
Deletions on Chromosome Y and Downregulation of the SRY Gene in Tumor Tissue Are Associated with Worse Survival of Glioblastoma Patients

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. IDH1/2 and MGMT Analysis
2.3. Droplet Digital PCR (ddPCR)
2.4. Statistical Analysis
2.5. Gene Set Enrichment Analysis (GSEA)
3. Results
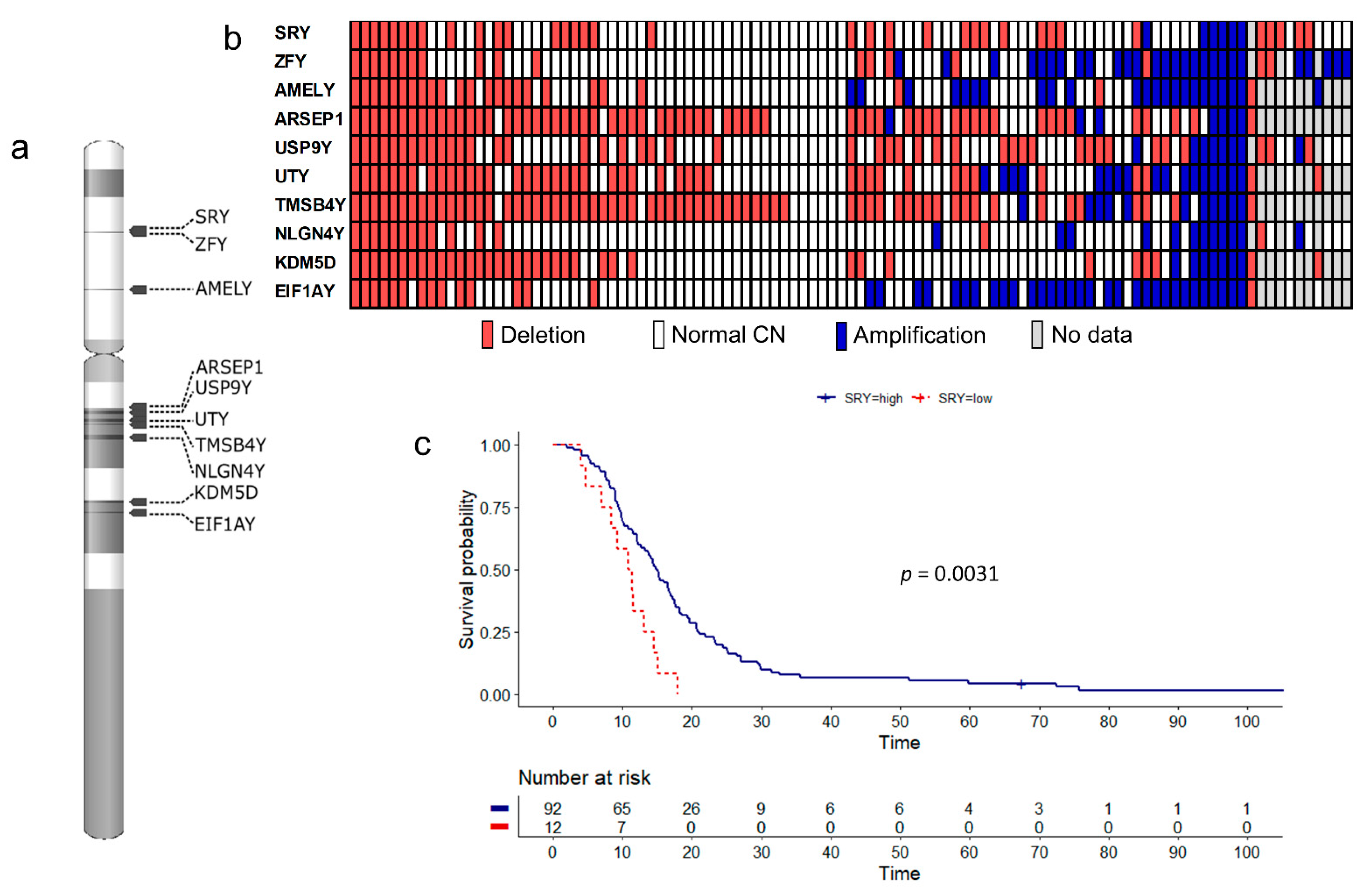
3.1. Copy Number Alterations
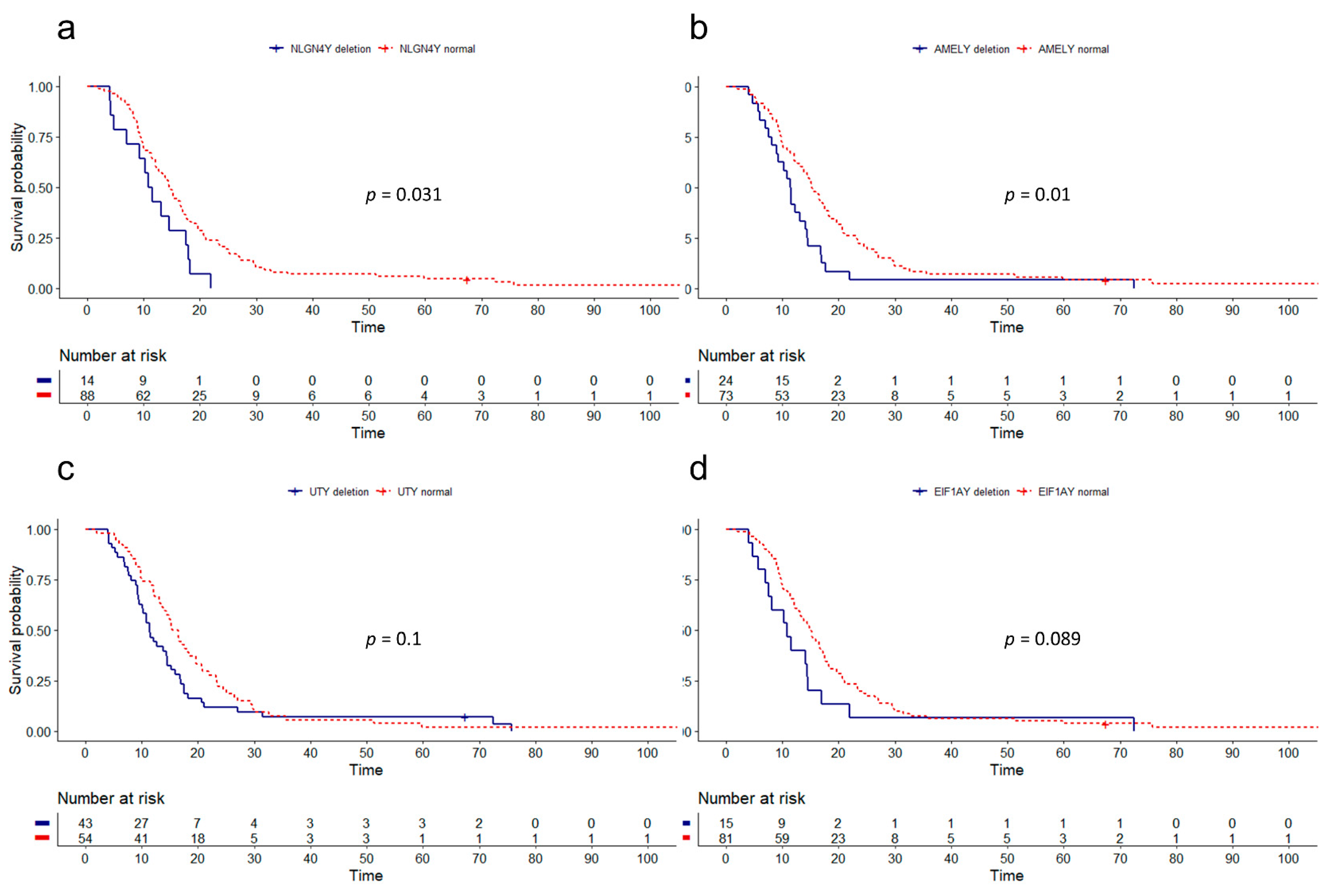
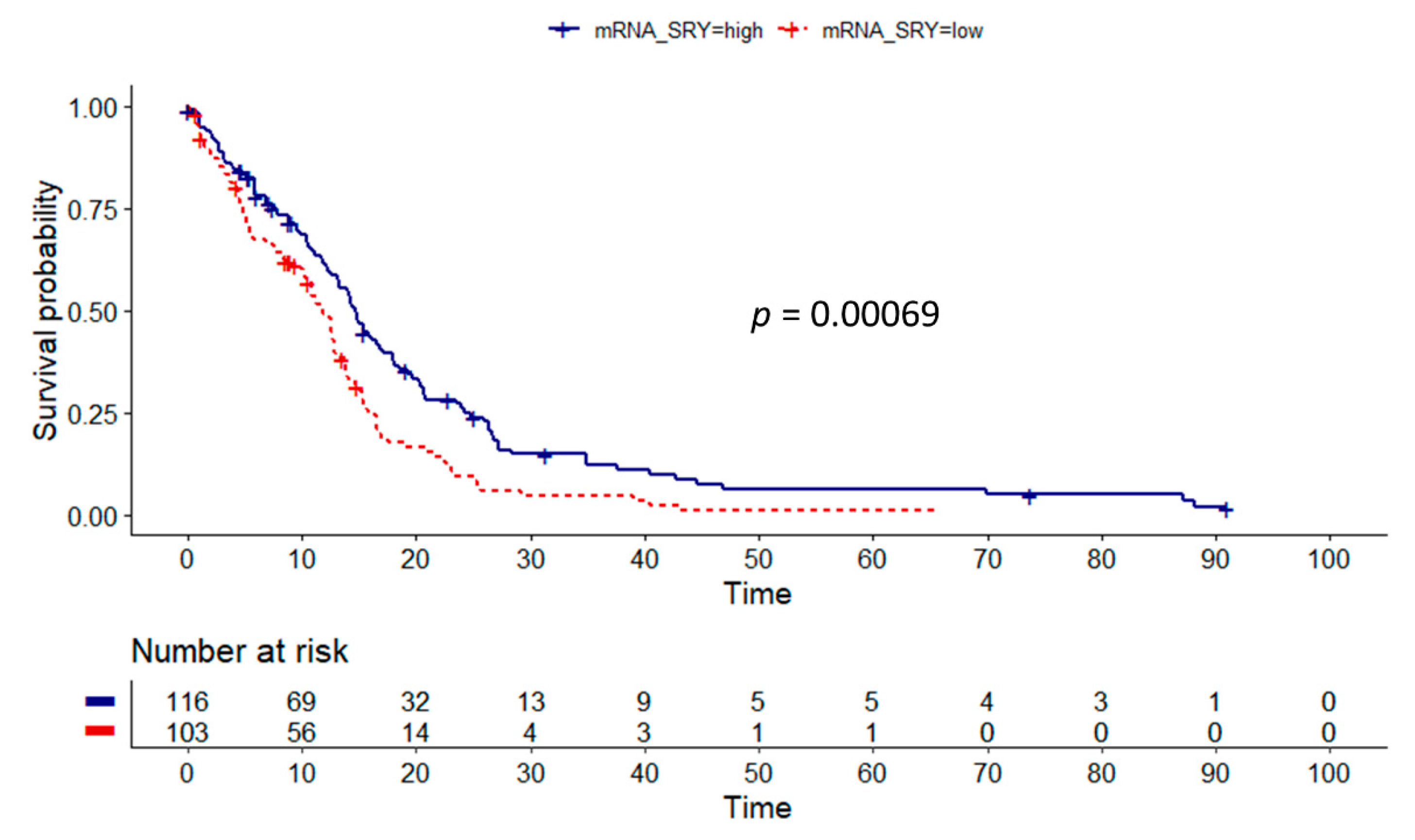
3.2. Influence of Gene Dosage Alterations and mRNA Expression on Survival
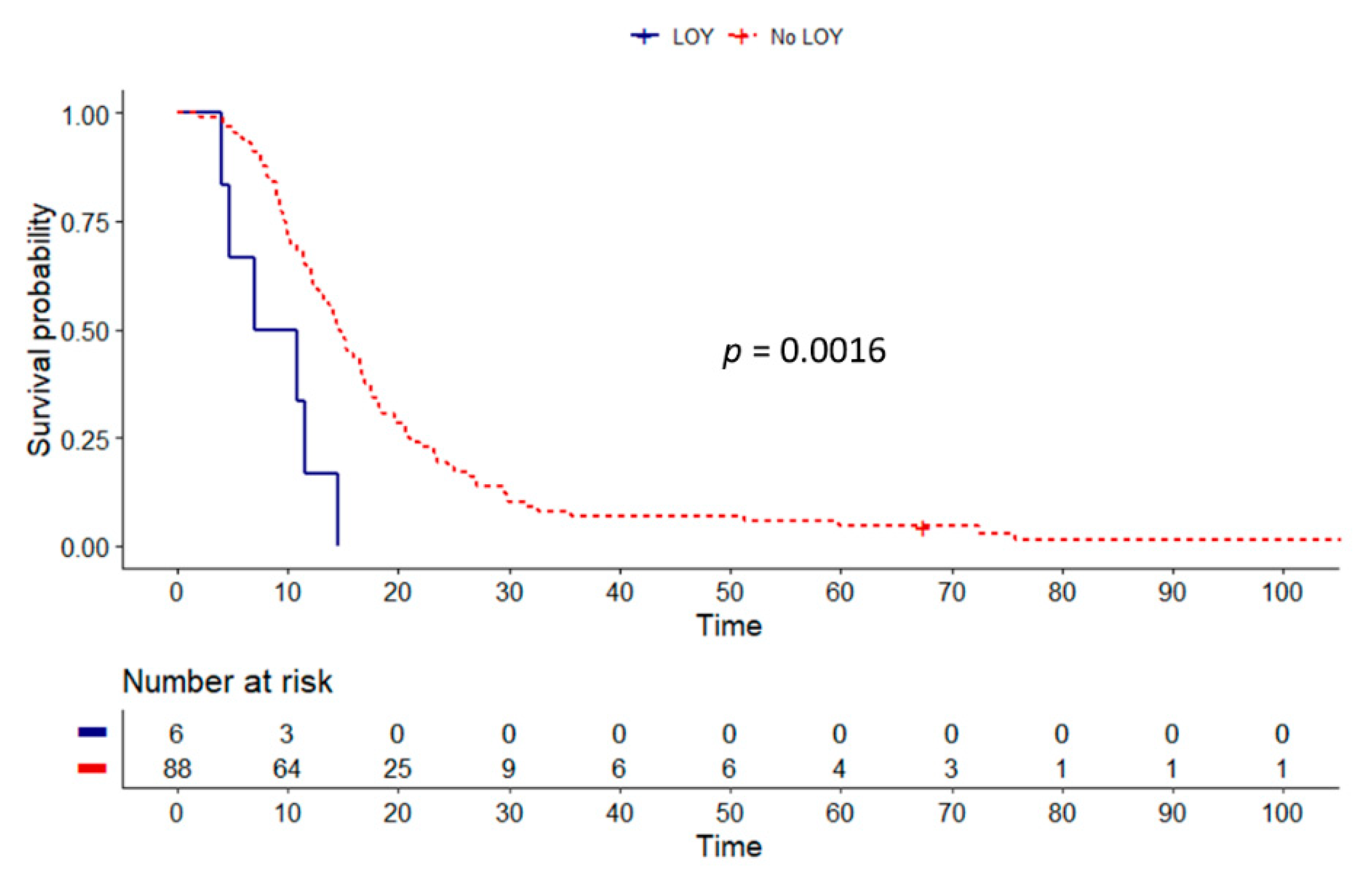
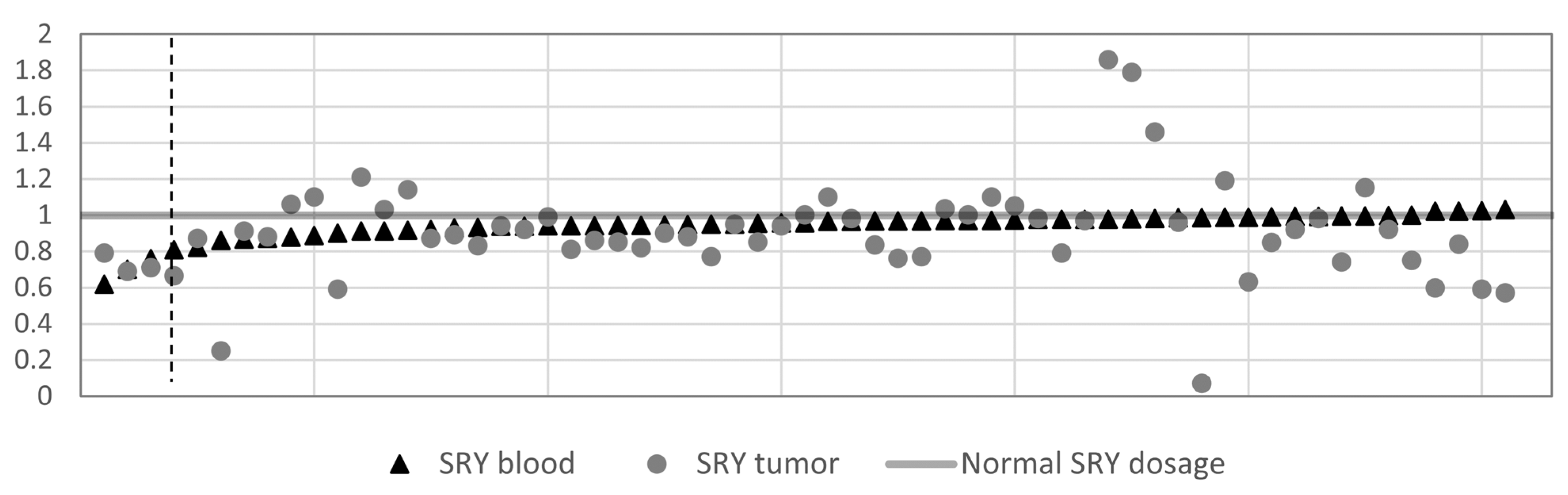
3.3. SRY Loss
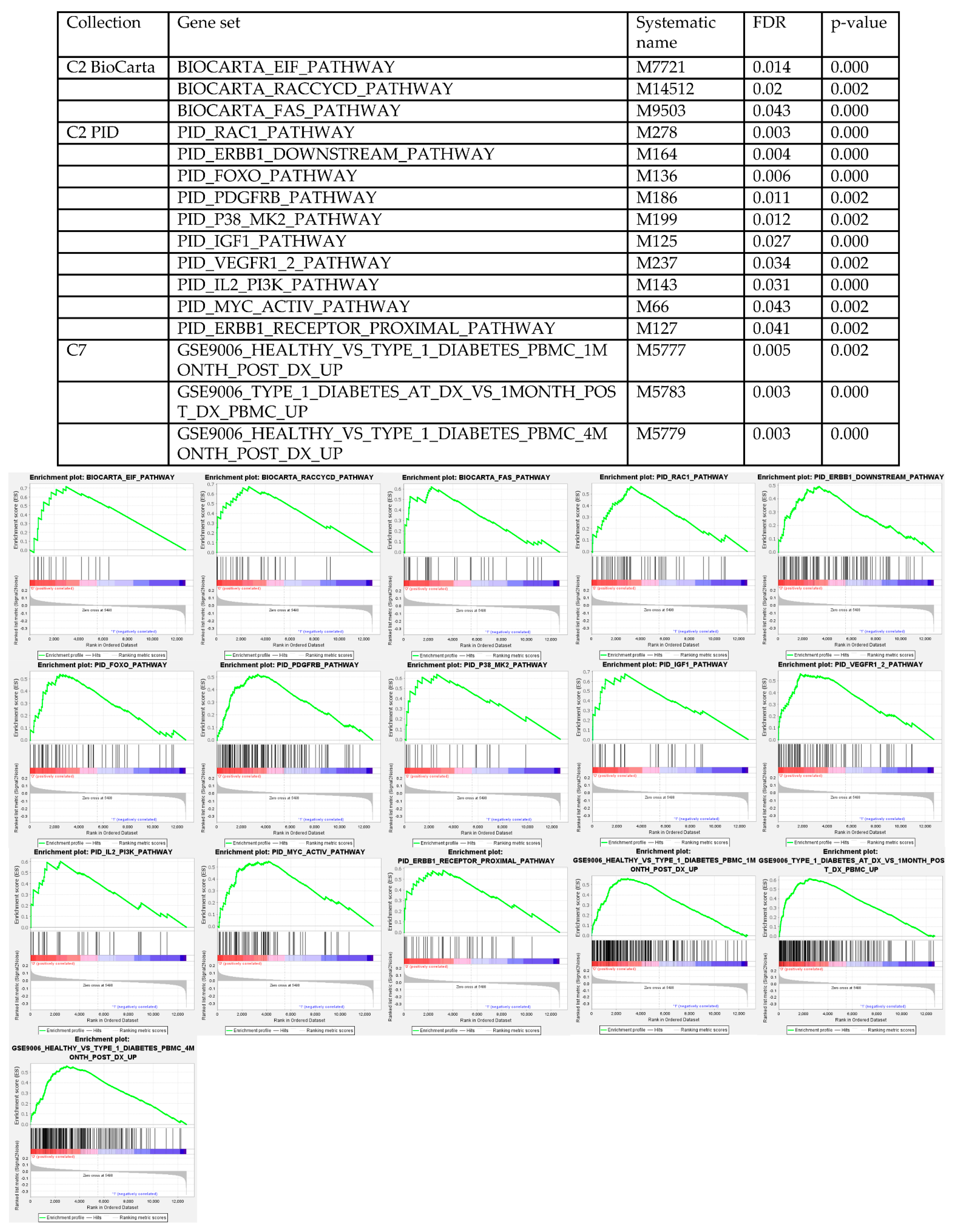
3.4. Gene Set Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GBM | Glioblastoma |
| IDH | Isocitrate dehydrogenase |
| LOY | Loss of chromosome Y |
| TCGA | The Cancer Genome Atlas |
| MGMT | O-6-methylguanine-DNA methyltransferase |
| SRY | Sex determining region Y gene |
| ddPCR | Droplet digital PCR |
| OS | Overall survival |
| CN | Copy number |
| GSEA | Gene Set Enrichment Analysis |
| ZFY | Zinc finger protein Y-linked |
| AMELY | Amelogenin Y-linked |
| ARSEP1 | Arylsulfatase L pseudogene 1 |
| USP9Y | Ubiquitin specific peptidase 9 Y-Linked |
| UTY | Ubiquitously transcribed tetratricopeptide repeat containing, Y-Linked |
| TMSB4Y | Thymosin beta 4 Y-linked |
| NLGN4Y | Neuroligin 4 Y-linked |
| KDM5D | Lysine demethylase 5D |
| EIF1AY | Eukaryotic translation initiation factor 1A Y-linked |
| LGG | Lower grade gliomas |
| c-Myc | MYC proto-oncogene |
| ErbB | Epidermal growth factor receptor family |
| EGFR | Epidermal growth factor receptor |
| PDGFR | Platelet derived growth factor receptor |
| VEGFR1/2 | Vascular endothelial growth factor receptor 1 and 2 |
| PI3K | Phosphoinositide 3 kinase |
| Akt | Protein kinase B |
| IGF1 | Insulin like growth factor 1 |
| eIF | Eukaryotic translation initiation factor |
| Fas | Fas cell surface death receptor |
| Rac1 | Rac family small GTPase 1 |
| FOXO | Forkhead box O1 |
References
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molinaro, A.M.; Taylor, J.W.; Wiencke, J.K.; Wrensch, M.R. Genetic and molecular epidemiology of adult diffuse glioma. Nat. Rev. Neurol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Radkiewicz, C.; Johansson, A.L.V.; Dickman, P.W.; Lambe, M.; Edgren, G. Sex differences in cancer risk and survival: A Swedish cohort study. Eur. J. Cancer 2017, 84, 130–140. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Gittleman, H.; Truitt, G.; Boscia, A.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2011-2015. Neuro Oncol. 2018, 20, iv1–iv86. [Google Scholar] [CrossRef] [Green Version]
- Gittleman, H.; Ostrom, Q.T.; Stetson, L.C.; Waite, K.; Hodges, T.R.; Wright, C.H.; Wright, J.; Rubin, J.B.; Berens, M.E.; Lathia, J.; et al. Sex is an important prognostic factor for glioblastoma but not for nonglioblastoma. Neurooncol. Pract. 2019, 6, 451–462. [Google Scholar] [CrossRef]
- Dorak, M.T.; Karpuzoglu, E. Gender differences in cancer susceptibility: An inadequately addressed issue. Front Genet. 2012, 3, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Liu, L.; Chen, H.; Wang, Y.; Xu, Y.; Mao, H.; Li, J.; Mills, G.B.; Shu, Y.; Li, L.; et al. Comprehensive Characterization of Molecular Differences in Cancer between Male and Female Patients. Cancer Cell 2016, 29, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Malladi, S.; Beck, A.H. Systematic Analysis of Sex-Linked Molecular Alterations and Therapies in Cancer. Sci. Rep. 2016, 6, 19119. [Google Scholar] [CrossRef] [Green Version]
- Yamada, K.; Kasama, M.; Kondo, T.; Shinoura, N.; Yoshioka, M. Chromosome studies in 70 brain tumors with special attention to sex chromosome loss and single autosomal trisomy. Cancer Genet. Cytogenet. 1994, 73, 46–52. [Google Scholar] [CrossRef]
- Lindstrom, E.; Salford, L.G.; Heim, S.; Mandahl, N.; Stromblad, S.; Brun, A.; Mitelman, F. Trisomy 7 and sex chromosome loss need not be representative of tumor parenchyma cells in malignant glioma. Genes Chromosom. Cancer 1991, 3, 474–479. [Google Scholar] [CrossRef]
- Hecht, B.K.; Turc-Carel, C.; Chatel, M.; Paquis, P.; Gioanni, J.; Attias, R.; Gaudray, P.; Hecht, F. Cytogenetics of malignant gliomas. II. The sex chromosomes with reference to X isodisomy and the role of numerical X/Y changes. Cancer Genet. Cytogenet. 1995, 84, 9–14. [Google Scholar] [CrossRef]
- Forsberg, L.A.; Rasi, C.; Malmqvist, N.; Davies, H.; Pasupulati, S.; Pakalapati, G.; Sandgren, J.; Diaz de Stahl, T.; Zaghlool, A.; Giedraitis, V.; et al. Mosaic loss of chromosome Y in peripheral blood is associated with shorter survival and higher risk of cancer. Nat. Genet. 2014, 46, 624–628. [Google Scholar] [CrossRef] [Green Version]
- Loftfield, E.; Zhou, W.; Graubard, B.I.; Yeager, M.; Chanock, S.J.; Freedman, N.D.; Machiela, M.J. Predictors of mosaic chromosome Y loss and associations with mortality in the UK Biobank. Sci. Rep. 2018, 8, 12316. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Machiela, M.J.; Freedman, N.D.; Rothman, N.; Malats, N.; Dagnall, C.; Caporaso, N.; Teras, L.T.; Gaudet, M.M.; Gapstur, S.M.; et al. Mosaic loss of chromosome Y is associated with common variation near TCL1A. Nat. Genet. 2016, 48, 563–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burki, T.K. Smoking and mosaic Y chromosome loss. Lancet Oncol. 2015, 16, e12. [Google Scholar] [CrossRef]
- Dumanski, J.P.; Lambert, J.C.; Rasi, C.; Giedraitis, V.; Davies, H.; Grenier-Boley, B.; Lindgren, C.M.; Campion, D.; Dufouil, C.; European Alzheimer’s Disease Initiative Investigators; et al. Mosaic Loss of Chromosome Y in Blood Is Associated with Alzheimer Disease. Am. J. Hum. Genet. 2016, 98, 1208–1219. [Google Scholar] [CrossRef] [Green Version]
- Wright, D.J.; Day, F.R.; Kerrison, N.D.; Zink, F.; Cardona, A.; Sulem, P.; Thompson, D.J.; Sigurjonsdottir, S.; Gudbjartsson, D.F.; Helgason, A.; et al. Genetic variants associated with mosaic Y chromosome loss highlight cell cycle genes and overlap with cancer susceptibility. Nat. Genet. 2017, 49, 674–679. [Google Scholar] [CrossRef] [PubMed]
- Machiela, M.J.; Dagnall, C.L.; Pathak, A.; Loud, J.T.; Chanock, S.J.; Greene, M.H.; McGlynn, K.A.; Stewart, D.R. Mosaic chromosome Y loss and testicular germ cell tumor risk. J. Hum. Genet. 2017, 62, 637–640. [Google Scholar] [CrossRef] [Green Version]
- Thompson, D.J.; Genovese, G.; Halvardson, J.; Ulirsch, J.C.; Wright, D.J.; Terao, C.; Davidsson, O.B.; Day, F.R.; Sulem, P.; Jiang, Y.; et al. Genetic predisposition to mosaic Y chromosome loss in blood. Nature 2019, 575, 652–657. [Google Scholar] [CrossRef]
- Bowman, R.L.; Wang, Q.; Carro, A.; Verhaak, R.G.; Squatrito, M. GlioVis data portal for visualization and analysis of brain tumor expression datasets. Neuro Oncol. 2017, 19, 139–141. [Google Scholar] [CrossRef] [Green Version]
- GlioVis Data Visualization Tools for Brain Tumor Datasets. GlioVis Explore. Available online: http://gliovis.bioinfo.cnio.es/ (accessed on 11 February 2011).
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hothorn, T.; Lausen, B. On the exact distribution of maximally selected rank statistics. Comput. Stat. Data Anal. 2003, 43, 121–137. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstrale, M.; Laurila, E.; et al. PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Gene Set Enrichment Analysis, M.S.D. Molecular Signatures Database v7.2. Available online: https://www.gsea-msigdb.org/gsea/msigdb (accessed on 5 November 2020).
- Matteoni, S.; Abbruzzese, C.; Villani, V.; Malorni, W.; Pace, A.; Matarrese, P.; Paggi, M.G. The influence of patient sex on clinical approaches to malignant glioma. Cancer Lett. 2020, 468, 41–47. [Google Scholar] [CrossRef]
- Zhang, H.; Liao, J.; Zhang, X.; Zhao, E.; Liang, X.; Luo, S.; Shi, J.; Yu, F.; Xu, J.; Shen, W.; et al. Sex difference of mutation clonality in diffuse glioma evolution. Neuro Oncol. 2019, 21, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Warrington, N.M.; Taylor, S.J.; Whitmire, P.; Carrasco, E.; Singleton, K.W.; Wu, N.; Lathia, J.D.; Berens, M.E.; Kim, A.H.; et al. Sex differences in GBM revealed by analysis of patient imaging, transcriptome, and survival data. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef] [Green Version]
- Ippolito, J.E.; Yim, A.K.; Luo, J.; Chinnaiyan, P.; Rubin, J.B. Sexual dimorphism in glioma glycolysis underlies sex differences in survival. JCI Insight 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiffgens, S.; Wilkens, L.; Brandes, A.A.; Meier, T.; Franceschi, E.; Ermani, M.; Hartmann, C.; Sandalcioglu, I.E.; Dumitru, C.A. Sex-specific clinicopathological significance of novel (Frizzled-7) and established (MGMT, IDH1) biomarkers in glioblastoma. Oncotarget 2016, 7, 55169–55180. [Google Scholar] [CrossRef] [Green Version]
- Hollows, R.; Wei, W.; Cazier, J.B.; Mehanna, H.; Parry, G.; Halford, G.; Murray, P. Association between loss of Y chromosome and poor prognosis in male head and neck squamous cell carcinoma. Head Neck 2019, 41, 993–1006. [Google Scholar] [CrossRef] [Green Version]
- Hirata, T.; Hishimoto, A.; Otsuka, I.; Okazaki, S.; Boku, S.; Kimura, A.; Horai, T.; Sora, I. Investigation of chromosome Y loss in men with schizophrenia. Neuropsychiatr. Dis. Treat. 2018, 14, 2115–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomdedeu, M.; Pereira, A.; Calvo, X.; Colomer, J.; Sole, F.; Arias, A.; Gomez, C.; Luno, E.; Cervera, J.; Arnan, M.; et al. Clinical and biological significance of isolated Y chromosome loss in myelodysplastic syndromes and chronic myelomonocytic leukemia. A report from the Spanish MDS Group. Leuk. Res. 2017, 63, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Caceres, A.; Jene, A.; Esko, T.; Perez-Jurado, L.A.; Gonzalez, J.R. Extreme down-regulation of chromosome Y and cancer risk in men. J. Natl. Cancer Inst. 2020. [Google Scholar] [CrossRef] [PubMed]
- Danielsson, M.; Halvardson, J.; Davies, H.; Torabi Moghadam, B.; Mattisson, J.; Rychlicka-Buniowska, E.; Jaszczynski, J.; Heintz, J.; Lannfelt, L.; Giedraitis, V.; et al. Longitudinal changes in the frequency of mosaic chromosome Y loss in peripheral blood cells of aging men varies profoundly between individuals. Eur. J. Hum. Genet. 2020, 28, 349–357. [Google Scholar] [CrossRef]
- Guo, X.; Dai, X.; Zhou, T.; Wang, H.; Ni, J.; Xue, J.; Wang, X. Mosaic loss of human Y chromosome: What, how and why. Hum. Genet. 2020, 139, 421–446. [Google Scholar] [CrossRef]
- Malmstrom, A.; Gronberg, B.H.; Marosi, C.; Stupp, R.; Frappaz, D.; Schultz, H.; Abacioglu, U.; Tavelin, B.; Lhermitte, B.; Hegi, M.E.; et al. Temozolomide versus standard 6-week radiotherapy versus hypofractionated radiotherapy in patients older than 60 years with glioblastoma: The Nordic randomised, phase 3 trial. Lancet Oncol. 2012, 13, 916–926. [Google Scholar] [CrossRef]
- Liu, C.; Ren, Y.F.; Dong, J.; Ke, M.Y.; Ma, F.; Monga, S.P.S.; Wu, R.; Lv, Y.; Zhang, X.F. Activation of SRY accounts for male-specific hepatocarcinogenesis: Implication in gender disparity of hepatocellular carcinoma. Cancer Lett. 2017, 410, 20–31. [Google Scholar] [CrossRef]
- Murakami, S.; Chishima, S.; Uemoto, H.; Sakamoto, E.; Sato, T.; Kurabe, N.; Kawasaki, Y.; Shibata, T.; Akiyama, H.; Tashiro, F. The male-specific factor Sry harbors an oncogenic function. Oncogene 2014, 33, 2978–2986. [Google Scholar] [CrossRef] [Green Version]
- Kido, T.; Sun, Z.; Lau, Y.C. Aberrant activation of the human sex-determining gene in early embryonic development results in postnatal growth retardation and lethality in mice. Sci. Rep. 2017, 7, 4113. [Google Scholar] [CrossRef]
- Taleahmad, S.; Alikhani, M.; Mollamohammadi, S.; Yousefi, M.; Taei, A.; Hassani, S.N.; Baharvand, H.; Salekdeh, G.H. Inhibition of Human Y Chromosome Gene, SRY, Promotes Naive State of Human Pluripotent Stem Cells. J. Proteome. Res. 2019, 18, 4254–4261. [Google Scholar] [CrossRef]
- Rajakulendran, N.; Rowland, K.J.; Selvadurai, H.J.; Ahmadi, M.; Park, N.I.; Naumenko, S.; Dolma, S.; Ward, R.J.; So, M.; Lee, L.; et al. Wnt and Notch signaling govern self-renewal and differentiation in a subset of human glioblastoma stem cells. Genes Dev. 2019, 33, 498–510. [Google Scholar] [CrossRef]
- Verhaak, R.G.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef] [Green Version]
- Kodric, K.; Zupan, J.; Kranjc, T.; Komadina, R.; Mlakar, V.; Marc, J.; Lovsin, N. Sex-determining region Y (SRY) attributes to gender differences in RANKL expression and incidence of osteoporosis. Exp. Mol. Med. 2019, 51, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.K.; Jin, X.; Sohn, Y.W.; Jin, X.; Jeon, H.Y.; Kim, E.J.; Ham, S.W.; Jeon, H.M.; Chang, S.Y.; Oh, S.Y.; et al. Tumoral RANKL activates astrocytes that promote glioma cell invasion through cytokine signaling. Cancer Lett. 2014, 353, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Machiela, M.J.; Zhou, W.; Karlins, E.; Sampson, J.N.; Freedman, N.D.; Yang, Q.; Hicks, B.; Dagnall, C.; Hautman, C.; Jacobs, K.B.; et al. Female chromosome X mosaicism is age-related and preferentially affects the inactivated X chromosome. Nat. Commun. 2016, 7, 11843. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.M.; Lundin, E.; Qian, X.; Mirzazadeh, M.; Halvardson, J.; Darj, E.; Feuk, L.; Nilsson, M.; Jazin, E. Spatial sexual dimorphism of X and Y homolog gene expression in the human central nervous system during early male development. Biol. Sex Differ. 2016, 7, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bramble, M.S.; Vashist, N.; Vilain, E. Sex steroid hormone modulation of neural stem cells: A critical review. Biol. Sex Differ. 2019, 10, 28. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Lee, J.E.; Kahng, J.Y.; Kim, S.H.; Park, J.S.; Yoon, S.J.; Um, J.Y.; Kim, W.K.; Lee, J.K.; Park, J.; et al. Human glioblastoma arises from subventricular zone cells with low-level driver mutations. Nature 2018, 560, 243–247. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Category | Number of Patients | Percentage |
|---|---|---|---|
| Age, years | ≤60 | 64 | 61% |
| >60 | 41 | 39% | |
| Surgery | Biopsy | 15 | 14% |
| Partial resection | 24 | 23% | |
| Large total resection | 66 | 63% | |
| Preoperative performance status WHO | 0–1 | 83 | 79% |
| 2–3 | 19 | 18% | |
| Missing | 3 | 3% | |
| RT dose (Gray) | 34 a | 7 | 6.7% |
| 40–50 b | 8 | 7.6% | |
| 54–60 | 89 | 84.7% | |
| Missing | 1 | 1% | |
| MGMT | Methylated | 36 | 65.7% |
| Unmethylated | 69 | 34.3% | |
| SRY CN | ≤0.6 | 12 | 11.4% |
| >0.6 | 92 | 87.6% | |
| Missing | 1 | 0.95% |
| Variables | Compared Groups | Hazard Ratio (95% Cl) | p-Value |
|---|---|---|---|
| Age | >60 vs. ≤60 | 1.702 (1.1–2.633) | 0.017 * |
| Preoperative performance status (WHO) | 2-3 vs. 0-1 | 0.937 (0.553–1.588) | 0.810 |
| Type of surgery | Biopsy/partial vs. large total | 1.131 (0.736–1.740) | 0.574 |
| MGMT status | Unmethylated vs. methylated | 2.071 (1.302–3.294) | 0.002 * |
| CN of SRY | ≤0.597 vs. >0.597 | 3.352 (1.695–6.629) | 0.001 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Łysiak, M.; Smits, A.; Roodakker, K.R.; Sandberg, E.; Dimberg, A.; Mudaisi, M.; Bratthäll, C.; Strandeus, M.; Milos, P.; Hallbeck, M.; et al. Deletions on Chromosome Y and Downregulation of the SRY Gene in Tumor Tissue Are Associated with Worse Survival of Glioblastoma Patients. Cancers 2021, 13, 1619. https://doi.org/10.3390/cancers13071619
Łysiak M, Smits A, Roodakker KR, Sandberg E, Dimberg A, Mudaisi M, Bratthäll C, Strandeus M, Milos P, Hallbeck M, et al. Deletions on Chromosome Y and Downregulation of the SRY Gene in Tumor Tissue Are Associated with Worse Survival of Glioblastoma Patients. Cancers. 2021; 13(7):1619. https://doi.org/10.3390/cancers13071619
Chicago/Turabian StyleŁysiak, Małgorzata, Anja Smits, Kenney Roy Roodakker, Elisabeth Sandberg, Anna Dimberg, Munila Mudaisi, Charlotte Bratthäll, Michael Strandeus, Peter Milos, Martin Hallbeck, and et al. 2021. "Deletions on Chromosome Y and Downregulation of the SRY Gene in Tumor Tissue Are Associated with Worse Survival of Glioblastoma Patients" Cancers 13, no. 7: 1619. https://doi.org/10.3390/cancers13071619