
Network Pharmacological Analysis through a Bioinformatics Approach of Novel NSC765600 and NSC765691 Compounds as Potential Inhibitors of CCND1/CDK4/PLK1/CD44 in Cancer Types

 ,
, 
 , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Pharmacokinetic (PK), Drug-Likeness, and Medicinal Chemical Analyses
2.2. Identifying the Molecular Targets of NSC765600 and NSC765691
2.3. Bioinformatics Predictions
2.4. Protein-Protein Interaction (PPI) Analysis
2.5. Analysis of Genomic Alterations of CCND1/CDK4/PLK1/CD44 in Multiple Cancer Types
2.6. In Silico Analysis of Molecular Docking of Receptors and Ligands
2.7. In Vitro Screening of NSC765600 and NSC765691 against Full National Cancer Institute (NC I)-60 Cell Panels of Human Tumor Cell Lines
2.8. Data Analysis
3. Results
3.1. NSC765600 and NSC765691 Adhere to the Required Drug-Likeness Criteria
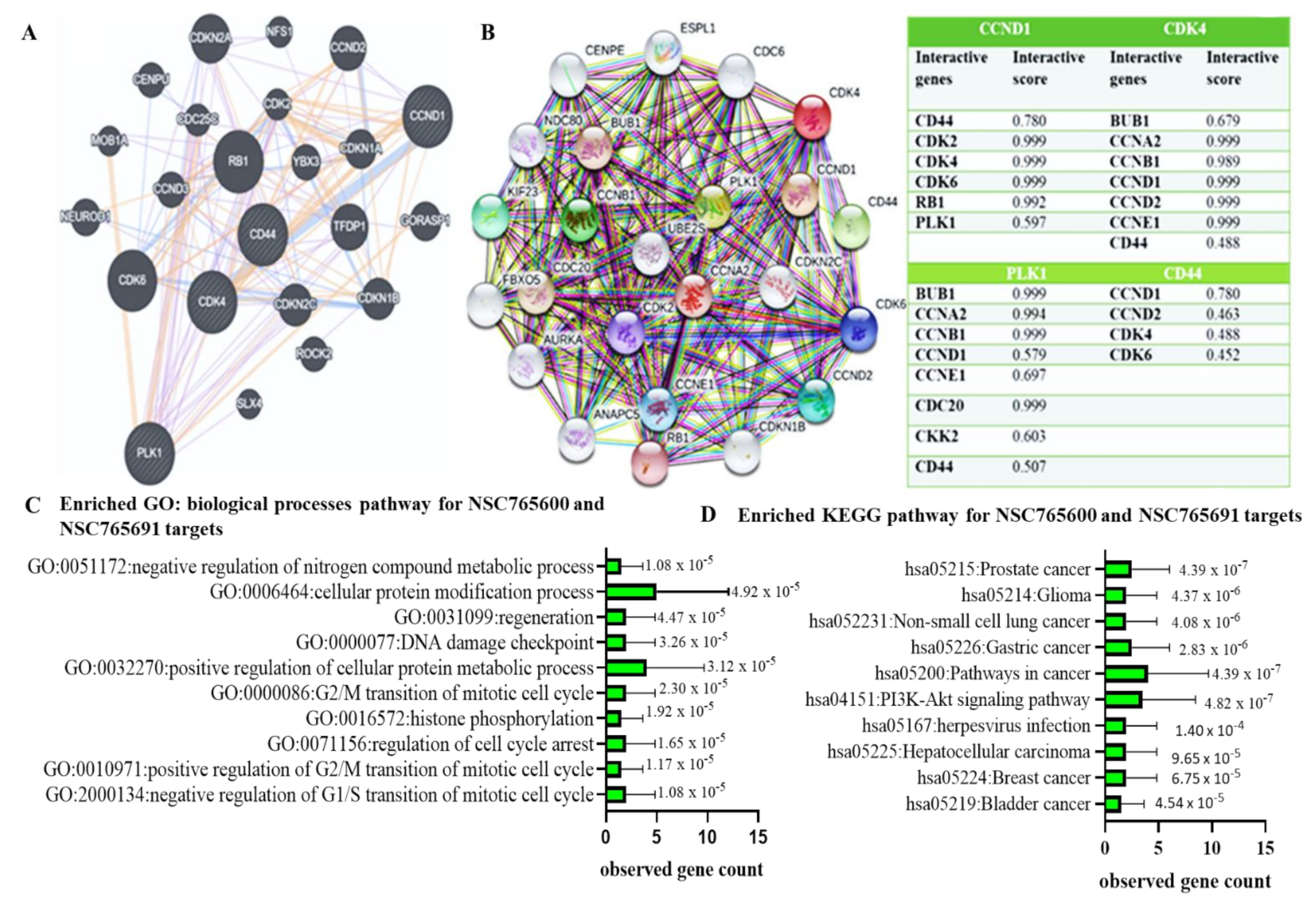
3.2. CCND1/CDK4/PLK1/CD44 Gene Signatures are Potential Drug Targets for NSC765600 and NSC765691
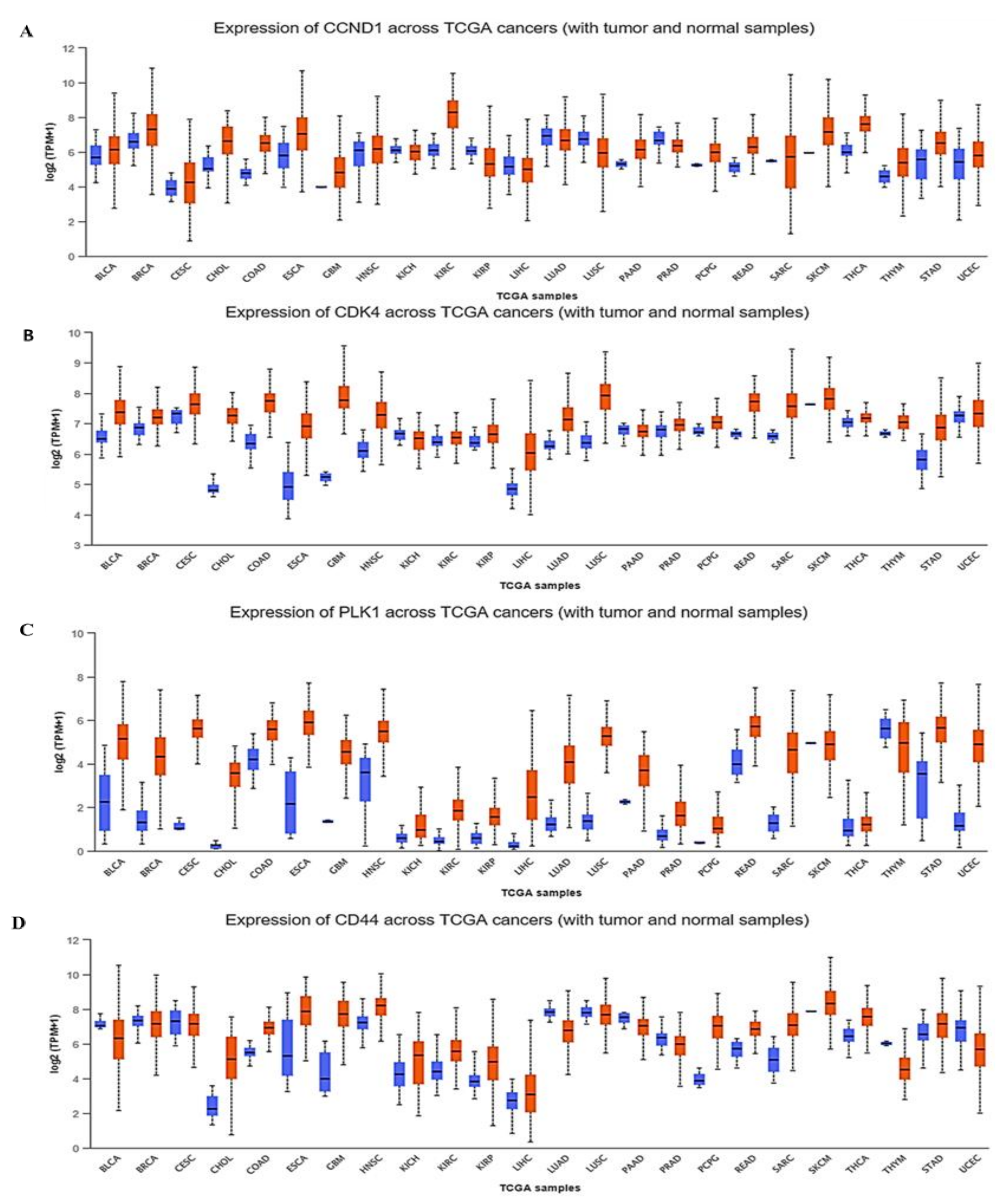
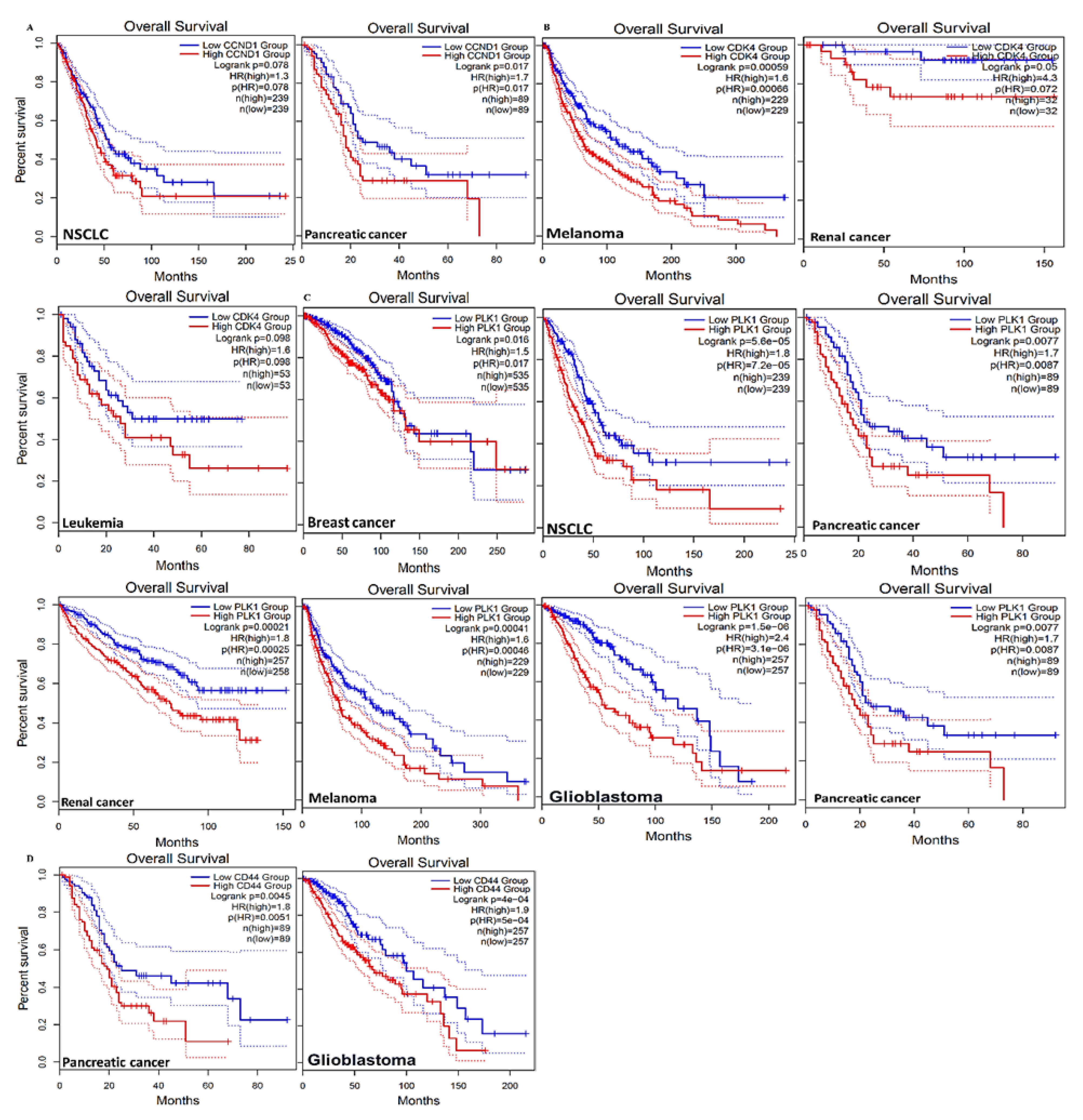
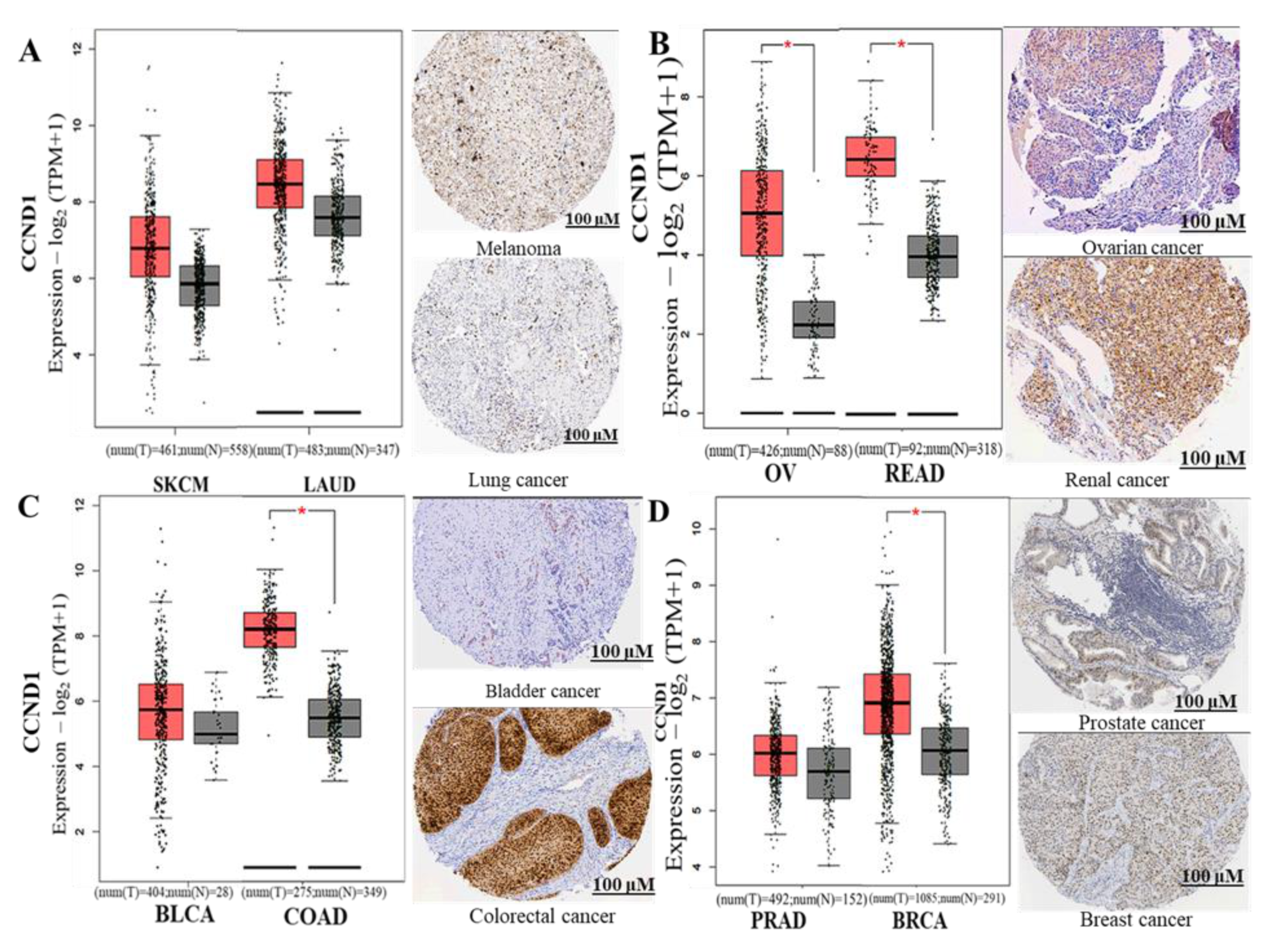
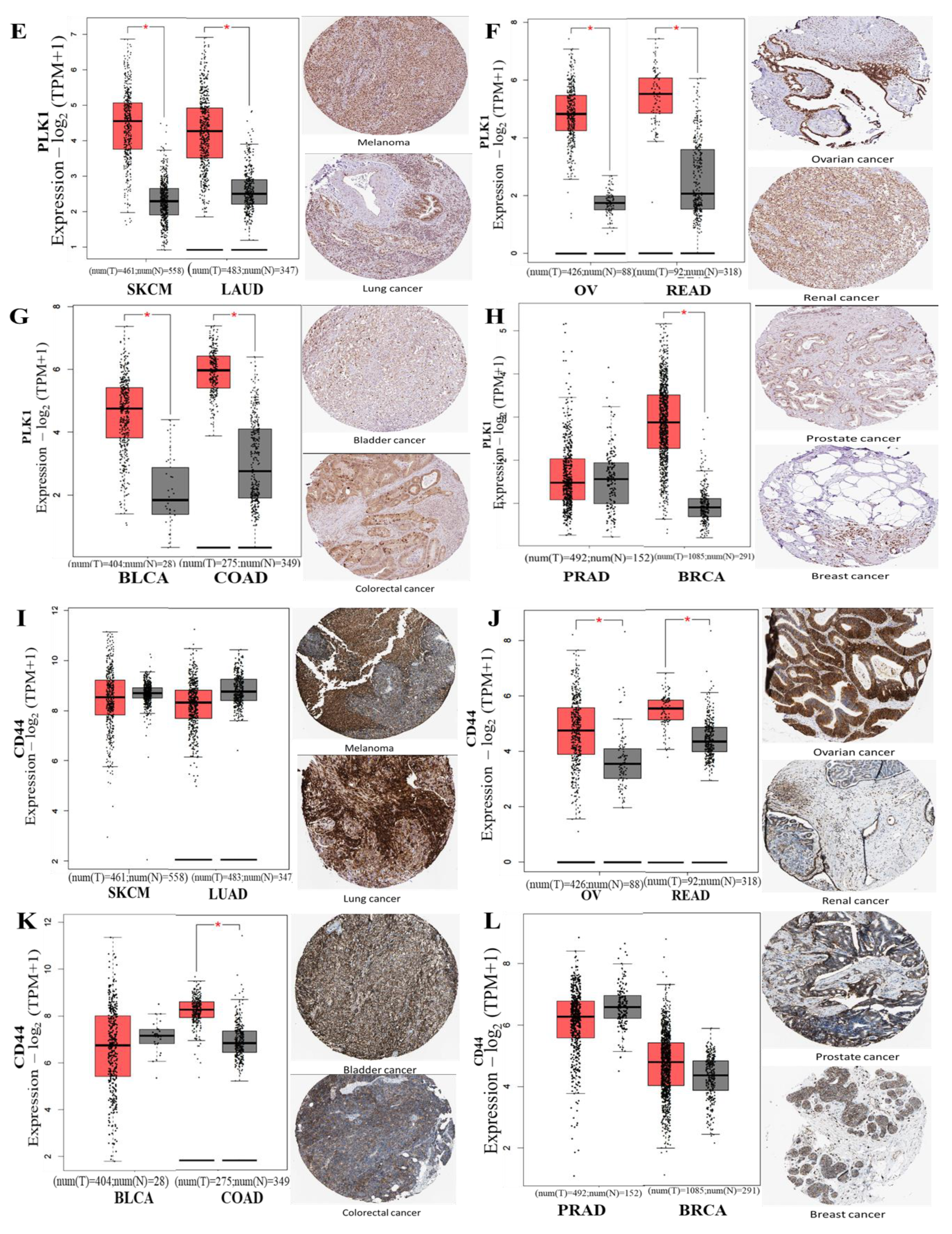
3.3. CCND1/CDK4/PLK1/CD44 are Overexpressed in Multiple Cancers and Associated with Poor Prognoses
3.4. CCND1/CDK4/PLK1/CD44 Genes are Altered in Multiple Cancer Types
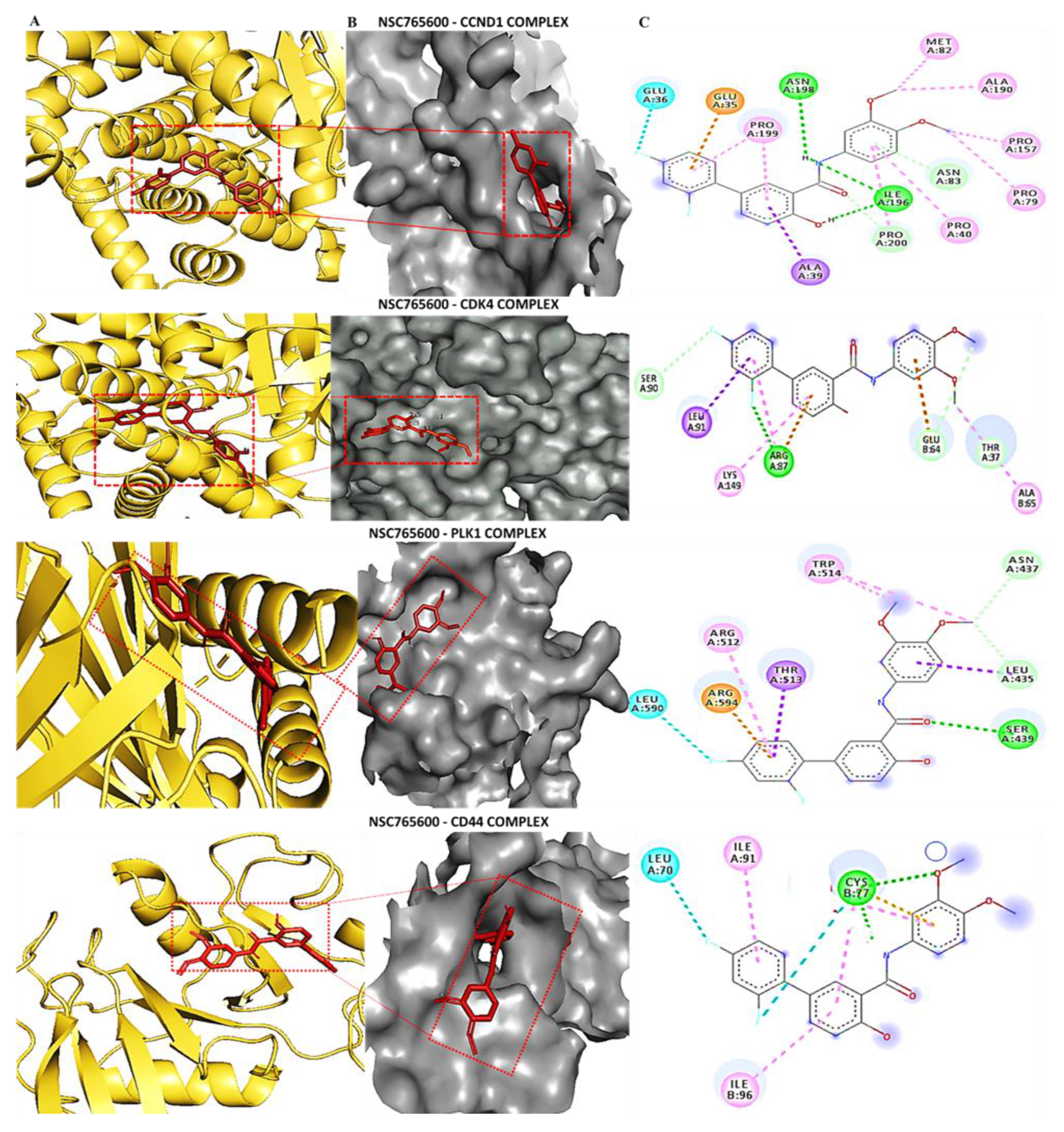
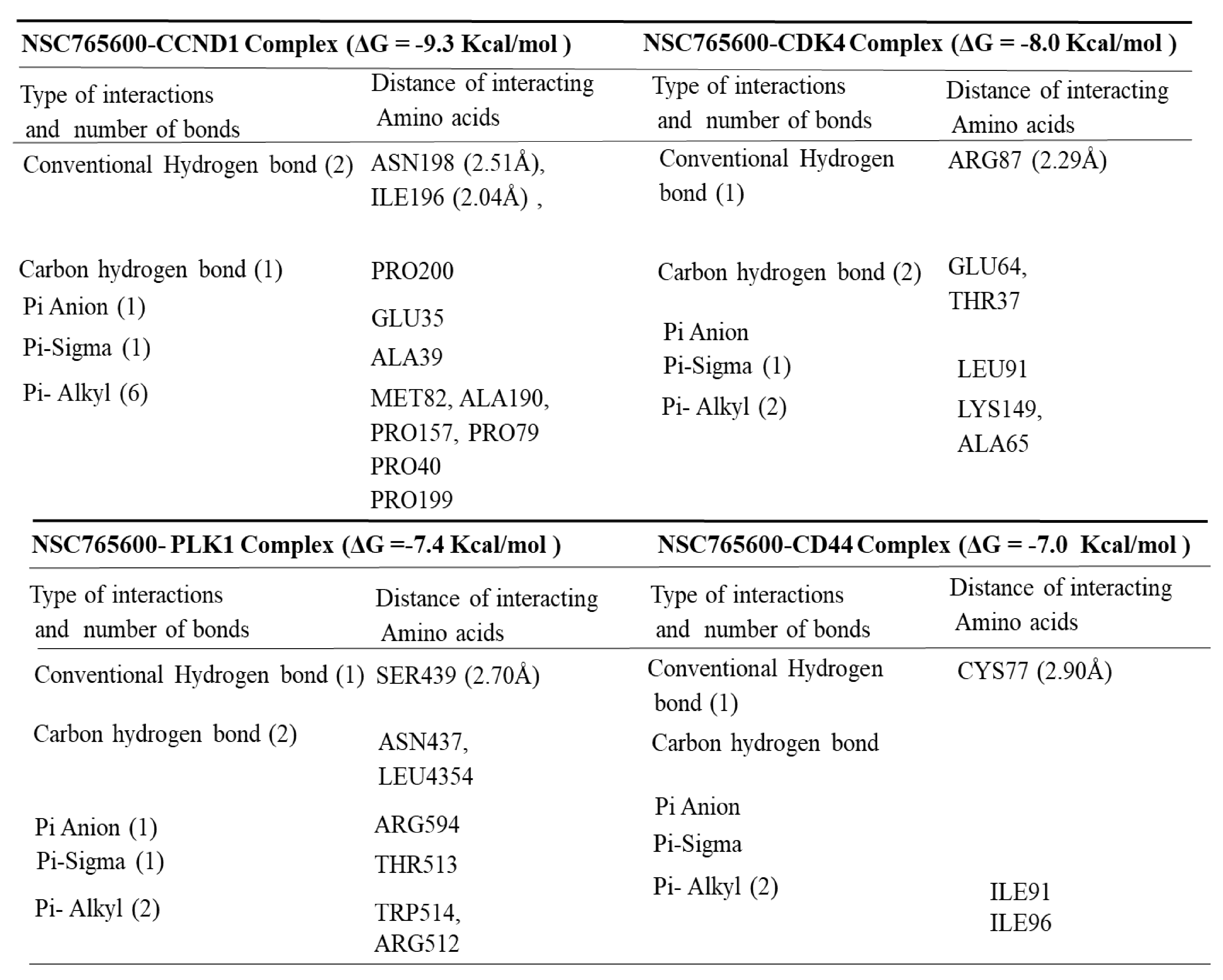
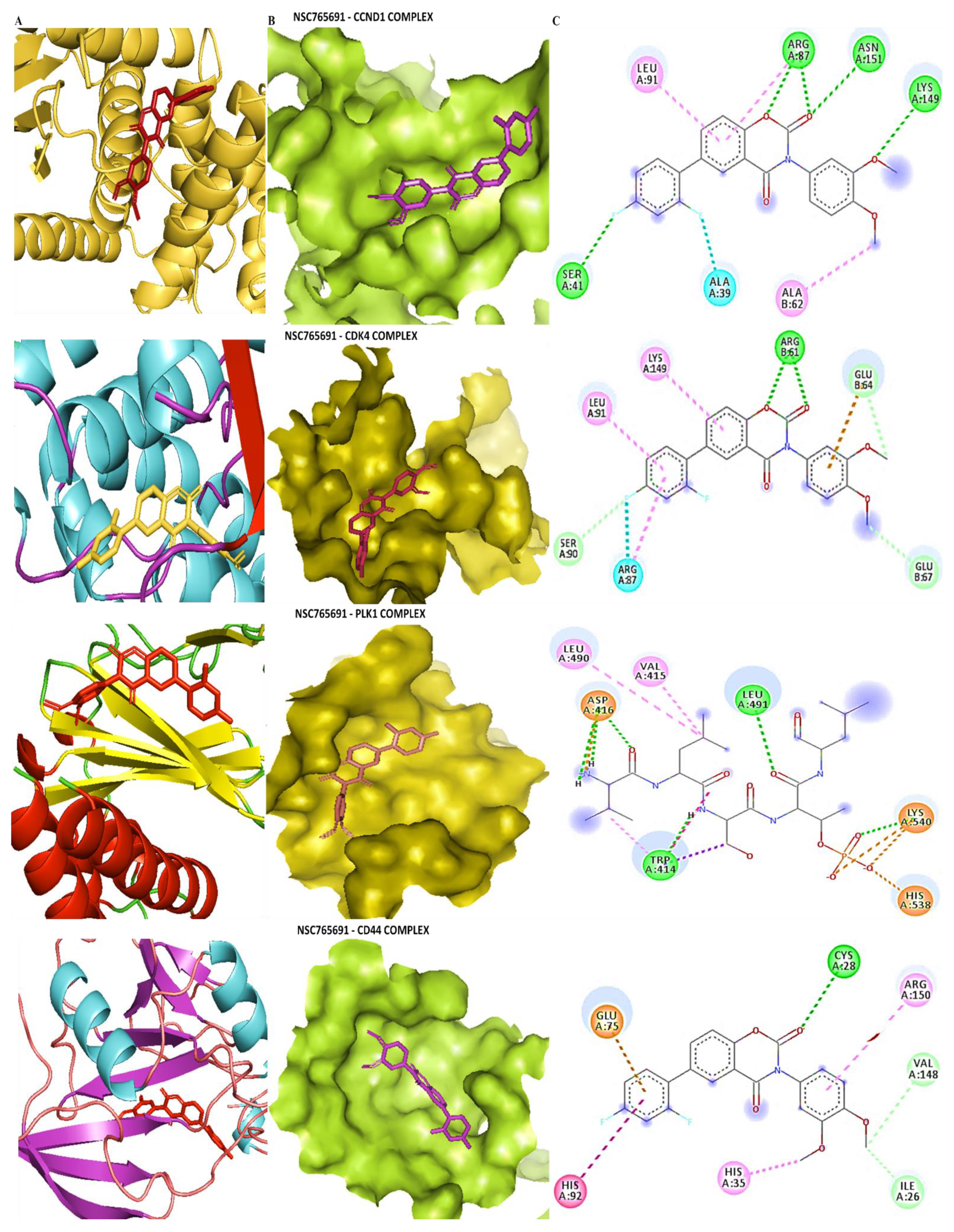
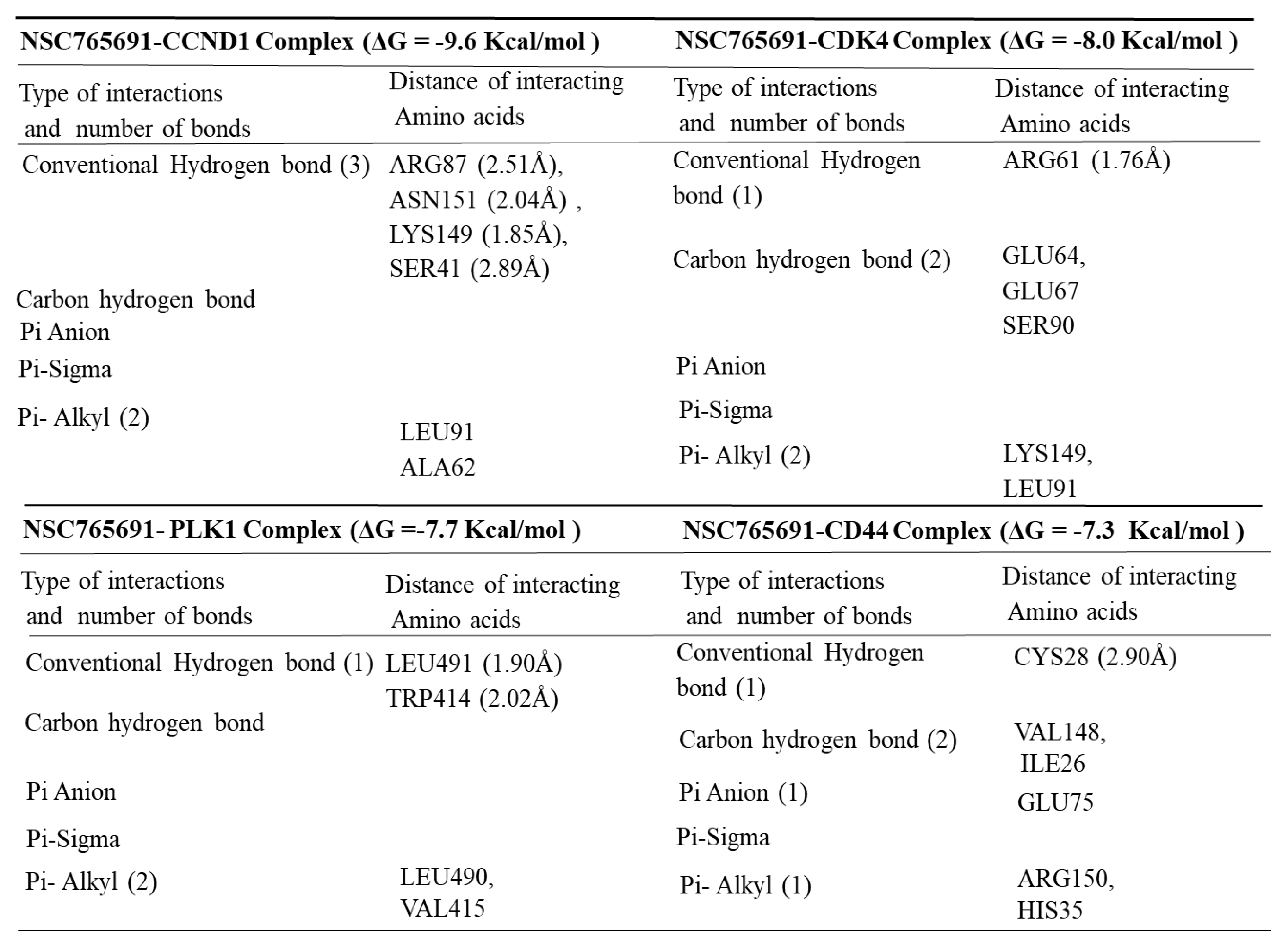
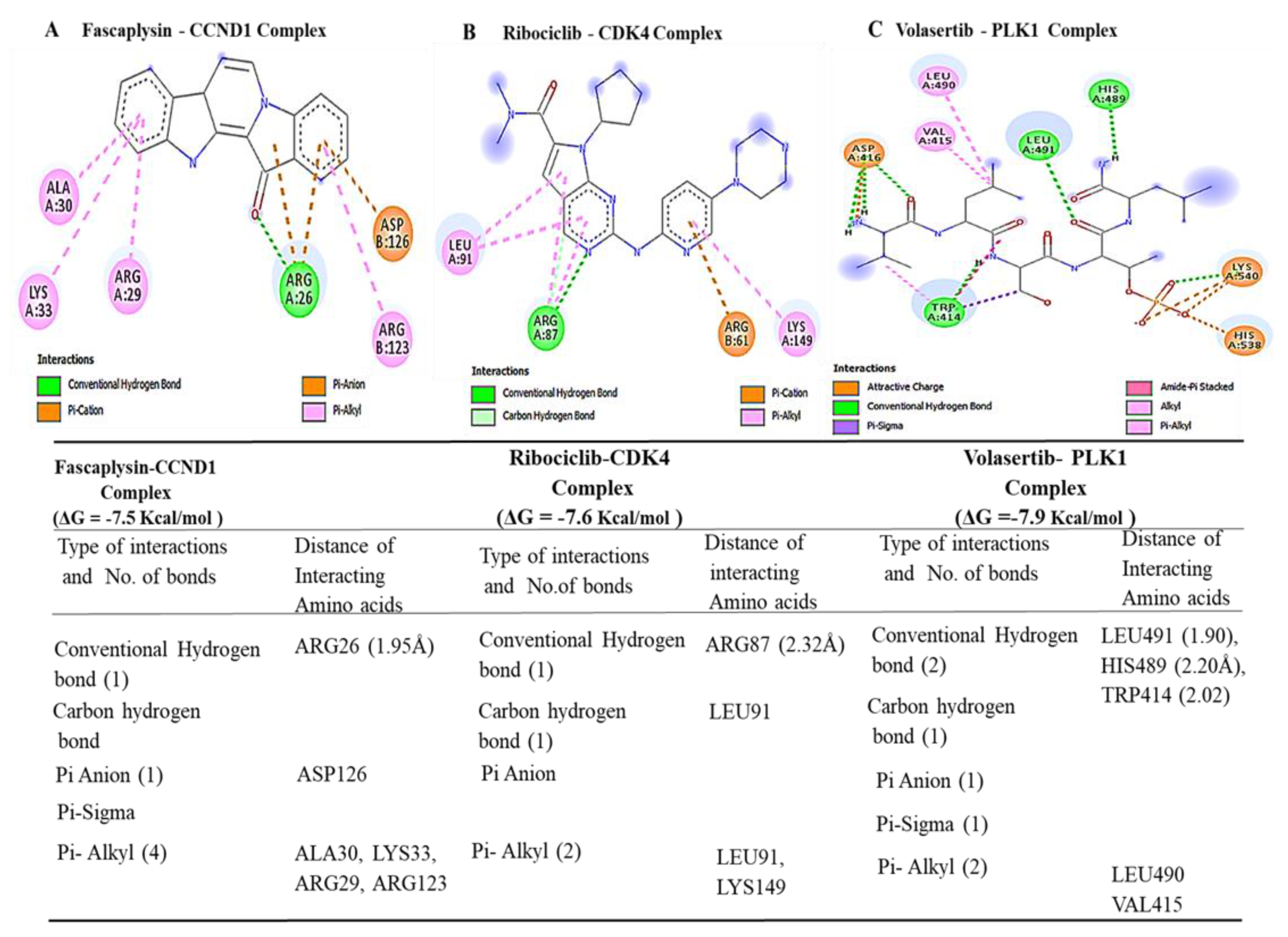
3.5. Determining Protein–Ligand Interactions (PLIs)
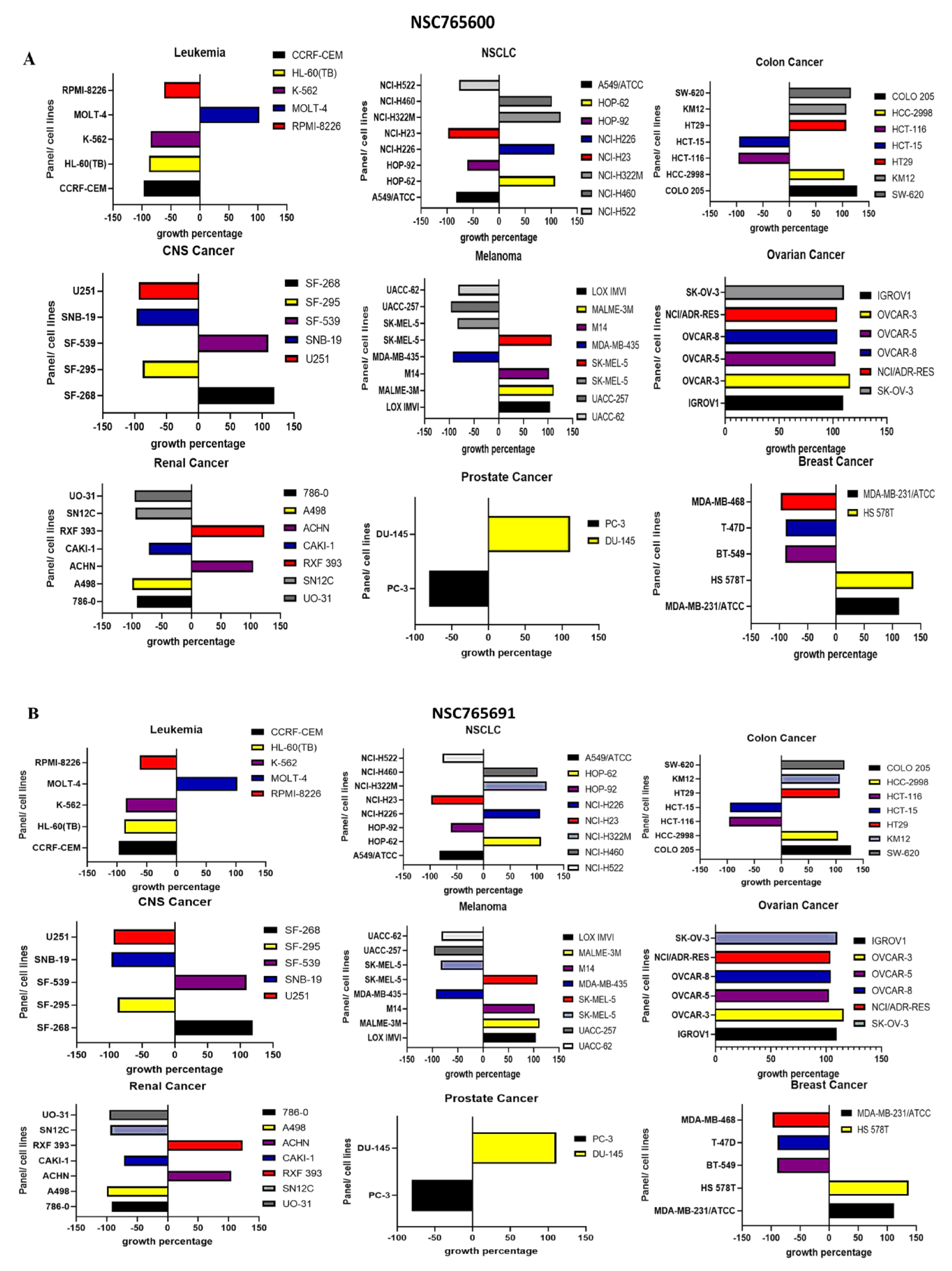
3.6. NSC765600 and NSC765691 Display Antiproliferative and Cytotoxic Effects against a Panel of 60 Human Tumor Cell Lines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christensen, J.F.; Simonsen, C.; Hojman, P. Exercise Training in Cancer Control and Treatment. Compr. Physiol. 2018, 9, 165–205. [Google Scholar] [CrossRef]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2014, 136, E359–E386. [Google Scholar] [CrossRef]
- Rizvi, S.M.D.; Alshammari, A.A.A.; Almawkaa, W.A.; Ahmed, A.B.F.; Katamesh, A.; AlAfnan, A.; Almutairi, T.J.; Alshammari, R.F. An oncoinformatics study to predict the inhibitory potential of recent FDA-approved anti-cancer drugs against human Polo-like kinase 1 enzyme: A step towards dual-target cancer medication. 3 Biotech 2019, 9, 70. [Google Scholar] [CrossRef]
- O’Donnell, J.S.; Teng, M.W.L.; Smyth, M.J. Cancer immunoediting and resistance to T cell-based immunotherapy. Nat. Rev. Clin. Oncol. 2019, 16, 1511–1567. [Google Scholar] [CrossRef] [PubMed]
- Zugazagoitia, J.; Guedes, C.; Ponce, S.; Ferrer, I.; Molina-Pinelo, S.; Paz-Ares, L. Current Challenges in Cancer Treatment. Clin. Ther. 2016, 38, 1551–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, S.J.; Reddy, E.P. CDK4: A Key Player in the Cell Cycle, Development, and Cancer. Genes Cancer 2012, 3, 658–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tchakarska, G.; Sola, B. The double dealing of cyclin D1. Cell Cycle 2020, 19, 163–178. [Google Scholar] [CrossRef] [Green Version]
- Malumbres, M.; Barbacid, M. To cycle or not to cycle: A critical decision in cancer. Nat. Rev. Cancer 2001, 1, 222–231. [Google Scholar] [CrossRef]
- Musgrove, E.A.; Caldon, C.E.; Barraclough, J.; Stone, A.; Sutherland, R.L. Cyclin D as a therapeutic target in cancer. Nat. Rev. Cancer 2011, 11, 558–572. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Research Network. Erratum: Corrigendum: Comprehensive genomic characterization defines human glioblastoma genes and core pathways. Nat. Cell Biol. 2013, 494, 506. [Google Scholar] [CrossRef]
- Cohen, E.E.W.; Soulières, D.; Le Tourneau, C.; Dinis, J.; Licitra, L.; Ahn, M.-J.; Soria, A.; Machiels, J.-P.; Mach, N.; Mehra, R.; et al. Pembrolizumab versus methotrexate, docetaxel, or cetuximab for recurrent or metastatic head-and-neck squamous cell carcinoma (KEYNOTE-040): A randomised, open-label, phase 3 study. Lancet 2019, 393, 156–167. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, Y.; Gao, X.; Li, Y.; Lin, J.; Chen, L.; Chang, L.; Chen, G.; Guan, Y.; Pan, L.K.; et al. CCND1 Amplification Contributes to Immunosuppression and is Associated with a Poor Prognosis to Immune Checkpoint Inhibitors in Solid Tumors. Front. Immunol. 2020, 11, 1620. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, X.; Cheng, Q.; Ning, D.; Ma, J.; Zhang, Z.; Chen, X.; Jiang, L. Effects of cyclin D1 gene silencing on cell proliferation, cell cycle, and apoptosis of hepatocellular carcinoma cells. J. Cell. Biochem. 2018, 119, 2368–2380. [Google Scholar] [CrossRef]
- Montalto, F.I.; De Amicis, F. Cyclin D1 in Cancer: A Molecular Connection for Cell Cycle Control, Adhesion and Invasion in Tumor and Stroma. Cells 2020, 9, 2648. [Google Scholar] [CrossRef] [PubMed]
- Diehl, J.A.; Zindy, F.; Sherr, C.J. Inhibition of cyclin D1 phosphorylation on threonine-286 prevents its rapid degradation via the ubiquitin-proteasome pathway. Genes Dev. 1997, 11, 957–972. [Google Scholar] [CrossRef] [Green Version]
- Malumbres, M.; Barbacid, M. Cell cycle, CDKs and cancer: A changing paradigm. Nat. Rev. Cancer 2009, 9, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.C.; Cardiff, R.D.; Zukerberg, L.; Lees, E.; Arnold, A.; Schmidt, E.V. Mammary hyperplasia and carcinoma in MMTV-cyclin D1 transgenic mice. Nat. Cell Biol. 1994, 369, 669–671. [Google Scholar] [CrossRef]
- Van De Weerdt, B.C.; Littler, D.R.; Klompmaker, R.; Huseinovic, A.; Fish, A.; Perrakis, A.; Medema, R.H. Polo-box domains confer target specificity to the Polo-like kinase family. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2008, 1783, 1015–1022. [Google Scholar] [CrossRef] [Green Version]
- Gutteridge, R.E.A.; Ndiaye, M.A.; Liu, X.; Ahmad, N. Plk1 Inhibitors in Cancer Therapy: From Laboratory to Clinics. Mol. Cancer Ther. 2016, 15, 1427–1435. [Google Scholar] [CrossRef] [Green Version]
- Wolf, G.; Elez, R.; Doermer, A.; Holtrich, U.; Ackermann, H.; Stutte, H.J.; Altmannsberger, H.-M.; Rübsamen-Waigmann, H.; Strebhardt, K. Prognostic significance of polo-like kinase (PLK) expression in non-small cell lung cancer. Oncogene 1997, 14, 543–549. [Google Scholar] [CrossRef]
- Knecht, R.; Elez, R.; Oechler, M.; Solbach, C.; Von Ilberg, C.; Strebhardt, K. Prognostic significance of polo-like kinase (PLK) expression in squamous cell carcinomas of the head and neck. Cancer Res. 1999, 59, 2794–2797. [Google Scholar]
- Montaudon, E.; Nikitorowicz-Buniak, J.; Sourd, L.; Morisset, L.; El Botty, R.; Huguet, L.; Dahmani, A.; Painsec, P.; Nemati, F.; Vacher, S.; et al. PLK1 inhibition exhibits strong anti-tumoral activity in CCND1-driven breast cancer metastases with acquired palbociclib resistance. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Medina-Aguilar, R.; Marchat, L.A.; Ocampo, E.A.; Gariglio, P.; Mena, J.G.; Sepúlveda, N.V.; Castillo, M.M.; López-Camarillo, C. Resveratrol inhibits cell cycle progression by targeting Aurora kinase A and Polo-like kinase 1 in breast cancer cells. Oncol. Rep. 2016, 35, 3696–3704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Xing, S.; Zhang, L.; Yu, J.-M.; Lin, C.; Yang, W.-J. Involvement of Polo-like kinase 1 (Plk1) in quiescence regulation of cancer stem-like cells of the gastric cancer cell lines. Oncotarget 2017, 8, 37633–37645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, H.; Hanison, J.; Nirmalan, N. “Omics”-Informed Drug and Biomarker Discovery: Opportunities, Challenges and Future Perspectives. Proteomes 2016, 4, 28. [Google Scholar] [CrossRef] [Green Version]
- Paananen, J.; Fortino, V. An omics perspective on drug target discovery platforms. Brief. Bioinform. 2020, 21, 1937–1953. [Google Scholar] [CrossRef] [Green Version]
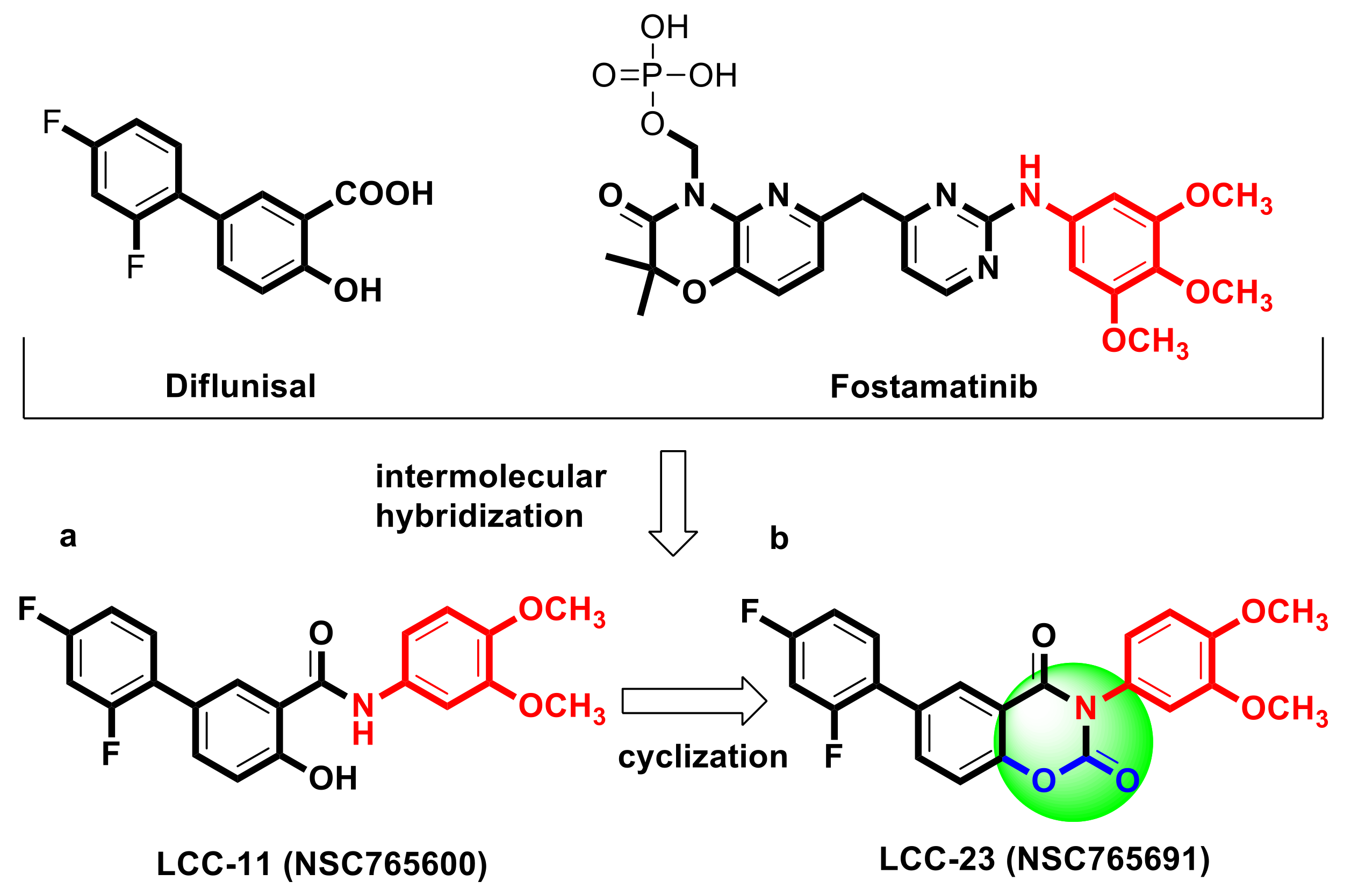
- Zhong, G.-X.; Hu, J.-Q.; Zhao, K.; Chen, L.-L.; Hu, W.-X.; Qiu, M.-Y. Synthesis and biological evaluation of amide derivatives of diflunisal as potential anti-inflammatory agents. Bioorg. Med. Chem. Lett. 2009, 19, 516–519. [Google Scholar] [CrossRef]
- Markham, A. Fostamatinib: First Global Approval. Drugs 2018, 78, 959–963. [Google Scholar] [CrossRef]
- Zhong, G.-X.; Chen, L.-L.; Li, H.-B.; Liu, F.-J.; Hu, J.-Q.; Hu, W.-X. Synthesis and biological evaluation of amide derivatives of diflunisal as potential anti-tumor agents. Bioorg. Med. Chem. Lett. 2009, 19, 4399–4402. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Lipinski, C.A. Lead- and drug-like compounds: The rule-of-five revolution. Drug Discov. Today Technol. 2004, 1, 337–341. [Google Scholar] [CrossRef]
- Martin, Y.C. A Bioavailability Score. J. Med. Chem. 2005, 48, 3164–3170. [Google Scholar] [CrossRef]
- Daina, A.; Zoete, V. A Boiled-Egg to Predict Gastrointestinal Absorption and Brain Penetration of Small Molecules. ChemMedChem 2016, 11, 1117–1121. [Google Scholar] [CrossRef] [Green Version]
- Gfeller, D.; Grosdidier, A.; Wirth, M.; Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: A web server for target prediction of bioactive small molecules. Nucleic Acids Res. 2014, 42, W32–W38. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, R.H. The NCI60 human tumour cell line anticancer drug screen. Nat. Rev. Cancer 2006, 6, 813–823. [Google Scholar] [CrossRef]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Olson, A.J. Using AutoDock for Ligand-Receptor Docking. Curr. Protoc. Bioinform. 2008, 24, 8.14.1–8.14.40. [Google Scholar] [CrossRef]
- Raman, E.P.; Paul, T.J.; Hayes, R.L.; Brooks, I.C.L. Automated, Accurate, and Scalable Relative Protein–Ligand Binding Free-Energy Calculations Using Lambda Dynamics. J. Chem. Theory Comput. 2020, 16, 7895–7914. [Google Scholar] [CrossRef] [PubMed]
- Küçükgüzel, Ş.G.; Küçükgüzel, I.; Tatar, E.; Rollas, S.; Şahin, F.; Güllüce, M.; De Clercq, E.; Kabasakal, L. Synthesis of some novel heterocyclic compounds derived from diflunisal hydrazide as potential anti-infective and anti-inflammatory agents. Eur. J. Med. Chem. 2007, 42, 893–901. [Google Scholar] [CrossRef]
- Liechti, C.; Séquin, U.; Bold, G.; Furet, P.; Meyer, T.; Traxler, P. Salicylanilides as inhibitors of the protein tyrosine kinase epidermal growth factor receptor. Eur. J. Med. Chem. 2004, 39, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, T.J.; Ertl, P.; Lewis, R. The graphical representation of ADME-related molecule properties for medicinal chemists. Drug Discov. Today 2011, 16, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Coussens, N.P.; Braisted, J.C.; Peryea, T.; Sittampalam, G.S.; Simeonov, A.; Hall, M.D. Small-Molecule Screens: A Gateway to Cancer Therapeutic Agents with Case Studies of Food and Drug Administration–Approved Drugs. Pharmacol. Rev. 2017, 69, 479–496. [Google Scholar] [CrossRef] [Green Version]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Nowell, P.C. The clonal evolution of tumor cell populations. Science 1976, 194, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Keller, T.H.; Pichota, A.; Yin, Z. A practical view of ‘druggability’. Curr. Opin. Chem. Biol. 2006, 10, 357–361. [Google Scholar] [CrossRef]
- Morris, G.M.; Lim-Wilby, M. Molecular Docking. Methods Mol. Biol. 2008, 443, 365–382. [Google Scholar] [CrossRef]
- Ma, R.; Ma, Z.-G.; Zhen, C.-L.; Shen, X.; Li, S.-L.; Li, L.; Zheng, Y.-F.; Dong, D.-L.; Sun, Z.-J. Design, synthesis and characterization of poly (methacrylic acid-niclosamide) and its effect on arterial function. Mater. Sci. Eng. C 2017, 77, 352–359. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physicochemical Properties Based on Bioavailability Radar of NSC765600 | Recommended Value | |
|---|---|---|
| Formula | C21H17F2NO4 | |
| Molecular weight | 385.36 g/mol | 150–500 g/mol |
| Fraction Csp3 | 0.10 | ≤1 |
| Number of rotatable bonds | 6 | ≤10 |
| Number of H-bond acceptors | 6 | ≤12 |
| Number of H-bond donors | 2 | ≤5 |
| Molar refractivity | 101.02 | |
| TPSA | 67.79 Å2 | ≤140 Å2 |
| Log Po/w (XLOGP3) | 5 | −0.7–5 |
| Log S (ESOL) | −5.89 | 0–6 |
| Pharmacokinetics | ||
| GI absorption | High | |
| BBB | Low | |
| Drug-likeness | ||
| Lipinski | Yes; 0 violation | |
| Ghose | Yes | |
| Veber | Yes | |
| Egan | Yes | |
| Muegge | Yes | |
| Bioavailability Score | 0.55 (55%) | |
| Medical Chemistry | ||
| Synthetic accessibility | 2.64 | 1 (easy to make) and 10 (difficult to make) |
| Physicochemical Properties Based on Bioavailability Radar of NSC765691 | Recommended Value | |
|---|---|---|
| Formula | C22H15F2NO5 | |
| Molecular weight | 411.36 g/mol | 150–500 g/mol |
| Fraction Csp3 | 0.09 | ≤1 |
| Number of rotatable bonds | 4 | ≤10 |
| Number of H-bond acceptors | 7 | ≤12 |
| Number of H-bond donors | 0 | ≤5 |
| Molar refractivity | 106.42 | |
| TPSA | 70.67 Å2 | ≤140 Å2 |
| Log Po/w (XLOGP3) | 4.53 | −0.7–5 |
| Log S (ESOL) | −5.52 | 0–6 |
| Pharmacokinetics | ||
| GI absorption | High | |
| BBB | Yes (0.215) | |
| Drug-likeness | ||
| Lipinski | Yes; 0 violation | |
| Ghose | Yes | |
| Veber | Yes | |
| Egan | Yes | |
| Muegge | Yes | |
| Bioavailability score | 0.55 (55%) | |
| Medical Chemistry | ||
| Synthetic accessibility | 3.48 | 1 (easy to make) and 10 (difficult to make) |
| Rank | p | CCLC | Target Descriptor | MW | p | CCLC | Target Descriptor | MW | |
|---|---|---|---|---|---|---|---|---|---|
| NSC765600 Fingerprint | 1 | 0.63 | 49 | Dinoterb | 240.21 | 0.46 | 52 | 4-ipomeanol | 168.19 |
| 2 | 0.62 | 50 | 8-[(4-tert-butylphenoxy)] | 342.4 | 0.41 | 49 | Piperazine | 86.14 | |
| 3 | 0.59 | 53 | Masterid | 360.5 | 0.4 | 44 | Amsacrine | 393.5 | |
| 4 | 0.59 | 52 | Nitrodan(usan) | 296.3 | 0.4 | 52 | Fluorodopan | 249.67 | |
| 5 | 0.58 | 41 | Resorcinol, 4-hexyl-(8ci) | 194.27 | 0.39 | 42 | Mustard | 159.08 | |
| 6 | 0.58 | 52 | Azd-4635 | 315.73 | 0.37 | 50 | Tamoxifen | 371.15 | |
| 7 | 0.57 | 50 | Chimaphilin | 186.21 | 0.35 | 52 | Topotecan | 421.4 | |
| 8 | 0.56 | 53 | 10074-g5 | 332.3 | 0.33 | 44 | Morpholino | 86.11 | |
| 9 | 0.56 | 53 | Gsk586581a | 381.4 | 0.32 | 52 | Procarbazine | 221.3 | |
| 10 | 0.55 | 50 | Tioconazole (usan) | 387.7 | 0.28 | 45 | Diaziquone | 364.35 | |
| Rank | r | CCLC | Target Descriptor | MW | r | CCLC | Target Descriptor | MW | |
| NSC765691 Fingerprint | 1 | 0.69 | 41 | Flavanone | 298.3 | 0.5 | 56 | 4-ipomeanol | 168.19 |
| 2 | 0.69 | 54 | 13668 | 217.69 | 0.49 | 53 | Piperazine | 86.14 | |
| 3 | 0.68 | 56 | Isomammein | 372.5 | 0.47 | 55 | Tamoxifen | 371.5 | |
| 4 | 0.66 | 57 | C.I. 37525 | 311.8 | 0.46 | 56 | Flavoneacetic | 280.5 | |
| 5 | 0.65 | 56 | 10074-g5 | 332.3 | 0.44 | 56 | Sulfoximine | 64.09 | |
| 6 | 0.64 | 47 | 4-(acetyl) amphilectolide | 302.4 | 0.43 | 45 | Mustard | 159.08 | |
| 7 | 0.64 | 53 | Chimaphilin | 186.21 | 0.42 | 55 | Bryostatin | 905 | |
| 8 | 0.64 | 57 | Niclosamide (usan) | 327.12 | 0.41 | 56 | Glycoxalic acid | 74.03 | |
| 9 | 0.63 | 46 | Thiazolobenzimidazole | 288.32 | 0.4 | 56 | Fluorodopan | 249.67 | |
| 10 | 0.63 | 56 | Azd-4635 | 315.73 | 0.4 | 56 | Merbarone | 263.27 |
| Target | Common Name | Uniprot ID | ChEMBL ID | Target Class |
|---|---|---|---|---|
| Serine/threonine-protein kinase MTOR | MTOR | P42345 | CHEMBL2842 | Kinase |
| PI3-kinase p85-alpha subunit | PIK3R1 | P27986 | CHEMBL2506 | Enzyme |
| Serotonin 2c (5-HT2c) receptor | HTR2C | P28335 | CHEMBL225 | Family A G protein-coupled receptor |
| Cyclin-dependent kinase 4 | CDK4 | P11802 | CHEMBL331 | Kinase |
| Matrix metalloproteinase 3 | MMP3 | P08254 | CHEMBL283 | Protease |
| Histone chaperone ASF1A | ASF1A | Q9Y294 | CHEMBL3392950 | Unclassified protein |
| PI3-kinase p110-delta subunit | PIK3CD | O00329 | CHEMBL3130 | Enzyme |
| Phosphodiesterase 5A | PDE5A | O76074 | CHEMBL1827 | Phosphodiesterase |
| CDK2/Cyclin A | CCNA2 CDK2 | P20248 P24941 | CHEMBL3038469 | Kinase |
| Nuclear factor NF-kappa-B | NFKB1 | P19838 | CHEMBL3251 | Other cytosolic protein |
| Cyclin-dependent kinase 2/cyclin E | CCNE2CDK2 CCNE1 | O96020 P24941 P24864 | CHEMBL2094126 | Other cytosolic protein |
| Serine/threonine-protein kinase PLK1 | PLK1 | P53350 | CHEMBL3024 | Kinase |
| Beta-glucuronidase | GUSB | P08236 | CHEMBL2728 | Enzyme |
| cAMP-dependent protein kinase alpha-catalytic subunit | PRKACA | P17612 | CHEMBL4101 | Kinase |
| Interleukin-8 receptor B | CXCR2 | P25025 | CHEMBL2434 | Family A G protein-coupled receptor |
| Dual specificity tyrosine-phosphorylation-regulated kinase 1B | DYRK1B | Q9Y463 | CHEMBL5543 | Kinase |
| Cyclin-dependent kinase 4/cyclin D1 | CCND1 CDK4 | P24385 P11802 | CHEMBL1907601 | Kinase |
| Cyclophilin A | PPIA | P62937 | CHEMBL1949 | Isomerase |
| Platelet-derived growth factor receptor alpha | PDGFRA | P16234 | CHEMBL2007 | Kinase |
| Glycogen synthase kinase-3 alpha | GSK3A | P49840 | CHEMBL2850 | Kinase |
| Target | Common Name | Uniprot ID | ChEMBL ID | Target Class |
|---|---|---|---|---|
| Serine/threonine-protein kinase | PLK1 | P53350 | CHEMBL3024 | Kinase |
| Nicotinamide phosphoribosyl transferase | NAMPT | P43490 | CHEMBL1744525 | Enzyme |
| Rho-associated protein kinase 1 | ROCK1 | Q13464 | CHEMBL3231 | Kinase |
| Monoamine oxidase B | MAOB | P27338 | CHEMBL2039 | Oxidoreductase |
| Focal adhesion kinase 1 | PTK2 | Q05397 | CHEMBL2695 | Kinase |
| Vascular endothelial growth factor receptor 2 | KDR | P35968 | CHEMBL279 | Kinase |
| Tyrosine-protein kinase TIE-2 | TEK | Q02763 | CHEMBL4128 | Kinase |
| Cyclin-dependent kinase 5/CDK5 activator 1 | CDK5R1 CDK5 | Q15078 Q00535 | CHEMBL1907600 | Kinase |
| Cyclin-dependent kinase 7 | CDK7 | P50613 | CHEMBL3055 | Kinase |
| Platelet-derived growth factor receptor alpha | PDGFRA | P16234 | CHEMBL2007 | Kinase |
| TGF-beta receptor type I | TGFBR1 | P36897 | CHEMBL4439 | Kinase |
| Phosphodiesterase 5A | PDE5A | O76074 | CHEMBL1827 | Phosphodiesterase |
| Cyclin-dependent kinase 4/cyclin D1 | CCND1 CDK4 | P24385 P11802 | CHEMBL1907601 | Kinase |
| Rho-associated protein kinase 2 | ROCK2 | O75116 | CHEMBL2973 | Kinase |
| Cyclin-dependent kinase 2 | CDK2 | P24941 | CHEMBL301 | Kinase |
| Cyclin-dependent kinase 1 | CDK1 | P06493 | CHEMBL308 | Kinase |
| Cyclin-dependent kinase 4 | CDK4 | P11802 | CHEMBL331 | Kinase |
| G-protein coupled receptor kinase 2 | GRK2 | P25098 | CHEMBL4079 | Kinase |
| Toll-like receptor (TLR7/TLR9) | TLR9 | Q9NR96 | CHEMBL5804 | Toll-like and Il-1 receptors |
| Glycogen synthase kinase-3 beta | GSK3B | P49841 | CHEMBL262 | Kinase |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mokgautsi, N.; Wang, Y.-C.; Lawal, B.; Khedkar, H.; Sumitra, M.R.; Wu, A.T.H.; Huang, H.-S. Network Pharmacological Analysis through a Bioinformatics Approach of Novel NSC765600 and NSC765691 Compounds as Potential Inhibitors of CCND1/CDK4/PLK1/CD44 in Cancer Types. Cancers 2021, 13, 2523. https://doi.org/10.3390/cancers13112523
Mokgautsi N, Wang Y-C, Lawal B, Khedkar H, Sumitra MR, Wu ATH, Huang H-S. Network Pharmacological Analysis through a Bioinformatics Approach of Novel NSC765600 and NSC765691 Compounds as Potential Inhibitors of CCND1/CDK4/PLK1/CD44 in Cancer Types. Cancers. 2021; 13(11):2523. https://doi.org/10.3390/cancers13112523
Chicago/Turabian StyleMokgautsi, Ntlotlang, Yu-Chi Wang, Bashir Lawal, Harshita Khedkar, Maryam Rachmawati Sumitra, Alexander T. H. Wu, and Hsu-Shan Huang. 2021. "Network Pharmacological Analysis through a Bioinformatics Approach of Novel NSC765600 and NSC765691 Compounds as Potential Inhibitors of CCND1/CDK4/PLK1/CD44 in Cancer Types" Cancers 13, no. 11: 2523. https://doi.org/10.3390/cancers13112523