
Nrf2/Keap1-Pathway Activation and Reduced Susceptibility to Chemotherapy Treatment by Acidification in Esophageal Adenocarcinoma Cells

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Treatment with Acidified Medium
2.3. Protein Isolation and Western Blotting
2.4. RNA Isolation and cDNA Synthesis
2.5. Quantitative RT-PCR
2.6. Viability Assay
2.7. ARE Luciferase Assay
2.8. 3D-Culture
2.9. Histology and Immunohistochemistry
2.10. Immunohistochemistry for HO1
2.11. Statistics
3. Results
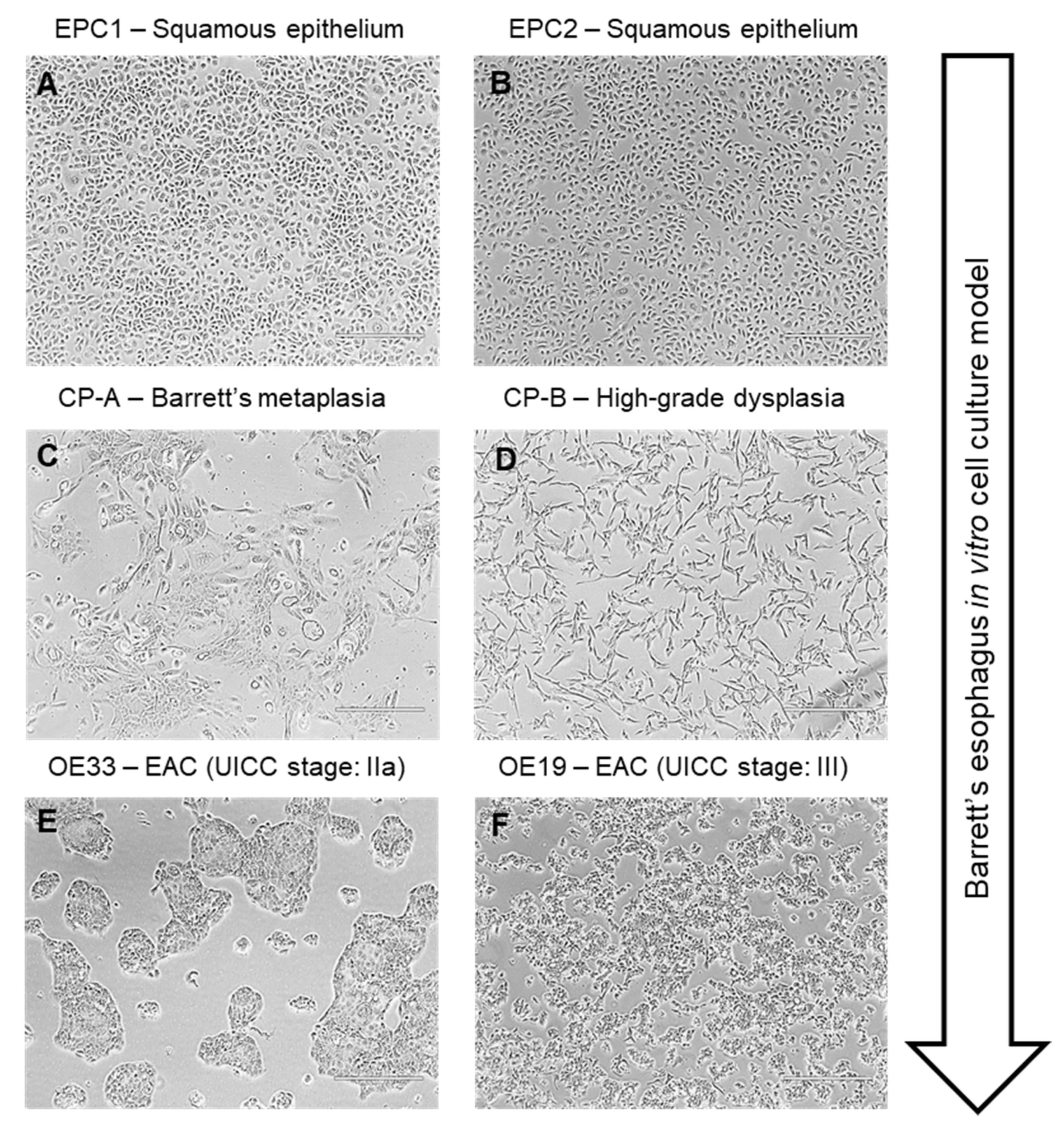
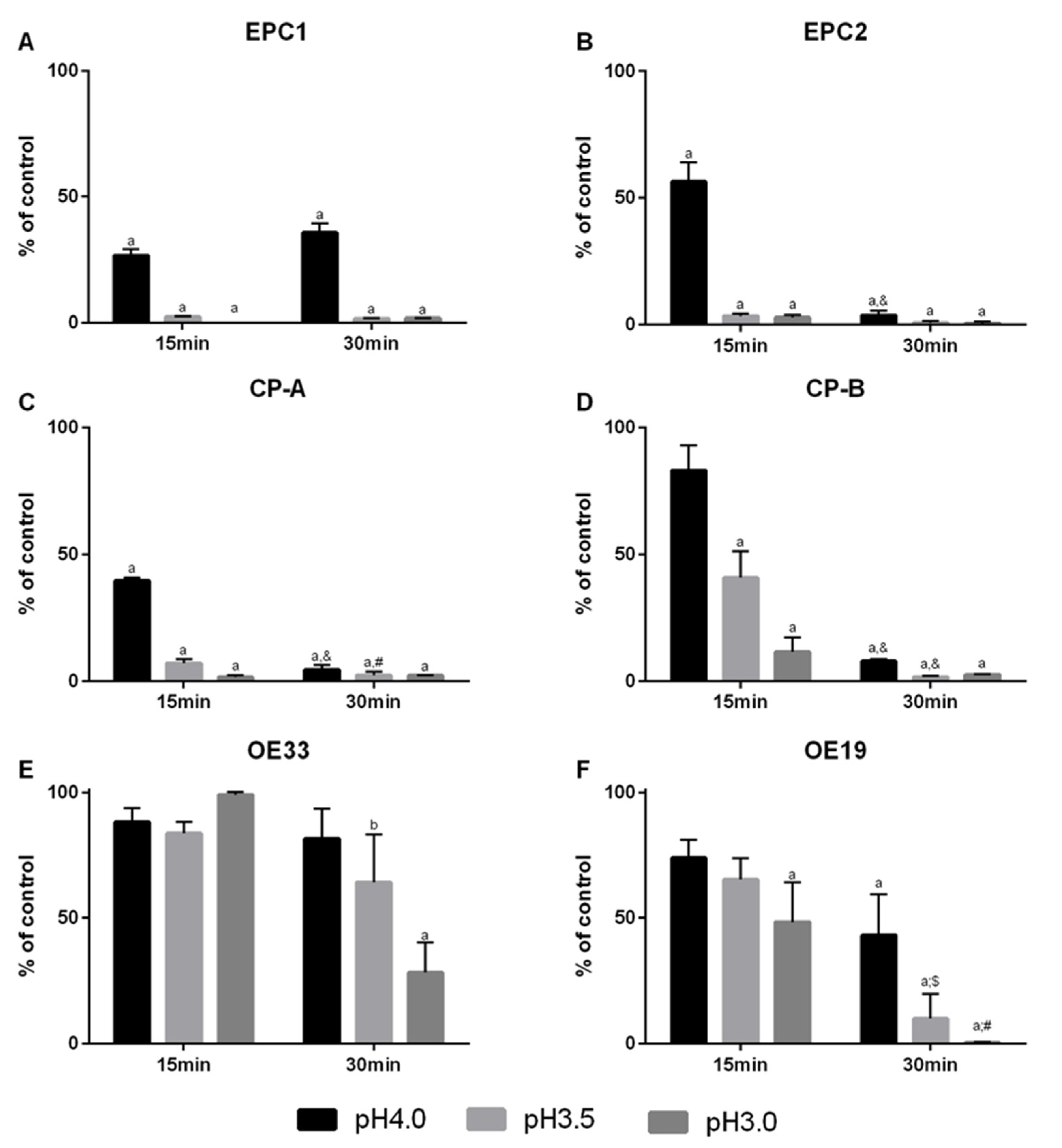
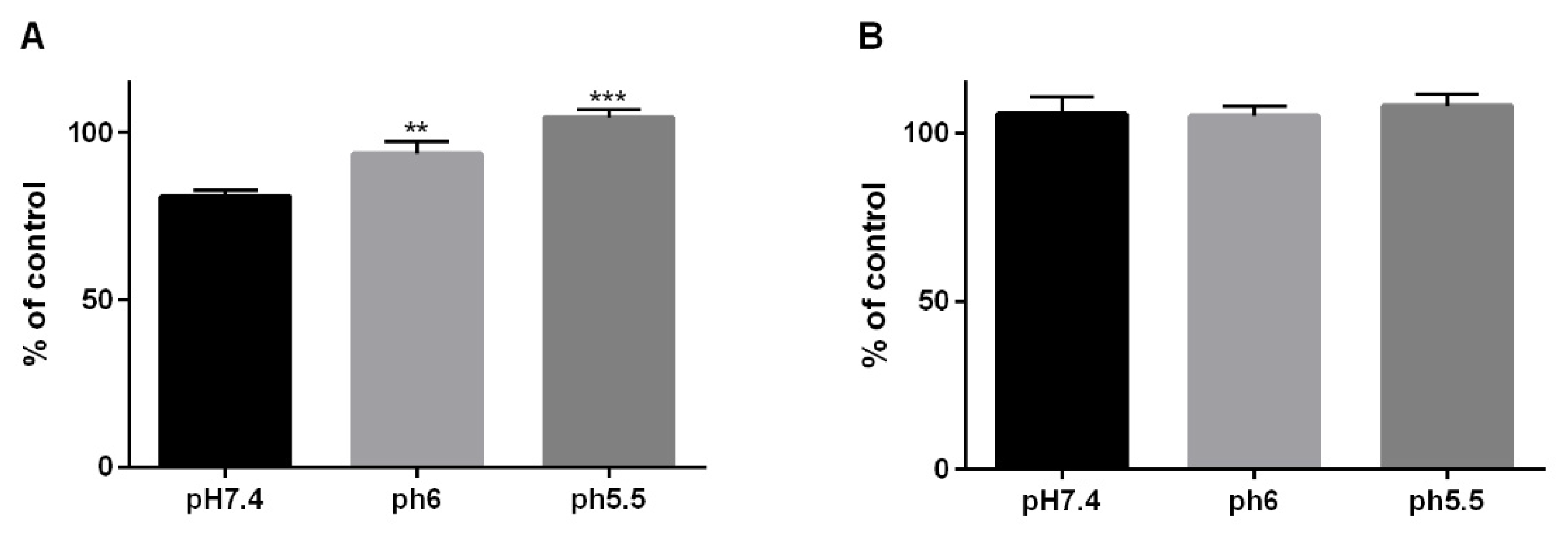
3.1. Cellular Viability in Cells of the Barrett’s Sequence by a Reflux-Like Acidic Exposure
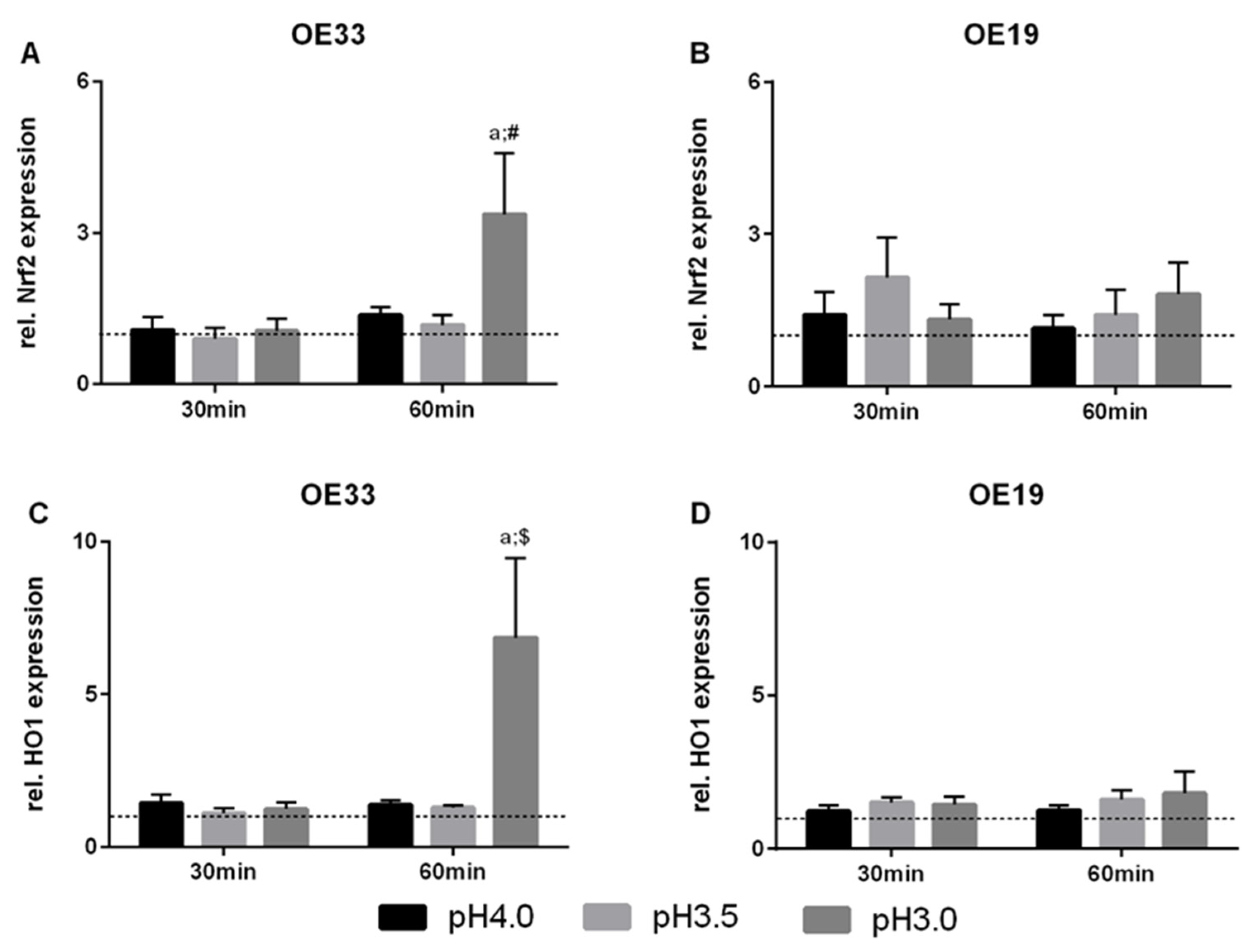
3.2. Nrf2 and HO1 Expression in EAC Cells
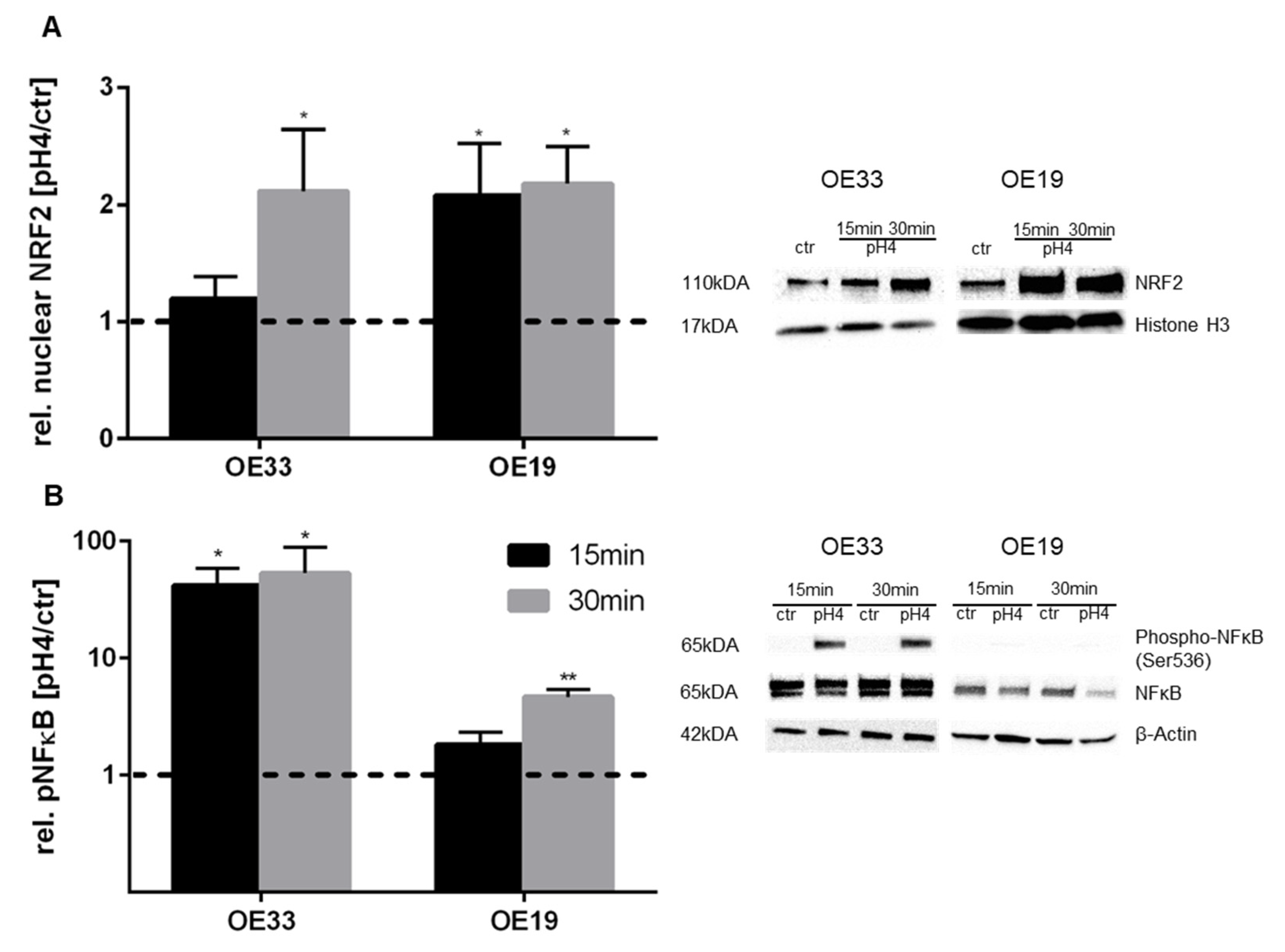
3.3. pH 4-Induced Nrf2 and NFκB Activation in EAC Cells
3.4. Dependency of 5-FU Effectiveness with Regard to a pH-Shift
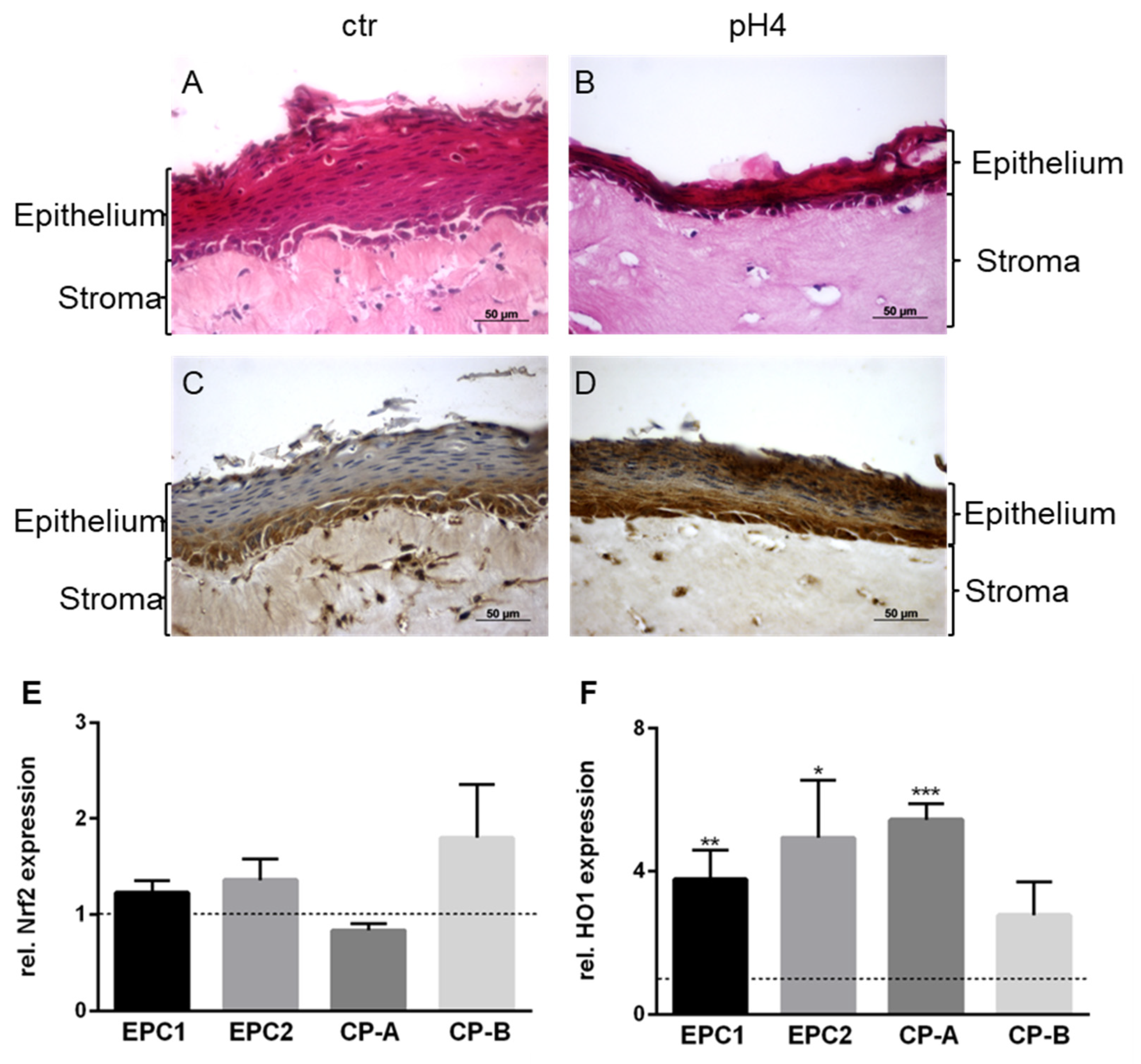
3.5. Organotypic 3D-Cultures and Nrf2 and HO1 Expression in Non-Malignant Cells with Chronical Acid Exposure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lordick, F.; Mariette, C.; Haustermans, K.; Obermannova, R.; Arnold, D.; Committee, E.G. Oesophageal cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2016, 27, v50–v57. [Google Scholar] [CrossRef]
- Al-Batran, S.E.; Hofheinz, R.D.; Pauligk, C.; Kopp, H.G.; Haag, G.M.; Luley, K.B.; Meiler, J.; Homann, N.; Lorenzen, S.; Schmalenberg, H.; et al. Histopathological regression after neoadjuvant docetaxel, oxaliplatin, fluorouracil, and leucovorin versus epirubicin, cisplatin, and fluorouracil or capecitabine in patients with resectable gastric or gastro-oesophageal junction adenocarcinoma (FLOT4-AIO): Results from the phase 2 part of a multicentre, open-label, randomised phase 2/3 trial. Lancet Oncol. 2016, 17, 1697–1708. [Google Scholar] [CrossRef] [PubMed]
- Al-Batran, S.-E.; Homann, N.; Schmalenberg, H.; Kopp, H.-G.; Haag, G.M.; Luley, K.B.; Schmiegel, W.H.; Folprecht, G.; Probst, S.; Prasnikar, N.; et al. Perioperative chemotherapy with docetaxel, oxaliplatin, and fluorouracil/leucovorin (FLOT) versus epirubicin, cisplatin, and fluorouracil or capecitabine (ECF/ECX) for resectable gastric or gastroesophageal junction (GEJ) adenocarcinoma (FLOT4-AIO): A multicenter, randomized phase 3 trial. J. Clin. Oncol. 2017, 35, 4004. [Google Scholar] [CrossRef]
- Hoeppner, J.; Lordick, F.; Brunner, T.; Glatz, T.; Bronsert, P.; Rothling, N.; Schmoor, C.; Lorenz, D.; Ell, C.; Hopt, U.T.; et al. ESOPEC: Prospective randomized controlled multicenter phase III trial comparing perioperative chemotherapy (FLOT protocol) to neoadjuvant chemoradiation (CROSS protocol) in patients with adenocarcinoma of the esophagus (NCT02509286). BMC Cancer 2016, 16, 503. [Google Scholar] [CrossRef] [Green Version]
- Ott, K.; Herrmann, K.; Krause, B.J.; Lordick, F. The Value of PET Imaging in Patients with Localized Gastroesophageal Cancer. Gastrointest. Cancer Res. 2008, 2, 287–294. [Google Scholar] [PubMed]
- Abdel-Latif, M.M.; O’Riordan, J.; Windle, H.J.; Carton, E.; Ravi, N.; Kelleher, D.; Reynolds, J.V. NF-kappaB activation in esophageal adenocarcinoma: Relationship to Barrett’s metaplasia, survival, and response to neoadjuvant chemoradiotherapy. Ann. Surg. 2004, 239, 491–500. [Google Scholar] [CrossRef]
- Spechler, S.J. Clinical practice. Barrett’s Esophagus. N. Engl. J. Med. 2002, 346, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Lagergren, J.; Smyth, E.; Cunningham, D.; Lagergren, P. Oesophageal cancer. Lancet 2017, 390, 2383–2396. [Google Scholar] [CrossRef] [Green Version]
- Bennett, C.; Moayyedi, P.; Corley, D.A.; DeCaestecker, J.; Falck-Ytter, Y.; Falk, G.; Vakil, N.; Sanders, S.; Vieth, M.; Inadomi, J.; et al. BOB CAT: A Large-Scale Review and Delphi Consensus for Management of Barrett’s Esophagus with No Dysplasia, Indefinite for, or Low-Grade Dysplasia. Am. J. Gastroenterol. 2015, 110, 662–682. [Google Scholar] [CrossRef] [Green Version]
- de Jonge, P.J.; van Blankenstein, M.; Grady, W.M.; Kuipers, E.J. Barrett’s oesophagus: Epidemiology, cancer risk and implications for management. Gut 2014, 63, 191–202. [Google Scholar] [CrossRef]
- Murakami, S.; Motohashi, H. Roles of Nrf2 in cell proliferation and differentiation. Free Radic. Biol. Med. 2015, 88, 168–178. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Castranova, V.; Shi, X. New insights into the role of nuclear factor-kappaB in cell growth regulation. Am. J. Pathol. 2001, 159, 387–397. [Google Scholar] [CrossRef]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Latif, M.M.; O’Riordan, J.M.; Ravi, N.; Kelleher, D.; Reynolds, J.V. Activated nuclear factor-kappa B and cytokine profiles in the esophagus parallel tumor regression following neoadjuvant chemoradiotherapy. Dis. Esophagus 2005, 18, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Harada, H.; Nakagawa, H.; Oyama, K.; Takaoka, M.; Andl, C.D.; Jacobmeier, B.; von Werder, A.; Enders, G.H.; Opitz, O.G.; Rustgi, A.K. Telomerase induces immortalization of human esophageal keratinocytes without p16INK4a inactivation. Mol. Cancer Res. 2003, 1, 729–738. [Google Scholar] [PubMed]
- Lyros, O.; Rafiee, P.; Nie, L.; Medda, R.; Jovanovic, N.; Otterson, M.F.; Behmaram, B.; Gockel, I.; Mackinnon, A.; Shaker, R. Wnt/beta-Catenin Signaling Activation beyond Robust Nuclear beta-Catenin Accumulation in Nondysplastic Barrett’s Esophagus: Regulation via Dickkopf-1. Neoplasia 2015, 17, 598–611. [Google Scholar] [CrossRef] [Green Version]
- Chemnitzer, O.; Gotzel, K.; Maurer, L.; Dietrich, A.; Eichfeld, U.; Lyros, O.; Jansen-Winkeln, B.; Hoffmeister, A.; Gockel, I.; Thieme, R. Response to TNF-alpha Is Increasing Along with the Progression in Barrett’s Esophagus. Dig. Dis. Sci. 2017, 62, 3391–3401. [Google Scholar] [CrossRef] [PubMed]
- Pendzialek, S.M.; Knelangen, J.M.; Schindler, M.; Gurke, J.; Grybel, K.J.; Gocza, E.; Fischer, B.; Santos, A.N. Trophoblastic microRNAs are downregulated in a diabetic pregnancy through an inhibition of Drosha. Mol. Cell Endocrinol. 2019, 480, 167–179. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Kalabis, J.; Wong, G.S.; Vega, M.E.; Natsuizaka, M.; Robertson, E.S.; Herlyn, M.; Nakagawa, H.; Rustgi, A.K. Isolation and characterization of mouse and human esophageal epithelial cells in 3D organotypic culture. Nat. Protoc. 2012, 7, 235–246. [Google Scholar] [CrossRef] [Green Version]
- Lyros, O.; Rafiee, P.; Nie, L.; Medda, R.; Jovanovic, N.; Schmidt, J.; Mackinnon, A.; Venu, N.; Shaker, R. Dickkopf-1, the Wnt antagonist, is induced by acidic pH and mediates epithelial cellular senescence in human reflux esophagitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G557–G574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman, A.; Condon, A.; Adler, E.; Minnella, M.; Bernstein, C.; Bernstein, H.; Dvorak, K. Protective effects of glycoursodeoxycholic acid in Barrett’s esophagus cells. Dis. Esophagus 2010, 23, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Torihata, Y.; Asanuma, K.; Iijima, K.; Mikami, T.; Hamada, S.; Asano, N.; Koike, T.; Imatani, A.; Masamune, A.; Shimosegawa, T. Estrogen-Dependent Nrf2 Expression Protects Against Reflux-Induced Esophagitis. Dig. Dis. Sci. 2018, 63, 345–355. [Google Scholar] [CrossRef]
- Perez, S.; Talens-Visconti, R.; Rius-Perez, S.; Finamor, I.; Sastre, J. Redox signaling in the gastrointestinal tract. Free Radic. Biol. Med. 2017, 104, 75–103. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Huo, X.; Rezaei, D.; Zhang, Q.; Zhang, X.; Yu, C.; Asanuma, K.; Cheng, E.; Pham, T.H.; Wang, D.H.; et al. In Barrett’s esophagus patients and Barrett’s cell lines, ursodeoxycholic acid increases antioxidant expression and prevents DNA damage by bile acids. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G129–G139. [Google Scholar] [CrossRef]
- Kobayashi, M.; Yamamoto, M. Molecular mechanisms activating the Nrf2-Keap1 pathway of antioxidant gene regulation. Antioxid. Redox Signal. 2005, 7, 385–394. [Google Scholar] [CrossRef]
- Hu, R.; Saw, C.L.; Yu, R.; Kong, A.N. Regulation of NF-E2-related factor 2 signaling for cancer chemoprevention: Antioxidant coupled with antiinflammatory. Antioxid. Redox Signal. 2010, 13, 1679–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, D.; Lu, H.; Zhu, S.; Zhou, Z.; Hu, T.; Chen, Z.; Zaika, A.; El-Rifai, W. NRF2 antioxidant response protects against acidic bile salts-induced oxidative stress and DNA damage in esophageal cells. Cancer Lett. 2019, 458, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network. Integrated genomic characterization of oesophageal carcinoma. Nature 2017, 541, 169–175. [CrossRef] [PubMed] [Green Version]
- Peng, D.; Zaika, A.; Que, J.; El-Rifai, W. The antioxidant response in Barrett’s tumorigenesis: A double-edged sword. Redox Biol. 2021, 41, 101894. [Google Scholar] [CrossRef]
- Tselepis, C.; Perry, I.; Dawson, C.; Hardy, R.; Darnton, S.J.; McConkey, C.; Stuart, R.C.; Wright, N.; Harrison, R.; Jankowski, J.A. Tumour necrosis factor-alpha in Barrett’s oesophagus: A potential novel mechanism of action. Oncogene 2002, 21, 6071–6081. [Google Scholar] [CrossRef] [Green Version]
- O’Donovan, T.R.; O’Sullivan, G.C.; McKenna, S.L. Induction of autophagy by drug-resistant esophageal cancer cells promotes their survival and recovery following treatment with chemotherapeutics. Autophagy 2011, 7, 509–524. [Google Scholar] [CrossRef] [Green Version]
- Falvey, C.M.; O’Donovan, T.R.; El-Mashed, S.; Nyhan, M.J.; O’Reilly, S.; McKenna, S.L. UBE2L6/UBCH8 and ISG15 attenuate autophagy in esophageal cancer cells. Oncotarget 2017, 8, 23479–23491. [Google Scholar] [CrossRef]
- Ajani, J.A.; Wang, X.; Song, S.; Suzuki, A.; Taketa, T.; Sudo, K.; Wadhwa, R.; Hofstetter, W.L.; Komaki, R.; Maru, D.M.; et al. ALDH-1 expression levels predict response or resistance to preoperative chemoradiation in resectable esophageal cancer patients. Mol. Oncol. 2014, 8, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Kastelein, F.; Spaander, M.C.; Steyerberg, E.W.; Biermann, K.; Valkhoff, V.E.; Kuipers, E.J.; Bruno, M.J.; ProBar Study, G. Proton pump inhibitors reduce the risk of neoplastic progression in patients with Barrett’s esophagus. Clin. Gastroenterol. Hepatol. 2013, 11, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Ouatu-Lascar, R.; Fitzgerald, R.C.; Triadafilopoulos, G. Differentiation and proliferation in Barrett’s esophagus and the effects of acid suppression. Gastroenterology 1999, 117, 327–335. [Google Scholar] [CrossRef]
- Booth, C.L.; Thompson, K.S. Barrett’s esophagus: A review of diagnostic criteria, clinical surveillance practices and new developments. J. Gastrointest. Oncol. 2012, 3, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Lagergren, J.; Bergstrom, R.; Lindgren, A.; Nyren, O. Symptomatic gastroesophageal reflux as a risk factor for esophageal adenocarcinoma. N. Engl. J. Med. 1999, 340, 825–831. [Google Scholar] [CrossRef]
- Weismuller, J.; Thieme, R.; Hoffmeister, A.; Weismuller, T.; Gockel, I. Barrett-Screening: Rational, current concepts and perspectives. Z. Gastroenterol. 2019, 57, 317–326. [Google Scholar] [CrossRef]
- Elias, P.S.; Castell, D.O. The Role of Acid Suppression in Barrett’s Esophagus. Am. J. Med. 2017, 130, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.H.; Tiwari, A.; Kim, M.E.; Clemons, N.J.; Regmi, N.L.; Hodges, W.A.; Berman, D.M.; Montgomery, E.A.; Watkins, D.N.; Zhang, X.; et al. Hedgehog signaling regulates FOXA2 in esophageal embryogenesis and Barrett’s metaplasia. J. Clin. Investig. 2014, 124, 3767–3780. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.J.; Suh, E.R.; Lynch, J.P. The role of Cdx proteins in intestinal development and cancer. Cancer Biol. Ther. 2004, 3, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Beck, F. The role of Cdx genes in the mammalian gut. Gut 2004, 53, 1394–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Williams, V.A.; Gellersen, O.; Jones, C.; Watson, T.J.; Peters, J.H. The pathogenesis of Barrett’s esophagus: Secondary bile acids upregulate intestinal differentiation factor CDX2 expression in esophageal cells. J. Gastrointest. Surg. 2007, 11, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Kazumori, H.; Ishihara, S.; Rumi, M.A.; Kadowaki, Y.; Kinoshita, Y. Bile acids directly augment caudal related homeobox gene Cdx2 expression in oesophageal keratinocytes in Barrett’s epithelium. Gut 2006, 55, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, M.; Caliot, E.; Pringault, E. Chronic acid exposure leads to activation of the cdx2 intestinal homeobox gene in a long-term culture of mouse esophageal keratinocytes. J. Cell Sci. 2003, 116, 1429–1436. [Google Scholar] [CrossRef] [Green Version]
- Huo, X.; Zhang, X.; Yu, C.; Cheng, E.; Zhang, Q.; Dunbar, K.B.; Pham, T.H.; Lynch, J.P.; Wang, D.H.; Bresalier, R.S.; et al. Aspirin prevents NF-kappaB activation and CDX2 expression stimulated by acid and bile salts in oesophageal squamous cells of patients with Barrett’s oesophagus. Gut 2018, 67, 606–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibodies for Westernblot Analysis | Dilution | Company | Catalogue Number |
|---|---|---|---|
| β-Actin | 1:2500 (0.5% LFM/PBS) | Sigma-Aldrich, Taufkirchen, Germany | A1978 |
| NF-κB p65 | 1:1000 (1% BSA/TBST) | Cell Signaling Technology, Danvers, USA | 8242 |
| Phospho-NFκB p65 (Ser536) | 1:1000 (1% BSA/TBST) | Cell Signaling Technology, Danvers, USA | 3033 |
| Nrf2 | 1:1000 (1% BSA/TBST) | Abnova, Taipeh, Taiwan | MAB20252 |
| Histone H3 | 1:1000 (1% BSA/TBST) | Cell Signaling Technology, Danvers, USA | 4499S |
| Secondary antibodies for Westernblot analysis | |||
| Goat anti-rabbit | 1:7500 (0.5% LFM) | Jackson Immuno Research, Suffolk, UK | 111-035-045 |
| Goat anti-mouse | 1:7500 (0.5% LFM). | Jackson Immuno Research, Suffolk, UK | 115-035-068 |
| Primary antibodies for Immunohistochemistry | |||
| HO1 | 1:100 (1% BSA/PBS) | GeneTex, Inc., Irvine, USA | GTX101147 |
| Secondary antibodies for Immunohistochemistry | |||
| Goat anti-rabbit | 1:500 (1% BSA/PBS) | Jackson Immuno Research, Suffolk, UK | 111-035-045 |
| Name | Forward | Reverse | Reference | Length (bp) |
|---|---|---|---|---|
| β-Actin | GTCTTCCCCTCCATCGTG | AGGGTGAGGATGCCTCTCTT | NM_001101.3 | 113 |
| NRF2 | TCTTGCCTCCAAAGTATGTCAA | ACACGGTCCACAGCTCATC | NM_006164 | 99 |
| HO1 | GAGTGTAAGGACCCATCGGA | GCCAGCAACAAAGTGCAAG | NM_002133 | 105 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Storz, L.; Walther, P.; Chemnitzer, O.; Lyros, O.; Niebisch, S.; Mehdorn, M.; Jansen-Winkeln, B.; Moulla, Y.; Büch, T.; Gockel, I.; et al. Nrf2/Keap1-Pathway Activation and Reduced Susceptibility to Chemotherapy Treatment by Acidification in Esophageal Adenocarcinoma Cells. Cancers 2021, 13, 2806. https://doi.org/10.3390/cancers13112806
Storz L, Walther P, Chemnitzer O, Lyros O, Niebisch S, Mehdorn M, Jansen-Winkeln B, Moulla Y, Büch T, Gockel I, et al. Nrf2/Keap1-Pathway Activation and Reduced Susceptibility to Chemotherapy Treatment by Acidification in Esophageal Adenocarcinoma Cells. Cancers. 2021; 13(11):2806. https://doi.org/10.3390/cancers13112806
Chicago/Turabian StyleStorz, Lucie, Philipp Walther, Olga Chemnitzer, Orestis Lyros, Stefan Niebisch, Matthias Mehdorn, Boris Jansen-Winkeln, Yusef Moulla, Thomas Büch, Ines Gockel, and et al. 2021. "Nrf2/Keap1-Pathway Activation and Reduced Susceptibility to Chemotherapy Treatment by Acidification in Esophageal Adenocarcinoma Cells" Cancers 13, no. 11: 2806. https://doi.org/10.3390/cancers13112806