Tumour Behaviour in Young Patients with Papillary Thyroid Cancer. Cancers. 2021; 13(13):3223. https://doi.org/10.3390/cancers13133223
Chicago/Turabian StyleJeong, Seonhyang, Seul-Gi Lee, Hyunji Kim, Gibbeum Lee, Sunmi Park, In-Kyu Kim, Jandee Lee, and Young-Suk Jo. 2021. "Simultaneous Expression of Long Non-Coding RNA FAL1 and Extracellular Matrix Protein 1 Defines Tumour Behaviour in Young Patients with Papillary Thyroid Cancer" Cancers 13, no. 13: 3223. https://doi.org/10.3390/cancers13133223