
Estrogens and Progestogens in Triple Negative Breast Cancer: Do They Harm?

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
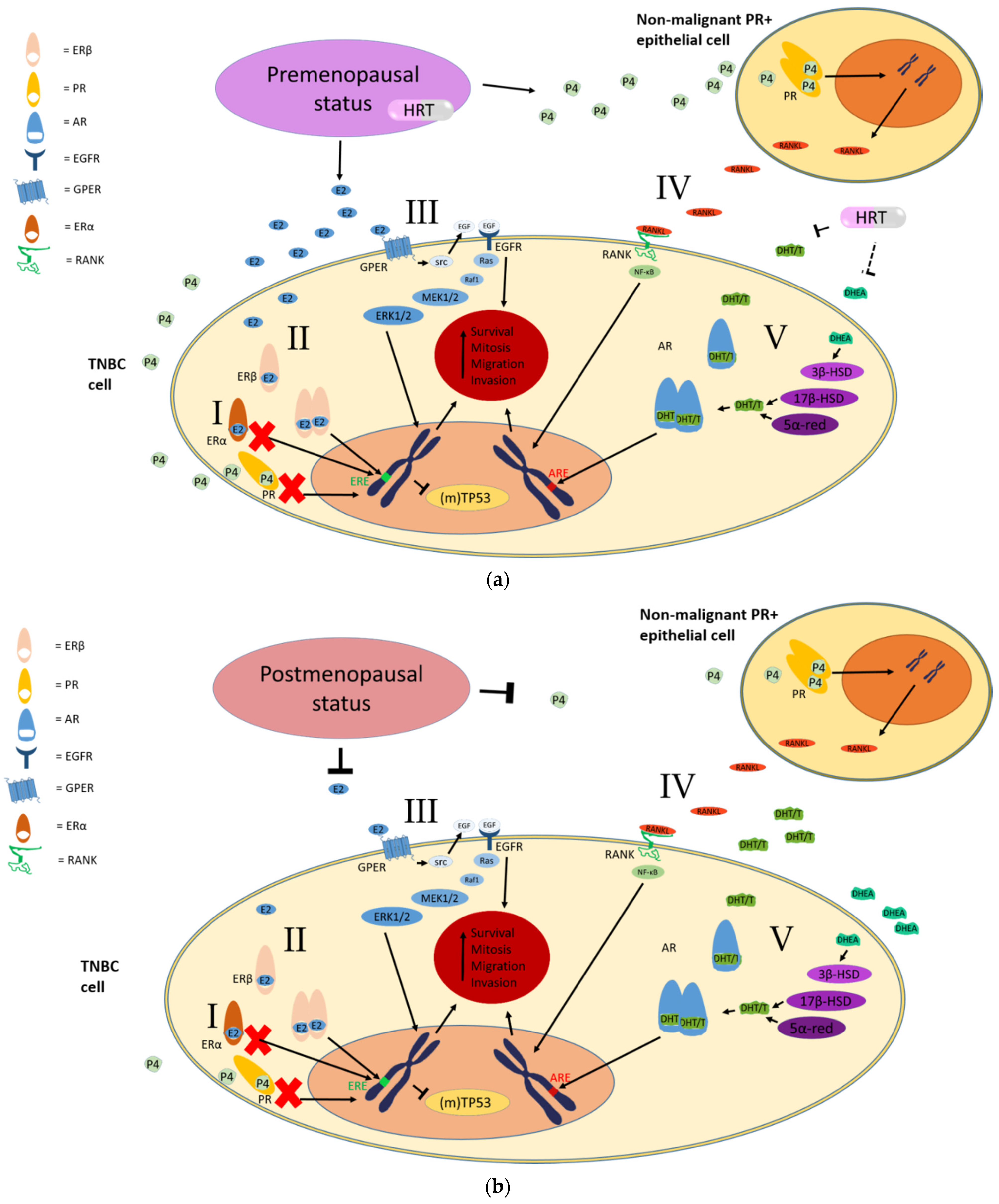
2. Clinical Evidence for a Role of Estrogen and Progesterone in TNBC
2.1. The Level of ER Expression in Primary TNBC
2.2. ER Conversion of Primary TNBC
2.3. Effect of Recovery of Ovarian Function (Endogenous Hormone Exposure) after Chemotherapy
2.4. Effect of Exogenous Hormone Exposure
3. Preclinical Evidence for a Role of Estrogen and Progesterone in TNBC
3.1. Estrogen Receptor Beta in TNBC
3.2. The G Protein-Coupled Estrogen Receptor (GPER1/GPR30)
3.3. The Effect of the RANK/RANKL Pathway
3.4. Androgens and the Androgen Receptor in TNBC
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bauer, K.R.; Brown, M.; Cress, R.D.; Parise, C.A.; Caggiano, V. Descriptive analysis of estrogen receptor (ER)-negative, progesterone receptor (PR)-negative, and HER2-negative invasive breast cancer, the so-called triple-negative phenotype. Cancer 2007, 109, 1721–1728. [Google Scholar] [CrossRef]
- Early Breast Cancer Trialists’ Collaborative Group (EBCTCG). Effects of chemotherapy and hormonal therapy for early breast cancer on recurrence and 15-year survival: An overview of the randomised trials. Lancet 2005, 365, 1687–1717. [Google Scholar] [CrossRef]
- Carter, J.; Lacchetti, C.; Andersen, B.L.; Barton, D.L.; Bolte, S.; Damast, S.; Diefenbach, M.A.; Duhamel, K.; Florendo, J.; Ganz, P.A.; et al. Interventions to Address Sexual Problems in People with Cancer: American Society of Clinical Oncology Clinical Practice Guideline Adaptation of Cancer Care Ontario Guideline. J. Clin. Oncol. 2018, 36, 492–511. [Google Scholar] [CrossRef] [PubMed]
- Kenemans, P.; Bundred, N.J.; Foidart, J.-M.; Kubista, E.; von Schoultz, B.; Sismondi, P.; Vassilopoulou-Sellin, R.; Yip, C.H.; Egberts, J.; Mol-Arts, M.; et al. Safety and efficacy of tibolone in breast-cancer patients with vasomotor symptoms: A double-blind, randomised, non-inferiority trial. Lancet Oncol. 2009, 10, 135–146. [Google Scholar] [CrossRef]
- Holmberg, L.; Anderson, H. HABITS (hormonal replacement therapy after breast cancer—Is it safe?), a randomised comparison: Trial stopped. Lancet 2004, 363, 453–455. [Google Scholar] [CrossRef]
- Writing Group for the Women’s Health Initiative Investigators. Risks and Benefits of Estrogen Plus Progestin in Healthy Postmenopausal Women: Principal Results from the Women’s Health Initiative Randomized Controlled Trial. JAMA 2002, 288, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Liu, J.; Chen, K.; Li, S.; Wang, Y.; Yang, Y.; Deng, H.; Jia, W.; Rao, N.; Liu, Q.; et al. What lies behind chemotherapy-induced amenorrhea for breast cancer patients: A meta-analysis. Breast Cancer Res. Treat. 2014, 145, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Muka, T.; Oliver-Williams, C.; Kunutsor, S.; Laven, J.S.E.; Fauser, B.C.J.M.; Chowdhury, R.; Kavousi, M.; Franco, O.H. Association of Age at Onset of Menopause and Time Since Onset of Menopause with Cardiovascular Outcomes, Intermediate Vascular Traits, and All-Cause Mortality. JAMA Cardiol. 2016, 1, 767–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, T.; Kogawa, T.; Dong, W.; Sahin, A.A.; Moulder, S.; Litton, J.K.; Tripathy, D.; Iwamoto, T.; Hunt, K.K.; Pusztai, L.; et al. Revisiting the definition of estrogen receptor positivity in HER2-negative primary breast cancer. Ann. Oncol. 2017, 28, 2420–2428. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Huo, L.; Koenig, K.B.; Mittendorf, E.A.; Meric-Bernstam, F.; Kuerer, H.M.; Bedrosian, I.; Buzdar, A.U.; Symmans, W.F.; Crow, J.R.; et al. Which threshold for ER positivity? A retrospective study based on 9639 patients. Ann. Oncol. 2014, 25, 1004–1011. [Google Scholar] [CrossRef]
- Cheang, M.C.; Martin, M.; Nielsen, T.O.; Prat, A.; Voduc, D.; Rodriguez-Lescure, A.; Ruiz, A.; Chia, S.; Shepherd, L.; Ruiz-Borrego, M.; et al. Defining Breast Cancer Intrinsic Subtypes by Quantitative Receptor Expression. Oncologist 2015, 20, 474–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrijver, W.; Suijkerbuijk, K.P.M.; van Gils, C.H.; van der Wall, E.; Moelans, C.B.; van Diest, P.J. Receptor Conversion in Distant Breast Cancer Metastases: A Systematic Review and Meta-analysis. J. Natl. Cancer Inst. 2018, 110, 568–580. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Yin, W.; Du, Y.; Shen, Z.; Lu, J. Prognostic impact of chemotherapy-induced amenorrhea on premenopausal breast cancer: A meta-analysis of the literature. Menopause 2015, 22, 1091–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambertini, M.; Campbell, C.; Bines, J.; Korde, L.A.; Izquierdo, M.; Fumagalli, D.; Del Mastro, L.; Ignatiadis, M.; Pritchard, K.; Wolff, A.C.; et al. Adjuvant Anti-HER2 Therapy, Treatment-Related Amenorrhea, and Survival in Premenopausal HER2-Positive Early Breast Cancer Patients. J. Natl. Cancer Inst. 2019, 111, 86–94. [Google Scholar] [CrossRef]
- Van Barele, M.; Heemskerk-Gerritsen, B.A.; Van Doorn, H.C.; Schmidt, M.K.; Hooning, M.J.; Jager, A. The impact of menstruation persistence or recovery after chemotherapy on survival in young patients with hormone receptor negative breast cancer. Breast 2020, 52, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Park, I.H.; Han, H.S.; Lee, H.; Lee, K.S.; Kang, H.S.; Lee, S.; Kim, S.W.; Jung, S.; Ro, J. Resumption or persistence of menstruation after cytotoxic chemotherapy is a prognostic factor for poor disease-free survival in premenopausal patients with early breast cancer. Ann. Oncol. 2012, 23, 2283–2289. [Google Scholar] [CrossRef]
- Metcalfe, K.; Lynch, H.T.; Foulkes, W.D.; Tung, N.; Kim-Sing, C.; Olopade, O.I.; Eisen, A.; Rosen, B.; Snyder, C.; Gershman, S.; et al. Effect of Oophorectomy on Survival After Breast Cancer inBRCA1andBRCA2Mutation Carriers. JAMA Oncol. 2015, 1, 306–313. [Google Scholar] [CrossRef]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-Negative Breast Cancer: Clinical Features and Patterns of Recurrence. Clin. Cancer Res. 2007, 13, 4429–4434. [Google Scholar] [CrossRef] [Green Version]
- Early Breast Cancer Trialists’ Collaborative Group; Clarke, M.J. Ovarian ablation for early breast cancer. Cochrane Database Syst. Rev. 1998, 2000, CD000485. [Google Scholar] [CrossRef]
- Liedtke, C.; Rody, A.; Gluz, O.; Baumann, K.; Beyer, D.; Kohls, E.-B.; Lausen, K.; Hanker, L.; Holtrich, U.; Becker, S.; et al. The prognostic impact of age in different molecular subtypes of breast cancer. Breast Cancer Res. Treat. 2015, 152, 667–673. [Google Scholar] [CrossRef]
- Liedtke, C.; Hess, K.R.; Karn, T.; Rody, A.; Kiesel, L.; Hortobagyi, G.N.; Pusztai, L.; Gonzalez-Angulo, A.M. The prognostic impact of age in patients with triple-negative breast cancer. Breast Cancer Res. Treat. 2013, 138, 591–599. [Google Scholar] [CrossRef]
- Partridge, A.H.; Hughes, M.E.; Warner, E.T.; Ottesen, R.A.; Wong, Y.-N.; Edge, S.B.; Theriault, R.L.; Blayney, D.W.; Niland, J.C.; Winer, E.P.; et al. Subtype-Dependent Relationship Between Young Age at Diagnosis and Breast Cancer Survival. J. Clin. Oncol. 2016, 34, 3308–3314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, C.K.; Hsu, D.S.; Broadwater, G.; Acharya, C.R.; Foekens, J.A.; Zhang, Y.; Wang, Y.; Marcom, P.K.; Marks, J.R.; Febbo, P.G.; et al. Young Age at Diagnosis Correlates with Worse Prognosis and Defines a Subset of Breast Cancers with Shared Patterns of Gene Expression. J. Clin. Oncol. 2008, 26, 3324–3330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, H.C.F.; Unger, J.M.; Phillips, K.-A.; Boyle, F.; Hitre, E.; Porter, D.; Francis, P.A.; Goldstein, L.J.; Gomez, H.L.; Vallejos, C.S.; et al. Goserelin for Ovarian Protection during Breast-Cancer Adjuvant Chemotherapy. N. Engl. J. Med. 2015, 372, 923–932. [Google Scholar] [CrossRef] [Green Version]
- Harvey, J.M.; Clark, G.M.; Osborne, C.K.; Allred, D.C. Estrogen Receptor Status by Immunohistochemistry Is Superior to the Ligand-Binding Assay for Predicting Response to Adjuvant Endocrine Therapy in Breast Cancer. J. Clin. Oncol. 1999, 17, 1474. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.; Harbeck, N.; Nap, M.; Molina, R.; Nicolini, A.; Senkus, E.; Cardoso, F. Clinical use of biomarkers in breast cancer: Updated guidelines from the European Group on Tumor Markers (EGTM). Eur. J. Cancer 2017, 75, 284–298. [Google Scholar] [CrossRef] [Green Version]
- Marotti, J.D.; Collins, L.C.; Hu, R.; Tamimi, R.M. Estrogen receptor-β expression in invasive breast cancer in relation to molecular phenotype: Results from the Nurses’ Health Study. Mod. Pathol. 2009, 23, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Dahlman-Wright, K.; Gustafsson, J.A. Estrogen receptor beta: An overview and update. Nuclear Recept. Signal. 2008, 6, e003. [Google Scholar] [CrossRef] [Green Version]
- Skliris, G.P.; Leygue, E.; Watson, P.H.; Murphy, L.C. Estrogen receptor alpha negative breast cancer patients: Estrogen receptor beta as a therapeutic target. J. Steroid Biochem. Mol. Biol. 2008, 109, 1–10. [Google Scholar] [CrossRef]
- Litwiniuk, M.M.; Rożnowski, K.; Filas, V.; Godlewski, D.D.; Stawicka, M.; Kaleta, R.; Bręborowicz, J. Expression of estrogen receptor beta in the breast carcinoma of BRCA1 mutation carriers. BMC Cancer 2008, 8, 100. [Google Scholar] [CrossRef] [Green Version]
- Andersson, S.; Sundberg, M.; Pristovsek, N.; Ibrahim, A.; Jonsson, P.; Katona, B.; Clausson, C.-M.; Zieba, A.; Ramström, M.; Söderberg, O.; et al. Insufficient antibody validation challenges oestrogen receptor beta research. Nat. Commun. 2017, 8, 15840. [Google Scholar] [CrossRef] [PubMed]
- Reese, J.M.; Suman, V.J.; Subramaniam, M.; Wu, X.; Negron, V.; Gingery, A.; Pitel, K.S.; Shah, S.S.; Cunliffe, H.E.; McCullough, A.E.; et al. ERβ1: Characterization, prognosis, and evaluation of treatment strategies in ERα-positive and -negative breast cancer. BMC Cancer 2014, 14, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawse, J.R.; Carter, J.M.; Aspros, K.G.M.; Bruinsma, E.S.; Koepplin, J.W.; Negron, V.; Subramaniam, M.; Ingle, J.N.; Rech, K.L.; Goetz, M.P. Optimized immunohistochemical detection of estrogen receptor beta using two validated monoclonal antibodies confirms its expression in normal and malignant breast tissues. Breast Cancer Res. Treat. 2020, 179, 241–249. [Google Scholar] [CrossRef]
- Leung, Y.-K.; Mak, P.; Hassan, S.; Ho, S.-M. Estrogen receptor (ER)-beta isoforms: A key to understanding ER-beta signaling. Proc. Natl. Acad. Sci. 2006, 103, 13162–13167. [Google Scholar] [CrossRef] [Green Version]
- Chantzi, Ν.Ι.; Tiniakos, D.G.; Palaiologou, M.; Goutas, N.; Filippidis, T.; Vassilaros, S.D.; Dhimolea, E.; Mitsiou, D.J.; Alexis, Μ.N. Estrogen receptor beta 2 is associated with poor prognosis in estrogen receptor alpha-negative breast carcinoma. J. Cancer Res. Clin. Oncol. 2013, 139, 1489–1498. [Google Scholar] [CrossRef] [PubMed]
- Groenendijk, F.H.; Zwart, W.; Floore, A.; Akbari, S.; Bernards, R. Estrogen receptor splice variants as a potential source of false-positive estrogen receptor status in breast cancer diagnostics. Breast Cancer Res. Treat. 2013, 140, 475–484. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.; Rajapaksa, G.; Nikolos, F.; Hao, R.; Katchy, A.; Mccollum, C.W.; Bondesson, M.; Quinlan, P.; Thompson, A.; Krishnamurthy, S.; et al. ERβ1 represses basal-like breast cancer epithelial to mesenchymal transition by destabilizing EGFR. Breast Cancer Res. 2012, 14, R148. [Google Scholar] [CrossRef] [Green Version]
- Hinsche, O.; Girgert, R.; Emons, G.; Gründker, C. Estrogen receptor? selective agonists reduce invasiveness of triple-negative breast cancer cells. Int. J. Oncol. 2014, 46, 878–884. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Huang, S.; Mei, S.; Yang, Z.; Xu, L.; Zhou, N.; Yang, Q.; Shen, Q.; Wang, W.; Le, X.; et al. Pharmacological activation of estrogen receptor beta augments innate immunity to suppress cancer metastasis. Proc. Natl. Acad. Sci. USA 2018, 115, E3673–E3681. [Google Scholar] [CrossRef] [Green Version]
- Shanle, E.K.; Zhao, Z.; Hawse, J.; Wisinski, K.; Keles, S.; Yuan, M.; Xu, W. Research Resource: Global Identification of Estrogen Receptor β Target Genes in Triple Negative Breast Cancer Cells. Mol. Endocrinol. 2013, 27, 1762–1775. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.K.; Abrahamsson, A.; Dabrosin, C. Fulvestrant inhibits growth of triple negative breast cancer and synergizes with tamoxifen in ERalpha positive breast cancer by up-regulation of ERbeta. Oncotarget 2016, 7, 56876–56888. [Google Scholar] [CrossRef] [PubMed]
- Bado, I.; Nikolos, F.; Rajapaksa, G.; Gustafsson, J.A.; Thomas, C. ERbeta decreases the invasiveness of triple-negative breast cancer cells by regulating mutant p53 oncogenic function. Oncotarget 2016, 7, 13599–13611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reese, J.M.; Bruinsma, E.S.; Monroe, D.G.; Negron, V.; Suman, V.J.; Ingle, J.N. ERbeta inhibits cyclin dependent kinases 1 and 7 in triple negative breast cancer. Oncotarget 2017, 8, 96506–96521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schüler-Toprak, S.; Häring, J.; Inwald, E.C.; Moehle, C.; Ortmann, O.; Treeck, O. Agonists and knockdown of estrogen receptor β differentially affect invasion of triple-negative breast cancer cells in vitro. BMC Cancer 2016, 16, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reese, J.M.; Bruinsma, E.S.; Nelson, A.W.; Chernukhin, I.; Carroll, J.S.; Li, Y.; Subramaniam, M.; Suman, V.J.; Negron, V.; Monroe, D.G.; et al. ERβ-mediated induction of cystatins results in suppression of TGFβ signaling and inhibition of triple-negative breast cancer metastasis. Proc. Natl. Acad. Sci. USA 2018, 115, E9580–E9589. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, U.K.; Oturkar, C.C.; Adams, C.; Wickramasekera, N.; Bansal, S.; Medisetty, R.; Miller, A.; Swetzig, W.M.; Silwal-Pandit, L.; Børresen-Dale, A.-L.; et al. TP53 Status as a Determinant of Pro- vs Anti-Tumorigenic Effects of Estrogen Receptor-Beta in Breast Cancer. J. Natl. Cancer Inst. 2019, 111, 1202–1215. [Google Scholar] [CrossRef] [PubMed]
- Koboldt, D.C.; Fulton, R.S.; McLellan, M.D.; Schmidt, H.; Kalicki-Veizer, J.; McMichael, J.F.; Fulton, L.L.; Dooling, D.J.; Ding, L.; Mardis, E.R.; et al. Comprehensive molecular portraits of human breast tumours. Nature 2012, 490, 61–70. [Google Scholar]
- Hui, L.; Zheng, Y.; Yan, Y.; Bargonetti, J.; Foster, D.A. Mutant p53 in MDA-MB-231 breast cancer cells is stabilized by elevated phospholipase D activity and contributes to survival signals generated by phospholipase D. Oncogene 2006, 25, 7305–7310. [Google Scholar] [CrossRef] [Green Version]
- Oliveras-Ferraros, C.; Vazquez-Martin, A.; López-Bonet, E.; Martín-Castillo, B.; Del Barco, S.; Brunet, J.; Menendez, J.A. Growth and molecular interactions of the anti-EGFR antibody cetuximab and the DNA cross-linking agent cisplatin in gefitinib-resistant MDA-MB-468 cells: New prospects in the treatment of triple-negative/basal-like breast cancer. Int. J. Oncol. 2008, 33, 1165–1176. [Google Scholar] [PubMed]
- Austin, D.; Hamilton, N.; Elshimali, Y.; Pietras, R.; Wu, Y.; Vadgama, J. Estrogen receptor-beta is a potential target for triple negative breast cancer treatment. Oncotarget 2018, 9, 33912–33930. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, N.; Márquez-Garbán, D.; Mah, V.; Fernando, G.; Elshimali, Y.; Garban, H.; Elashoff, D.; Vadgama, J.; Goodglick, L.; Pietras, R. Biologic Roles of Estrogen Receptor-β and Insulin-Like Growth Factor-2 in Triple-Negative Breast Cancer. BioMed Res. Int. 2015, 2015, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, C.; Lopes-Coelho, F.; Ramos, C.; Martins, F.; Santos, I.; Rodrigues, A.; Silva, F.; André, S.; Serpa, J. Unraveling FATP1, regulated by ER-β, as a targeted breast cancer innovative therapy. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Piperigkou, Z.; Bouris, P.; Onisto, M.; Franchi, M.; Kletsas, D.; Theocharis, A.D.; Karamanos, N.K. Estrogen receptor beta modulates breast cancer cells functional properties, signaling and expression of matrix molecules. Matrix Biol. 2016, 56, 4–23. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Dey, P.; Ziegler, Y.; Jiao, X.; Kim, S.H.; Katzenellenbogen, J.A.; Katzenellenbogen, B.S. Contrasting activities of estrogen receptor beta isoforms in triple negative breast cancer. Breast Cancer Res. Treat. 2021, 185, 281–292. [Google Scholar] [CrossRef]
- Nelson, A.W.; Groen, A.J.; Miller, J.L.; Warren, A.Y.; Holmes, K.A.; Tarulli, G.A.; Tilley, W.D.; Katzenellenbogen, B.S.; Hawse, J.R.; Gnanapragasam, V.J.; et al. Comprehensive assessment of estrogen receptor beta antibodies in cancer cell line models and tissue reveals critical limitations in reagent specificity. Mol. Cell. Endocrinol. 2017, 440, 138–150. [Google Scholar] [CrossRef] [Green Version]
- Novelli, F.; Milella, M.; Melucci, E.; Di Benedetto, A.; Sperduti, I.; Perrone-Donnorso, R.; Perracchio, L.; Venturo, I.; Nisticò, C.; Fabi, A.; et al. A divergent role for estrogen receptor-beta in node-positive and node-negative breast cancer classified according to molecular subtypes: An observational prospective study. Breast Cancer Res. 2008, 10, R74. [Google Scholar] [CrossRef] [Green Version]
- Wimberly, H.; Han, G.; Pinnaduwage, D.; Murphy, L.C.; Yang, X.R.; Andrulis, I.L.; Sherman, M.; Figueroa, J.; Rimm, D.L. ERβ splice variant expression in four large cohorts of human breast cancer patient tumors. Breast Cancer Res. Treat. 2014, 146, 657–667. [Google Scholar] [CrossRef]
- Shanle, E.K.; Onitilo, A.A.; Huang, W.; Kim, K.; Zang, C.; Engel, J.M.; Xu, W.; Wisinski, K.B. Prognostic significance of full-length estrogen receptor beta expression in stage I-III triple negative breast cancer. Am. J. Transl. Res. 2015, 7, 1246–1259. [Google Scholar]
- Wang, J.; Zhang, C.; Chen, K.; Tang, H.; Tang, J.; Song, C.; Xie, X. ERβ1 inversely correlates with PTEN/PI3K/AKT pathway and predicts a favorable prognosis in triple-negative breast cancer. Breast Cancer Res. Treat. 2015, 152, 255–269. [Google Scholar] [CrossRef]
- Yu, K.-D.; Zhu, R.; Zhan, M.; Rodriguez, A.A.; Yang, W.; Wong, S.; Makris, A.; Lehmann, B.D.; Chen, X.; Mayer, I.; et al. Identification of Prognosis-Relevant Subgroups in Patients with Chemoresistant Triple-Negative Breast Cancer. Clin. Cancer Res. 2013, 19, 2723–2733. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Li, X.; Blanchard, A.; Bramwell, V.H.C.; Pritchard, K.I.; Tu, D.; Shepherd, L.; Myal, Y.; Penner, C.; Watson, P.H.; et al. Expression of both Estrogen Receptor-beta 1 (ER-β1) and its co-regulator Steroid Receptor RNA Activator Protein (SRAP) are predictive for benefit from tamoxifen therapy in patients with Estrogen Receptor-alpha (ER-α)-Negative Early Breast Cancer (EBC). Ann. Oncol. 2013, 24, 1986–1993. [Google Scholar] [CrossRef] [PubMed]
- Gruvberger-Saal, S.K.; Bendahl, P.-O.; Saal, L.H.; Laakso, M.; Hegardt, C.; Edén, P.; Peterson, C.; Malmström, P.; Isola, J.; Borg, Å.; et al. Estrogen Receptor β Expression Is Associated with Tamoxifen Response in ERα-Negative Breast Carcinoma. Clin. Cancer Res. 2007, 13, 1987–1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakopoulou, L.; Lazaris, A.C.; Panayotopoulou, E.G.; Giannopoulou, I.; Givalos, N.; Markaki, S.; Keramopoulos, A. The favourable prognostic value of oestrogen receptor immunohistochemical expression in breast cancer. J. Clin. Pathol. 2004, 57, 523–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honma, N.; Horii, R.; Iwase, T.; Saji, S.; Younes, M.; Takubo, K.; Matsuura, M.; Ito, Y.; Akiyama, F.; Sakamoto, G. Clinical Importance of Estrogen Receptor-β Evaluation in Breast Cancer Patients Treated with Adjuvant Tamoxifen Therapy. J. Clin. Oncol. 2008, 26, 3727–3734. [Google Scholar] [CrossRef] [PubMed]
- Wisinski, K.B.; Xu, W.; Tevaarwerk, A.J.; Saha, S.; Kim, K.; Traynor, A.; Dietrich, L.; Hegeman, R.; Patel, D.; Blank, J.; et al. Targeting Estrogen Receptor Beta in a Phase 2 Study of High-Dose Estradiol in Metastatic Triple-Negative Breast Cancer: A Wisconsin Oncology Network Study. Clin. Breast Cancer 2016, 16, 256–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watters, J.J.; Campbell, J.S.; Cunningham, M.J.; Krebs, E.G.; Dorsa, D.M. Rapid Membrane Effects of Steroids in Neuroblastoma Cells: Effects of Estrogen on Mitogen Activated Protein Kinase Signalling Cascade and c-fos Immediate Early Gene Transcription. Endocrinology 1997, 138, 4030–4033. [Google Scholar] [CrossRef] [PubMed]
- Revankar, C.M.; Cimino, D.F.; Sklar, L.A.; Arterburn, J.B.; Prossnitz, E.R. A Transmembrane Intracellular Estrogen Receptor Mediates Rapid Cell Signaling. Science 2005, 307, 1625–1630. [Google Scholar] [CrossRef] [Green Version]
- Filardo, E.J.; Quinn, J.A.; Bland, K.I.; Frackelton, A.R. Estrogen-Induced Activation of Erk-1 and Erk-2 Requires the G Protein-Coupled Receptor Homolog, GPR30, and Occurs via Trans-Activation of the Epidermal Growth Factor Receptor through Release of HB-EGF. Mol. Endocrinol. 2000, 14, 1649–1660. [Google Scholar] [CrossRef]
- Prossnitz, E.R.; Barton, M. Signaling, physiological functions and clinical relevance of the G protein-coupled estrogen receptor GPER. Prostaglandins Other Lipid Mediat. 2009, 89, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Filardo, E.J. Epidermal growth factor receptor (EGFR) transactivation by estrogen via the G-protein-coupled receptor, GPR30: A novel signaling pathway with potential significance for breast cancer. J. Steroid Biochem. Mol. Biol. 2002, 80, 231–238. [Google Scholar] [CrossRef]
- Lappano, R.; Rigiracciolo, D.; De Marco, P.; Avino, S.; Cappello, A.R.; Rosano, C.; Maggiolini, M.; De Francesco, E.M. Recent Advances on the Role of G Protein-Coupled Receptors in Hypoxia-Mediated Signaling. AAPS J. 2016, 18, 305–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, D.P.; Lappano, R.; Albanito, L.; Madeo, A.; Maggiolini, M.; Picard, D. Estrogenic GPR30 signalling induces proliferation and migration of breast cancer cells through CTGF. EMBO J. 2009, 28, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marjon, N.A.; Hu, C.; Hathaway, H.J.; Prossnitz, E.R. G Protein–Coupled Estrogen Receptor Regulates Mammary Tumorigenesis and Metastasis. Mol. Cancer Res. 2014, 12, 1644–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.; Liu, M.; Luo, H.; Wu, C.; Tang, X.; Tang, S.; Hu, P.; Yan, Y.; Wang, Z.; Tu, G. GPER mediates enhanced cell viability and motility via non-genomic signaling induced by 17β-estradiol in triple-negative breast cancer cells. J. Steroid Biochem. Mol. Biol. 2014, 143, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Girgert, R.; Emons, G.; Gründker, C. 17β-estradiol-induced growth of triple-negative breast cancer cells is prevented by the reduction of GPER expression after treatment with gefitinib. Oncol. Rep. 2016, 37, 1212–1218. [Google Scholar] [CrossRef] [PubMed]
- Girgert, R.; Emons, G.; Gründker, C. Inactivation of GPR30 reduces growth of triple-negative breast cancer cells: Possible application in targeted therapy. Breast Cancer Res. Treat. 2012, 134, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.-J.; Wei, W.-D.; Jiang, G.-M.; Liu, H.; Yang, X.; Wu, Y.-M.; Liu, H.; Wong, C.K.; Du, J.; Wang, H.-S. Activation of GPER suppresses epithelial mesenchymal transition of triple negative breast cancer cells via NF-κB signals. Mol. Oncol. 2016, 10, 775–788. [Google Scholar] [CrossRef] [Green Version]
- Samartzis, E.P.; Noske, A.; Meisel, A.; Varga, Z.; Fink, D.; Imesch, P. The G Protein-Coupled Estrogen Receptor (GPER) Is Expressed in Two Different Subcellular Localizations Reflecting Distinct Tumor Properties in Breast Cancer. PLoS ONE 2014, 9, e83296. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Sun, P.; Zhang, Y.; You, X.; Li, P.; Wang, T. Estrogen stimulated migration and invasion of estrogen receptor-negative breast cancer cells involves an ezrin-dependent crosstalk between G protein-coupled receptor 30 and estrogen receptor beta signaling. Steroids 2016, 111, 113–120. [Google Scholar] [CrossRef]
- Talia, M.; De Francesco, E.M.; Rigiracciolo, D.C.; Muoio, M.G.; Muglia, L.; Belfiore, A.; Maggiolini, M.; Sims, A.H.; Lappano, R. The G Protein-Coupled Estrogen Receptor (GPER) Expression Correlates with Pro-Metastatic Pathways in ER-Negative Breast Cancer: A Bioinformatics Analysis. Cells 2020, 9, 622. [Google Scholar] [CrossRef] [Green Version]
- Fata, J.E.; Kong, Y.-Y.; Li, J.; Sasaki, T.; Irie-Sasaki, J.; Moorehead, R.A.; Elliott, R.; Scully, S.; Voura, E.B.; Lacey, D.L.; et al. The Osteoclast Differentiation Factor Osteoprotegerin-Ligand Is Essential for Mammary Gland Development. Cell 2000, 103, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Beleut, M.; Rajaram, R.D.; Caikovski, M.; Ayyanan, A.; Germano, D.; Choi, Y.; Schneider, P.; Brisken, C. Two distinct mechanisms underlie progesterone-induced proliferation in the mammary gland. Proc. Natl. Acad. Sci. USA 2010, 107, 2989–2994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sflomos, G.; Brisken, C. A new Achilles Heel in breast cancer? Oncotarget 2013, 4, 1126–1127. [Google Scholar] [CrossRef] [Green Version]
- Kiesel, L.; Kohl, A. Role of the RANK/RANKL pathway in breast cancer. Maturitas 2016, 86, 10–16. [Google Scholar] [CrossRef]
- Schramek, D.; Leibbrandt, A.; Sigl, V.; Kenner, L.; Pospisilik, J.A.; Lee, H.J.; Hanada, R.; Joshi, P.A.; Aliprantis, A.; Glimcher, L.; et al. Osteoclast differentiation factor RANKL controls development of progestin-driven mammary cancer. Nature 2010, 468, 98–102. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Suarez, E.; Jacob, A.P.; Jones, J.; Miller, R.; Roudier-Meyer, M.P.; Erwert, R.; Pinkas, J.; Branstetter, D.G.; Dougall, W.C. RANK ligand mediates progestin-induced mammary epithelial proliferation and carcinogenesis. Nat. Cell Biol. 2010, 468, 103–107. [Google Scholar] [CrossRef]
- Tsubaki, M.; Komai, M.; Fujimoto, S.-I.; Itoh, T.; Imano, M.; Sakamoto, K.; Shimaoka, H.; Takeda, T.; Ogawa, N.; Mashimo, K.; et al. Activation of NF-κB by the RANKL/RANK system up-regulates snail and twist expressions and induces epithelial-to-mesenchymal transition in mammary tumor cell lines. J. Exp. Clin. Cancer Res. 2013, 32, 62. [Google Scholar] [CrossRef] [Green Version]
- Palafox, M.; Ferrer, I.; Pellegrini, P.; Vila, S.; Hernandez-Ortega, S.; Urruticoechea, A.; Climent, F.; Soler, M.T.; Muñoz, P.; Viñals, F.; et al. RANK Induces Epithelial–Mesenchymal Transition and Stemness in Human Mammary Epithelial Cells and Promotes Tumorigenesis and Metastasis. Cancer Res. 2012, 72, 2879–2888. [Google Scholar] [CrossRef] [Green Version]
- Reyes, M.E.; Fujii, T.; Branstetter, D.; Krishnamurthy, S.; Masuda, H.; Wang, X.; Reuben, J.M.; Woodward, W.A.; Edwards, B.J.; Hortobagyi, G.N.; et al. Poor prognosis of patients with triple-negative breast cancer can be stratified by RANK and RANKL dual expression. Breast Cancer Res. Treat. 2017, 164, 57–67. [Google Scholar] [CrossRef]
- Vidula, N.; Yau, C.; Li, J.; Esserman, L.J.; Rugo, H.S. Receptor activator of nuclear factor kappa B (RANK) expression in primary breast cancer correlates with recurrence-free survival and development of bone metastases in I-SPY1 (CALGB 150007/150012; ACRIN 6657). Breast Cancer Res. Treat. 2017, 165, 129–138. [Google Scholar] [CrossRef]
- Pfitzner, B.M.; Branstetter, D.; Loibl, S.; Denkert, C.; Lederer, B.; Schmitt, W.D.; Dombrowski, F.; Werner, M.; Rüdiger, T.; Dougall, W.C.; et al. RANK expression as a prognostic and predictive marker in breast cancer. Breast Cancer Res. Treat. 2014, 145, 307–315. [Google Scholar] [CrossRef]
- Tan, W.; Zhang, W.; Strasner, A.; Grivennikov, S.; Cheng, J.Q.; Hoffman, R.M.; Karin, M. Tumour-infiltrating regulatory T cells stimulate mammary cancer metastasis through RANKL–RANK signalling. Nature 2011, 470, 548–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.S.; Lee, A.; Chae, B.J.; Bae, J.-S.; Song, B.J.; Jung, S.S. Expression of receptor activator of nuclear factor kappa-B as a poor prognostic marker in breast cancer. J. Surg. Oncol. 2014, 110, 807–812. [Google Scholar] [CrossRef]
- Yao, W.; Dai, W.; Shahnazari, M.; Pham, A.; Chen, Z.; Chen, H.; Guan, M.; Lane, N.E. Inhibition of the Progesterone Nuclear Receptor during the Bone Linear Growth Phase Increases Peak Bone Mass in Female Mice. PLoS ONE 2010, 5, e11410. [Google Scholar] [CrossRef] [Green Version]
- Corona, S.P.; Roviello, G.; Strina, C.; Milani, M.; Allevi, G.; Aguggini, S.; Zanoni, D.; Generali, D. Could gonadotropin-releasing hormone analogs be helpful in the treatment of triple-negative breast cancer? Future Oncol. 2017, 13, 2473–2477. [Google Scholar] [CrossRef] [PubMed]
- Köster, F.; Buchholz, S.; Seitz, S.; Schally, A.V.; Engel, J.; Rick, F.G.; Szalontay, L.; Hohla, F.; Krishan, A.; Papadia, A.; et al. Triple-negative breast cancers express receptors for luteinizing hormone-releasing hormone (LHRH) and respond to LHRH antagonist Cetrorelix with growth inhibition. Int. J. Oncol. 2009, 35, 789–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gründker, C.; Föst, C.; Fister, S.; Nolte, N.; Günthert, A.R.; Emons, G. Gonadotropin-releasing hormone type II antagonist induces apoptosis in MCF-7 and triple-negative MDA-MB-231 human breast cancer cells in vitro and in vivo. Breast Cancer Res. 2010, 12, R49. [Google Scholar] [CrossRef] [Green Version]
- Schubert, A.; Hawighorst, T.; Emons, G.; Gründker, C. Agonists and antagonists of GnRH-I and -II reduce metastasis formation by triple-negative human breast cancer cells in vivo. Breast Cancer Res. Treat. 2011, 130, 783–790. [Google Scholar] [CrossRef]
- Kwok, C.W.; Treeck, O.; Buchholz, S.; Seitz, S.; Ortmann, O.; Engel, J.B. Receptors for luteinizing hormone-releasing hormone (GnRH) as therapeutic targets in triple negative breast cancers (TNBC). Target. Oncol. 2014, 10, 365–373. [Google Scholar] [CrossRef]
- Lim, E.; Fab, K.C.; Vaillant, F.; Wu, D.; Forrest, N.C.; Pal, B.; Hart, A.H.; Asselin-Labat, M.-L.; Gyorki, D.E.; Ward, T.; et al. Aberrant luminal progenitors as the candidate target population for basal tumor development in BRCA1 mutation carriers. Nat. Med. 2009, 15, 907–913. [Google Scholar] [CrossRef]
- Molyneux, G.; Geyer, F.C.; Magnay, F.-A.; McCarthy, A.; Kendrick, H.; Natrajan, R.; MacKay, A.; Grigoriadis, A.; Tutt, A.; Ashworth, A.; et al. BRCA1 Basal-like Breast Cancers Originate from Luminal Epithelial Progenitors and Not from Basal Stem Cells. Cell Stem Cell 2010, 7, 403–417. [Google Scholar] [CrossRef] [Green Version]
- Nolan, E.; Kathleen Cuningham Foundation Consortium for research into Familial Breast cancer (kConFab); Vaillant, F.; Branstetter, D.; Pal, B.; Giner, G.; Whitehead, L.; Lok, S.W.; Mann, G.B.; Rohrbach, K.; et al. RANK ligand as a potential target for breast cancer prevention in BRCA1-mutation carriers. Nat. Med. 2016, 22, 933–939. [Google Scholar] [CrossRef]
- Barton, V.N.; D’Amato, N.C.; Gordon, M.A.; Christenson, J.L.; Elias, A.; Richer, J.K. Androgen Receptor Biology in Triple Negative Breast Cancer: A Case for Classification as AR+ or Quadruple Negative Disease. Horm. Cancer 2015, 6, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, B.D.; Jovanović, B.; Chen, X.; Estrada, M.V.; Johnson, K.N.; Shyr, Y.; Moses, H.L.; Sanders, M.E.; Pietenpol, J.A. Refinement of Triple-Negative Breast Cancer Molecular Subtypes: Implications for Neoadjuvant Chemotherapy Selection. PLoS ONE 2016, 11, e0157368. [Google Scholar] [CrossRef]
- Rahim, B.; O’Regan, R. AR Signaling in Breast Cancer. Cancers 2017, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Vera-Badillo, F.E.; Templeton, A.J.; De Gouveia, P.; Diaz-Padilla, I.; Bedard, P.L.; Al-Mubarak, M.; Seruga, B.; Tannock, I.F.; Ocana, A.; Amir, E. Androgen Receptor Expression and Outcomes in Early Breast Cancer: A Systematic Review and Meta-Analysis. J. Natl. Cancer Inst. 2014, 106, djt319. [Google Scholar] [CrossRef]
- Wang, C.; Pan, B.; Zhu, H.; Zhou, Y.; Mao, F.; Lin, Y.; Xu, Q.; Sun, Q. Prognostic value of androgen receptor in triple negative breast cancer: A meta-analysis. Oncotarget 2016, 7, 46482–46491. [Google Scholar] [CrossRef] [Green Version]
- Swerdloff, R.S.; Dudley, R.E.; Page, S.T.; Wang, C.; Salameh, W.A. Dihydrotestosterone: Biochemistry, Physiology, and Clinical Implications of Elevated Blood Levels. Endocr. Rev. 2017, 38, 220–254. [Google Scholar] [CrossRef]
- Chen, F.; Knecht, K.; Birzin, E.; Fisher, J.; Wilkinson, H.; Mojena, M.; Moreno, C.T.; Schmidt, A.; Harada, S.-I.; Freedman, L.P.; et al. Direct Agonist/Antagonist Functions of Dehydroepiandrosterone. Endocrinology 2005, 146, 4568–4576. [Google Scholar] [CrossRef] [Green Version]
- Labrie, F.; Bélanger, A.; Cusan, L.; Candas, B. Physiological Changes in Dehydroepiandrosterone Are Not Reflected by Serum Levels of Active Androgens and Estrogens but of Their Metabolites: Intracrinology. J. Clin. Endocrinol. Metab. 1997, 82, 2403–2409. [Google Scholar] [CrossRef] [PubMed]
- Recchione, C.; Venturelli, E.; Manzari, A.; Cavalleri, A.; Martinetti, A.; Secreto, G. Testosterone, dihydrotestosterone and oestradiol levels in postmenopausal breast cancer tissues. J. Steroid Biochem. Mol. Biol. 1995, 52, 541–546. [Google Scholar] [CrossRef]
- Page, S.T.; Lin, D.W.; Mostaghel, E.A.; Marck, B.T.; Wright, J.L.; Wu, J.; Amory, J.K.; Nelson, P.S.; Matsumoto, A.M. Dihydrotestosterone Administration Does Not Increase Intraprostatic Androgen Concentrations or Alter Prostate Androgen Action in Healthy Men: A Randomized-Controlled Trial. J. Clin. Endocrinol. Metab. 2011, 96, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-C.; Chen, C.-K.; Chen, M.-J.; Lin, J.-C.; Lin, C.-H.; Huang, W.-C.; Cheng, S.-P.; Chen, S.-N.; Liu, C.-L. Expression of 3?-Hydroxysteroid Dehydrogenase Type 1 in Breast Cancer is Associated with Poor Prognosis Independent of Estrogen Receptor Status. Ann. Surg. Oncol. 2017, 24, 4033–4041. [Google Scholar] [CrossRef]
- Von Wahlde, M.-K.; Hülsewig, C.; Rückert, C.; Götte, M.; Kiesel, L.; Bernemann, C. The anti-androgen drug dutasteride renders triple negative breast cancer cells more sensitive to chemotherapy via inhibition of HIF-1α-/VEGF-signaling. Gynecol. Endocrinol. 2014, 31, 160–164. [Google Scholar] [CrossRef]
- Barton, V.N.; D’Amato, N.C.; Gordon, M.A.; Lind, H.T.; Spoelstra, N.S.; Babbs, B.L.; Heinz, R.E.; Elias, A.; Jedlicka, P.; Jacobsen, B.M.; et al. Multiple Molecular Subtypes of Triple-Negative Breast Cancer Critically Rely on Androgen Receptor and Respond to Enzalutamide In Vivo. Mol. Cancer Ther. 2015, 14, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Yang, F.; Zhang, W.; Song, W.; Liu, Y.; Guan, X. The Androgen Receptor Promotes Cellular Proliferation by Suppression of G-Protein Coupled Estrogen Receptor Signaling in Triple-Negative Breast Cancer. Cell. Physiol. Biochem. 2017, 43, 2047–2061. [Google Scholar] [CrossRef]
- Blankvoort, B.; de Groene, E.; van Meeteren-Kreikamp, A.; Witkamp, R.; Rodenburg, R.; Aarts, J. Development of an Androgen Reporter Gene Assay (AR-LUX) Utilizing a Human Cell Line with an Endogenously Regulated Androgen Receptor. Anal. Biochem. 2001, 298, 93–102. [Google Scholar] [CrossRef]
- Kemppainen, J.A.; Langley, E.; Wong, C.-I.; Bobseine, K.; Kelce, W.R.; Wilson, E.M. Distinguishing Androgen Receptor Agonists and Antagonists: Distinct Mechanisms of Activation by Medroxyprogesterone Acetate and Dihydrotestosterone. Mol. Endocrinol. 1999, 13, 440–454. [Google Scholar] [CrossRef]
- Sasagawa, S.; Shimizu, Y.; Kami, H.; Takeuchi, T.; Mita, S.; Imada, K.; Kato, S.; Mizuguchi, K. Dienogest is a selective progesterone receptor agonist in transactivation analysis with potent oral endometrial activity due to its efficient pharmacokinetic profile. Steroids 2008, 73, 222–231. [Google Scholar] [CrossRef]
- Casson, P.R. Effect of Postmenopausal Estrogen Replacement on Circulating Androgens. Obstet. Gynecol. 1997, 90, 995–998. [Google Scholar] [CrossRef]
- Simon, J.A. Estrogen replacement therapy: Effects on the endogenous androgen milieu. Fertil. Steril. 2002, 77, 77–82. [Google Scholar] [CrossRef]
- Ali, I.; James, T.; Meston, N.; Wojnarowska, F.; Shine, B. Age and hormone replacement therapy as factors influencing androgen levels in the postmenopausal female. Clin. Chem. Lab. Med. 2009, 47, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Samuel, S.L.; Zhai, G.; Rana, S.; Taylor, M.; Umphrey, H.R.; Oelschlager, D.K.; Buchsbaum, D.J.; Zinn, K.R. Combination therapy with anti-DR5 antibody and tamoxifen for triple negative breast cancer. Cancer Biol. Ther. 2014, 15, 1053–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, V.M.; Jeyakumar, M.; Carlson, K.E.; Katzenellenbogen, J.A. Diarylpropionitrile (DPN) Enantiomers: Synthesis and Evaluation of Estrogen Receptor β-Selective Ligands. J. Med. Chem. 2011, 55, 528–537. [Google Scholar] [CrossRef] [Green Version]
- Weiser, M.J.; Wu, T.J.; Handa, R.J. Estrogen Receptor-β Agonist Diarylpropionitrile: Biological Activities of R- and S-Enantiomers on Behavior and Hormonal Response to Stress. Endocrinology 2008, 150, 1817–1825. [Google Scholar] [CrossRef]
- Duong, B.N.; Elliott, S.; Frigo, D.E.; Melnik, L.I.; Vanhoy, L.; Tomchuck, S.; Lebeau, H.P.; David, O.; Beckman, B.S.; Alam, J.; et al. AKT Regulation of Estrogen Receptor β Transcriptional Activity in Breast Cancer. Cancer Res. 2006, 66, 8373–8381. [Google Scholar] [CrossRef] [Green Version]
- Shaaban, A.M.; Green, A.R.; Karthik, S.; Alizadeh, Y.; Hughes, T.A.; Harkins, L.; Ellis, I.O.; Robertson, J.F.; Paish, E.C.; Saunders, P.T.; et al. Nuclear and Cytoplasmic Expression of ERβ1, ERβ2, and ERβ5 Identifies Distinct Prognostic Outcome for Breast Cancer Patients. Clin. Cancer Res. 2008, 14, 5228–5235. [Google Scholar] [CrossRef] [Green Version]
- Baek, J.-M.; Chae, B.-J.; Song, B.-J.; Jung, S.-S. The potential role of estrogen receptor β2 in breast cancer. Int. J. Surg. 2015, 14, 17–22. [Google Scholar] [CrossRef]
- Faria, M.; Karami, S.; Granados-Principal, S.; Dey, P.; Verma, A.; Choi, D.S.; Elemento, O.; Bawa-Khalfe, T.; Chang, J.C.; Strom, A.M.; et al. The ERβ4 variant induces transformation of the normal breast mammary epithelial cell line MCF-10A; the ERβ variants ERβ2 and ERβ5 increase aggressiveness of TNBC by regulation of hypoxic signaling. Oncotarget 2018, 9, 12201–12211. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Tang, L.; Xu, Y.; Sun, Q.; Yang, F.; Guan, X. ERβ1 inhibits metastasis of androgen receptor-positive triple-negative breast cancer by suppressing ZEB1. J. Exp. Clin. Cancer Res. 2017, 36, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anestis, A.; Sarantis, P.; Theocharis, S.; Zoi, I.; Tryfonopoulos, D.; Korogiannos, A.; Koumarianou, A.; Xingi, E.; Thomaidou, D.; Kontos, M.; et al. Estrogen receptor beta increases sensitivity to enzalutamide in androgen receptor-positive triple-negative breast cancer. J. Cancer Res. Clin. Oncol. 2019, 145, 1221–1233. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| ERβ | Direct Effect | |
|---|---|---|
| Favorable | Unfavorable | |
| ||
| ERβ | Indirect Effect | |
| Favorable | Unfavorable | |
| ||
| RANK | Indirect Effect | |
| Favorable | Unfavorable | |
| AR | Direct Effect | |
| Favorable | Unfavorable | |
| AR | Indirect Effect | |
| Favorable | Unfavorable | |
| GPER | Direct Effect | |
| Favorable | Unfavorable | |
| ||
| GPER | Indirect Effect | |
| Favorable | Unfavorable | |
| ||
| ERα and PR | In the event of receptor conversion, possible stimulation of growth and metastatic potential [12] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Barele, M.; Heemskerk-Gerritsen, B.A.M.; Louwers, Y.V.; Vastbinder, M.B.; Martens, J.W.M.; Hooning, M.J.; Jager, A. Estrogens and Progestogens in Triple Negative Breast Cancer: Do They Harm? Cancers 2021, 13, 2506. https://doi.org/10.3390/cancers13112506
van Barele M, Heemskerk-Gerritsen BAM, Louwers YV, Vastbinder MB, Martens JWM, Hooning MJ, Jager A. Estrogens and Progestogens in Triple Negative Breast Cancer: Do They Harm? Cancers. 2021; 13(11):2506. https://doi.org/10.3390/cancers13112506
Chicago/Turabian Stylevan Barele, Mark, Bernadette A. M. Heemskerk-Gerritsen, Yvonne V. Louwers, Mijntje B. Vastbinder, John W. M. Martens, Maartje J. Hooning, and Agnes Jager. 2021. "Estrogens and Progestogens in Triple Negative Breast Cancer: Do They Harm?" Cancers 13, no. 11: 2506. https://doi.org/10.3390/cancers13112506