
Combined Effects of Myeloid Cells in the Neuroblastoma Tumor Microenvironment


UCL Institute of Child Health, Developmental Biology and Cancer Section, University College London, London WC1N 1EH, UK
*
Author to whom correspondence should be addressed.
Cancers 2021, 13(7), 1743; https://doi.org/10.3390/cancers13071743
Submission received: 15 March 2021
/
Revised: 2 April 2021
/
Accepted: 2 April 2021
/
Published: 6 April 2021
(This article belongs to the Special Issue The Tumor Microenvironment of Neuroblastoma)
Abstract
:Simple Summary
Neuroblastoma is a type of childhood solid cancer often characterized by resistance to treatment and with poor prognosis. The tumors contain a mixture of cell types as well as cancer cells, and one of the most common is a type of blood cells called myeloid cells. In this review article we present a summary of what is known about the function of myeloid cells within neuroblastoma tumors.
Abstract
Despite multimodal treatment, survival chances for high-risk neuroblastoma patients remain poor. Immunotherapeutic approaches focusing on the activation and/or modification of host immunity for eliminating tumor cells, such as chimeric antigen receptor (CAR) T cells, are currently in development, however clinical trials have failed to reproduce the preclinical results. The tumor microenvironment is emerging as a major contributor to immune suppression and tumor evasion in solid cancers and thus has to be overcome for therapies relying on a functional immune response. Among the cellular components of the neuroblastoma tumor microenvironment, suppressive myeloid cells have been described as key players in inhibition of antitumor immune responses and have been shown to positively correlate with more aggressive disease, resistance to treatments, and overall poor prognosis. This review article summarizes how neuroblastoma-driven inflammation induces suppressive myeloid cells in the tumor microenvironment and how they in turn sustain the tumor niche through suppressor functions, such as nutrient depletion and generation of oxidative stress. Numerous preclinical studies have suggested a range of drug and cellular therapy approaches to overcome myeloid-derived suppression in neuroblastoma that warrant evaluation in future clinical studies.
1. Introduction
As one of the more recently introduced treatments in neuroblastoma, immunotherapy targeting tumors via the abundantly expressed tumor-associated antigen disialoganglioside GD2 has shown promising results [1]. While consolidative therapy with anti-GD2 monoclonal antibodies (mAb), such as dinutuximab, has already been integrated into standard protocols for high-risk patients, ongoing clinical studies investigating chimeric antigen receptor (CAR) T cells in neuroblastoma have revealed limited responses despite preclinical reports of efficacy [2]. Demonstrated issues included minimal CAR T expansion as well as reduced immune activation when targeting GD2 with CAR T cells [3,4,5,6], suggesting the influence of an immunosuppressive mechanism at play.
One of the reasons neuroblastoma is hard to treat with immunity-based approaches is its inflammatory character, which leads to the formation of a highly immunosuppressive microenvironment consisting of soluble factors, stromal cells, and alternatively activated immune effectors. Studies investigating neuroblastoma in transgenic TH-MYCN mouse models have shown that tumors undergo a transition from early lesions highly infiltrated with lymphocytes to progressively increasing numbers of myeloid populations in advanced tumors [7,8]. This was further confirmed in immunohistochemical analyses of human tumor samples, where neuroblastoma and other pediatric solid tumors were heavily infiltrated with Cluster of differentiation (CD) 68+ myeloid cells [9]. We now know that these myeloid populations predominantly consist of type 2 tumor-associated macrophages (TAM) and myeloid-derived suppressor cells (MDSC) [10,11,12]. This is reflected in a 14-gene signature associated with poor prognosis in neuroblastoma, which contains five genes related to myeloid populations and their suppressive mechanisms (CD14, CD33, IL-10, CD16, IL-6R) [10].
Therefore, detailed understanding of the induction and suppressive mechanisms of these tumor microenvironment (TME)-associated myeloid populations in solid malignancies, including neuroblastoma, is crucial for the design of novel immunotherapeutic treatments and combinatorial approaches. This review will discuss cellular components of the immunosuppressive microenvironment of neuroblastoma, with a focus on TAM- and MDSC-mediated suppression, and how these myeloid populations can be targeted in the immunotherapy of neuroblastoma.
2. The Suppressive Tumor Microenvironment of Neuroblastoma
Neuroblastoma is characterized by its complex immunosuppressive microenvironment. The combined effects of tumor cells, stromal components, and infiltrating immune cells leads to an inflammatory, hypoxic, vascular, and angiogenic niche that supports tumorigenesis and immune evasion [13,14,15]. Inhibitory immune effectors like TAMs, MDSCs, and Treg cells infiltrate tumors as the lesion progresses, contributing to increasingly immunosuppressive sites [8]. Indeed, Asgharzadeh et al. have identified 14 genes associated with poor outcome in metastatic neuroblastoma, of which 5 were inflammation-related genes and 9 were tumor-related genes [10].
Defects in the antigen presenting machinery (APM) and low levels of major histocompatibility complex (MHC) I molecules in neuroblastoma hinder engagement with cytotoxic T cells and their subsequent activation [16,17,18]. T cell function is further inhibited through secretion of soluble factors like transforming growth factor-β (TGF-β), interleukin (IL)-10, and galectin-1 [15,19,20,21]. Moreover, MYCN-dependent downregulation of NKG2D ligands, such as MIC-A, MIC-B, ULBP-1, ULBP-2, and ULBP-3, as well as release of their soluble forms support evasion of innate immune responses, including NK cell activation [22,23,24]. Studies in both murine and human neuroblastoma have demonstrated that the tumor cells can also suppress immunity through arginase (Arg)-2 activity [25]. The resulting depletion of L-arginine leads to downregulation of the CD3z chain on T cells, which induces proliferative arrest of the cells, as well as further contributes to the induction of immunosuppressive phenotypes in the tumor niche [25,26].
Abundantly found in the tumor stroma, mesenchymal stem/stromal cells (MSC) and cancer-associated fibroblasts (CAF) strongly support tumor survival through their angiogenic and immunosuppressive effects. It has been reported in many tumors that MSCs can promote cell invasiveness, migration, and endothelial-to-mesenchymal transition (EMT) of cancer cells. Indeed, studies have found that MSCs isolated from neuroblastoma tumors carried transcriptomic profiles associated with EMT, which in turn were associated with poor prognosis, and expressed high levels of C-X-C Motif Chemokine Ligand 12 (CXCL12), promoting neuroblastoma metastasis and invasiveness [27,28,29]. MSCs also protect the tumor from oxidative stress through the production and activation of antioxidant enzymes against hydrogen peroxide in the TME [30]. In neuroblastoma, CAFs were first recognized as α-smooth muscle actin (α-SMA)-positive cells, which were associated with an increase in microvascular proliferation and poor clinical outcome [31,32]. CAFs are activated by soluble factors in the tumor proximity, such as TGF-β, and in turn release abundant tumor-promoting and angiogenic factors, thus contributing to the establishment of a refractory TME, tumor growth, and disease progression [33]. CAFs promote vascular proliferation through the angiogenic factors, vascular endothelial growth factor A (VEGF-A) and IL-8, and suppress NK and T cells through secretion of anti-inflammatory cytokines like TGF-β and IL-6 [34,35]. Furthermore, the produced factors activate signal transducer and activator of transcription (STAT)3 and extracellular regulated kinase (ERK 1/2)-mediated signaling in neuroblastoma cells, supporting proliferation, survival, and chemokine resistance of the tumor [36]. Like MSCs, CAFs also release chemokines, such as C-C Motif Chemokine Ligand 2 (CCL2), to recruit further infiltrating immune cells to the TME. Indeed, it has been reported that in neuroblastoma, CAFs and TAMs are often in close proximity with reciprocal functionality [32,37]. While CAFs recruit TAMs via CCL2 and shift differentiation towards the M2 phenotype via production of prostaglandin E 2 (PGE2) [38,39], TAMs in turn enhance CAF-mediated STAT3 activation through release of the soluble agonistic IL-6 receptor (IL-6R), and promote CAF invasiveness and proliferation [30,36]. Moreover, Hashimoto et al. have reported that the number of TAMs and the area of CAFs in neuroblastoma tumors were positively correlated with clinical stage and MYCN amplification [37].
In the solid tumor TME, infiltrating T cells can acquire an immunoregulatory phenotype. Regulatory T cells (Tregs) suppress antitumor immunity via pleiotropic inhibitory mechanisms and thus support immune evasion. Indeed, in neuroblastoma mouse models, depletion of Tregs increased the efficacy of T cell-mediated immunotherapy [40,41]. Tregs contribute to the immunosuppressive milieu by secreting inhibitory cytokines, such as IL-10, TGF-β, and IL-35, and disrupt maturation of antigen-presenting cells (APC) through CTLA-4 engagement. Furthermore, Tregs express granzyme and perforin, which mediate cytotoxic effects on effector T cells [33,42,43]. Through their expression of high-affinity receptors for important inflammatory cytokines, such as IL-2, IL-7, IL-12, and IL-15, Tregs act as competitive sinks and further suppress function and activation of cytotoxic T cells [44,45,46].
The local accumulation of C-C motif chemokines, such as CCL2, CCL7, and CXCL1, as well as the hypoxic milieu of neuroblastoma, results in the recruitment of high numbers of myeloid cells [37,47]. As a result of the local availability of cytokines and growth factors, myeloid differentiation is often disrupted in solid tumors, which leads to the generation of heterogeneous immature populations with suppressive functions, generally described as MDSC [48,49]. Moreover, the populations that do undergo differentiation in the solid TME acquire suppressive phenotypes and disrupted functions. Dendritic cells in the TME demonstrate defective antigen presentation function caused by the local effects of cytokines and growth factors, such as IL-6, IL-10, and VEGF [44], and downregulate both MHC and costimulatory receptor expression on their surfaces, resulting in less T cell priming and activation [50]. Studies in solid tumors show that hypoxic factors in the TME, such as hypoxia inducible factor 1α (HIF1α), promote differentiation of MDSCs into TAMs, further contributing to the range of immunosuppressive cell populations [47,51]. Considering their wide array of suppressive mechanisms, their prognostic potential, and their abundance in the neuroblastoma environment, suppressive myeloid populations, including MDSCs and TAMs, present an interesting target for improving immunotherapy of solid tumors. The next sections therefore will focus on their role in cancer progression and immune suppression, and how preclinical studies are attempting to overcome myeloid-mediated suppression in neuroblastoma.
3. Myeloid Cell Populations in Neuroblastoma
3.1. Myeloid Heterogeneity
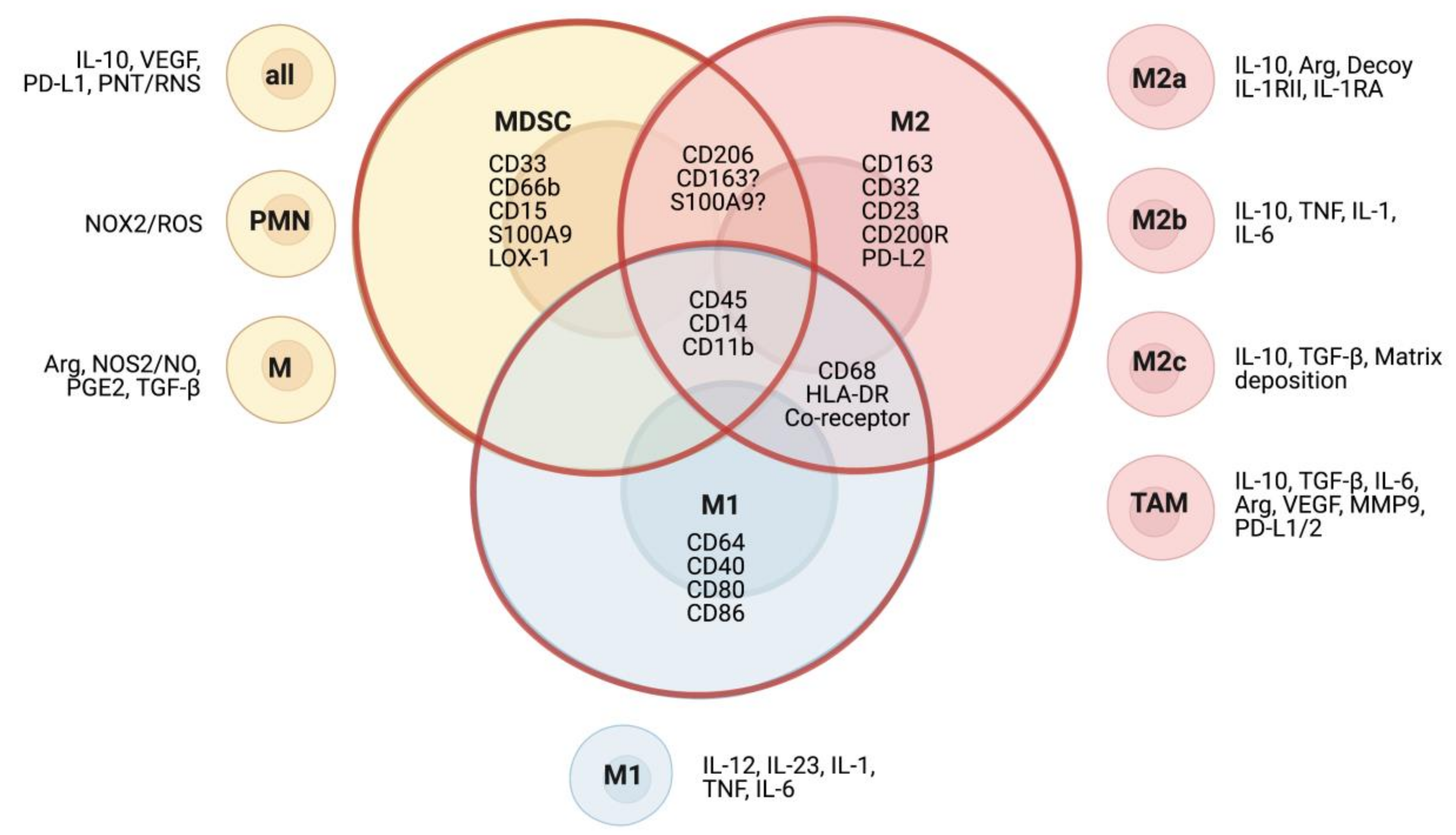
In the TME, cells of myeloid origin are of heterogeneous character and are generally of an immature and immunosuppressive phenotype. Due to the transitional expression of surface markers, phenotypes of the different subsets are difficult to dissect and often overlap (Figure 1). In mice, MDSC can be defined by the markers of Gr-1, Ly6C, and Ly6G, and are discriminated from TAM via the marker F4/80 [52]. In humans, the distinction between the two myeloid subsets is less clear. While MDSC are generally considered immature myeloid cells with low expression of class II MHC, and which do not express the macrophage marker CD68 [53], the marker is not as heterogeneously expressed among macrophages as once thought and may thus lead to false categorization as MDSC. Another marker classically used for the characterization of suppressive macrophages is CD163, but recent studies have found that CD163 can be detected on a variety of human anti-inflammatory myeloid cells, as it is upregulated by IL-10, glucocorticoids, and M-CSF, and has been found on tumor-infiltrating cells with negligible expression of CD68 [54,55,56,57]. Furthermore, in acute liver failure syndromes, a subset of circulating MDSC with anti-inflammatory functions has also been found to express CD163 [58]. The expression of markers considered specific for MDSC and TAM seems to vary [59] and underlines that further efforts have to be made into characterizing subsets correctly, such as recently proposed immunofluorescent multiparameter assays [56].
3.1.1. Tumor-Associated Macrophages
Macrophages are a class of heterogeneous immune cells of myeloid origin which primarily exert phagocytic functions in immune reactions and tissue remodeling [60]. Depending on environmental stimuli, macrophage polarization can result in two main phenotypes: M1 and M2. While M1 macrophages are characterized by their immunogenic functions and are typically activated by immunostimulatory signals lipopolysaccharide (LPS) or interferon gamma (IFN-γ), the M2 phenotype is stimulated by IL-4 and IL-13 and plays an immunoregulatory role in wound healing and tissue remodeling, exerted through the secretion of VEGF, matrix metalloproteinase 9 (MMP9), IL-10, and TGF-β [61,62]. M1 and M2 macrophages do not only differ in their functions but can also be distinguished by their genetic profile [63,64], metabolism [65,66], and epigenetic modifications [67]. Macrophages with M2 polarization can be further characterized into subtypes with differing profiles (Figure 1). M2a macrophages are mainly involved in wound healing through secretion of TGF-β and other factors. Stimulated by toll-like receptor (TLR) or interleukin 1 receptor (IL-1R) agonists, M2b macrophages exert immunoregulatory functions via release of cytokines, such as IL-10, in cancer and other diseases. Lastly, M2c macrophages are involved in tissue remodeling and dampen the immune response through the release of TGF-β and IL-10 [62,68,69]. Importantly, macrophages are a highly heterogeneous and fluid population, which can easily transition between different phenotypes depending on environmental stimuli [70]. The binary M1 and M2 classification is an oversimplification, however due to the current limitations in clearly differentiating subtypes and activation states, it will be used in the subsequent sections of this review.
Macrophages in the TME are collectively described as TAMs and typically mirror the M2 phenotype. Interestingly, their presence has been found to correlate with worse prognosis in neuroblastoma and other solid tumors [10,71]. Furthermore, studies suggest that TAMs can promote tumor progression by upregulating oncogenic MYC expression through activation of the STAT3 pathway in non-MYCN amplified neuroblastoma cells [72].
Expressing M2 macrophage markers like CD163 and CD206 (Table 1), TAMs suppress the immune response through the secretion of regulatory cytokines, such as IL-10 and TGF-β, and promote tumorigenesis via factors like VEGF and MMPs [73]. Within tumors, distinct subpopulations of TAMs can be found, depending on their location, with varying degrees of M2-like character and functions. For example, perivascular TAMs contribute to metastasis, while TAMs at the tumor–stroma interface promote angiogenesis [74,75]. Recent studies in lung carcinoma and other solid tumors have found that hypoxia within the TME promotes differentiation of MDSC to TAMs and shapes their phenotype towards lowered hypoxia-sensitive gene expression and angiogenic activity but does not change the M2-like character [51,76]. This heterogeneity among TAMs within tumors is often overlooked in preclinical studies, which tend to oversimplify TAMs as a single homogeneous population, limiting the informative value of their results [62].
3.1.2. Myeloid-Derived Suppressor Cells
MDSC, a heterogeneous population of immune cells with myeloid lineage, have been known to exist in cancer for over 30 years, but only recently have they caught more attention towards their critical function in shaping the TME [77,78]. Santilli et al. have demonstrated that co-injecting immunocompetent mice with neuroblastoma cells and MDSC greatly increases tumor growth [11]. Furthermore, studies on rhabdomyosarcoma in mice have shown that disruption of CXCR2-dependent MDSC trafficking to the tumor resulted in significantly increased antitumor effects [79]. MDSC can also be found under normal physiological conditions, where they regulate immune tolerance. For example, MDSC mediate maternal–fetal tolerance in pregnancy and have also been found to accumulate with age [80,81,82]. In cancer, the major factor driving MDSC accumulation is the chronic, low-grade inflammation of the TME, where the alternatively activated myeloid cells act through a wide range of mechanisms mediating tumor progression and immune inhibition [83].
Generally, MDSC in cancer describes immature cells of myeloid origin with immunosuppressive character [77]. They have been observed both in mice and in human disease and can be grouped into at least two subpopulations: mononuclear MDSC (M-MDSC) and granulocytic or polymorphonuclear MDSC (PMN-MDSC). Correspondingly, M-MDSC are morphologically similar to mononuclear blood monocytes, while PMN-MDSC resemble polymorphonuclear neutrophils [53]. All MDSC in mouse and human are negative for T cell, B cell, and NK cell lineage markers [78] (Table 1). In mice, MDSC can be characterized by their surface expression of CD11b combined with Ly6C or Ly6G, which are known collectively as Gr-1. Mouse M-MDSC are CD11b+ Ly6C+ Ly6G−/low, whereas PMN-MDSC are CD11b+ Ly6G+ [84,85]. In human, there are no MDSC-specific markers, so that they can only be distinguished using a combination of common immune markers. Human M-MDSC are CD33+ CD11b+ CD14+ CD15− HLA-DR−/low and PMN-MDSC are CD33dim CD11b+ CD15+ CD14− HLA-DR−/low. Furthermore, there is a third population described in human, early-stage MDSC (eMDSC), which are CD33+ CD11b+ HLA-DR−/low but lack myeloid lineage markers (CD14, CD15, CD66b) [53,86,87]. While M-MDSC can be phenotypically distinguished from healthy monocytes by absence or low expression of HLA-DR, there has been an ongoing uncertainty about the differences between PMN-MDSC and neutrophils [88,89]. One way to separate the two populations is by standard Ficoll density gradients (1.077 g/L), where neutrophils band at a higher density than PMN-MDSC, albeit some activated neutrophils may be included in the low-density fraction [90]. More recent advances in whole-genome analysis and flow cytometry of samples from head and neck cancer patients have revealed the lectin-type oxidized low-density lipoprotein receptor 1 (LOX1) as a marker found in human PMN-MDSC, but not in neutrophils [91]. In mice, Cd84 and junctional adhesion molecule have been discussed as markers distinguishing PMN-MDSC from neutrophils [92].
3.2. Contribution of Myeloid Cells to the Tumor Microenvironment
3.2.1. The Role of TAMs
Recruited by chemoattractants including CCL2, CCL20, CXCL12, and CSF-1 [37,93,94,95], macrophages in the proximity of solid tumors are quickly reprogrammed towards an M2-like phenotype in a process driven by the local hypoxia [96]. The polarized TAMs then contribute to tumor progression, angiogenesis, and immunosuppression by expressing factors such as IL-6, VEGF, inducible nitric oxide synthase (iNOS), and arginase [62,96]. TAMs have been characterized in a range of solid tumors, which can provide indications for their contributions in neuroblastoma tumorigenesis.
Contribution to Tumor Progression
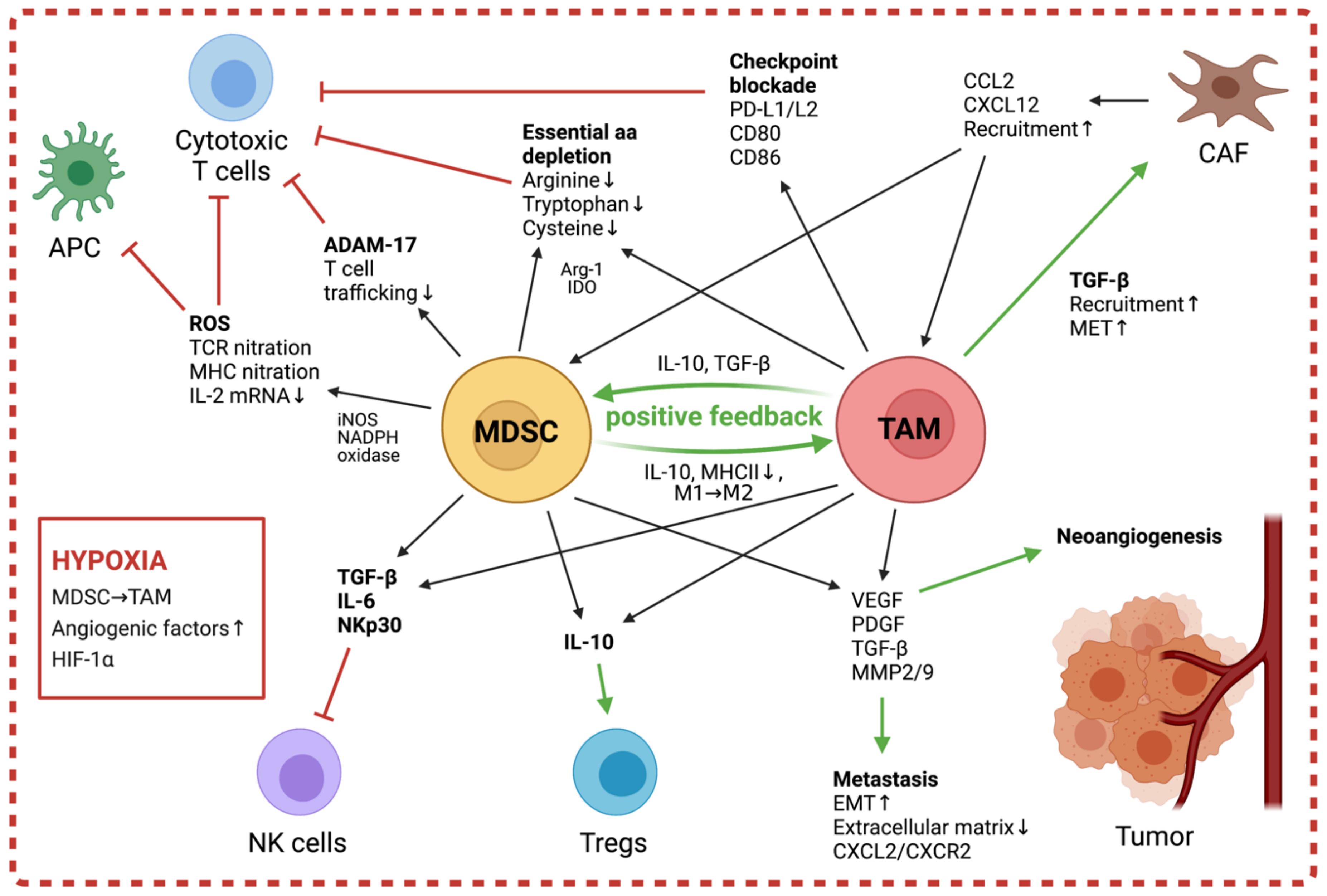
Progression of neuroblastoma and other solid tumors is promoted by TAMs through the secretion of factors like VEGF, Platelet-derived growth factor (PDGF), TGF-β, MMP2, and MMP9, which lead to extracellular matrix remodeling and increased neoangiogenesis [97,98] (Figure 2). It has been demonstrated that the pro-angiogenic niche in tumors result from the combined effects of TAMs and local hypoxia, and that in neuroblastoma, HIF2a and TAMs are markers of the perivascular niche [99,100]. The upregulated levels of HIF1α/2α in TAMs further promote angiogenesis through inducing expression of pro-angiogenic factors, including VEGF, TNFα, and MMP [93,96]. These factors released by TAMs also contribute to tumor metastasis and facilitate both the tumor’s release from the primary site and the establishment at secondary sites by increasing vessel permeability [73]. Furthermore, Hashimoto et al. have shown that in neuroblastoma, TAMs enhance tumor invasion through increased expression of CXCL2 and interactions with its receptor CXCR2 on neuroblastoma cells [37].
Contribution to the Immunosuppressive Microenvironment
TAMs contribute to the immune suppression in the TME both through stimulation of other suppressive populations, such as Tregs and MDSC, and through direct inhibition of effectors including NK cells and cytotoxic T cells [62] (Figure 2).
Studies in neuroblastoma have demonstrated that tumor cells induce suppressive macrophage populations, which inhibit effector NK cells via secretion of IL-6 and TGF-β and thus dampen the efficiency of anti-GD2-targeted immunotherapy [19,101]. TAMs also exert a suppressive effect on infiltrating cytotoxic T cells, which is mediated both by their release of inhibitory cytokines as well as through direct interactions with T cell surface receptors. In human lung squamous-cell carcinomas, melanoma, and other solid tumors, TAMs simulate APM interactions and thus prevent the T cell effectors from binding to functional APCs, while activating checkpoint blockade through engagement with inhibitory co-receptors PD-1 and CTLA-4 [102,103,104]. Moreover, TAMs express the enzymes Arg-1 and indoleamine 2,3-dioxygenase (IDO)-1/2, leading to a local metabolism of L-arginine and L-tryptophan respectively, which are essential for T cell activation [105]. Through their release of soluble factors, TAMs actively shape the immunosuppressive character of the TME by affecting other suppressive populations. Indeed, TAM-secreted CCL2 recruits Tregs, which in turn promote differentiation of M2 macrophages and exert their immunosuppressive functions via IL-10 and TGF-β release [105,106]. In prostate carcinoma, the secreted TGF-β also mediates recruitment and enhancement of CAFs in the TME by facilitating mesenchymal-to-epithelial transition (MET) [107]. This increase in local CAFs leads to higher levels of stromal cell-derived factor 1 (SDF1/CXCL12), thus further enforcing infiltration of TAMs and endothelial progenitor cells to promote angiogenesis [37,95,108]. Via the release of CCL2, TAMs recruit further myeloid populations to the tumor, and through the released factors IL-10 and TGF-β, mitigate induction of immunosuppressive MDSC and suppression of APC recruitment and function [109]. In turn, the MDSC produce more IL-10 and can quickly differentiate into M2-like macrophages, providing a positive feedback towards more immunosuppression, as shown in several solid tumor models [51] (Figure 2).
3.2.2. The Role of MDSC
There is only limited data available on the clinical significance of MDSC in neuroblastoma. Studies in neuroblastoma mouse models have reported the accumulation of MDSC during tumor progression and that MDSC-released factors including Reactive oxygen species (ROS)-, Arg-1-, and TGF-β-mediated tumor growth [7,110]. Indeed, Santilli et al. observed a drastic increase of immunosuppressive MDSC-like cells in the blood and tumors of neuroblastoma patients [11].
The predominant population in tumor-bearing mice and cancer patients are often PMN-MDSC, however there are differences between cancer types and inconclusive reports about which population is the most suppressive [49,78]. It has been shown in mice that M-MDSC can be considered more immune suppressive than PMN-MDSC on a per-cell basis. Studies in patients with head and neck and urothelial cancer, however, have demonstrated a more suppressive per-cell activity of the PMN-MDSC populations [86]. To our knowledge, there are no studies comparing the suppressive potential of MDSC subsets in neuroblastoma, so the predominant suppressive subset is not known.
Recruitment and Activation of MDSC
During hematopoiesis, hematopoietic stem cells (HSC) in the bone marrow differentiate into common myeloid progenitor cells (CMPs), which further give rise to granulocytes, DC, and macrophages. When exposed to stress conditions, such as cancer-associated inflammation, this healthy differentiation process is perturbed, leading to the accumulation of immature myeloid cells during so-called emergency myelopoiesis. Excessive levels of tumor-produced granulocyte colony-stimulating factor (G-CSF) and granulocyte-macrophage colony-stimulating factor (GM-CSF) cause the faulty myelopoiesis through downregulation of the transcription factor interferon regulatory factor 8 (IRF8) and alternatively activate these MDSC, equipping them with potent immunosuppressive activity [111,112,113].
The accumulation of MDSC in tumor-bearing individuals has been shown to depend on the inflammatory mediators IL-1β, IL-6, and Prostaglandin-E2 (PGE2) [114,115,116,117,118]. A crucial pathway for this is via the key MDSC activator STAT3. Activation of STAT3 induces upregulation of p47phox and gp91, which in turn increases the production of reactive oxygen species by MDSC, as shown in subcutaneous tumor models in mice [119,120]. STAT3 activation in MDSC is facilitated by G-CSF and GM-CSF [121,122,123,124], hypoxia in the TME [51], and IL-1β and IL-6 [114,115,125,126].
Another activator of MDSC is the highly proinflammatory alarmin S100A8/A9 complex, which has been demonstrated to activate Nuclear Factor Kappa B (NF-κB) and STAT3 pathways by binding TLR4 and carboxylated N-glycans on the receptor for advanced glycation end products (RAGE) on MDSC in colon carcinoma-bearing mice [78,127]. Sinha et al. have also demonstrated that S100A8/A9 acts as a chemoattractant for MDSC in solid tumors and that MDSC secrete S100A8 and S100A9 in the serum of tumor-bearing mice, thus creating an autocrine feedback loop [128]. Given its impact on MDSC accumulation and activation, S100A8/A9 has been suggested as a novel biomarker for human MDSC [129], which is yet to be elucidated in neuroblastoma.
Promotion of Tumor Progression
MDSC not only exert significant immunosuppressive effects but can also promote tumor progression through non-immunogenic mechanisms (Figure 2). Studies in tumor-bearing mice and patients with triple-negative breast cancer demonstrated MDSC-mediated induction of angiogenesis and metastasis through the production of Mmp9 [130,131]. The protein promotes neo-angiogenesis through increasing VEGF release and facilitates invasion of tumor cells through the degradation of the extracellular matrix of healthy tissue. Metastasis is further supported through increased epithelial–mesenchymal transition induced by MDSC, which has been reported to be dependent on TGF-β, Epidermal growh factor and hepatocyte growth factor (EGF, and HGF) signaling pathways in a melanoma mouse model [132]. In mice with lung adenomas and adenocarcinomas, MDSC protect tumor cells from senescence by secreting IL-1 receptor agonist, thus blocking the production of pro-senescence factors and promoting tumor growth [133].
Contribution to the Immunosuppressive Microenvironment
Early studies in breast cancer models revealed that MDSC exert their immunological functions through the release of soluble mediators, which rely on cell contact because of their short half-life and concentration [134]. More recent investigations have additionally demonstrated the existence of MDSC-derived exosomes with suppressive activity and the ability to shape other suppressive populations in the TME [135,136,137]. MDSC influence a wide range of immune cells within the TME through their heterogeneous mechanisms. While T cells are their key target, with suppression of T cell proliferation or cytokine release considered one of the hallmarks of defining MDSC [53], other effectors of immunity, including NK cells, APCs, Tregs, and B cells, have also been described to be targeted by MDSC-mediated measures (Figure 2). Importantly, although MDSC possess a multitude of mechanisms to interfere with pathways of the immune system, myeloid populations in the TME are heterogeneous and the predominant mechanisms may change depending on differentiation stage, extracellular milieu, and cellular composition of tumors [78].
MDSC suppress T cells through (1) limiting amino acid availability, (2) disruption of T cell functions through production of reactive species, and (3) prevention of trafficking. A key MDSC marker, Arg-1, attenuates T cell activation through depletion of L-arginine, which is essential for manufacture of the T cell receptor (TCR) ζ chain [138,139,140]. Furthermore, MDSC reduce the availability of L-tryptophan by expressing IDO [141] and decrease extracellular levels of the essential amino acid cysteine by hoarding the crucial building block cystine and suppressing cysteine release in APCs [142]. Alongside arg-1, reactive oxygen/nitrogen species (ROS/RNS) production is found in both human and mouse MDSC and therefore serves as a MDSC biomarker [53]. MDSC subsets can generate both ROS and RNS, of which ROS are predominantly generated by PMN-MDSC [49,143]. ROS production involves phosphorylation of STAT3, which in turn upregulates expression of p47phox and gp91, two subunits of the ROS-generating NADPH oxidase. This facilitates superoxide to react with nitric oxide (NO), generated by iNOS-mediated breakdown of L-arginine, to produce peroxynitrite (PNT) [53,120,144]. PNT inhibits T cell activation through nitration of the TCR as well as MHC nitration on APCs, preventing antigen recognition [145,146]. Moreover, PNT nitrates chemokines important for T cell recruitment and infiltration [147]. Studies in breast cancer have shown that MDSC also prevent T cell activation in the lymph node through expression of the enzyme A disintegrin and metalloprotease 17 (ADAM-17), which cleaves L-selectin, a crucial surface molecule for T cell extravasation from the blood and lymphatics into the lymph nodes [148,149]. Furthermore, in hepatocellular carcinoma, MDSC disrupt the T cell stimulatory activity of DCs through secretion of IL-10, which inhibits TLR-ligand-induced IL-12 production of DCs [150]. Studies in mice models of metastatic colon cancer have demonstrated that MDSC also mediate the development of immune-suppressive Tregs through CD40–CD40L interaction in an IFN-γ- and IL-10-dependent process [151,152]. In mouse and human carcinomas, MDSC-mediated suppression of NK cell functions is cell contact-dependent and mainly exerted via engagement with NKp30 on NK cells [153,154]. In mice, a novel Ly6C-negative MDSC subset has been reported to target NK cells after activation by tumor-released IL-1β [154,155], whereas in melanoma patients, M-MDSC are activated by PGE2 and impair NK cell-mediated cytotoxicity through the release of TGF-β [117].
Lastly, the predominant myeloid populations within the TME work together in sustaining the suppressive niche through a feedback-loop dependent on IL-1β and MDSC-expressed TLR4. The crosstalk between MDSC and macrophages enhances TAM protumor activity: MDSC inhibit antitumor M1-like macrophages by secretion of IL-10 and induce downregulation of MHC class II on macrophages, thus attenuating M2-like phenotypes. In turn, macrophages promote MDSC production of IL-10, creating a positive feedback loop towards the creation of TAM [156,157,158]. Furthermore, when entering hypoxic regions of solid tumors, M-MDSC rapidly differentiate to TAM [51].
4. Therapeutic Strategies to Target Suppressive Myeloid Cells
The potent immunoregulatory microenvironment of solid cancers, like neuroblastoma, poses an important hurdle to overcome in immunotherapy. With their heterogeneous range of suppressive mechanisms, myeloid cells have become a major target to consider for combinatorial approaches with CAR T cells, checkpoint inhibitors, and NK cell-based therapies. Indeed, it has been demonstrated that not only do myeloid phenotypes positively correlate with CAR T therapy failure [159], but also that ablation of these populations can improve antitumor effects of various immunotherapeutics [160,161,162,163]. Strategies to overcome myeloid-mediated suppression of the immune response can be generally categorized into (1) prevention of recruitment to the tumor, (2) depletion of myeloid populations or disruption of their suppressive mechanisms, and (3) repolarization towards immunostimulatory phenotypes. An overview of published studies targeting myeloid cells and their underlying strategies can be found in Table 2.
4.1. Disruption of Myeloid Recruitment
Both MDSC and TAM are recruited to the TME by chemoattractant molecules, so interference with binding of these molecules to their corresponding receptors can prevent the accumulation of myeloid cells close to the tumor and subsequently their immunoinhibitory and tumor-promoting effects. An interesting target is the CSF-1/CSF-1R axis, as it is involved in myeloid recruitment to the TME and differentiation into M2 phenotypes [164]. Indeed, studies in various solid malignancies have shown that blockade of the axis with monoclonal antibodies or CSF-1R inhibitors can lead to reduced myeloid numbers at the tumor site [165,166] and increased numbers of macrophages with antitumor activity [167]. Studying the spontaneous neuroblastoma TH-MYCN mouse model, Mao et al. have demonstrated that CSF-1R inhibition efficiently slows tumor progression in tumor-bearing mice and enhances PD-1/PD-L1 antibody therapy [160]. Furthermore, Webb et al. have shown that reduction of myeloid cells via CSF-1R blockade can also improve the efficiency of chemotherapy in a human neuroblastoma model in T cell-deficient mice, indicating a beneficial effect independent from the adaptive immune response [94]. The CCR2-CCL-2 axis is another pathway important in tumor-mediated myeloid recruitment. Albeit not investigated in neuroblastoma, studies in solid tumors show that the axis can be disrupted either by inhibiting CCR2 [168,169] or via anti-CCL2 antibodies [170,171,172], thereby preventing the recruitment of suppressive myeloid cells and initiation of their tumor-promoting effects. Furthermore, Carlson et al. have provided evidence that the tumor-associated inflammation in neuroblastoma can pose another therapeutic target to prevent the accumulation of suppressive effectors in the TME [7]. When treating TH-MYCN mice with low-dose aspirin, they reported a significant reduction in tumor burden, tumor-associated innate cells, including MDSC and TAM, and intra-tumoral TGF-β.
4.2. Depletion of Myeloid Populations
Alternatively, suppression exerted by myeloid cells in the TME can be counteracted either by disrupting their inhibitory mechanisms or depleting the myeloid populations entirely. Several agents have been reported to have cytotoxic effects on myeloid cells in solid tumors, among them chemotherapeutics, such as gemcitabine [173,174,175], 5-fluorouracil [176,177], and trabectedin [178,179]. The latter caused selective depletion of monocytes and macrophages in mouse models of fibrosarcoma, ovarian carcinoma, and Lewis lung carcinoma, as well as in patients with soft tissue sarcoma, while sparing lymphocytes and neutrophils [178]. Siebert et al., moreover, have reported beneficial effects of anti-CD11b antibody therapy in neuroblastoma-bearing mice in combination with anti-GD2 antibody therapy [180]. The receptor tyrosine kinase inhibitor sunitinib has also been suggested to induce myeloid depletion via interference with STAT3 signaling in several solid tumors, including renal cell carcinoma, melanoma, and fibrosarcoma [181,182,183,184]. Renal cell carcinoma patients treated with sunitinib have superior expansion of intratumor lymphocytes and decreased numbers of MDSC both in the tumor and in circulation [181,183]. In pediatric patients with solid malignancies, sunitinib treatment also results in decreased monocyte counts [185], however effects in neuroblastoma patients remain to be investigated.
One of the key inhibitory mechanisms of suppressive myeloid cells which has an established role in neuroblastoma is arginase activity [25]. Indeed, inhibition of arginase recovers T cell proliferation in vitro and reduces tumor growth in mouse models of colon, lung, skin, and breast cancer [186,187], and has shown beneficial effects in combination with checkpoint blockade, cell-based immunotherapy, and chemotherapy [186], indicating a potential for similar effects in neuroblastoma. The depletion of the essential amino acid tryptophan mediated by suppressive myeloid cells can also be overcome in various solid tumors through treatment with IDO inhibitors [188,189] and COX2 inhibitors [8,190,191].
4.3. Repolarization of Myeloid Cells
Suppressive myeloid cells can also be reprogrammed into their immunostimulatory counterparts by a range of therapeutics. These strategies typically function by blocking receptors inducing inhibitory signals or by providing exogenous ligands for receptors activating pro-inflammatory pathways [192]. One strategy to repolarize myeloid cells is targeting histone deacetylase (HDAC) proteins or epigenetic reader proteins [193,194]. One example for this is the HDAC inhibitor vorinostat, which has been shown to promote M1 repolarization of TAM as well as decreasing number and function of MDSC in an in vivo neuroblastoma study [195]. Kroesen et al. have further demonstrated that the drug mediates a synergistic effect with anti-GD2 monoclonal antibodies, making it an interesting candidate for combination immunotherapy. Similarly, HDAC inhibitor, entinostat, has been reported to suppress MDSC-mediated effects and enhance checkpoint inhibitor therapy in mouse models of breast cancer, pancreatic cancer, lung, and renal cell carcinoma [196,197]. Another agent described to induce the repolarization of myeloid cells is all-trans retinoic acid (ATRA), which upregulates expression of glutathione synthase in MDSC. The resulting accumulation of intracellular glutathione neutralizes their high ROS production and stimulates MDSC to differentiate into mature myeloid cells [198], which can improve immunotherapies, as seen with checkpoint inhibitors in melanoma patients [199] and cancer vaccines in patients with lung cancer [200,201]. Other approaches involving macrophage repolarization in neuroblastoma, including MSC-associated delivery of IFN-γ [202], inhibition of PGE2 [38], and viral delivery of CXCR4 antagonist [203], have all been shown to increase immunostimulatory M1 populations in the TME.

{kind=link}
{kind=link}
Table 2.
Treatment strategies targeting myeloid cells in the tumor microenvironment. Table indicates way of action, therapeutics, the studied myeloid population, and references to publications. Asterisks indicate studies performed in neuroblastoma models.
Table 2.
Treatment strategies targeting myeloid cells in the tumor microenvironment. Table indicates way of action, therapeutics, the studied myeloid population, and references to publications. Asterisks indicate studies performed in neuroblastoma models.
| Therapeutic Target | Point of Action | Treatment | Studied Population | Reference |
|---|---|---|---|---|
| CCR2/CCL2 | Recruitment | CCR2 inhibitor | TAM/MDSC | [168,169] |
| anti-CCL2 antibody | [170,171,172] | |||
| CSF-1/CSF-1R | Recruitment/ Repolarization | anti-CSF-1R antibody | TAM, MDSC | [94] *, [165,204,205,206] |
| CSF-1R inhibitor | TAM, MDSC | [94] *, [160] *, [166,167,206,207,208,209] | ||
| RAGE | Recruitment | Anti-RAE antibody | MDSC | [128,210] |
| Epigenetics | Repolarization | BRD inhibitor | TAM | [193], [194] * |
| Histone deacetylase inhibitor | TAM, MDSC | [195] *, [196,197] | ||
| Glutathione synthase (GSS) | Repolarization/ Survival | All-trans retinoic acid (ATRA) | MDSC | [162,198,199,200,201,211] |
| Syk-Rac2/PI3Kγ | Repolarization | Syk/PI3Kγ inhibitor | TAM, MDSC | [212,213] |
| CD40 | Repolarization | CD40 agonist | TAM | [214] *, [215] *, [216] *, [217] *, [218] |
| PGE2 | Repolarization | Prostaglandin E Synthase-1 inhibitor | TAM | [38] * |
| 67 kDa laminin receptor | Repolarization | Polyphenon E | MDSC | [11] * |
| Bisphosphonates | Repolarization/ Survival | Liposome-encapsulated bisphosphonates, Zoledronic acid | TAM | [219,220,221,222,223] |
| JAK2/STAT3 and NF-κB | Repolarization/ Survival/ Function | JAK/STAT inhibitor | TAM | [72] * |
| Sunitinib | MDSC | [181,182,183,184,224] | ||
| Curcumin | MDSC | [225,226,227,228] | ||
| Withaferin A | MDSC | [229] | ||
| Chemotherapy | Survival | 5-fluorouracil (5-FU) | MDSC | [176,177], [180] * |
| Gemcitabine | MDSC | [173,174,175,177,230] | ||
| Trabectidin (ET-743) | TAM | [178], [179] * | ||
| CD11b | Survival | anti-CD11b antibody | MDSC | [180] * |
| CD1d | Survival | Vα24-invariant NKT cells | TAM | [19] * |
| CD105 | Survival | anti-CD105 antibody | TAM | [231] * |
| CD38 | Survival | Anti-CD38 antibody | M-MDSC/Tregs | [232,233,234] |
| Bcl-2/Bcl-xL | Survival | Bcl-2 inhibitor | MDSC | [235] |
| Folate receptor β (FRβ) | Survival | anti-FRβ CAR T cells | TAM | [161] |
| CD33/CD16 | Survival | Bispecific killer cell enhancer (BiKE) | MDSC | [236] |
| Gemtuzumab | MDSC | [163] * | ||
| TRAIL-R2 | Survival | Agonist TRAIL-R2 antibody | MDSC | [237,238] |
| CD124 (IL-4Rα) | Survival | anti-IL4Rα aptamer | TAM/MDSC | [239] |
| CD47 | Survival/Function | Anti-CD47 antibody | MDSC | [240,241] |
| Arg-1 | Function | Arginase inhibitor | MDSC | [186,187] |
| IDO | Function/ Recruitment | IDO inhibitor | MDSC | [188,189] |
| COX2 (CXCR4/CXCL12) | Recruitment/ Function | COX2 inhibitor | MDSC | [7] *, [8] *, [190,191,242,243,244] |
| PDE-5 | Function | PDE-5 inhibitor | MDSC | [245,246,247,248,249,250,251] |
| Checkpoint inhibitors | Function | Anti-PD-1 antibody Anti-CTLA-4 antibody | TAM/MDSC/ Tregs | [252,253] |
| NOX2 | Function | NOX2 inhibitor | MDSC | [254] |
| VEGFR | Function | VEGFR antagonist | MDSC | [255] |
5. Conclusions
In summary, myeloid populations in the TME of neuroblastoma accrue in a combined force, suppressing host immunity and immunotherapy, which needs to be considered when developing novel treatment approaches. Increasing numbers of myeloid cells within progressed lesions and the diagnostic value of myeloid markers in neuroblastoma underline the importance that these cellular players have in tumor progression and immune inhibition. Several studies have demonstrated that immunotherapy benefits from combinatorial approaches targeting myeloid-derived suppression alongside the tumor cells, and a combined effort will have to be put into finding the most effective combinations, which may change depending on therapeutic approach and disease stage. Further investigation may be required to more clearly dissect myeloid subpopulations and transitional stages to be able to pinpoint precise targets and predominant pathways when developing future interventions for neuroblastoma.
Author Contributions
J.F. and I.L. prepared the manuscript; J.F. and J.A. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the GOSH NIHR Biomedical Research Centre and Great Ormond Street Hospital Charity for John Anderson and Research into Childhood Cancer, to Jennifer Frosch.
Data Availability Statement
Not applicable.
Acknowledgments
All figures were created with BioRender.com (accessed on 5 April 2021).
Conflicts of Interest
The authors declare no relevant conflict of interest. J.A. funded by Great Ormond Street Charity and the Great Ormond Street NIHR biomedical research centre.
Abbreviations
List of abbreviations sorted in chronological order.
| mAb | monoclonal antibodies |
| CD | Cluster of differentiation |
| TAM | tumor-associated macrophages |
| MDSC | myeloid-derived suppressor cells |
| TME | tumor microenvironment |
| APM | antigen presenting machinery |
| MHC | major histocompatibility complex |
| TGF-β | transforming growth factor-β |
| IL | interleukin |
| Arg | arginase |
| MSC | mesenchymal stem/stromal cells |
| CAF | cancer-associated fibroblasts |
| EMT | endothelial-to-mesenchymal transition |
| CXCL12 | C-X-C Motif Chemokine Ligand 12 |
| α-SMA | α-smooth muscle actin |
| VEGF | Vascular endothelial growth factor |
| STAT | signal transducer and activator of transcription |
| ERK | extracellular regulated kinase |
| CCL2 | C-C Motif Chemokine Ligand 2 |
| PGE2 | prostaglandin E 2 |
| Tregs | Regulatory T cells |
| APC | antigen-presenting cells |
| HIF1α | hypoxia inducible factor 1α |
| LPS | lipopolysaccharide |
| IFN-γ | interferon gamma |
| MMP9 | matrix metalloproteinase 9 |
| TLR | toll-like receptor |
| M-MDSC | mononuclear MDSC |
| PMN-MDSC | granulocytic or polymorphonuclear MDSC |
| eMDSC | early-stage MDSC |
| LOX1 | lectin-type oxidized low-density lipoprotein receptor 1 |
| iNOS | inducible nitric oxide synthase |
| PDGF | Platelet-derived growth factor |
| IDO | indoleamine 2,3-dioxygenase |
| MET | mesenchymal-to-epithelial transition |
| SDF1 | stromal cell-derived factor 1 |
| HSC | hematopoietic stem cells |
| CMP | common myeloid progenitor cells |
| G-CSF | granulocyte colony-stimulating factor |
| GM-CSF | granulocyte-macrophage colony-stimulating factor |
| IRF8 | Interferon regulatory factor 8 |
| RAGE | receptor for advanced glycation end products |
| TCR | T cell receptor |
| ROS | reactive oxygen species |
| RNS | reactive nitrogen species |
| NO | nitric oxide |
| PNT | peroxynitrite |
| HDAC | histone deacetylase |
| ATRA | all-trans retinoic acid |
References
- Whittle, S.B.; Smith, V.; Doherty, E.; Zhao, S.; Mccarty, S.; Zage, P.E. Overview and recent advances in the treatment of neuroblastoma. Expert Rev. Anticancer. Ther. 2017, 17, 369–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, W.A.; June, C.H. The Principles of Engineering Immune Cells to Treat Cancer. Cell 2017, 168, 724–740. [Google Scholar] [CrossRef] [Green Version]
- Pule, M.A.; Savoldo, B.; Myers, G.D.; Rossig, C.; Russell, H.V.; Dotti, G.; Huls, M.H.; Liu, E.; Gee, A.P.; Mei, Z.; et al. Virus-specific T cells engineered to coexpress tumor-specific receptors: Persistence and antitumor activity in individuals with neuroblastoma. Nat. Med. 2008, 14, 1264–1270. [Google Scholar] [CrossRef]
- Louis, C.U.; Savoldo, B.; Dotti, G.; Pule, M.; Yvon, E.; Myers, G.D.; Rossig, C.; Russell, H.V.; Diouf, O.; Liu, E.; et al. Antitumor activity and long-term fate of chimeric antigen receptor–positive T cells in patients with neuroblastoma. Blood 2011, 118, 6050–6056. [Google Scholar] [CrossRef]
- Heczey, A.; Louis, C.U.; Savoldo, B.; Dakhova, O.; Durett, A.; Grilley, B.; Liu, H.; Wu, M.F.; Mei, Z.; Gee, A.; et al. CAR T Cells Administered in Combination with Lymphodepletion and PD-1 Inhibition to Patients with Neuroblastoma. Mol. Ther. 2017, 25, 2214–2224. [Google Scholar] [CrossRef] [Green Version]
- Straathof, K.; Flutter, B.; Wallace, R.; Jain, N.; Loka, T.; Depani, S.; Wright, G.; Thomas, S.; Cheung, G.W.-K.; Gileadi, T.; et al. Antitumor activity without on-target off-tumor toxicity of GD2–chimeric antigen receptor T cells in patients with neuroblastoma. Sci. Transl. Med. 2020, 12, eabd6169. [Google Scholar] [CrossRef]
- Carlson, L.-M.; Rasmuson, A.; Idborg, H.; Segerström, L.; Jakobsson, P.-J.; Sveinbjörnsson, B.; Kogner, P. Low-dose aspirin delays an inflammatory tumor progression in vivo in a transgenic mouse model of neuroblastoma. Carcinogenesis 2013, 34, 1081–1088. [Google Scholar] [CrossRef]
- Carlson, L.-M.; Kogner, P. Neuroblastoma-related inflammation. Oncoimmunology 2013, 2, e24658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apps, J.R.; Hasan, F.; Campus, O.; Behjati, S.; Jacques, T.S.; Sebire, N.J.; Anderson, J. The immune environment of paediatric solid malignancies: Evidence from an immunohistochemical study of clinical cases. Fetal Pediatr. Pathol. 2013, 32, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Asgharzadeh, S.; Salo, J.A.; Ji, L.; Oberthuer, A.; Fischer, M.; Berthold, F.; Hadjidaniel, M.; Liu, C.W.-Y.; Metelitsa, L.S.; Pique-Regi, R.; et al. Clinical Significance of Tumor-Associated Inflammatory Cells in Metastatic Neuroblastoma. J. Clin. Oncol. 2012, 30, 3525–3532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santilli, G.; Piotrowska, I.; Cantilena, S.; Chayka, O.; D’Alicarnasso, M.; Morgenstern, D.A.; Himoudi, N.; Pearson, K.; Anderson, J.; Thrasher, A.J.; et al. Polyphenol E Enhances the Antitumor Immune Response in Neuroblastoma by Inactivating Myeloid Suppressor Cells. Clin. Cancer Res. 2013, 19, 1116–1125. [Google Scholar] [CrossRef] [Green Version]
- Santilli, G.; Anderson, J.; Thrasher, A.J.; Sala, A. Catechins and antitumor immunity. Oncoimmunology 2013, 2, e24443. [Google Scholar] [CrossRef] [Green Version]
- Epistoia, V.; Emorandi, F.; Ebianchi, G.; Epezzolo, A.; Eprigione, I.; Eraffaghello, L. Immunosuppressive Microenvironment in Neuroblastoma. Front. Oncol. 2013, 3, 167. [Google Scholar] [CrossRef] [Green Version]
- Borriello, L.; Seeger, R.C.; Asgharzadeh, S.; de Clerck, Y.A. More than the genes, the tumor microenvironment in neuroblastoma. Cancer Lett. 2016, 380, 304–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanichapol, T.; Chutipongtanate, S.; Anurathapan, U.; Hongeng, S. Immune Escape Mechanisms and Future Prospects for Immunotherapy in Neuroblastoma. BioMed Res. Int. 2018, 2018, 1812535. [Google Scholar] [CrossRef] [PubMed]
- Prigione, I.; Corrias, M.V.; Airoldi, I.; Raffaghello, L.; Morandi, F.; Bocca, P.; Cocco, C.; Ferrone, S.; Pistoia, V. Immunogenicity of Human Neuroblastoma. Ann. N. Y. Acad. Sci. 2004, 1028, 69–80. [Google Scholar] [CrossRef]
- Jungbluth, A.A.; Garrido, F.; Cabrera, T.; Meyen-Southard, S.; Ernestus, K.; Berthold, F. Expression of MHC class I, MHC class II, and cancer germline antigens in neuroblastoma. Cancer Immunol. Immunother. 2005, 54, 400–406. [Google Scholar] [CrossRef]
- Corrias, M.; Occhino, M.; Croce, M.; de Ambrosis, A.; Pistillo, M.; Bocca, P.; Pistoia, V.; Ferrini, S. Lack of HLA-class I antigens in human neuroblastoma cells: Analysis of its relationship to TAP and tapasin expression. Tissue Antigens 2001, 57, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Asgharzadeh, S.; Salo, J.; Engell, K.; Wu, H.-W.; Sposto, R.; Ara, T.; Silverman, A.M.; Declerck, Y.A.; Seeger, R.C.; et al. Vα24-invariant NKT cells mediate antitumor activity via killing of tumor-associated macrophages. J. Clin. Investig. 2009, 119, 1524–1536. [Google Scholar] [CrossRef] [Green Version]
- Asgharzadeh, S.; Pique-Regi, R.; Sposto, R.; Wang, H.; Yang, Y.; Shimada, H.; Matthay, K.; Buckley, J.; Ortega, A.; Seeger, R.C. Prognostic Significance of Gene Expression Profiles of Metastatic Neuroblastomas Lacking MYCN Gene Amplification. J. Natl. Cancer Inst. 2006, 98, 1193–1203. [Google Scholar] [CrossRef]
- Soldati, R.; Berger, E.; Zenclussen, A.C.; Jorch, G.; Lode, H.N.; Salatino, M.; Rabinovich, G.A.; Fest, S. Neuroblastoma triggers an immunoevasive program involving galectin-1-dependent modulation of T cell and dendritic cell compartments. Int. J. Cancer 2011, 131, 1131–1141. [Google Scholar] [CrossRef]
- Brandetti, E.; Veneziani, I.; Melaiu, O.; Pezzolo, A.; Castellano, A.; Boldrini, R.; Ferretti, E.; Fruci, D.; Moretta, L.; Pistoia, V.; et al. MYCN is an immunosuppressive oncogene dampening the expression of ligands for NK-cell-activating receptors in human high-risk neuroblastoma. Oncoimmunology 2017, 6, e1316439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raffaghello, L.; Prigione, I.; Airoldi, I.; Camoriano, M.; Morandi, F.; Bocca, P.; Gambini, C.; Ferrone, S.; Pistoia, V. Mechanisms of immune evasion of human neuroblastoma. Cancer Lett. 2005, 228, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Raffaghello, L.; Prigione, I.; Airoldi, I.; Camoriano, M.; Levreri, I.; Gambini, C.; Pende, D.; Steinle, A.; Ferrone, S.; Pistoia, V. Downregulation and/or Release of NKG2D Ligands as Immune Evasion Strategy of Human Neuroblastoma. Neoplasia 2004, 6, 558–568. [Google Scholar] [CrossRef] [Green Version]
- Mussai, F.; Egan, S.A.; Hunter, S.; Webber, H.; Fisher, J.; Wheat, R.; McConville, C.; Sbirkov, Y.; Wheeler, K.A.; Bendle, G.; et al. Neuroblastoma Arginase Activity Creates an Immunosuppressive Microenvironment That Impairs Autologous and Engineered Immunity. Cancer Res. 2015, 75, 3043–3053. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, P.C.; Quiceno, D.G.; Ochoa, A.C. l-arginine availability regulates T-lymphocyte cell-cycle progression. Blood 2006, 109, 1568–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelizzo, G.; Veschi, V.; Mantelli, M.; Croce, S.; Di Benedetto, V.; D’Angelo, P.; Maltese, A.; Catenacci, L.; Apuzzo, T.; Scavo, E.; et al. Microenvironment in neuroblastoma: Isolation and characterization of tumor-derived mesenchymal stromal cells. BMC Cancer 2018, 18, 1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiura, Y.; Shimada, H.; Seeger, R.C.; Laug, W.E.; Declerck, Y.A. Matrix metalloproteinases-2 and -9 are expressed in human neuroblastoma: Contribution of stromal cells to their production and correlation with metastasis. Cancer Res. 1998, 58, 2209–2216. [Google Scholar]
- Ribatti, M.; Marimpietri, D.; Pastorino, F.; Brignole, C.; Nico, B.; Vacca, A.; Ponzoni, M. Angiogenesis in Neuroblastoma. Ann. N. Y. Acad. Sci. 2004, 1028, 133–142. [Google Scholar] [CrossRef]
- Lanza, C.; Morando, S.; Voci, A.; Canesi, L.; Principato, M.C.; Serpero, L.D.; Mancardi, G.; Uccelli, A.; Vergani, L. Neuroprotective mesenchymal stem cells are endowed with a potent antioxidant effectin vivo. J. Neurochem. 2009, 110, 1674–1684. [Google Scholar] [CrossRef]
- Zeine, R.; Salwen, H.R.; Peddinti, R.; Tian, Y.; Guerrero, L.; Yang, Q.; Chlenski, A.; Cohn, S.L. Presence of cancer-associated fibroblasts inversely correlates with Schwannian stroma in neuroblastoma tumors. Mod. Pathol. 2009, 22, 950–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borriello, L.; Nakata, R.; Sheard, M.A.; Fernandez, G.E.; Sposto, R.; Malvar, J.; Blavier, L.; Shimada, H.; Asgharzadeh, S.; Seeger, R.C.; et al. Cancer-Associated Fibroblasts Share Characteristics and Protumorigenic Activity with Mesenchymal Stromal Cells. Cancer Res. 2017, 77, 5142–5157. [Google Scholar] [CrossRef] [Green Version]
- Blavier, L.; Yang, R.-M.; Declerck, Y.A. The Tumor Microenvironment in Neuroblastoma: New Players, New Mechanisms of Interaction and New Perspectives. Cancers 2020, 12, 2912. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R. The biology and function of fibroblasts in cancer. Nat. Rev. Cancer 2016, 16, 582–598. [Google Scholar] [CrossRef] [PubMed]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; DeNardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 2020, 20, 174–186. [Google Scholar] [CrossRef] [Green Version]
- Ara, T.; Nakata, R.; Sheard, M.A.; Shimada, H.; Buettner, R.; Groshen, S.G.; Ji, L.; Yu, H.; Jove, R.; Seeger, R.C.; et al. Critical Role of STAT3 in IL-6–Mediated Drug Resistance in Human Neuroblastoma. Cancer Res. 2013, 73, 3852–3864. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, O.; Yoshida, M.; Koma, Y.; Yanai, T.; Hasegawa, D.; Kosaka, Y.; Nishimura, N.; Yokozaki, H. Collaboration of cancer-associated fibroblasts and tumour-associated macrophages for neuroblastoma development. J. Pathol. 2016, 240, 211–223. [Google Scholar] [CrossRef]
- Kock, A.; Larsson, K.; Bergqvist, F.; Eissler, N.; Elfman, L.H.; Raouf, J.; Korotkova, M.; Johnsen, J.I.; Jakobsson, P.-J.; Kogner, P. Inhibition of Microsomal Prostaglandin E Synthase-1 in Cancer-Associated Fibroblasts Suppresses Neuroblastoma Tumor Growth. EBioMedicine 2018, 32, 84–92. [Google Scholar] [CrossRef]
- Silzle, T.; Kreutz, M.; Dobler, M.A.; Brockhoff, G.; Knuechel, R.; Kunz-Schughart, L.A. Tumor-associated fibroblasts recruit blood monocytes into tumor tissue. Eur. J. Immunol. 2003, 33, 1311–1320. [Google Scholar] [CrossRef]
- Johnson, B.D.; Jing, W.; Orentas, R.J. CD25+ Regulatory T Cell Inhibition Enhances Vaccine-induced Immunity to Neuroblastoma. J. Immunother. 2007, 30, 203–214. [Google Scholar] [CrossRef]
- Rigo, V.; Corrias, M.V.; Orengo, A.M.; Brizzolara, A.; Emionite, L.; Fenoglio, D.; Filaci, G.; Croce, M.; Ferrini, S. Recombinant IL-21 and anti-CD4 antibodies cooperate in syngeneic neuroblastoma immunotherapy and mediate long-lasting immunity. Cancer Immunol. Immunother. 2014, 63, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Zou, W. Regulatory T cells, tumour immunity and immunotherapy. Nat. Rev. Immunol. 2006, 6, 295–307. [Google Scholar] [CrossRef]
- Ohue, Y.; Nishikawa, H. Regulatory T (Treg) cells in cancer: Can Treg cells be a new therapeutic target? Cancer Sci. 2019, 110, 2080–2089. [Google Scholar] [CrossRef] [PubMed]
- Kerkar, S.P.; Restifo, N.P. Cellular Constituents of Immune Escape within the Tumor Microenvironment: Figure 1. Cancer Res. 2012, 72, 3125–3130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, S.; Ahlers, J.D.; Vortmeyer, A.O.; Terabe, M.; Tsukui, T.; Carbone, D.P.; Liotta, L.A.; Berzofsky, J.A. A model for CD8+ CTL tumor immunosurveillance and regulation of tumor escape by CD4 T cells through an effect on quality of CTL. J. Immunol. 1999, 163, 184–193. [Google Scholar] [PubMed]
- Gattinoni, L.; Finkelstein, S.E.; Klebanoff, C.A.; Antony, P.A.; Palmer, D.C.; Spiess, P.J.; Hwang, L.N.; Yu, Z.; Wrzesinski, C.; Heimann, D.M.; et al. Removal of homeostatic cytokine sinks by lymphodepletion enhances the efficacy of adoptively transferred tumor-specific CD8+ T cells. J. Exp. Med. 2005, 202, 907–912. [Google Scholar] [CrossRef]
- Kumar, V.; Patel, S.; Tcyganov, E.; Gabrilovich, D.I. The Nature of Myeloid-Derived Suppressor Cells in the Tumor Microenvironment. Trends Immunol. 2016, 37, 208–220. [Google Scholar] [CrossRef] [Green Version]
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-derived suppressor cells as regulators of the immune system. Nat. Rev. Immunol. 2009, 9, 162–174. [Google Scholar] [CrossRef]
- Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Bronte, V. Coordinated regulation of myeloid cells by tumours. Nat. Rev. Immunol. 2012, 12, 253–268. [Google Scholar] [CrossRef] [Green Version]
- Gabrilovich, D.I. Mechanisms and functional significance of tumour-induced dendritic-cell defects. Nat. Rev. Immunol. 2004, 4, 941–952. [Google Scholar] [CrossRef]
- Corzo, C.A.; Condamine, T.; Lu, L.; Cotter, M.J.; Youn, J.-I.; Cheng, P.; Cho, H.-I.; Celis, E.; Quiceno, D.G.; Padhya, T.; et al. HIF-1α regulates function and differentiation of myeloid-derived suppressor cells in the tumor microenvironment. J. Exp. Med. 2010, 207, 2439–2453. [Google Scholar] [CrossRef] [PubMed]
- Bergenfelz, C.; Leandersson, K. The Generation and Identity of Human Myeloid-Derived Suppressor Cells. Front. Oncol. 2020, 10, 109. [Google Scholar] [CrossRef] [Green Version]
- Bronte, V.; Brandau, S.; Chen, S.-H.; Colombo, M.P.; Frey, A.B.; Greten, T.F.; Mandruzzato, S.; Murray, P.J.; Ochoa, A.; Ostrand-Rosenberg, S.; et al. Recommendations for myeloid-derived suppressor cell nomenclature and characterization standards. Nat. Commun. 2016, 7, 12150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medrek, C.; Pontén, F.; Jirström, K.; Leandersson, K. The presence of tumor associated macrophages in tumor stroma as a prognostic marker for breast cancer patients. BMC Cancer 2012, 12, 306. [Google Scholar] [CrossRef] [PubMed]
- Ni, C.; Yang, L.; Xu, Q.; Yuan, H.; Wang, W.; Xia, W.; Gong, D.; Zhang, W.; Yu, K. CD68- and CD163-positive tumor infiltrating macrophages in non-metastatic breast cancer: A retrospective study and meta-analysis. J. Cancer 2019, 10, 4463–4472. [Google Scholar] [CrossRef] [Green Version]
- Saylor, J.; Ma, Z.; Goodridge, H.S.; Huang, F.; Cress, A.E.; Pandol, S.J.; Shiao, S.L.; Vidal, A.C.; Wu, L.; Nickols, N.G.; et al. Spatial Mapping of Myeloid Cells and Macrophages by Multiplexed Tissue Staining. Front. Immunol. 2018, 9, 2925. [Google Scholar] [CrossRef] [Green Version]
- Sulahian, T.H.; Högger, P.; Wahner, A.E.; Wardwell, K.; Goulding, N.J.; Sorg, C.; Droste, A.; Stehling, M.; Wallace, P.K.; Morganelli, P.M.; et al. Human monocytes express cd163, which is upregulated by il-10 and identical to p155. Cytokine 2000, 12, 1312–1321. [Google Scholar] [CrossRef]
- Triantafyllou, E.; Woollard, K.J.; McPhail, M.J.W.; Antoniades, C.G.; Possamai, L.A. The Role of Monocytes and Macrophages in Acute and Acute-on-Chronic Liver Failure. Front. Immunol. 2018, 9, 2948. [Google Scholar] [CrossRef]
- Yang, J.; Anholts, J.; Kolbe, U.; Stegehuis-Kamp, J.A.; Claas, F.H.J.; Eikmans, M. Calcium-Binding Proteins S100A8 and S100A9: Investigation of Their Immune Regulatory Effect in Myeloid Cells. Int. J. Mol. Sci. 2018, 19, 1833. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhang, X. Pivotal regulators of tissue homeostasis and cancer: Macrophages. Exp. Hematol. Oncol. 2017, 6, 1–8. [Google Scholar] [CrossRef]
- Gordon, S.; Martinez, F.O. Alternative Activation of Macrophages: Mechanism and Functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.X.; Joshi, S. “Re-educating” Tumor Associated Macrophages as a Novel Immunotherapy Strategy for Neuroblastoma. Front. Immunol. 2020, 11, 1947. [Google Scholar] [CrossRef]
- Martinez, F.O.; Gordon, S.; Locati, M.; Mantovani, A. Transcriptional Profiling of the Human Monocyte-to-Macrophage Differentiation and Polarization: New Molecules and Patterns of Gene Expression. J. Immunol. 2006, 177, 7303–7311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS–) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef] [PubMed]
- Rath, M.; Müler, I.; Kropf, P.; Closs, E.I.; Munder, M. Metabolism via Arginase or Nitric Oxide Synthase: Two Competing Arginine Pathways in Macrophages. Front. Immunol. 2014, 5, 532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viola, A.; Munari, F.; Sánchez-Rodríguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, M.; Wen, H.; Corsa, C.A.S.; Liu, T.; Coelho, A.L.; Allen, R.M.; Carson, I.W.F.; Cavassani, K.A.; Li, X.; Lukacs, N.W.; et al. Epigenetic regulation of the alternatively activated macrophage phenotype. Blood 2009, 114, 3244–3254. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, S.; Wu, H.; Rong, X.; Guo, J. M2b macrophage polarization and its roles in diseases. J. Leukoc. Biol. 2019, 106, 345–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, Y. Tumor-associated macrophages: From basic research to clinical application. J. Hematol. Oncol. 2017, 10, 58. [Google Scholar] [CrossRef] [Green Version]
- Hadjidaniel, M.D.; Muthugounder, S.; Hung, L.T.; Sheard, M.A.; Shirinbak, S.; Chan, R.Y.; Nakata, R.; Borriello, L.; Malvar, J.; Kennedy, R.J.; et al. Tumor-associated macrophages promote neuroblastoma via STAT3 phosphorylation and up-regulation of c-MYC. Oncotarget 2017, 8, 91516–91529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanmee, T.; Ontong, P.; Konno, K.; Itano, N.; Chanmee, T. Tumor-Associated Macrophages as Major Players in the Tumor Microenvironment. Cancers (Basel) 2014, 6, 1670–1690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laoui, D.; van Overmeire, E.; Movahedi, K.; Bossche, J.V.D.; Schouppe, E.; Mommer, C.; Nikolaou, A.; Morias, Y.; De Baetselier, P.; van Ginderachter, J.A. Mononuclear phagocyte heterogeneity in cancer: Different subsets and activation states reaching out at the tumor site. Immunobiology 2011, 216, 1192–1202. [Google Scholar] [CrossRef]
- Schouppe, E.; De Baetselier, P.; van Ginderachter, J.A.; Sarukhan, A. Instruction of myeloid cells by the tumor microenvironment: Open questions on the dynamics and plasticity of different tumor-associated myeloid cell populations. OncoImmunology 2012, 1, 1135–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laoui, D.; van Overmeire, E.; Di Conza, G.; Aldeni, C.; Keirsse, J.; Morias, Y.; Movahedi, K.; Houbracken, I.; Schouppe, E.; Elkrim, Y.; et al. Tumor Hypoxia Does Not Drive Differentiation of Tumor-Associated Macrophages but Rather Fine-Tunes the M2-like Macrophage Population. Cancer Res. 2014, 74, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Gabrilovich, D.I.; Bronte, V.; Chen, S.-H.; Colombo, M.P.; Ochoa, A.; Ostrand-Rosenberg, S.; Schreiber, H. The Terminology Issue for Myeloid-Derived Suppressor Cells. Cancer Res. 2007, 67, 425. [Google Scholar] [CrossRef] [Green Version]
- Ostrand-Rosenberg, S. Myeloid-Derived Suppressor Cells: Facilitators of Cancer and Obesity-Induced Cancer. Annu. Rev. Cancer Biol. 2021, 5, 17–38. [Google Scholar] [CrossRef]
- Highfill, S.L.; Cui, Y.; Giles, A.J.; Smith, J.P.; Zhang, H.; Morse, E.; Kaplan, R.N.; Mackall, C.L. Disruption of CXCR2-Mediated MDSC Tumor Trafficking Enhances Anti-PD1 Efficacy. Sci. Transl. Med. 2014, 6, 237ra67. [Google Scholar] [CrossRef]
- Köstlin, N.; Kugel, H.; Spring, B.; Leiber, A.; Marmé, A.; Henes, M.; Rieber, N.; Hartl, D.; Poets, C.F.; Gille, C. Granulocytic myeloid derived suppressor cells expand in human pregnancy and modulate T-cell responses. Eur. J. Immunol. 2014, 44, 2582–2591. [Google Scholar] [CrossRef]
- Ostrand-Rosenberg, S.; Sinha, P.; Figley, C.; Long, R.; Park, D.; Carter, D.; Clements, V.K. Frontline Science: Myeloid-derived suppressor cells (MDSCs) facilitate maternal–fetal tolerance in mice. J. Leukoc. Biol. 2017, 101, 1091–1101. [Google Scholar] [CrossRef]
- Verschoor, C.P.; Johnstone, J.; Millar, J.; Dorrington, M.G.; Habibagahi, M.; Lelic, A.; Loeb, M.; Bramson, J.L.; Bowdish, D.M.E. Blood CD33(+)HLA-DR(−) myeloid-derived suppressor cells are increased with age and a history of cancer. J. Leukoc. Biol. 2013, 93, 633–637. [Google Scholar] [CrossRef]
- Ostrand-Rosenberg, S.; Sinha, P. Myeloid-Derived Suppressor Cells: Linking Inflammation and Cancer. J. Immunol. 2009, 182, 4499–4506. [Google Scholar] [CrossRef] [PubMed]
- Movahedi, K.; Guilliams, M.; Bossche, J.V.D.; Bergh, R.V.D.; Gysemans, C.; Beschin, A.; De Baetselier, P.; van Ginderachter, J.A. Identification of discrete tumor-induced myeloid-derived suppressor cell subpopulations with distinct T cell–suppressive activity. Blood 2008, 111, 4233–4244. [Google Scholar] [CrossRef] [PubMed]
- Youn, J.-I.; Nagaraj, S.; Collazo, M.; Gabrilovich, D.I. Subsets of Myeloid-Derived Suppressor Cells in Tumor-Bearing Mice. J. Immunol. 2008, 181, 5791–5802. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.; Bruderek, K.; Kaspar, C.; Höing, B.; Kanaan, O.; Dominas, N.; Hussain, T.; Droege, F.; Eyth, C.; Hadaschik, B.; et al. Clinical Relevance and Suppressive Capacity of Human Myeloid-Derived Suppressor Cell Subsets. Clin. Cancer Res. 2018, 24, 4834–4844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassetta, L.; Baekkevold, E.S.; Brandau, S.; Bujko, A.; Cassatella, M.A.; Dorhoi, A.; Krieg, C.; Lin, A.; Loré, K.; Marini, O.; et al. Deciphering myeloid-derived suppressor cells: Isolation and markers in humans, mice and non-human primates. Cancer Immunol. Immunotherapy 2019, 68, 687–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fridlender, Z.G.; Sun, J.; Kim, S.; Kapoor, V.; Cheng, G.; Ling, L.; Worthen, G.S.; Albelda, S.M. Polarization of Tumor-Associated Neutrophil Phenotype by TGF-β: “N1” versus “N2” TAN. Cancer Cell 2009, 16, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Sagiv, J.Y.; Michaeli, J.; Assi, S.; Mishalian, I.; Kisos, H.; Levy, L.; Damti, P.; Lumbroso, D.; Polyansky, L.; Sionov, R.V.; et al. Phenotypic diversity and plasticity in circulating neutrophil subpopulations in cancer. Cell Rep. 2015, 10, 562–573. [Google Scholar] [CrossRef] [Green Version]
- Dumitru, C.A.; Moses, K.; Trellakis, S.; Lang, S.; Brandau, S. Neutrophils and granulocytic myeloid-derived suppressor cells: Immunophenotyping, cell biology and clinical relevance in human oncology. Cancer Immunol. Immunotherapy 2012, 61, 1155–1167. [Google Scholar] [CrossRef]
- Condamine, T.; Dominguez, G.A.; Youn, J.-I.; Kossenkov, A.V.; Mony, S.; Alicea-Torres, K.; Tcyganov, E.; Hashimoto, A.; Nefedova, Y.; Lin, C.; et al. Lectin-type oxidized LDL receptor-1 distinguishes population of human polymorphonuclear myeloid-derived suppressor cells in cancer patients. Sci. Immunol. 2016, 1, aaf8943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alshetaiwi, H.; Pervolarakis, N.; McIntyre, L.L.; Ma, D.; Nguyen, Q.; Rath, J.A.; Nee, K.; Hernandez, G.; Evans, K.; Torosian, L.; et al. Defining the emergence of myeloid-derived suppressor cells in breast cancer using single-cell transcriptomics. Sci. Immunol. 2020, 5, eaay6017. [Google Scholar] [CrossRef] [PubMed]
- Abraham, D.; Zins, K.; Sioud, M.; Lucas, T.; Schäfer, R.; Stanley, E.R.; Aharinejad, S. Stromal cell-derived CSF-1 blockade prolongs xenograft survival of CSF-1-negative neuroblastoma. Int. J. Cancer 2009, 126, 1339–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, M.W.; Sun, J.; Sheard, M.A.; Liu, W.; Wu, H.; Jackson, J.R.; Malvar, J.; Sposto, R.; Daniel, D.; Seeger, R.C. Colony stimulating factor 1 receptor blockade improves the efficacy of chemotherapy against human neuroblastoma in the absence of T lymphocytes. Int. J. Cancer 2018, 143, 1483–1493. [Google Scholar] [CrossRef] [PubMed]
- Orimo, A.; Gupta, P.B.; Sgroi, D.C.; Arenzana-Seisdedos, F.; Delaunay, T.; Naeem, R.; Carey, V.J.; Richardson, A.L.; Weinberg, R.A. Stromal Fibroblasts Present in Invasive Human Breast Carcinomas Promote Tumor Growth and Angiogenesis through Elevated SDF-1/CXCL12 Secretion. Cell 2005, 121, 335–348. [Google Scholar] [CrossRef]
- Kumar, V.; Gabrilovich, D.I. Hypoxia-inducible factors in regulation of immune responses in tumour microenvironment. Immunology 2014, 143, 512–519. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D.; Vacca, A.; Nico, B.; de Falco, G.; Montaldo, P.G.; Ponzoni, M. Angiogenesis and anti-angiogenesis in neuroblastoma. Eur. J. Cancer 2002, 38, 750–757. [Google Scholar] [CrossRef]
- Wyckoff, J.B.; Wang, Y.; Lin, E.Y.; Li, J.-F.; Goswami, S.; Stanley, E.R.; Segall, J.E.; Pollard, J.W.; Condeelis, J. Direct Visualization of Macrophage-Assisted Tumor Cell Intravasation in Mammary Tumors. Cancer Res. 2007, 67, 2649–2656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietras, A.; Gisselsson, D.; Øra, I.; Noguera, R.; Beckman, S.; Navarro, S.; Påhlman, S. High levels of HIF-2α highlight an immature neural crest-like neuroblastoma cell cohort located in a perivascular niche. J. Pathol. 2007, 214, 482–488. [Google Scholar] [CrossRef]
- Löfstedt, T.; Fredlund, E.; Holmquist-Mengelbier, L.; Pietras, A.; Ovenberger, M.; Poellinger, L.; Påhlman, S. Hypoxia Inducible Factor-2α in Cancer. Cell Cycle 2007, 6, 919–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Sun, J.; Sheard, M.A.; Tran, H.C.; Wan, Z.; Liu, W.Y.; Asgharzadeh, S.; Sposto, R.; Wu, H.W.; Seeger, R.C. Lenalidomide overcomes suppression of human natural killer cell anti-tumor functions by neuroblastoma microenvironment-associated IL-6 and TGFβ1. Cancer Immunol. Immunotherapy 2013, 62, 1637–1648. [Google Scholar] [CrossRef] [Green Version]
- Peranzoni, E.; Lemoine, J.; Vimeux, L.; Feuillet, V.; Barrin, S.; Kantari-Mimoun, C.; Bercovici, N.; Guérin, M.; Biton, J.; Ouakrim, H.; et al. Macrophages impede CD8 T cells from reaching tumor cells and limit the efficacy of anti–PD-1 treatment. Proc. Natl. Acad. Sci. USA 2018, 115, E4041–E4050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noman, M.Z.; de Santis, G.; Janji, B.; Hasmim, M.; Karray, S.; Dessen, P.; Bronte, V.; Chouaib, S. PD-L1 is a novel direct target of HIF-1α, and its blockade under hypoxia enhanced MDSC-mediated T cell activation. J. Exp. Med. 2014, 211, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Durden, D.L. Combinatorial Approach to Improve Cancer Immunotherapy: Rational Drug Design Strategy to Simultaneously Hit Multiple Targets to Kill Tumor Cells and to Activate the Immune System. J. Oncol. 2019, 2019, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A.; Marchesi, F.; Malesci, A.; Laghi, L.; Allavena, P. Tumour-associated macrophages as treatment targets in oncology. Nat. Rev. Clin. Oncol. 2017, 14, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K.; Mantovani, A. Macrophage plasticity and interaction with lymphocyte subsets: Cancer as a paradigm. Nat. Immunol. 2010, 11, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Comito, G.; Giannoni, E.; Segura, C.P.; Barcellos-de-Souza, P.; Raspollini, M.R.; Baroni, G.; Lanciotti, M.; Serni, S.; Chiarugi, P. Cancer-associated fibroblasts and M2-polarized macrophages synergize during prostate carcinoma progression. Oncogene 2013, 33, 2423–2431. [Google Scholar] [CrossRef] [Green Version]
- Komohara, Y.; Takeya, M. CAFs and TAMs: Maestros of the tumour microenvironment. J. Pathol. 2016, 241, 313–315. [Google Scholar] [CrossRef]
- Malekghasemi, S.; Majidi, J.; Baghbanzadeh, A.; Abdolalizadeh, J.; Baradaran, B.; Aghebati-Maleki, L. Tumor-Associated Macrophages: Protumoral Macrophages in Inflammatory Tumor Microenvironment. Adv. Pharm. Bull. 2020, 10, 556–565. [Google Scholar] [CrossRef]
- Bianchi, G.; Vuerich, M.; Pellegatti, P.; Marimpietri, D.; Emionite, L.; Marigo, I.; Bronte, V.; Di Virgilio, F.; Pistoia, V.; Raffaghello, L. ATP/P2X7 axis modulates myeloid-derived suppressor cell functions in neuroblastoma microenvironment. Cell Death Dis. 2014, 5, e1135. [Google Scholar] [CrossRef] [Green Version]
- Stewart, T.J.; Liewehr, D.J.; Steinberg, S.M.; Greeneltch, K.M.; Abrams, S.I. Modulating the Expression of IFN Regulatory Factor 8 Alters the Protumorigenic Behavior of CD11b+Gr-1+Myeloid Cells. J. Immunol. 2009, 183, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Waight, J.D.; Netherby, C.; Hensen, M.L.; Miller, A.; Hu, Q.; Liu, S.; Bogner, P.N.; Farren, M.R.; Lee, K.P.; Liu, K.; et al. Myeloid-derived suppressor cell development is regulated by a STAT/IRF-8 axis. J. Clin. Investig. 2013, 123, 4464–4478. [Google Scholar] [CrossRef]
- Netherby, C.S.; Abrams, S.I. Mechanisms overseeing myeloid-derived suppressor cell production in neoplastic disease. Cancer Immunol. Immunother. 2017, 66, 989–996. [Google Scholar] [CrossRef]
- Bunt, S.K.; Sinha, P.; Clements, V.K.; Leips, J.; Ostrand-Rosenberg, S. Inflammation Induces Myeloid-Derived Suppressor Cells that Facilitate Tumor Progression. J. Immunol. 2005, 176, 284–290. [Google Scholar] [CrossRef] [Green Version]
- Bunt, S.K.; Yang, L.; Sinha, P.; Clements, V.K.; Leips, J.; Ostrand-Rosenberg, S. Reduced Inflammation in the Tumor Microenvironment Delays the Accumulation of Myeloid-Derived Suppressor Cells and Limits Tumor Progression. Cancer Res. 2007, 67, 10019–10026. [Google Scholar] [CrossRef] [Green Version]
- Sinha, P.; Clements, V.K.; Fulton, A.M.; Ostrand-Rosenberg, S. Prostaglandin E2 Promotes Tumor Progression by Inducing Myeloid-Derived Suppressor Cells. Cancer Res. 2007, 67, 4507–4513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Y.; Sarhan, D.; Steven, A.; Seliger, B.; Kiessling, R.; Lundqvist, A. Inhibition of Tumor-Derived Prostaglandin-E2 Blocks the Induction of Myeloid-Derived Suppressor Cells and Recovers Natural Killer Cell Activity. Clin. Cancer Res. 2014, 20, 4096–4106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Krelin, Y.; Dvorkin, T.; Bjorkdahl, O.; Segal, S.; Dinarello, C.A.; Voronov, E.; Apte, R.N. CD11b+/Gr-1+Immature Myeloid Cells Mediate Suppression of T Cells in Mice Bearing Tumors of IL-1β-Secreting Cells. J. Immunol. 2005, 175, 8200–8208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nefedova, Y.; Nagaraj, S.; Rosenbauer, A.; Murocacho, C.A.; Sebti, S.M.; Gabrilovich, D.I. Regulation of Dendritic Cell Differentiation and Antitumor Immune Response in Cancer by Pharmacologic-Selective Inhibition of the Janus-Activated Kinase 2/Signal Transducers and Activators of Transcription 3 Pathway. Cancer Res. 2005, 65, 9525–9535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corzo, C.A.; Cotter, M.J.; Cheng, P.; Cheng, F.; Kusmartsev, S.; Sotomayor, E.; Padhya, T.; McCaffrey, T.V.; McCaffrey, J.C.; Gabrilovich, D.I. Mechanism Regulating Reactive Oxygen Species in Tumor-Induced Myeloid-Derived Suppressor Cells. J. Immunol. 2009, 182, 5693–5701. [Google Scholar] [CrossRef]
- Waight, J.D.; Hu, Q.; Miller, A.; Liu, S.; Abrams, S.I. Tumor-Derived G-CSF Facilitates Neoplastic Growth through a Granulocytic Myeloid-Derived Suppressor Cell-Dependent Mechanism. PLoS ONE 2011, 6, e27690. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, T.; Ebihara, S.; Asada, M.; Kanda, A.; Sasaki, H.; Yamaya, M. Granulocyte colony-stimulating factor promotes tumor angiogenesis via increasing circulating endothelial progenitor cells and Gr1+CD11b+ cells in cancer animal models. Int. Immunol. 2005, 18, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bronte, V.; Chappell, D.B.; Apolloni, E.; Cabrelle, A.; Wang, M.; Hwu, P.; Restifo, N.P. Unopposed production of granulocyte-macrophage colony-stimulating factor by tumors inhibits CD8+ T cell responses by dysregulating antigen-presenting cell maturation. J. Immunol. 1999, 162, 5728–5737. [Google Scholar] [PubMed]
- Dolcetti, L.; Peranzoni, E.; Ugel, S.; Marigo, I.; Gomez, A.F.; Mesa, C.; Geilich, M.; Winkels, G.; Traggiai, E.; Casati, A.; et al. Hierarchy of immunosuppressive strength among myeloid-derived suppressor cell subsets is determined by GM-CSF. Eur. J. Immunol. 2010, 40, 22–35. [Google Scholar] [CrossRef]
- Arman, A.; Auron, P.E. Interleukin 1 (IL-1) Induces the Activation of Stat3. In Tissue Engineering, Stem Cells, and Gene Therapies; Elçin, Y.M., Ed.; Springer: Boston, MA, USA, 2003; pp. 297–307. [Google Scholar]
- Nakajima, K.; Yamanaka, Y.; Nakae, K.; Kojima, H.; Ichiba, M.; Kiuchi, N.; Kitaoka, T.; Fukada, T.; Hibi, M.; Hirano, T. A central role for Stat3 in IL-6-induced regulation of growth and differentiation in M1 leukemia cells. EMBO J. 1996, 15, 3651–3658. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Corzo, C.A.; Luetteke, N.; Yu, B.; Nagaraj, S.; Bui, M.M.; Ortiz, M.; Nacken, W.; Sorg, C.; Vogl, T.; et al. Inhibition of dendritic cell differentiation and accumulation of myeloid-derived suppressor cells in cancer is regulated by S100A9 protein. J. Exp. Med. 2008, 205, 2235–2249. [Google Scholar] [CrossRef]
- Sinha, P.; Okoro, C.; Foell, D.; Freeze, H.H.; Ostrand-Rosenberg, S.; Srikrishna, G. Proinflammatory S100 Proteins Regulate the Accumulation of Myeloid-Derived Suppressor Cells. J. Immunol. 2008, 181, 4666–4675. [Google Scholar] [CrossRef] [Green Version]
- Wagner, N.B.; Weide, B.; Gries, M.; Reith, M.; Tarnanidis, K.; Schuermans, V.; Kemper, C.; Kehrel, C.; Funder, A.; Lichtenberger, R.; et al. Tumor microenvironment-derived S100A8/A9 is a novel prognostic biomarker for advanced melanoma patients and during immunotherapy with anti-PD-1 antibodies. J. Immunother. Cancer 2019, 7, 343. [Google Scholar] [CrossRef]
- Yang, L.; de Busk, L.M.; Fukuda, K.; Fingleton, B.; Green-Jarvis, B.; Shyr, Y.; Matrisian, L.M.; Carbone, D.P.; Lin, P. Expansion of myeloid immune suppressor Gr+CD11b+ cells in tumor-bearing host directly promotes tumor angiogenesis. Cancer Cell 2004, 6, 409–421. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Wilkes, D.W.; Samuel, N.; Blanco, M.A.; Nayak, A.; Alicea-Torres, K.; Gluck, C.; Sinha, S.; Gabrilovich, D.; Chakrabarti, R. ΔNp63-driven recruitment of myeloid-derived suppressor cells promotes metastasis in triple-negative breast cancer. J. Clin. Investig. 2018, 128, 5095–5109. [Google Scholar] [CrossRef] [Green Version]
- Toh, B.; Wang, X.; Keeble, J.; Sim, W.J.; Khoo, K.; Wong, W.-C.; Kato, M.; Prevost-Blondel, A.; Thiery, J.-P.; Abastado, J.-P. Mesenchymal Transition and Dissemination of Cancer Cells Is Driven by Myeloid-Derived Suppressor Cells Infiltrating the Primary Tumor. PLoS Biol. 2011, 9, e1001162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Mitri, D.; Toso, A.; Chen, J.J.; Sarti, M.; Pinton, S.; Jost, T.R.; D’Antuono, R.; Montani, E.; Garcia-Escudero, R.; Guccini, I.; et al. Tumour-infiltrating Gr-1+ myeloid cells antagonize senescence in cancer. Nat. Cell Biol. 2014, 515, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Sinha, P.; Clements, V.K.; Ostrand-Rosenberg, S. Interleukin-13–regulated M2 Macrophages in Combination with Myeloid Suppressor Cells Block Immune Surveillance against Metastasis. Cancer Res. 2005, 65, 11743–11751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, M.; Choksawangkarn, W.; Edwards, N.; Ostrand-Rosenberg, S.; Fenselau, C. Exosomes from Myeloid-Derived Suppressor Cells Carry Biologically Active Proteins. J. Proteome Res. 2014, 13, 836–843. [Google Scholar] [CrossRef] [Green Version]
- Geis-Asteggiante, L.; Belew, A.T.; Clements, V.K.; Edwards, N.J.; Ostrand-Rosenberg, S.; El-Sayed, N.M.; Fenselau, C. Differential Content of Proteins, mRNAs, and miRNAs Suggests that MDSC and Their Exosomes May Mediate Distinct Immune Suppressive Functions. J. Proteome Res. 2018, 17, 486–498. [Google Scholar] [CrossRef]
- Chauhan, S.; Danielson, S.; Clements, V.; Edwards, N.; Ostrand-Rosenberg, S.; Fenselau, C. Surface Glycoproteins of Exosomes Shed by Myeloid-Derived Suppressor Cells Contribute to Function. J. Proteome Res. 2017, 16, 238–246. [Google Scholar] [CrossRef]
- Bronte, V.; Zanovello, P. Regulation of immune responses by L-arginine metabolism. Nat. Rev. Immunol. 2005, 5, 641–654. [Google Scholar] [CrossRef]
- Ezernitchi, A.V.; Vaknin, I.; Cohen-Daniel, L.; Levy, O.; Manaster, E.; Halabi, A.; Pikarsky, E.; Shapira, L.; Baniyash, M. TCR ζ Down-Regulation under Chronic Inflammation Is Mediated by Myeloid Suppressor Cells Differentially Distributed between Various Lymphatic Organs. J. Immunol. 2006, 177, 4763–4772. [Google Scholar] [CrossRef] [Green Version]
- Raber, P.; Ochoa, A.C.; Rodríguez, P.C. Metabolism of L-arginine by myeloid-derived suppressor cells in cancer: Mechanisms of T cell suppression and therapeutic perspectives. Immunol. Investig. 2012, 41, 614–634. [Google Scholar] [CrossRef]
- Yu, J.; Du, W.; Yan, F.; Wang, Y.; Li, H.; Cao, S.; Yu, W.; Shen, C.; Liu, J.; Ren, X. Myeloid-Derived Suppressor Cells Suppress Antitumor Immune Responses through IDO Expression and Correlate with Lymph Node Metastasis in Patients with Breast Cancer. J. Immunol. 2013, 190, 3783–3797. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, M.K.; Sinha, P.; Clements, V.K.; Rodriguez, P.; Ostrand-Rosenberg, S. Myeloid-Derived Suppressor Cells Inhibit T-Cell Activation by Depleting Cystine and Cysteine. Cancer Res. 2010, 70, 68–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaraj, S.; Schrum, A.G.; Cho, H.-I.; Celis, E.; Gabrilovich, D.I. Mechanism of T Cell Tolerance Induced by Myeloid-Derived Suppressor Cells. J. Immunol. 2010, 184, 3106–3116. [Google Scholar] [CrossRef]
- Schmielau, J.; Finn, O.J. Activated granulocytes and granulocyte-derived hydrogen peroxide are the underlying mechanism of suppression of t-cell function in advanced cancer patients. Cancer Res. 2001, 61, 4756–4760. [Google Scholar] [PubMed]
- Lu, T.; Ramakrishnan, R.; Altiok, S.; Youn, J.-I.; Cheng, P.; Celis, E.; Pisarev, V.; Sherman, S.; Sporn, M.B.; Gabrilovich, D. Tumor-infiltrating myeloid cells induce tumor cell resistance to cytotoxic T cells in mice. J. Clin. Investig. 2011, 121, 4015–4029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaraj, S.; Gupta, K.; Pisarev, V.M.; Kinarsky, L.; Sherman, S.; Kang, L.; Herber, D.L.; Schneck, J.P.; Gabrilovich, D.I. Altered recognition of antigen is a mechanism of CD8+ T cell tolerance in cancer. Nat. Med. 2007, 13, 828–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molon, B.; Ugel, S.; Del Pozzo, F.; Soldani, C.; Zilio, S.; Avella, D.; De Palma, A.; Mauri, P.; Monegal, A.; Rescigno, M.; et al. Chemokine nitration prevents intratumoral infiltration of antigen-specific T cells. J. Exp. Med. 2011, 208, 1949–1962. [Google Scholar] [CrossRef]
- Hanson, E.M.; Clements, V.K.; Sinha, P.; Ilkovitch, D.; Ostrand-Rosenberg, S. Myeloid-Derived Suppressor Cells Down-Regulate L-Selectin Expression on CD4+ and CD8+ T Cells. J. Immunol. 2009, 183, 937–944. [Google Scholar] [CrossRef] [Green Version]
- Ku, A.W.; Muhitch, J.B.; Powers, C.A.; Diehl, M.; Kim, M.; Fisher, D.T.; Sharda, A.P.; Clements, V.K.; O’Loughlin, K.; Minderman, H.; et al. Tumor-induced MDSC act via remote control to inhibit L-selectin-dependent adaptive immunity in lymph nodes. eLife 2016, 5, e17375. [Google Scholar] [CrossRef]
- Hu, C.-E.; Gan, J.; Zhang, R.-D.; Cheng, Y.-R.; Huang, G.-J. Up-regulated myeloid-derived suppressor cell contributes to hepatocellular carcinoma development by impairing dendritic cell function. Scand. J. Gastroenterol. 2010, 46, 156–164. [Google Scholar] [CrossRef]
- Huang, B.; Pan, P.-Y.; Li, Q.; Sato, A.I.; Levy, D.E.; Bromberg, J.; Divino, C.M.; Chen, S.-H. Gr-1+CD115+ Immature Myeloid Suppressor Cells Mediate the Development of Tumor-Induced T Regulatory Cells and T-Cell Anergy in Tumor-Bearing Host. Cancer Res. 2006, 66, 1123–1131. [Google Scholar] [CrossRef] [Green Version]
- Pan, P.-Y.; Ma, G.; Weber, K.J.; Ozao-Choy, J.; Wang, G.; Yin, B.; Divino, C.M.; Chen, S.-H. Immune Stimulatory Receptor CD40 Is Required for T-Cell Suppression and T Regulatory Cell Activation Mediated by Myeloid-Derived Suppressor Cells in Cancer. Cancer Res. 2010, 70, 99–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoechst, B.; Voigtlaender, T.; Ormandy, L.; Gamrekelashvili, J.; Zhao, F.; Wedemeyer, H.; Lehner, F.; Manns, M.P.; Greten, T.F.; Korangy, F. Myeloid derived suppressor cells inhibit natural killer cells in patients with hepatocellular carcinoma via the NKp30 receptor. Hepatology 2009, 50, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yu, S.; Kappes, J.; Wang, J.; Grizzle, W.E.; Zinn, K.R.; Zhang, H.-G. Expansion of spleen myeloid suppressor cells represses NK cell cytotoxicity in tumor-bearing host. Blood 2007, 109, 4336–4342. [Google Scholar] [CrossRef] [PubMed]
- Elkabets, M.; Ribeiro, V.S.G.; Dinarello, C.A.; Ostrand-Rosenberg, S.; Di Santo, J.P.; Apte, R.N.; Vosshenrich, C.A.J. IL-1β regulates a novel myeloid-derived suppressor cell subset that impairs NK cell development and function. Eur. J. Immunol. 2010, 40, 3347–3357. [Google Scholar] [CrossRef] [PubMed]
- Beury, D.W.; Parker, K.H.; Nyandjo, M.; Sinha, P.; Carter, K.A.; Ostrand-Rosenberg, S. Cross-talk among myeloid-derived suppressor cells, macrophages, and tumor cells impacts the inflammatory milieu of solid tumors. J. Leukoc. Biol. 2014, 96, 1109–1118. [Google Scholar] [CrossRef]
- Bunt, S.K.; Clements, V.K.; Hanson, E.M.; Sinha, P.; Ostrand-Rosenberg, S. Inflammation enhances myeloid-derived suppressor cell cross-talk by signaling through Toll-like receptor 4. J. Leukoc. Biol. 2009, 85, 996–1004. [Google Scholar] [CrossRef] [Green Version]
- Sinha, P.; Clements, V.K.; Bunt, S.K.; Albelda, S.M.; Ostrand-Rosenberg, S. Cross-Talk between Myeloid-Derived Suppressor Cells and Macrophages Subverts Tumor Immunity toward a Type 2 Response. J. Immunol. 2007, 179, 977–983. [Google Scholar] [CrossRef]
- Jain, M.D.; Zhao, H.; Wang, X.; Atkins, R.; Menges, M.; Reid, K.; Spitler, K.; Faramand, R.; Bachmeier, C.A.; Dean, E.A.; et al. Tumor interferon signaling and suppressive myeloid cells associate with CAR T cell failure in large B cell lymphoma. Blood 2021. [Google Scholar] [CrossRef]
- Mao, Y.; Eissler, N.; le Blanc, K.; Johnsen, J.I.; Kogner, P.; Kiessling, R. Targeting Suppressive Myeloid Cells Potentiates Checkpoint Inhibitors to Control Spontaneous Neuroblastoma. Clin. Cancer Res. 2016, 22, 3849–3859. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Garcia, A.; Lynn, R.C.; Poussin, M.; Eiva, M.A.; Shaw, L.C.; O’Connor, R.S.; Minutolo, N.G.; Casado-Medrano, V.; Lopez, G.; Matsuyama, T.; et al. CAR-T cell-mediated depletion of immunosuppressive tumor-associated macrophages promotes endogenous antitumor immunity and augments adoptive immunotherapy. Nat. Commun. 2021, 12, 877. [Google Scholar] [CrossRef]
- Long, A.H.; Highfill, S.L.; Cui, Y.; Smith, J.P.; Walker, A.J.; Ramakrishna, S.; El-Etriby, R.; Galli, S.; Tsokos, M.G.; Orentas, R.J.; et al. Reduction of MDSCs with All-trans Retinoic Acid Improves CAR Therapy Efficacy for Sarcomas. Cancer Immunol. Res. 2016, 4, 869–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fultang, L.; Panetti, S.; Ng, M.; Collins, P.; Graef, S.; Rizkalla, N.; Booth, S.; Lenton, R.; Noyvert, B.; Shannon-Lowe, C.; et al. MDSC targeting with Gemtuzumab ozogamicin restores T cell immunity and immunotherapy against cancers. EBioMedicine 2019, 47, 235–246. [Google Scholar] [CrossRef] [Green Version]
- Caescu, C.I.; Guo, X.; Tesfa, L.; Bhagat, T.D.; Verma, A.; Zheng, D.; Stanley, E.R. Colony stimulating factor-1 receptor signaling networks inhibit mouse macrophage inflammatory responses by induction of microRNA-21. Blood 2015, 125, e1–e13. [Google Scholar] [CrossRef] [PubMed]
- Ries, C.H.; Cannarile, M.A.; Hoves, S.; Benz, J.; Wartha, K.; Runza, V.; Rey-Giraud, F.; Pradel, L.P.; Feuerhake, F.; Klaman, I.; et al. Targeting Tumor-Associated Macrophages with Anti-CSF-1R Antibody Reveals a Strategy for Cancer Therapy. Cancer Cell 2014, 25, 846–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manthey, C.L.; Johnson, D.L.; Illig, C.R.; Tuman, R.W.; Zhou, Z.; Baker, J.F.; Chaikin, M.A.; Donatelli, R.R.; Franks, C.F.; Zeng, L.; et al. JNJ-28312141, a novel orally active colony-stimulating factor-1 receptor/FMS-related receptor tyrosine kinase-3 receptor tyrosine kinase inhibitor with potential utility in solid tumors, bone metastases, and acute myeloid leukemia. Mol. Cancer Ther. 2009, 8, 3151–3161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyonteck, S.M.; Akkari, L.; Schuhmacher, A.J.; Bowman, R.L.; Sevenich, L.; Quail, D.F.; Olson, O.C.; Quick, M.L.; Huse, J.T.; Teijeiro, V.; et al. CSF-1R inhibition alters macrophage polarization and blocks glioma progression. Nat. Med. 2013, 19, 1264–1272. [Google Scholar] [CrossRef] [Green Version]
- Qian, B.-Z.; Li, J.; Zhang, H.; Kitamura, T.; Zhang, J.; Campion, L.R.; Kaiser, E.A.; Snyder, L.A.; Pollard, J.W. CCL2 recruits inflammatory monocytes to facilitate breast-tumour metastasis. Nature 2011, 475, 222–225. [Google Scholar] [CrossRef] [Green Version]
- Sanford, D.E.; Belt, B.A.; Panni, R.Z.; Mayer, A.; Deshpande, A.D.; Carpenter, D.; Mitchem, J.B.; Plambeck-Suess, S.M.; Worley, L.A.; Goetz, B.D.; et al. Inflammatory Monocyte Mobilization Decreases Patient Survival in Pancreatic Cancer: A Role for Targeting the CCL2/CCR2 Axis. Clin. Cancer Res. 2013, 19, 3404–3415. [Google Scholar] [CrossRef] [Green Version]
- Nywening, T.M.; Wang-Gillam, A.; Sanford, D.E.; Belt, B.A.; Panni, R.Z.; Cusworth, B.M.; Toriola, A.T.; Nieman, R.K.; Worley, L.A.; Yano, M.; et al. Targeting tumour-associated macrophages with CCR2 inhibition in combination with FOLFIRINOX in patients with borderline resectable and locally advanced pancreatic cancer: A single-centre, open-label, dose-finding, non-randomised, phase 1b trial. Lancet Oncol. 2016, 17, 651–662. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, S.K.; Papadopoulos, K.; Fong, P.C.; Patnaik, A.; Messiou, C.; Olmos, D.; Wang, G.; Tromp, B.J.; Puchalski, T.A.; Balkwill, F.; et al. A first-in-human, first-in-class, phase I study of carlumab (CNTO 888), a human monoclonal antibody against CC-chemokine ligand 2 in patients with solid tumors. Cancer Chemother. Pharmacol. 2013, 71, 1041–1050. [Google Scholar] [CrossRef]
- Pienta, K.J.; Machiels, J.-P.; Schrijvers, D.; Alekseev, B.; Shkolnik, M.; Crabb, S.J.; Li, S.; Seetharam, S.; Puchalski, T.A.; Takimoto, C.; et al. Phase 2 study of carlumab (CNTO 888), a human monoclonal antibody against CC-chemokine ligand 2 (CCL2), in metastatic castration-resistant prostate cancer. Investig. New Drugs 2013, 31, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, E.; Wenthe, J.; Irenaeus, S.; Loskog, A.; Ullenhag, G. Gemcitabine reduces MDSCs, tregs and TGFβ-1 while restoring the teff/treg ratio in patients with pancreatic cancer. J. Transl. Med. 2016, 14, 282. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, E.; Kapoor, V.; Jassar, A.S.; Kaiser, L.R.; Albelda, S.M. Gemcitabine Selectively Eliminates Splenic Gr-1+/CD11b+ Myeloid Suppressor Cells in Tumor-Bearing Animals and Enhances Antitumor Immune Activity. Clin. Cancer Res. 2005, 11, 6713–6721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, H.K.; Graham, L.; Cha, E.; Morales, J.K.; Manjili, M.H.; Bear, H.D. Gemcitabine directly inhibits myeloid derived suppressor cells in BALB/c mice bearing 4T1 mammary carcinoma and augments expansion of T cells from tumor-bearing mice. Int. Immunopharmacol. 2009, 9, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.; Mignot, G.; Chalmin, F.; Ladoire, S.; Bruchard, M.; Chevriaux, A.; Martin, F.; Apetoh, L.; Rébé, C.; Ghiringhelli, F. 5-Fluorouracil Selectively Kills Tumor-Associated Myeloid-Derived Suppressor Cells Resulting in Enhanced T Cell–Dependent Antitumor Immunity. Cancer Res. 2010, 70, 3052–3061. [Google Scholar] [CrossRef] [Green Version]
- Bruchard, M.; Mignot, G.; Derangère, V.; Chalmin, F.; Chevriaux, A.; Végran, F.; Boireau, W.; Simon, B.; Ryffel, B.; Connat, J.-L.; et al. Chemotherapy-triggered cathepsin B release in myeloid-derived suppressor cells activates the Nlrp3 inflammasome and promotes tumor growth. Nat. Med. 2013, 19, 57–64. [Google Scholar] [CrossRef]
- Germano, G.; Frapolli, R.; Belgiovine, C.; Anselmo, A.; Pesce, S.; Liguori, M.; Erba, E.; Uboldi, S.; Zucchetti, M.; Pasqualini, F.; et al. Role of Macrophage Targeting in the Antitumor Activity of Trabectedin. Cancer Cell 2013, 23, 249–262. [Google Scholar] [CrossRef] [Green Version]
- D’Incalci, M.; Colombo, T.; Ubezio, P.; Nicoletti, I.; Giavazzi, R.; Erba, E.; Ferrarese, L.; Meco, D.; Riccardi, R.; Sessa, C.; et al. The combination of yondelis and cisplatin is synergistic against human tumor xenografts. Eur. J. Cancer 2003, 39, 1920–1926. [Google Scholar] [CrossRef]
- Siebert, N.; Zumpe, M.; von Lojewski, L.; Troschke-Meurer, S.; Marx, M.; Lode, H.N. Reduction of CD11b+ myeloid suppressive cells augments anti-neuroblastoma immune response induced by the anti-GD2 antibody ch14.18/CHO. Oncoimmunology 2020, 9, 1836768. [Google Scholar] [CrossRef]
- Guislain, A.; Gadiot, J.; Kaiser, A.; Jordanova, E.S.; Broeks, A.; Sanders, J.; van Boven, H.; de Gruijl, T.D.; Haanen, J.B.A.G.; Bex, A.; et al. Sunitinib pretreatment improves tumor-infiltrating lymphocyte expansion by reduction in intratumoral content of myeloid-derived suppressor cells in human renal cell carcinoma. Cancer Immunol. Immunother. 2015, 64, 1241–1250. [Google Scholar] [CrossRef]
- Zhao, Q.; Guo, J.; Wang, G.; Chu, Y.; Hu, X. Suppression of immune regulatory cells with combined therapy of celecoxib and sunitinib in renal cell carcinoma. Oncotarget 2017, 8, 1668–1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, J.S.; Zea, A.H.; Rini, B.I.; Ireland, J.L.; Elson, P.; Cohen, P.; Golshayan, A.; Rayman, P.A.; Wood, L.; Garcia, J.; et al. Sunitinib Mediates Reversal of Myeloid-Derived Suppressor Cell Accumulation in Renal Cell Carcinoma Patients. Clin. Cancer Res. 2009, 15, 2148–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hooren, L.; Georganaki, M.; Huang, H.; Mangsbo, S.M.; Dimberg, A. Sunitinib enhances the antitumor responses of agonistic CD40-antibody by reducing MDSCs and synergistically improving endothelial activation and T-cell recruitment. Oncotarget 2016, 7, 50277–50289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, S.G.; Shusterman, S.; Ingle, A.M.; Ahern, C.H.; Reid, J.M.; Wu, B.; Baruchel, S.; Glade-Bender, J.; Ivy, P.; Grier, H.E.; et al. Phase I and Pharmacokinetic Study of Sunitinib in Pediatric Patients with Refractory Solid Tumors: A Children’s Oncology Group Study. Clin. Cancer Res. 2011, 17, 5113–5122. [Google Scholar] [CrossRef] [Green Version]
- Steggerda, S.M.; Bennett, M.K.; Chen, J.; Emberley, E.; Huang, T.; Janes, J.R.; Silinda, N.; MacKinnon, A.L.; Makkouk, A.; Marguier, G.; et al. Inhibition of arginase by CB-1158 blocks myeloid cell-mediated immune suppression in the tumor microenvironment. J. Immunother. Cancer 2017, 5, 101. [Google Scholar] [CrossRef]
- Miret, J.J.; Kirschmeier, P.; Koyama, S.; Zhu, M.; Li, Y.Y.; Naito, Y.; Wu, M.; Malladi, V.S.; Huang, W.; Walker, W.; et al. Suppression of Myeloid Cell Arginase Activity leads to Therapeutic Response in a NSCLC Mouse Model by Activating Anti-Tumor Immunity. J. Immunother. Cancer 2019, 7, 32. [Google Scholar] [CrossRef]
- Li, A.; Barsoumian, H.B.; Schoenhals, J.E.; Caetano, M.S.; Wang, X.; Menon, H.; Valdecanas, D.R.; Niknam, S.; Younes, A.I.; Cortez, M.A.; et al. IDO1 Inhibition Overcomes Radiation-Induced “Rebound Immune Suppression” by Reducing Numbers of IDO1-Expressing Myeloid-Derived Suppressor Cells in the Tumor Microenvironment. Int. J. Radiat. Oncol. 2019, 104, 903–912. [Google Scholar] [CrossRef]
- Prendergast, G.C.; Malachowski, W.J.; Mondal, A.; Scherle, P.; Muller, A.J. Chapter Four—Indoleamine 2,3-Dioxygenase and Its Therapeutic Inhibition in Cancer. In International Review of Cell and Molecular Biology; Galluzzi, L., Ed.; Academic Press: Cambridge, MA, USA, 2018; Volume 336, pp. 175–203. [Google Scholar]
- Fujita, M.; Kohanbash, G.; Fellows-Mayle, W.; Hamilton, R.L.; Komohara, Y.; Decker, S.A.; Ohlfest, J.R.; Okada, H. COX-2 Blockade Suppresses Gliomagenesis by Inhibiting Myeloid-Derived Suppressor Cells. Cancer Res. 2011, 71, 2664–2674. [Google Scholar] [CrossRef] [Green Version]
- Veltman, J.D.; Lambers, M.E.; van Nimwegen, M.; Hendriks, R.W.; Hoogsteden, H.C.; Aerts, J.G.; Hegmans, J.P. COX-2 inhibition improves immunotherapy and is associated with decreased numbers of myeloid-derived suppressor cells in mesothelioma. Celecoxib influences MDSC function. BMC Cancer 2010, 10, 464. [Google Scholar] [CrossRef] [Green Version]
- Davidov, V.; Jensen, G.; Mai, S.; Chen, S.-H.; Pan, P.-Y. Analyzing One Cell at a TIME: Analysis of Myeloid Cell Contributions in the Tumor Immune Microenvironment. Front. Immunol. 2020, 11, 1842. [Google Scholar] [CrossRef]
- Joshi, S.; Singh, A.R.; Liu, K.X.; Pham, T.V.; Zulcic, M.; Skola, D.; Chun, H.B.; Glass, C.K.; Morales, G.A.; Garlich, J.R.; et al. SF2523: Dual PI3K/BRD4 Inhibitor Blocks Tumor Immunosuppression and Promotes Adaptive Immune Responses in Cancer. Mol. Cancer Ther. 2019, 18, 1036–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puissant, A.; Frumm, S.M.; Alexe, G.; Bassil, C.F.; Qi, J.; Chanthery, Y.H.; Nekritz, E.A.; Zeid, R.; Gustafson, W.C.; Greninger, P.; et al. Targeting MYCN in neuroblastoma by BET bromodomain inhibition. Cancer Discov. 2013, 3, 308–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroesen, M.; Büll, C.; Gielen, P.R.; Brok, I.C.; Armandari, I.; Wassink, M.; Looman, M.W.G.; Boon, L.; Brok, M.H.D.; Hoogerbrugge, P.M.; et al. Anti-GD2 mAb and Vorinostat synergize in the treatment of neuroblastoma. OncoImmunology 2016, 5, e1164919. [Google Scholar] [CrossRef] [PubMed]
- Christmas, B.J.; Rafie, C.I.; Hopkins, A.C.; Scott, B.A.; Ma, H.S.; Cruz, K.A.; Woolman, S.; Armstrong, T.D.; Connolly, R.M.; Azad, N.A.; et al. Entinostat Converts Immune-Resistant Breast and Pancreatic Cancers into Checkpoint-Responsive Tumors by Reprogramming Tumor-Infiltrating MDSCs. Cancer Immunol. Res. 2018, 6, 1561–1577. [Google Scholar] [CrossRef] [Green Version]
- Orillion, A.; Hashimoto, A.; Damayanti, N.; Shen, L.; Adelaiye-Ogala, R.; Arisa, S.; Chintala, S.; Ordentlich, P.; Kao, C.; Elzey, B.; et al. Entinostat Neutralizes Myeloid-Derived Suppressor Cells and Enhances the Antitumor Effect of PD-1 Inhibition in Murine Models of Lung and Renal Cell Carcinoma. Clin. Cancer Res. 2017, 23, 5187–5201. [Google Scholar] [CrossRef] [Green Version]
- Nefedova, Y.; Fishman, M.; Sherman, S.; Wang, X.; Beg, A.A.; Gabrilovich, D.I. Mechanism of All-Trans Retinoic Acid Effect on Tumor-Associated Myeloid-Derived Suppressor Cells. Cancer Res. 2007, 67, 11021–11028. [Google Scholar] [CrossRef] [Green Version]
- Tobin, R.P.; Jordan, K.R.; Robinson, W.A.; Davis, D.; Borges, V.F.; Gonzalez, R.; Lewis, K.D.; McCarter, M.D. Targeting myeloid-derived suppressor cells using all-trans retinoic acid in melanoma patients treated with Ipilimumab. Int. Immunopharmacol. 2018, 63, 282–291. [Google Scholar] [CrossRef]
- Iclozan, C.; Antonia, S.; Chiappori, A.; Chen, D.-T.; Gabrilovich, D. Therapeutic regulation of myeloid-derived suppressor cells and immune response to cancer vaccine in patients with extensive stage small cell lung cancer. Cancer Immunol. Immunother. 2013, 62, 909–918. [Google Scholar] [CrossRef] [Green Version]
- Kusmartsev, S.; Cheng, F.; Yu, B.; Nefedova, Y.; Sotomayor, E.; Lush, R.; Gabrilovich, D. All-trans-retinoic acid eliminates immature myeloid cells from tumor-bearing mice and improves the effect of vaccination. Cancer Res. 2003, 63, 4441–4449. [Google Scholar]
- Relation, T.; Yi, T.; Guess, A.J.; La Perle, K.; Otsuru, S.; Hasgur, S.; Dominici, M.; Breuer, C.; Horwitz, E.M. Intratumoral Delivery of Interferonγ-Secreting Mesenchymal Stromal Cells Repolarizes Tumor-Associated Macrophages and Suppresses Neuroblastoma Proliferation In Vivo. Stem Cells 2018, 36, 915–924. [Google Scholar] [CrossRef] [Green Version]
- Komorowski, M.; Tisonczyk, J.; Kolakowska, A.; Drozdz, R.; Kozbor, D. Modulation of the Tumor Microenvironment by CXCR4 Antagonist-Armed Viral Oncotherapy Enhances the Antitumor Efficacy of Dendritic Cell Vaccines against Neuroblastoma in Syngeneic Mice. Viruses 2018, 10, 455. [Google Scholar] [CrossRef] [Green Version]
- Cassier, P.A.; Italiano, A.; Gomez-Roca, C.A.; le Tourneau, C.; Toulmonde, M.; Cannarile, M.A.; Ries, C.; Brillouet, A.; Müller, C.; Jegg, A.-M.; et al. CSF1R inhibition with emactuzumab in locally advanced diffuse-type tenosynovial giant cell tumours of the soft tissue: A dose-escalation and dose-expansion phase 1 study. Lancet Oncol. 2015, 16, 949–956. [Google Scholar] [CrossRef]
- Gomez-Roca, C.; Italiano, A.; Le Tourneau, C.; Cassier, P.; Toulmonde, M.; D’Angelo, S.; Campone, M.; Weber, K.; Loirat, D.; Cannarile, M.; et al. Phase I study of emactuzumab single agent or in combination with paclitaxel in patients with advanced/metastatic solid tumors reveals depletion of immunosuppressive M2-like macrophages. Ann. Oncol. 2019, 30, 1381–1392. [Google Scholar] [CrossRef]
- Zhu, Y.; Knolhoff, B.L.; Meyer, M.A.; Nywening, T.M.; West, B.L.; Luo, J.; Wang-Gillam, A.; Goedegebuure, S.P.; Linehan, D.C.; DeNardo, D.G. CSF1/CSF1R Blockade Reprograms Tumor-Infiltrating Macrophages and Improves Response to T-cell Checkpoint Immunotherapy in Pancreatic Cancer Models. Cancer Res. 2014, 74, 5057–5069. [Google Scholar] [CrossRef] [Green Version]
- Tap, W.D.; Gelderblom, H.; Palmerini, E.; Desai, J.; Bauer, S.; Blay, J.-Y.; Alcindor, T.; Ganjoo, K.; Martín-Broto, J.; Ryan, C.W.; et al. Pexidartinib versus placebo for advanced tenosynovial giant cell tumour (ENLIVEN): A randomised phase 3 trial. Lancet 2019, 394, 478–487. [Google Scholar] [CrossRef]
- Eissler, N.; Mao, Y.; Brodin, D.; Reuterswärd, P.; Svahn, H.A.; Johnsen, J.I.; Kiessling, R.; Kogner, P. Regulation of myeloid cells by activated T cells determines the efficacy of PD-1 blockade. OncoImmunology 2016, 5, e1232222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priceman, S.J.; Sung, J.L.; Shaposhnik, Z.; Burton, J.B.; Torres-Collado, A.X.; Moughon, D.L.; Johnson, M.; Lusis, A.J.; Cohen, D.A.; Iruela-Arispe, M.L.; et al. Targeting distinct tumor-infiltrating myeloid cells by inhibiting CSF-1 receptor: Combating tumor evasion of antiangiogenic therapy. Blood 2010, 115, 1461–1471. [Google Scholar] [CrossRef] [Green Version]
- Turovskaya, O.; Foell, D.; Sinha, P.; Vogl, T.; Newlin, R.; Nayak, J.; Nguyen, M.; Olsson, A.; Nawroth, P.P.; Bierhaus, A.; et al. RAGE, carboxylated glycans and S100A8/A9 play essential roles in colitis-associated carcinogenesis. Carcinogenesis 2008, 29, 2035–2043. [Google Scholar] [CrossRef]
- Mirza, N.; Fishman, M.; Fricke, I.; Dunn, M.; Neuger, A.M.; Frost, T.J.; Lush, R.M.; Antonia, S.; Gabrilovich, D.I. All-trans-Retinoic Acid Improves Differentiation of Myeloid Cells and Immune Response in Cancer Patients. Cancer Res. 2006, 66, 9299–9307. [Google Scholar] [CrossRef] [Green Version]
- Joshi, S.; Liu, K.X.; Zulcic, M.; Singh, A.R.; Skola, D.; Glass, C.K.; Sanders, P.D.; Sharabi, A.B.; Pham, T.V.; Tamayo, P.; et al. Macrophage Syk–PI3Kγ Inhibits Antitumor Immunity: SRX3207, a Novel Dual Syk–PI3K Inhibitory Chemotype Relieves Tumor Immunosuppression. Mol. Cancer Ther. 2020, 19, 755–764. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Skora, A.D.; Li, Z.; Liu, Q.; Tam, A.J.; Blosser, R.L.; Diaz, L.A.; Papadopoulos, N.; Kinzler, K.W.; Vogelstein, B.; et al. Eradication of metastatic mouse cancers resistant to immune checkpoint blockade by suppression of myeloid-derived cells. Proc. Natl. Acad. Sci. USA 2014, 111, 11774–11779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhtoiarov, I.N.; Sondel, P.M.; Wigginton, J.M.; Buhtoiarova, T.N.; Yanke, E.M.; Mahvi, D.A.; Rakhmilevich, A.L. Anti-tumour synergy of cytotoxic chemotherapy and anti-CD40 plus CpG-ODN immunotherapy through repolarization of tumour-associated macrophages. Immunology 2010, 132, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Buhtoiarov, I.N.; Lum, H.; Berke, G.; Paulnock, D.M.; Sondel, P.M.; Rakhmilevich, A.L. CD40 Ligation Activates Murine Macrophages via an IFN-γ-Dependent Mechanism Resulting in Tumor Cell Destruction In Vitro. J. Immunol. 2005, 174, 6013–6022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lum, H.D.; Buhtoiarov, I.N.; Schmidt, B.E.; Berke, G.; Paulnock, D.M.; Sondel, P.M.; Rakhmilevich, A.L. In vivo CD40 ligation can induce T cell-independent antitumor effects that involve macrophages. J. Leukoc. Biol. 2006, 79, 1181–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voeller, J.; Erbe, A.K.; Slowinski, J.; Rasmussen, K.; Carlson, P.M.; Hoefges, A.; Vandenheuvel, S.; Stuckwisch, A.; Wang, X.; Gillies, S.D.; et al. Combined innate and adaptive immunotherapy overcomes resistance of immunologically cold syngeneic murine neuroblastoma to checkpoint inhibition. J. Immunother. Cancer 2019, 7, 344. [Google Scholar] [CrossRef] [PubMed]
- Beatty, G.L.; Chiorean, E.G.; Fishman, M.P.; Saboury, B.; Teitelbaum, U.R.; Sun, W.; Huhn, R.D.; Song, W.; Li, D.; Sharp, L.L.; et al. CD40 Agonists Alter Tumor Stroma and Show Efficacy Against Pancreatic Carcinoma in Mice and Humans. Science 2011, 331, 1612–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rooijen, N.; Sanders, A.; Berg, T.K.V.D. Apoptosis of macrophages induced by liposome-mediated intracellular delivery of clodronate and propamidine. J. Immunol. Methods 1996, 193, 93–99. [Google Scholar] [CrossRef]
- Van Rooijen, N.; Sanders, A. Liposome mediated depletion of macrophages: Mechanism of action, preparation of liposomes and applications. J. Immunol. Methods 1994, 174, 83–93. [Google Scholar] [CrossRef]
- Sousa, S.; Auriola, S.; Mönkkönen, J.; Määttä, J. Liposome encapsulated zoledronate favours M1-like behaviour in murine macrophages cultured with soluble factors from breast cancer cells. BMC Cancer 2015, 15, 4. [Google Scholar] [CrossRef] [Green Version]
- Zeisberger, S.M.; Odermatt, B.; Marty, C.; Zehnder-Fjällman, A.H.M.; Ballmer-Hofer, K.; Schwendener, R.A. Clodronate-liposome-mediated depletion of tumour-associated macrophages: A new and highly effective antiangiogenic therapy approach. Br. J. Cancer 2006, 95, 272–281. [Google Scholar] [CrossRef]
- Veltman, J.; Lambers, M.; van Nimwegen, M.; Hendriks, R.; Hoogsteden, H.; Hegmans, J.; Aerts, J. Zoledronic acid impairs myeloid differentiation to tumour-associated macrophages in mesothelioma. Br. J. Cancer 2010, 103, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Draghiciu, O.; Nijman, H.W.; Hoogeboom, B.N.; Meijerhof, T.; Daemen, T. Sunitinib depletes myeloid-derived suppressor cells and synergizes with a cancer vaccine to enhance antigen-specific immune responses and tumor eradication. OncoImmunology 2015, 4, e989764. [Google Scholar] [CrossRef] [Green Version]
- Tu, S.P.; Jin, H.; Shi, J.D.; Zhu, L.M.; Suo, Y.; Lu, G.; Liu, A.; Wang, T.C.; Yang, C.S. Curcumin Induces the Differentiation of Myeloid-Derived Suppressor Cells and Inhibits Their Interaction with Cancer Cells and Related Tumor Growth. Cancer Prev. Res. 2012, 5, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Wolff, J.E.; Brown, R.E.; Buryanek, J.; Pfister, S.; Vats, T.S.; Rytting, M.E. Preliminary experience with personalized and targeted therapy for pediatric brain tumors. Pediatr. Blood Cancer 2011, 59, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Ramos, I.; Asafu-Adjei, D.; Quispe-Tintaya, W.; Chandra, D.; Jahangir, A.; Zang, X.; Aggarwal, B.B.; Gravekamp, C. Curcumin improves the therapeutic efficacy of Listeriaat-Mage-b vaccine in correlation with improved T-cell responses in blood of a triple-negative breast cancer model 4T1. Cancer Med. 2013, 2, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Donatelli, S.S.; Gilvary, D.L.; Tejera, M.M.; Eksioglu, E.A.; Chen, X.; Coppola, M.; Wei, S.; Djeu, J.Y. Therapeutic targeting of myeloid-derived suppressor cells involves a novel mechanism mediated by clusterin. Sci. Rep. 2016, 6, 29521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, P.; Ostrand-Rosenberg, S. Myeloid-derived suppressor cell function is reduced by Withaferin A, a potent and abundant component of Withania somnifera root extract. Cancer Immunol. Immunother. 2013, 62, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Annels, N.E.; Shaw, V.E.; Gabitass, R.F.; Billingham, L.; Corrie, P.; Eatock, M.; Valle, J.; Smith, D.; Wadsley, J.; Cunningham, D.; et al. The effects of gemcitabine and capecitabine combination chemotherapy and of low-dose adjuvant GM-CSF on the levels of myeloid-derived suppressor cells in patients with advanced pancreatic cancer. Cancer Immunol. Immunother. 2013, 63, 175–183. [Google Scholar] [CrossRef]
- Wu, H.-W.; Sheard, M.A.; Malvar, J.; Fernandez, G.E.; Declerck, Y.A.; Blavier, L.; Shimada, H.; Theuer, C.P.; Sposto, R.; Seeger, R.C. Anti-CD105 Antibody Eliminates Tumor Microenvironment Cells and Enhances Anti-GD2 Antibody Immunotherapy of Neuroblastoma with Activated Natural Killer Cells. Clin. Cancer Res. 2019, 25, 4761–4774. [Google Scholar] [CrossRef] [Green Version]
- Karakasheva, T.A.; Dominguez, G.A.; Hashimoto, A.; Lin, E.W.; Chiu, C.; Sasser, K.; Lee, J.W.; Beatty, G.L.; Gabrilovich, D.I.; Rustgi, A.K. CD38+ M-MDSC expansion characterizes a subset of advanced colorectal cancer patients. JCI Insight 2018, 3. [Google Scholar] [CrossRef]
- Chen, L.; Diao, L.; Yang, Y.; Yi, X.; Rodriguez, B.L.; Li, Y.; Villalobos, P.A.; Cascone, T.; Liu, X.; Tan, L.; et al. CD38-Mediated Immunosuppression as a Mechanism of Tumor Cell Escape from PD-1/PD-L1 Blockade. Cancer Discov. 2018, 8, 1156–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krejcik, J.; Casneuf, T.; Nijhof, I.S.; Verbist, B.; Bald, J.; Plesner, T.; Syed, K.; Liu, K.; van de Donk, N.W.C.J.; Weiss, B.M.; et al. Daratumumab depletes CD38+ immune regulatory cells, promotes T-cell expansion, and skews T-cell repertoire in multiple myeloma. Blood 2016, 128, 384–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Bardhan, K.; Paschall, A.V.; Yang, D.; Waller, J.L.; Park, M.A.; Nayak-Kapoor, A.; Samuel, T.A.; Abrams, S.I.; Liu, K. Deregulation of Apoptotic Factors Bcl-xL and Bax Confers Apoptotic Resistance to Myeloid-derived Suppressor Cells and Contributes to Their Persistence in Cancer. J. Biol. Chem. 2013, 288, 19103–19115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleason, M.K.; Ross, J.A.; Warlick, E.D.; Lund, T.C.; Verneris, M.R.; Wiernik, A.; Spellman, S.; Haagenson, M.D.; Lenvik, A.J.; Litzow, M.R.; et al. CD16xCD33 bispecific killer cell engager (BiKE) activates NK cells against primary MDS and MDSC CD33+ targets. Blood 2014, 123, 3016–3026. [Google Scholar] [CrossRef]
- Condamine, T.; Kumar, V.; Ramachandran, I.R.; Youn, J.-I.; Celis, E.; Finnberg, N.; El-Deiry, W.S.; Winograd, R.; Vonderheide, R.H.; English, N.R.; et al. ER stress regulates myeloid-derived suppressor cell fate through TRAIL-R–mediated apoptosis. J. Clin. Investig. 2014, 124, 2626–2639. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, G.A.; Condamine, T.; Mony, S.; Hashimoto, A.; Wang, F.; Liu, Q.; Forero, A.; Bendell, J.; Witt, R.; Hockstein, N.; et al. Selective Targeting of Myeloid-Derived Suppressor Cells in Cancer Patients Using DS-8273a, an Agonistic TRAIL-R2 Antibody. Clin. Cancer Res. 2017, 23, 2942–2950. [Google Scholar] [CrossRef] [Green Version]
- Roth, F.; de la Fuente, A.C.; Vella, J.L.; Zoso, A.; Inverardi, L.; Serafini, P. Aptamer-Mediated Blockade of IL4Rα Triggers Apoptosis of MDSCs and Limits Tumor Progression. Cancer Res. 2012, 72, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Theruvath, J.; Smith, B.; Linde, M.H.; Sotillo, E.; Heitzeneder, S.; Marjon, K.; Tousley, A.; Lattin, J.; Banuelos, A.; Dhingra, S.; et al. Abstract PR07: GD2 is a macrophage checkpoint molecule and combined GD2/CD47 blockade results in synergistic effects and tumor clearance in xenograft models of neuroblastoma and osteosarcoma. Cancer Res. Oral Present. Proffered Abstr. 2020, 80, PR07. [Google Scholar] [CrossRef]
- Wu, L.; Yu, G.-T.; Deng, W.-W.; Mao, L.; Yang, L.-L.; Ma, S.-R.; Bu, L.-L.; Kulkarni, A.B.; Zhang, W.-F.; Zhang, L.; et al. Anti-CD47 treatment enhances anti-tumor T-cell immunity and improves immunosuppressive environment in head and neck squamous cell carcinoma. OncoImmunology 2017, 7, e1397248. [Google Scholar] [CrossRef]
- Obermajer, N.; Muthuswamy, R.; Odunsi, K.O.; Edwards, R.P.; Kalinski, P. PGE2-Induced CXCL12 Production and CXCR4 Expression Controls the Accumulation of Human MDSCs in Ovarian Cancer Environment. Cancer Res. 2011, 71, 7463–7470. [Google Scholar] [CrossRef] [Green Version]
- Talmadge, J.E.; Hood, K.C.; Zobel, L.C.; Shafer, L.R.; Coles, M.; Toth, B. Chemoprevention by cyclooxygenase-2 inhibition reduces immature myeloid suppressor cell expansion. Int. Immunopharmacol. 2007, 7, 140–151. [Google Scholar] [CrossRef]
- Obermajer, N.; Muthuswamy, R.; Lesnock, J.; Edwards, R.P.; Kalinski, P. Positive feedback between PGE2 and COX2 redirects the differentiation of human dendritic cells toward stable myeloid-derived suppressor cells. Blood 2011, 118, 5498–5505. [Google Scholar] [CrossRef] [PubMed]
- Karakhanova, S.; Link, J.; Heinrich, M.; Shevchenko, I.; Yang, Y.; Hassenpflug, M.; Bunge, H.; von Ahn, K.; Brecht, R.; Mathes, A.; et al. Characterization of myeloid leukocytes and soluble mediators in pancreatic cancer: Importance of myeloid-derived suppressor cells. Oncoimmunology 2015, 4, e998519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, C.; Sevko, A.; Ramacher, M.; Bazhin, A.V.; Falk, C.S.; Osen, W.; Borrello, I.; Kato, M.; Schadendorf, D.; Baniyash, M.; et al. Chronic inflammation promotes myeloid-derived suppressor cell activation blocking antitumor immunity in transgenic mouse melanoma model. Proc. Natl. Acad. Sci. USA 2011, 108, 17111–17116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serafini, P.; Meckel, K.; Kelso, M.; Noonan, K.; Califano, J.; Koch, W.; Dolcetti, L.; Bronte, V.; Borrello, I. Phosphodiesterase-5 inhibition augments endogenous antitumor immunity by reducing myeloid-derived suppressor cell function. J. Exp. Med. 2006, 203, 2691–2702. [Google Scholar] [CrossRef]
- Califano, J.A.; Khan, Z.; Noonan, K.A.; Rudraraju, L.; Zhang, Z.; Wang, H.; Goodman, S.; Gourin, C.G.; Marshall, L.; Fakhry, C.; et al. Tadalafil Augments Tumor Specific Immunity in Patients with Head and Neck Squamous Cell Carcinoma. Clin. Cancer Res. 2015, 21, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Hassel, J.C.; Jiang, H.; Bender, C.; Winkler, J.; Sevko, A.; Shevchenko, I.; Halama, N.; Dimitrakopoulou-Strauss, A.; Haefeli, W.E.; Jäger, D.; et al. Tadalafil has biologic activity in human melanoma. Results of a pilot trial with Tadalafil in patients with metastatic Melanoma (TaMe). OncoImmunology 2017, 6, e1326440. [Google Scholar] [CrossRef] [Green Version]
- Tai, L.-H.; Alkayyal, A.A.; Leslie, A.L.; Sahi, S.; Bennett, S.; de Souza, C.T.; Baxter, K.; Angka, L.; Xu, R.; Kennedy, M.A.; et al. Phosphodiesterase-5 inhibition reduces postoperative metastatic disease by targeting surgery-induced myeloid derived suppressor cell-dependent inhibition of natural killer cell cytotoxicity. OncoImmunology 2018, 7, e1431082. [Google Scholar] [CrossRef]
- Weed, D.T.; Vella, J.L.; Reis, I.M.; de La Fuente, A.C.; Gomez, C.; Sargi, Z.; Nazarian, R.; Califano, J.; Borrello, I.; Serafini, P. Tadalafil Reduces Myeloid-Derived Suppressor Cells and Regulatory T Cells and Promotes Tumor Immunity in Patients with Head and Neck Squamous Cell Carcinoma. Clin. Cancer Res. 2015, 21, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Meyer, C.; Cagnon, L.; Costa-Nunes, C.M.; Baumgaertner, P.; Montandon, N.; Leyvraz, L.; Michielin, O.; Romano, E.; Speiser, D.E. Frequencies of circulating MDSC correlate with clinical outcome of melanoma patients treated with ipilimumab. Cancer Immunol. Immunother. 2014, 63, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Tarhini, A.A.; Edington, H.; Butterfield, L.H.; Lin, Y.; Shuai, Y.; Tawbi, H.; Sander, C.; Yin, Y.; Holtzman, M.; Johnson, J.; et al. Immune Monitoring of the Circulation and the Tumor Microenvironment in Patients with Regionally Advanced Melanoma Receiving Neoadjuvant Ipilimumab. PLoS ONE 2014, 9, e87705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiktorin, H.G.; Nilsson, M.S.; Kiffin, R.; Sander, F.E.; Lenox, B.; Rydström, A.; Hellstrand, K.; Martner, A. Histamine targets myeloid-derived suppressor cells and improves the anti-tumor efficacy of PD-1/PD-L1 checkpoint blockade. Cancer Immunol. Immunother. 2019, 68, 163–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, H.; Cai, P.; Li, Q.; Wang, W.; Sun, Y.; Xu, Q.; Gu, Y. Axitinib augments antitumor activity in renal cell carcinoma via STAT3-dependent reversal of myeloid-derived suppressor cell accumulation. Biomed. Pharmacother. 2014, 68, 751–756. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phenotypic surface markers and functional subsets of myeloid cells in humans. Venn diagram shows typical cellular markers of MDSC, M1, and M2 macrophages. Markers shared between cell types are shown in overlapping areas of the diagram.
Figure 1.
Phenotypic surface markers and functional subsets of myeloid cells in humans. Venn diagram shows typical cellular markers of MDSC, M1, and M2 macrophages. Markers shared between cell types are shown in overlapping areas of the diagram.

Figure 2.
Suppressive mechanisms exerted by myeloid cells in the tumor microenvironment (TME) and their effects on its cellular components. Green arrows indicate stimulatory effects and red bar headed lines indicate inhibitory effects on targets.
Figure 2.
Suppressive mechanisms exerted by myeloid cells in the tumor microenvironment (TME) and their effects on its cellular components. Green arrows indicate stimulatory effects and red bar headed lines indicate inhibitory effects on targets.

Table 1.
Subsets of myeloid cells in mouse and human. Table shows minimal markers for the phenotyping of subsets in macrophages and MDSC in mouse and human.
Table 1.
Subsets of myeloid cells in mouse and human. Table shows minimal markers for the phenotyping of subsets in macrophages and MDSC in mouse and human.
| Cell Type | Mouse | Human |
|---|---|---|
| Macrophages | F4/80+ CD11b+ M1: CD80high CD206mid M2/TAM: CD80mid CD206high | CD68+ CD11b+ CD14+ M1: HLA-DR+ CD80+ CD86+ M2/TAM: CD206+ CD163+ |
| MDSC | Gr1+ CD11b+ MHC class IIlow/− PMN-MDSC: CD11b+ Ly6Clo Ly6G+ M-MDSC: CD11b+ Ly6Chi Ly6G− | No general MDSC marker PMN-MDSC: CD14− CD11b+ CD15+ (or CD66b+) LOX-1+ M-MDSC: CD11b+ CD14+ HLA-DRlo/− CD15− e-MDSC: Lin−(CD3/14/15/19/56) HLA-DR− CD33+ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Frosch, J.; Leontari, I.; Anderson, J. Combined Effects of Myeloid Cells in the Neuroblastoma Tumor Microenvironment. Cancers 2021, 13, 1743. https://doi.org/10.3390/cancers13071743
AMA Style
Frosch J, Leontari I, Anderson J. Combined Effects of Myeloid Cells in the Neuroblastoma Tumor Microenvironment. Cancers. 2021; 13(7):1743. https://doi.org/10.3390/cancers13071743
Chicago/Turabian StyleFrosch, Jennifer, Ilia Leontari, and John Anderson. 2021. "Combined Effects of Myeloid Cells in the Neuroblastoma Tumor Microenvironment" Cancers 13, no. 7: 1743. https://doi.org/10.3390/cancers13071743
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.