
Genetic Patterns Found in the Nuclear Localization Signals (NLSs) Associated with EBV-1 and EBV-2 Provide New Insights into Their Contribution to Different Cell-Type Specificities

 ,
,  , , ,
, , ,  and
and {kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fresh-Frozen Gastric Cancer Tissues for PCR Analysis
2.2. DNA Extraction and PCR Assay
2.3. Sequence Dataset
2.4. Recombination and Phylogenetic Analyses
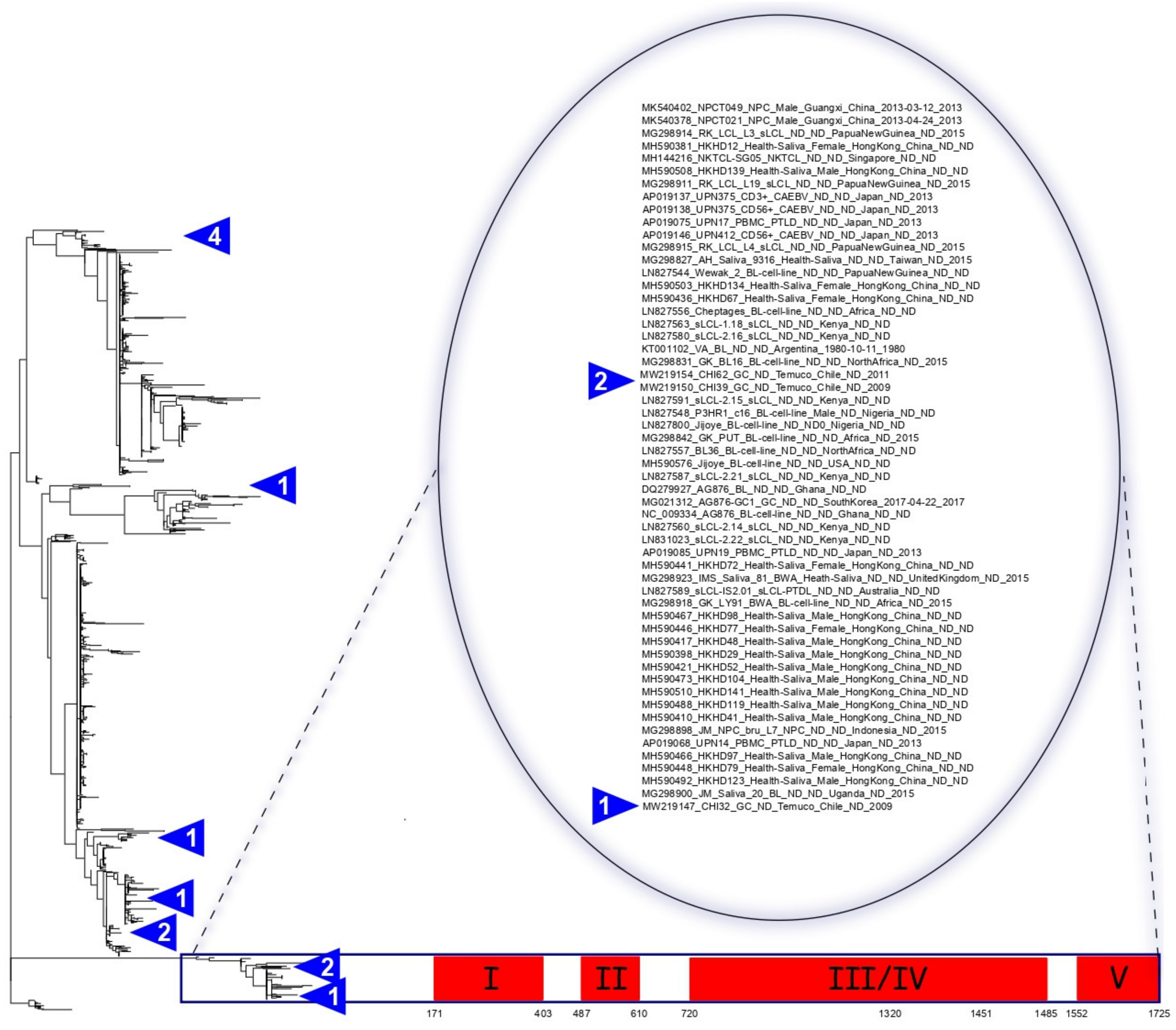
3. Results
3.1. EBV-Associated Gastric Cancer from Chile
3.2. EBNA3A Recovery from Genomes
3.3. Classifying EBV According to Traditional Classifications
3.4. South American EBV Characteristics
3.5. Two Non-Canonical NLSs Could Be Related to the Lower Efficiency of EBV-2 in Converting B Cells into Lymphoblastoid Cell Lines
3.6. Characteristic Deletions of EBV-2 in the EBNA3A Gene
3.7. Unique Type 2 Signatures Retained in the Intron Region of EBNA3A
3.8. EBNA3A Recombination Events Reside Exclusively in EBV-2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vetsika, E.-K.; Callan, M. Infectious mononucleosis and Epstein-Barr virus. Expert Rev. Mol. Med. 2004, 6, 1–16. [Google Scholar] [CrossRef]
- Gunvén, P.; Klein, G.; Henle, G.; Henle, W.; Clifford, P. Epstein–Barr Virus in Burkitt’s Lymphoma and Nasopharyngeal Carcinoma: Antibodies to EBV associated Membrane and Viral Capsid Antigens in Burkitt Lymphoma Patients. Nat. Cell Biol. 1970, 228, 1053–1056. [Google Scholar] [CrossRef] [PubMed]
- Tsao, S.W.; Yip, Y.L.; Tsang, C.M.; Pang, P.S.; Lau, V.M.Y.; Zhang, G.; Lo, K.W. Etiological factors of nasopharyngeal carcinoma. Oral Oncol. 2014, 50, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Corvalan, A.; Koriyama, C.; Akiba, S.; Eizuru, Y.; Backhouse, C.; Palma, M.; Argandoña, J.; Tokunaga, M. Epstein-Barr virus in gastric carcinoma is associated with location in the cardia and with a diffuse histology: A study in one area of Chile. Int. J. Cancer 2001, 94, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Baer, R.; Bankier, A.T.; Biggin, M.D.; Deininger, P.; Farrell, P.J.; Gibson, T.J.; Hatfull, G.; Hudson, G.S.; Satchwell, S.C.; Séguin, C.; et al. DNA sequence and expression of the B95-8 Epstein—Barr virus genome. Nat. Cell Biol. 1984, 310, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Palser, A.L.; Grayson, N.E.; White, R.E.; Corton, C.; Correia, S.; Abdullah, M.M.B.; Watson, S.J.; Cotten, M.; Arrand, J.R.; Murray, P.G.; et al. Genome Diversity of Epstein-Barr Virus from Multiple Tumor Types and Normal Infection. J. Virol. 2015, 89, 5222–5237. [Google Scholar] [CrossRef] [Green Version]
- Sample, J.; Young, L.; Martin, B.; Chatman, T.; Kieff, E.; Rickinson, A. Epstein-Barr virus types 1 and 2 differ in their EBNA-3A, EBNA-3B, and EBNA-3C genes. J. Virol. 1990, 64, 4084–4092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucchesi, W.; Brady, G.; Dittrich-Breiholz, O.; Kracht, M.; Russ, R.; Farrell, P.J. Differential Gene Regulation by Epstein-Barr Virus Type 1 and Type 2 EBNA2. J. Virol. 2008, 82, 7456–7466. [Google Scholar] [CrossRef] [Green Version]
- Ponnusamy, R.; Khatri, R.; Correia, P.B.; Wood, C.D.; Mancini, E.J.; Farrell, P.J.; West, M.J. Increased association between Epstein-Barr virus EBNA2 from type 2 strains and the transcriptional repressor BS69 restricts EBNA2 activity. PLoS Pathog. 2019, 15, e1007458. [Google Scholar] [CrossRef]
- Coleman, C.B.; Lang, J.; Sweet, L.A.; Smith, N.A.; Freed, B.M.; Pan, Z.; Haverkos, B.; Pelanda, R.; Rochford, R. Epstein-Barr Virus Type 2 Infects T Cells and Induces B Cell Lymphomagenesis in Humanized Mice. J. Virol. 2018, 92, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Coleman, C.B.; Wohlford, E.M.; Smith, N.A.; King, C.A.; Ritchie, J.A.; Baresel, P.C.; Kimura, H.; Rochford, R. Epstein-Barr Virus Type 2 Latently Infects T Cells, Inducing an Atypical Activation Characterized by Expression of Lymphotactic Cytokines. J. Virol. 2015, 89, 2301–2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cludts, I.; Farrell, P.J. Multiple Functions within the Epstein-Barr Virus EBNA-3A Protein. J. Virol. 1998, 72, 1862–1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, R.E.; Groves, I.J.; Turro, E.; Yee, J.; Kremmer, E.; Allday, M.J. Extensive Co-Operation between the Epstein-Barr Virus EBNA3 Proteins in the Manipulation of Host Gene Expression and Epigenetic Chromatin Modification. PLoS ONE 2010, 5, e13979. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, M.; Holthaus, A.M.; Calderwood, M.A.; Lai, C.-Y.; Krastins, B.; Sarracino, D.; Johannsen, E. The EBNA3 Family of Epstein-Barr Virus Nuclear Proteins Associates with the USP46/USP12 Deubiquitination Complexes to Regulate Lymphoblastoid Cell Line Growth. PLoS Pathog. 2015, 11, e1004822. [Google Scholar] [CrossRef] [PubMed]
- Allday, M.J.; Bazot, Q.; White, R.E. The EBNA3 Family: Two Oncoproteins and a Tumour Suppressor that Are Central to the Biology of EBV in B Cells. Curr. Top. Microbiol. Immunol. 2015, 391, 61–117. [Google Scholar] [CrossRef]
- Maruo, S.; Johannsen, E.; Illanes, D.; Cooper, A.; Zhao, B.; Kieff, E. Epstein-Barr Virus Nuclear Protein 3A Domains Essential for Growth of Lymphoblasts: Transcriptional Regulation through RBP-Jk/CBF1 Is Critical. J. Virol. 2005, 79, 10171–10179. [Google Scholar] [CrossRef] [Green Version]
- Hickabottom, M.; Parker, G.A.; Freemont, P.S.; Crook, T.; Allday, M.J. Two Nonconsensus Sites in the Epstein-Barr Virus Oncoprotein EBNA3A Cooperate to Bind the Co-repressor Carboxyl-terminal-binding Protein (CtBP). J. Biol. Chem. 2002, 277, 47197–47204. [Google Scholar] [CrossRef] [Green Version]
- Bazot, Q.; Paschos, K.; Allday, M.J. Epstein-Barr Virus (EBV) Latent Protein EBNA3A Directly Targets and Silences the STK39 Gene in B Cells Infected by EBV. J. Virol. 2018, 92, e01918-17. [Google Scholar] [CrossRef] [Green Version]
- Burgess, A.; Buck, M.; Krauer, K.; Sculley, T. Nuclear localization of the Epstein–Barr virus EBNA3B protein. J. Gen. Virol. 2006, 87, 789–793. [Google Scholar] [CrossRef]
- Cai, M.; Liao, Z.; Chen, T.; Wang, P.; Zou, X.; Wang, Y.; Xu, Z.; Jiang, S.; Huang, J.; Chen, D.; et al. Characterization of the subcellular localization of Epstein-Barr virus encoded proteins in live cells. Oncotarget 2017, 8, 70006–70034. [Google Scholar] [CrossRef] [Green Version]
- Buck, M.; Burgess, A.; Stirzaker, R.; Krauer, K.; Sculley, T. Epstein–Barr virus nuclear antigen 3A contains six nuclear-localization signals. J. Gen. Virol. 2006, 87, 2879–2884. [Google Scholar] [CrossRef]
- Zanella, L.; Riquelme, I.; Buchegger, K.; Abanto, M.; Ili, C.; Brebi, P. A reliable Epstein-Barr Virus classification based on phylogenomic and population analyses. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- De Jesus, O.; Smith, P.R.; Spender, L.C.; Karstegl, C.E.; Niller, H.H.; Huang, D.; Farrell, P.J. Updated Epstein–Barr virus (EBV) DNA sequence and analysis of a promoter for the BART (CST, BARF0) RNAs of EBV. J. Gen. Virol. 2003, 84, 1443–1450. [Google Scholar] [CrossRef] [PubMed]
- Dolan, A.; Addison, C.; Gatherer, D.; Davison, A.J.; McGeoch, D.J. The genome of Epstein–Barr virus type 2 strain AG. Virology 2006, 350, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Croucher, N.J.; Page, A.; Connor, T.R.; Delaney, A.J.; Keane, J.A.; Bentley, S.D.; Parkhill, J.; Harris, S.R. Rapid phylogenetic analysis of large samples of recombinant bacterial whole genome sequences using Gubbins. Nucleic Acids Res. 2015, 43, e15. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. Evaluation of methods for detecting recombination from DNA sequences: Computer simulations. Proc. Natl. Acad. Sci. USA 2001, 98, 13757–13762. [Google Scholar] [CrossRef] [Green Version]
- Padidam, M.; Sawyer, S.; Fauquet, C.M. Possible Emergence of New Geminiviruses by Frequent Recombination. Virology 1999, 265, 218–225. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, M.J.; Armstrong, J.S.; Gibbs, A.J. Sister-Scanning: A Monte Carlo procedure for assessing signals in recombinant sequences. Bioinformation 2000, 16, 573–582. [Google Scholar] [CrossRef]
- Boni, M.F.; Posada, D.; Feldman, M.W. An Exact Nonparametric Method for Inferring Mosaic Structure in Sequence Triplets. Genetics 2007, 176, 1035–1047. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, M.M.B.; Palermo, R.D.; Palser, A.L.; Grayson, N.E.; Kellam, P.; Correia, S.; Szymula, A.; White, R.E. Heterogeneity of the Epstein-Barr Virus (EBV) Major Internal Repeat Reveals Evolutionary Mechanisms of EBV and a Functional Defect in the Prototype EBV Strain B95. J. Virol. 2017, 91, 00920-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berenstein, A.J.; Lorenzetti, M.A.; Preciado, M.V.; José, B.A.; Alejandro, L.M.; Victoria, P.M. Recombination rates along the entire Epstein Barr virus genome display a highly heterogeneous landscape. Infect. Genet. Evol. 2018, 65, 96–103. [Google Scholar] [CrossRef]
- Görzer, I.; Niesters, H.G.; Cornelissen, J.J.; Puchhammer-Stöckl, E. Characterization of Epstein-Barr virus Type I variants based on linked polymorphism among EBNA3A, -3B, and -3C genes. Virus Res. 2006, 118, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Gantuz, M.; Lorenzetti, M.A.; Chabay, P.A.; Preciado, M.V. A novel recombinant variant of latent membrane protein 1 from Epstein Barr virus in Argentina denotes phylogeographical association. PLoS ONE 2017, 12, e0174221. [Google Scholar] [CrossRef] [PubMed]
- Kwok, H.; Wu, C.W.; Palser, A.L.; Kellam, P.; Sham, P.C.; Kwong, D.L.W.; Chiang, A.K.S. Genomic Diversity of Epstein-Barr Virus Genomes Isolated from Primary Nasopharyngeal Carcinoma Biopsy Samples. J. Virol. 2014, 88, 10662–10672. [Google Scholar] [CrossRef] [Green Version]
- Santpere, G.; Darre, F.; Blanco, S.; Alcami, A.; Villoslada, P.; Albà, M.M.; Navarro, A. Genome-Wide Analysis of Wild-Type Epstein–Barr Virus Genomes Derived from Healthy Individuals of the 1000 Genomes Project. Genome Biol. Evol. 2014, 6, 846–860. [Google Scholar] [CrossRef] [Green Version]
- Young, L.S.; Rickinson, A.B. Epstein–Barr virus: 40 years on. Nat. Rev. Cancer 2004, 4, 757–768. [Google Scholar] [CrossRef]
- Saha, A.; Robertson, E.S. Epstein-Barr Virus–Associated B-cell Lymphomas: Pathogenesis and Clinical Outcomes. Clin. Cancer Res. 2011, 17, 3056–3063. [Google Scholar] [CrossRef] [Green Version]
- Tomkinson, B.; Robertson, E.; Kieff, E. Epstein-Barr virus nuclear proteins EBNA-3A and EBNA-3C are essential for B-lymphocyte growth transformation. J. Virol. 1993, 67, 2014–2025. [Google Scholar] [CrossRef] [Green Version]
- Young, L.S.; Murray, P.G. Epstein–Barr virus and oncogenesis: From latent genes to tumours. Oncogene 2003, 22, 5108–5121. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.-S.; Kieff, E. Epstein–Barr virus latent genes. Exp. Mol. Med. 2015, 47, e131. [Google Scholar] [CrossRef] [Green Version]
- Grossman, S.R.; Johannsen, E.; Tong, X.; Yalamanchili, R.; Kieff, E. The Epstein-Barr virus nuclear antigen 2 transactivator is directed to response elements by the J kappa recombination signal binding protein. Proc. Natl. Acad. Sci. USA 1994, 91, 7568–7572. [Google Scholar] [CrossRef] [Green Version]
- Hertle, M.L.; Popp, C.; Petermann, S.; Maier, S.; Kremmer, E.; Lang, R.; Mages, J.; Kempkes, B. Differential Gene Expression Patterns of EBV Infected EBNA-3A Positive and Negative Human B Lymphocytes. PLoS Pathog. 2009, 5, e1000506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruo, S.; Zhao, B.; Johannsen, E.; Kieff, E.; Zou, J.; Takada, K. Epstein-Barr virus nuclear antigens 3C and 3A maintain lymphoblastoid cell growth by repressing p16INK4Aand p14ARFexpression. Proc. Natl. Acad. Sci. USA 2011, 108, 1919–1924. [Google Scholar] [CrossRef] [Green Version]
- Murer, A.; McHugh, D.; Caduff, N.; Kalchschmidt, J.; Barros, M.; Zbinden, A.; Capaul, R.; Niedobitek, G.; Allday, M.; Chijioke, O.; et al. EBV persistence without its EBNA3A and 3C oncogenes in vivo. PLoS Pathog. 2018, 14, e1007039. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Welch, R.; Zhao, B.; Ta, T.; Keleş, S.; Johannsen, E. Epstein-Barr Virus Nuclear Antigen 3 (EBNA3) Proteins Regulate EBNA2 Binding to Distinct RBPJ Genomic Sites. J. Virol. 2016, 90, 2906–2919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kienzle, N.; Young, D.B.; Liaskou, D.; Buck, M.; Greco, S.; Sculley, T.B. Intron retention may regulate expression of Epstein-Barr virus nuclear antigen 3 family genes. J. Virol. 1999, 73, 1195–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimber, U.; Adldinger, H.K.; Lenoir, G.M.; Vuillaume, M.; Knebel-Doeberitz, M.V.; Laux, G.; Deśgranges, C.; Wittmann, P.; Freese, U.-K.; Schneider, U.; et al. Geographical prevalence of two types of Epstein-Barr virus. Virology 1986, 154, 56–66. [Google Scholar] [CrossRef]
- Correia, S.; Bridges, R.; Wegner, F.; Venturini, C.; Palser, A.; Middeldorp, J.; Cohen, J.I.; Lorenzetti, M.A.; Bassano, I.; White, R.E.; et al. Sequence Variation of Epstein-Barr Virus: Viral Types, Geography, Codon Usage, and Diseases. J. Virol. 2018, 92, e01132-18. [Google Scholar] [CrossRef] [Green Version]
- Khanna, R.; Slade, R.W.; Poulsen, L.; Moss, D.J.; Burrows, S.R.; Nicholls, J.; Burrows, J.M. Evolutionary dynamics of genetic variation in Epstein-Barr virus isolates of diverse geographical origins: Evidence for immune pressure-independent genetic drift. J. Virol. 1997, 71, 8340–8346. [Google Scholar] [CrossRef] [Green Version]
- Mandage, R.; Telford, M.; Rodríguez, J.A.; Farré, X.; Layouni, H.; Marigorta, U.M.; Cundiff, C.; Heredia-Genestar, J.M.; Navarro, A.; Santpere, G. Genetic factors affecting EBV copy number in lymphoblastoid cell lines derived from the 1000 Genome Project samples. PLoS ONE 2017, 12, e0179446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Midgley, R.S.; Bell, A.I.; McGeoch, D.J.; Rickinson, A.B. Latent Gene Sequencing Reveals Familial Relationships among Chinese Epstein-Barr Virus Strains and Evidence for Positive Selection of A11 Epitope Changes. J. Virol. 2003, 77, 11517–11530. [Google Scholar] [CrossRef] [Green Version]
- Bazot, Q.; Deschamps, T.; Tafforeau, L.; Siouda, M.; Leblanc, P.; Harth-Hertle, M.L.; Rabourdin-Combe, C.; Lotteau, V.; Kempkes, B.; Tommasino, M.; et al. Epstein–Barr virus nuclear antigen 3A protein regulates CDKN2B transcription via interaction with MIZ. Nucleic Acids Res. 2014, 42, 9700–9716. [Google Scholar] [CrossRef] [PubMed]
- Krauer, K.; Buck, M.; Flanagan, J.; Belzer, D.; Sculley, T. Identification of the nuclear localization signals within the Epstein–Barr virus EBNA-6 protein. J. Gen. Virol. 2004, 85, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Makkerh, J.P.; Dingwall, C.; Laskey, R.A. Comparative mutagenesis of nuclear localization signals reveals the importance of neutral and acidic amino acids. Curr. Biol. 1996, 6, 1025–1027. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.-Y.; Jans, P.; Jans, D.A. Negative charge at the protein kinase CK2 site enhances recognition of the SV40 large T-antigen NLS by importin: Effect of conformation. FEBS Lett. 1998, 440, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Yao, Q.Y.; Tierney, R.J.; Croom-Carter, D.; Cooper, G.M.; Ellis, C.J.; Rowe, M.; Rickinson, A.B. Isolation of intertypic recombinants of Epstein-Barr virus from T-cell-immunocompromised individuals. J. Virol. 1996, 70, 4895–4903. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Kim, K.P. Log-likelihood-ratio-based detection ordering in V-BLAST. IEEE Trans. Commun. 2006, 54, 302–307. [Google Scholar] [CrossRef]
- McGeoch, D.J.; Gatherer, D. Lineage structures in the genome sequences of three Epstein–Barr virus strains. Virology 2007, 359, 1–5. [Google Scholar] [CrossRef]
- Midgley, R.S.; Blake, N.W.; Yao, Q.Y.; Croom-Carter, D.; Cheung, S.T.; Leung, S.F.; Chan, A.T.C.; Johnson, P.J.; Huang, D.; Rickinson, A.B.; et al. Novel Intertypic Recombinants of Epstein-Barr Virus in the Chinese Population. J. Virol. 2000, 74, 1544–1548. [Google Scholar] [CrossRef] [Green Version]
- Coleman, C.B.; Daud, I.I.; Ogolla, S.O.; Ritchie, J.A.; Smith, N.A.; Sumba, P.O.; Dent, A.E.; Rochford, R. Epstein-Barr Virus Type 2 Infects T Cells in Healthy Kenyan Children. J. Infect. Dis. 2017, 216, 670–677. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zanella, L.; Reyes, M.E.; Riquelme, I.; Abanto, M.; León, D.; Viscarra, T.; Ili, C.; Brebi, P. Genetic Patterns Found in the Nuclear Localization Signals (NLSs) Associated with EBV-1 and EBV-2 Provide New Insights into Their Contribution to Different Cell-Type Specificities. Cancers 2021, 13, 2569. https://doi.org/10.3390/cancers13112569
Zanella L, Reyes ME, Riquelme I, Abanto M, León D, Viscarra T, Ili C, Brebi P. Genetic Patterns Found in the Nuclear Localization Signals (NLSs) Associated with EBV-1 and EBV-2 Provide New Insights into Their Contribution to Different Cell-Type Specificities. Cancers. 2021; 13(11):2569. https://doi.org/10.3390/cancers13112569
Chicago/Turabian StyleZanella, Louise, María Elena Reyes, Ismael Riquelme, Michel Abanto, Daniela León, Tamara Viscarra, Carmen Ili, and Priscilla Brebi. 2021. "Genetic Patterns Found in the Nuclear Localization Signals (NLSs) Associated with EBV-1 and EBV-2 Provide New Insights into Their Contribution to Different Cell-Type Specificities" Cancers 13, no. 11: 2569. https://doi.org/10.3390/cancers13112569