Marked Increased Production of Acute Phase Reactants by Skeletal Muscle during Cancer Cachexia

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
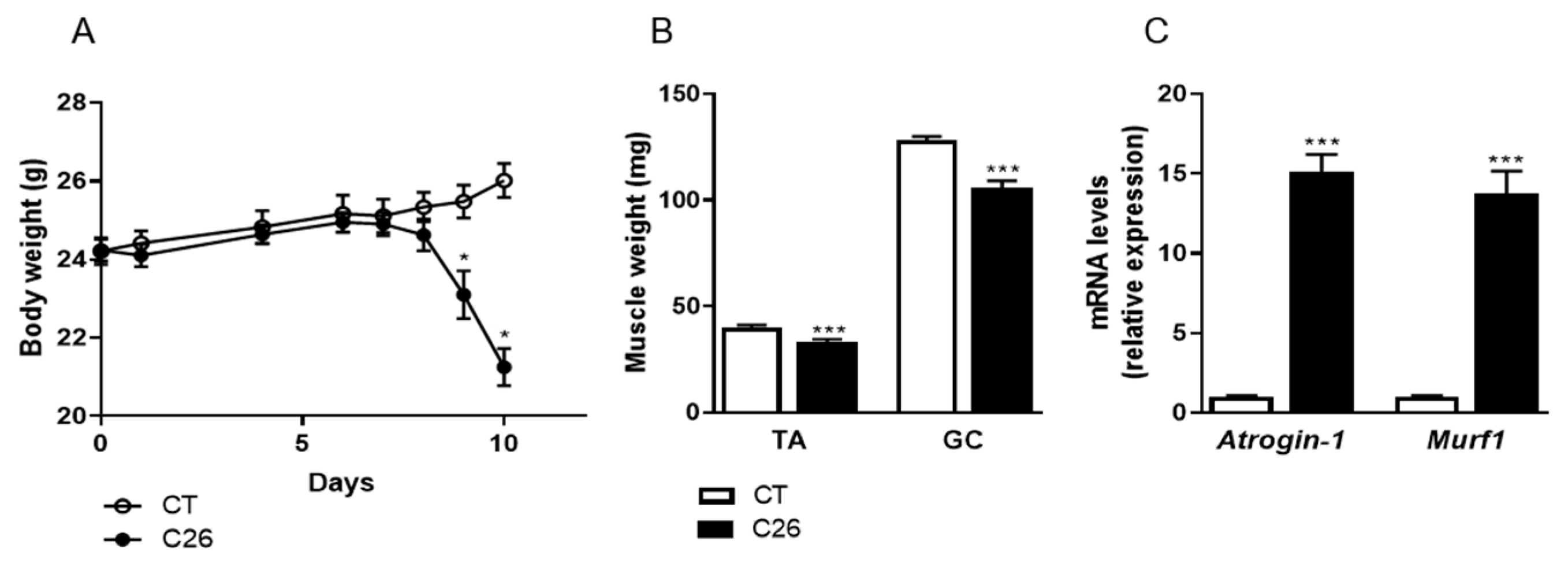
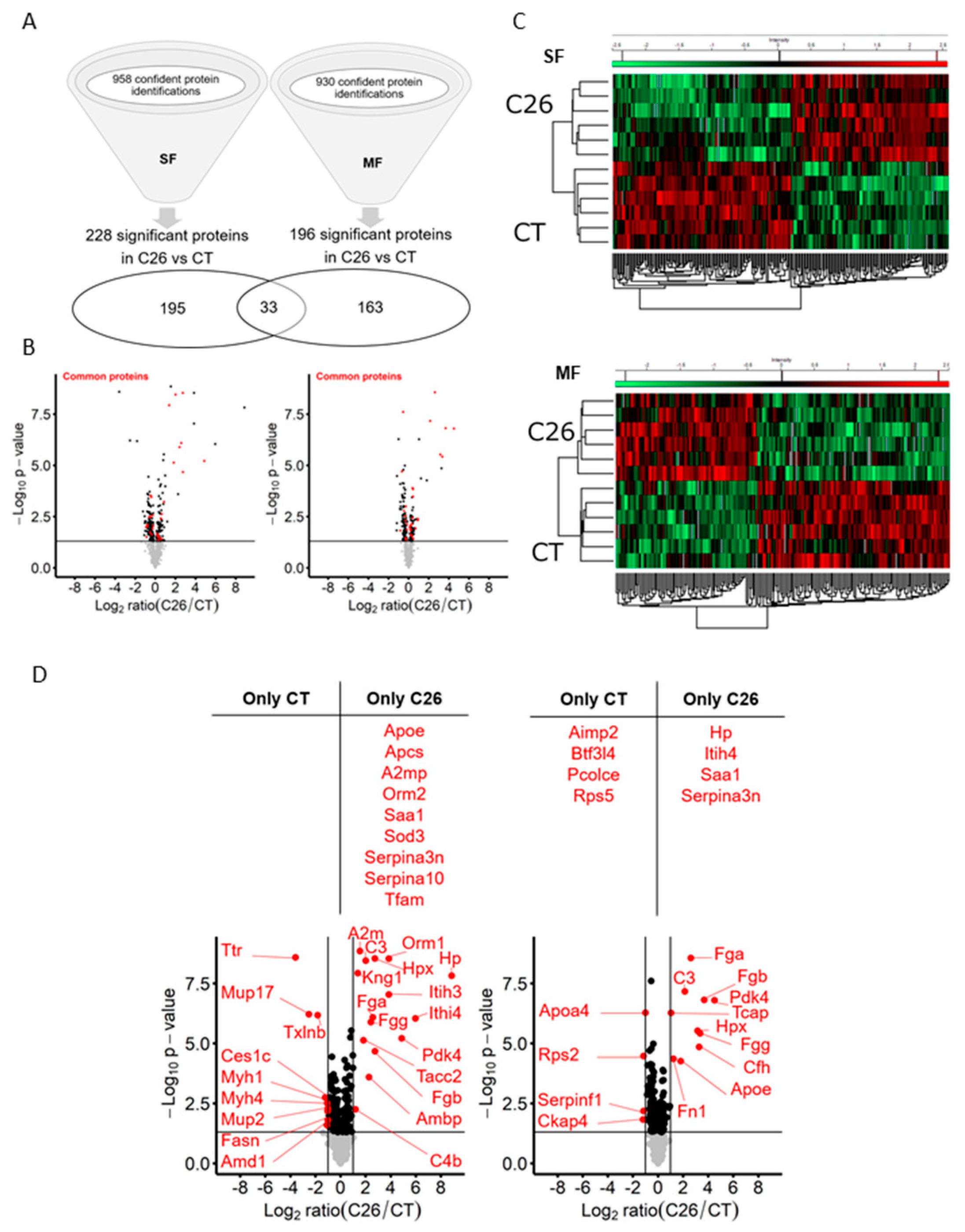
2.1. C26 Colon Carcinoma Induced-Cachexia Is Associated with Profound Changes in the Skeletal Muscle Proteome
2.2. Mitochondrial Dysfunction, Ribosome Depletion, and Acute Phase Response Take Place in the Skeletal Muscle of C26 Mice
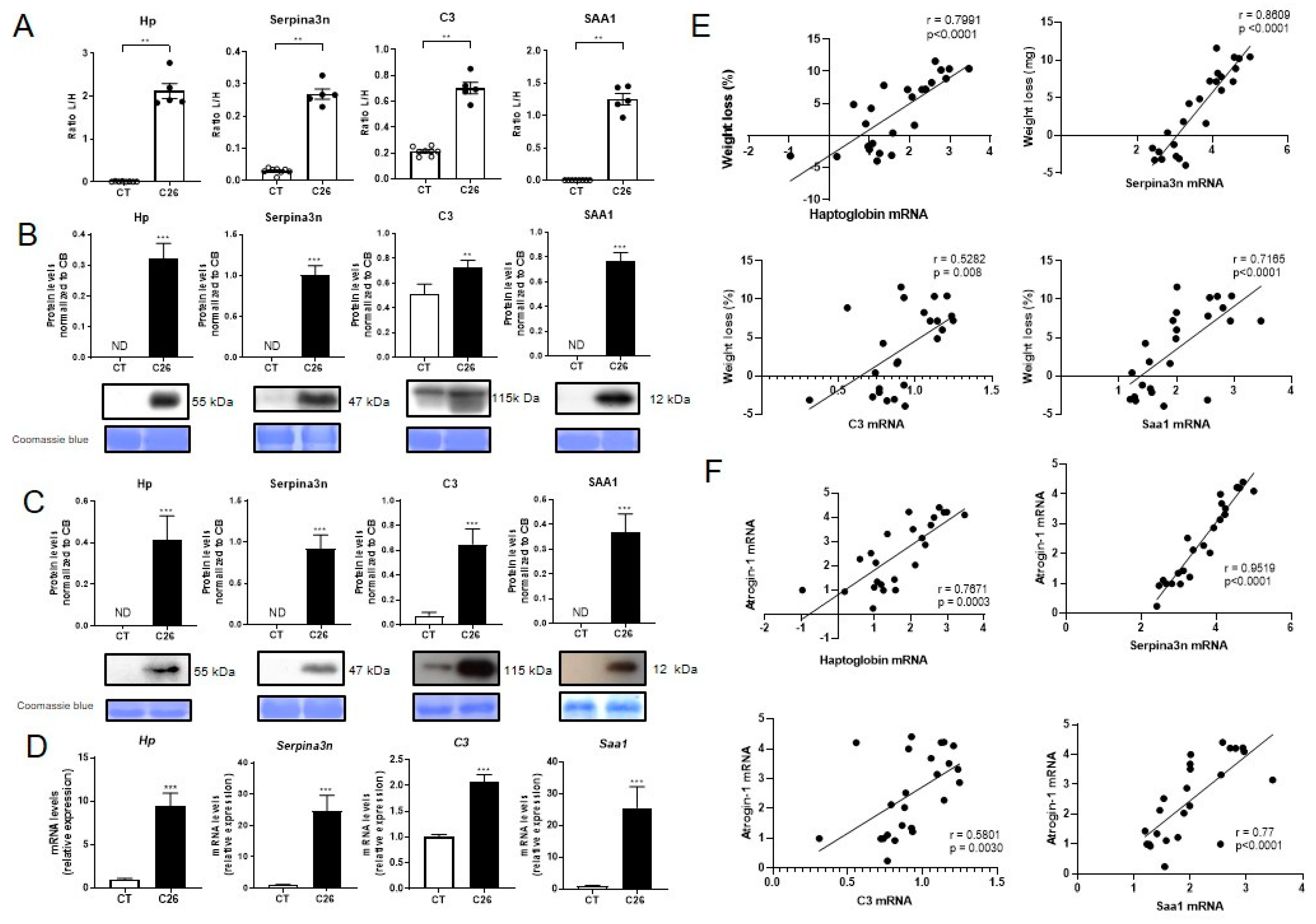
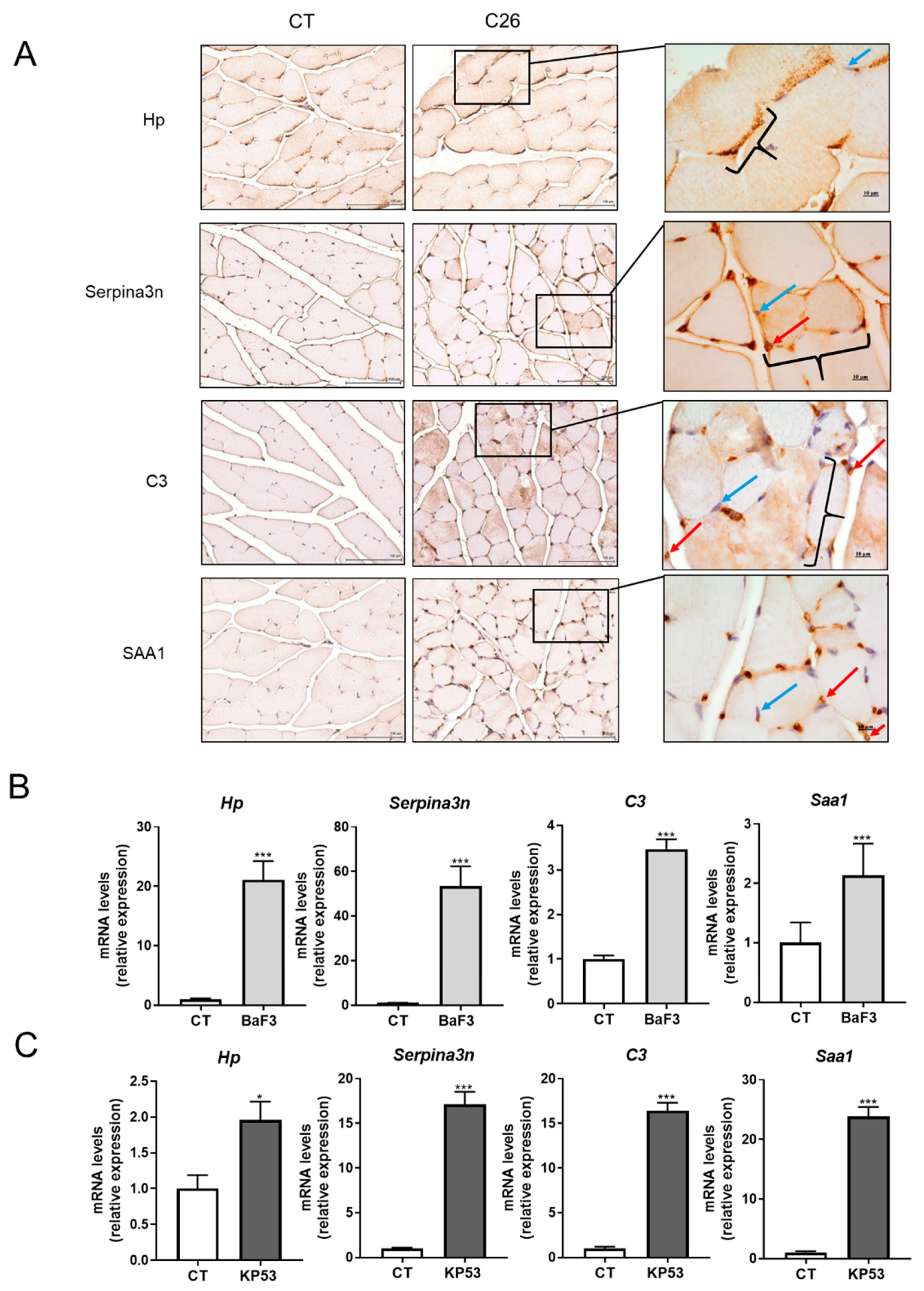
2.3. Skeletal Muscle Is a Source of Several Acute Phase Reactants during Cancer Cachexia
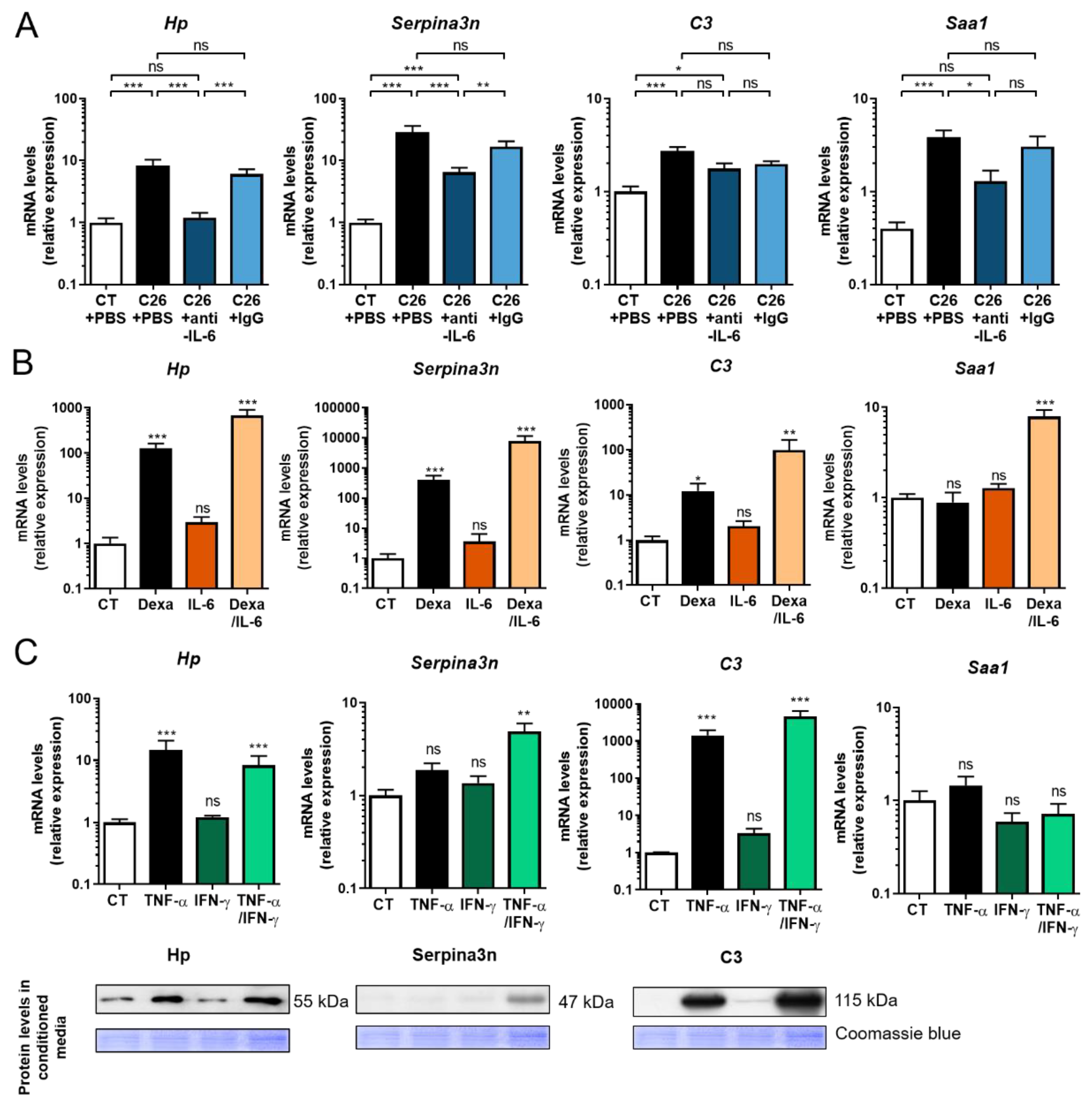
2.4. Anti-IL-6 Antibody Prevents Cancer-Induced Muscular Production of Acute Phase Reactants in C26 Mice
2.5. Glucocorticoids and Proinflammatory Cytokines Stimulate the Production of Acute Phase Reactants by Skeletal Muscle Cells
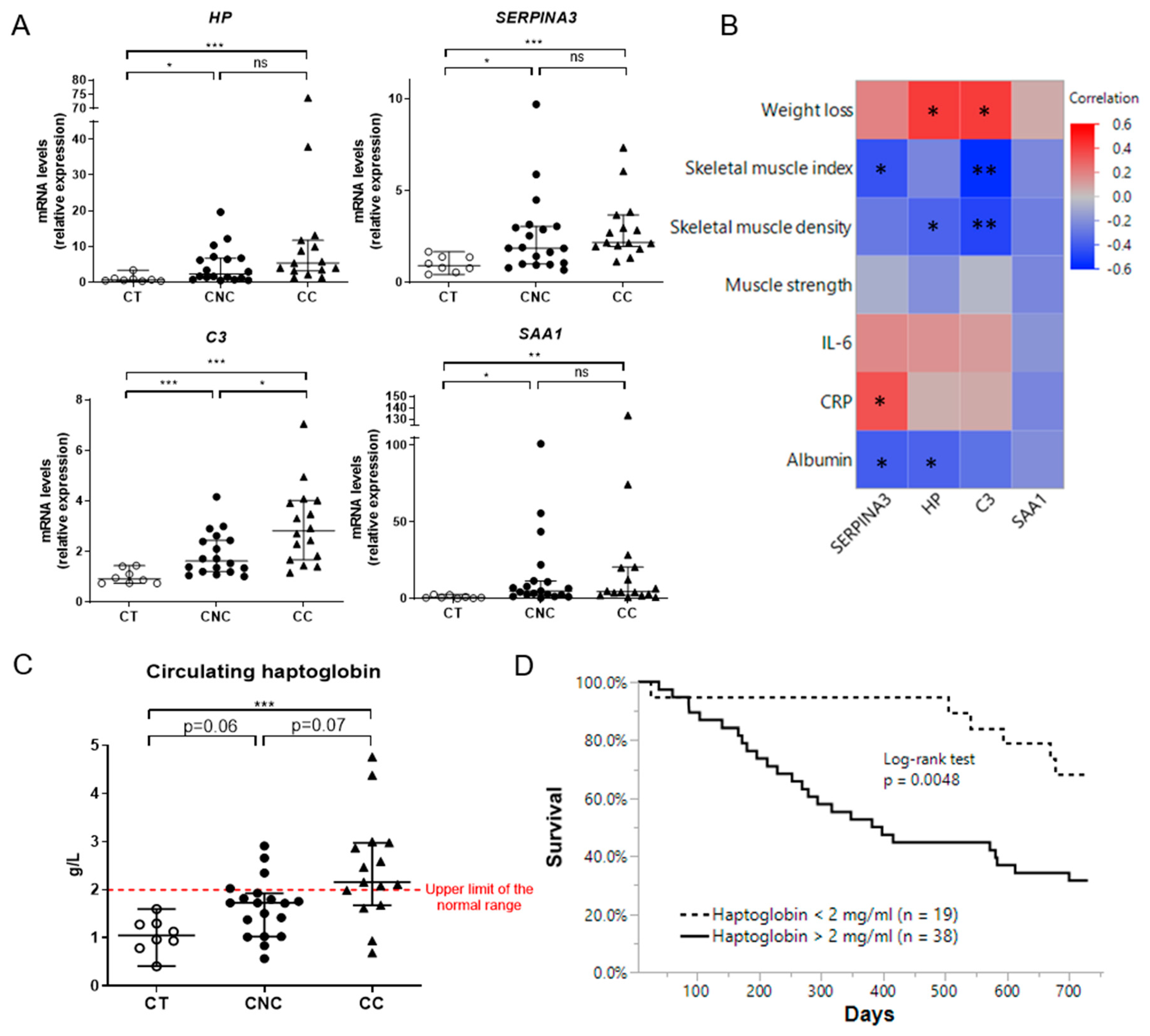
2.6. Muscle Expression of Acute Phase Reactants Is Increased in Cancer Patients
3. Discussion
4. Materials and Methods
4.1. Animals
4.1.1. C26 Mouse Model of Cancer Cachexia
4.1.2. BaF3 Mouse Model of Cancer Cachexia
4.1.3. KP53 Mouse Model of Cancer Cachexia
4.2. Proteomic Discovery Analysis
4.2.1. Sample Preparation, Muscle Protein Extraction, and Pre-Fractionation
4.2.2. Muscle Protein Extract Digestion
4.2.3. Sample Protein Digests Reconditioning and Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS) Analysis
Sample Reconditioning
LC-MS/MS
MS/MS Data Acquisition
Raw Data Analysis
Differential Analysis
4.3. Gene Ontology Analysis and Identification of Enriched Pathways
4.4. Targeted Proteomics Using Multiple Reaction Monitoring
4.5. Putative Secretomic Analysis
4.6. C2C12 Culture
4.7. Western Blot Analyses
4.8. mRNA Analysis by RT-qPCR
4.9. Immunohistochemistry
4.10. Human Samples
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Argiles, J.M.; Busquets, S.; Stemmler, B.; Lopez-Soriano, F.J. Cancer cachexia: Understanding the molecular basis. Nat. Rev. Cancer 2014, 14, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, G.W.P.D.; Farkas, J.; Dora, E.; von Haehling, S.; Lainscak, M. Cancer Cachexia and Related Metabolic Dysfunction. Int. J. Mol. Sci. 2020, 21, 2321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiessel, D.L.; Baracos, V.E. Barriers to cancer nutrition therapy: Excess catabolism of muscle and adipose tissues induced by tumour products and chemotherapy. Proc. Nutr. Soc. 2018, 77, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Fearon, K.F.; Strasser, S.D.; Anker, I.; Bosaeus, E.; Bruera, R.L.; Fainsinger, R.L.; Jatoi, A.; Loprinzi, C.; MacDonald, N.; Mantovani, G.; et al. Definition and classification of cancer cachexia: An international consensus. Lancet Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef]
- Fearon, K.C.; Glass, D.J.; Guttridge, D.C. Cancer cachexia: Mediators, signaling, and metabolic pathways. Cell Metab. 2012, 16, 153–166. [Google Scholar] [CrossRef] [Green Version]
- Klimek, M.E.B.; Aydogdu, T.; Link, M.J.; Pons, M.; Koniaris, L.G.; Zimmers, T.A. Acute inhibition of myostatin-family proteins preserves skeletal muscle in mouse models of cancer cachexia. Biochem. Biophys. Res. Commun. 2010, 391, 1548–1554. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, J.L.; Lu, Y.; Song, K.S.; Kwak, K.S.; Jiao, Q.; Rosenfeld, R.; Chen, Q.; Boone, T.; Simonet, W.S.; et al. Reversal of cancer cachexia and muscle wasting by ActRIIB antagonism leads to prolonged survival. Cell 2010, 142, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, B.K. Muscle as a secretory organ. Compr. Physiol. 2013, 3, 1337–1362. [Google Scholar] [CrossRef]
- Whitham, M.; Febbraio, M.A. The ever-expanding myokinome: Discovery challenges and therapeutic implications. Nat. Rev. Drug Discov. 2016, 15, 719–729. [Google Scholar] [CrossRef]
- Bodine, S.C.; Baehr, L.M. Skeletal muscle atrophy and the E3 ubiquitin ligases MuRF1 and MAFbx/atrogin-1. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E469–E484. [Google Scholar] [CrossRef] [Green Version]
- Acharyya, S.; Ladner, K.J.; Nelsen, L.L.; Damrauer, J.; Reiser, P.J.; Swoap, S.; Guttridge, D.C. Cancer cachexia is regulated by selective targeting of skeletal muscle gene products. J. Clin. Investig. 2004, 114, 370–380. [Google Scholar] [CrossRef]
- Bonetto, A.; Rupert, J.E.; Barreto, R.; Zimmers, T.A. The Colon-26 Carcinoma Tumor-bearing Mouse as a Model for the Study of Cancer Cachexia. J. Vis. Exp. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertaggia, E.; Scabia, G.; Dalise, S.; Lo Verso, F.; Santini, F.; Vitti, P.; Chisari, C.; Sandri, M.; Maffei, M. Haptoglobin is required to prevent oxidative stress and muscle atrophy. PLoS ONE 2014, 9, e100745. [Google Scholar] [CrossRef] [PubMed]
- Passey, S.L.; Bozinovski, S.; Vlahos, R.; Anderson, G.P.; Hansen, M.J. Serum Amyloid A Induces Toll-Like Receptor 2-Dependent Inflammatory Cytokine Expression and Atrophy in C2C12 Skeletal Muscle Myotubes. PLoS ONE 2016, 11, e0146882. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Du, J.; Hu, Z.; Han, G.; Delafontaine, P.; Garcia, G.; Mitch, W.E. IL-6 and serum amyloid A synergy mediates angiotensin II-induced muscle wasting. J. Am. Soc. Nephrol. 2009, 20, 604–612. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, J.; Wang, S. Serum Amyloid A Induces a Vascular Smooth Muscle Cell Phenotype Switch through the p38 MAPK Signaling Pathway. BioMed Res. Int. 2017, 2017, 4941379. [Google Scholar] [CrossRef]
- Tjondrokoesoemo, A.; Schips, T.; Kanisicak, O.; Sargent, M.A.; Molkentin, J.D. Genetic overexpression of Serpina3n attenuates muscular dystrophy in mice. Hum. Mol. Genet. 2016, 25, 1192–1202. [Google Scholar] [CrossRef] [Green Version]
- Remy, K.E.; Cortes-Puch, I.; Solomon, S.B.; Sun, J.; Pockros, B.M.; Feng, J.; Lertora, J.J.; Hantgan, R.R.; Liu, X.; Perlegas, A.; et al. Haptoglobin improves shock, lung injury, and survival in canine pneumonia. JCI Insight 2018, 3. [Google Scholar] [CrossRef]
- Parajuli, P.; Kumar, S.; Loumaye, A.; Singh, P.; Eragamreddy, S.; Nguyen, T.L.; Ozkan, S.; Razzaque, M.S.; Prunier, C.; Thissen, J.P.; et al. Twist1 Activation in Muscle Progenitor Cells Causes Muscle Loss Akin to Cancer Cachexia. Dev. Cell 2018, 45, 712–725.e6. [Google Scholar] [CrossRef] [Green Version]
- Bindels, L.B.; Beck, R.; Schakman, O.; Martin, J.C.; De Backer, F.; Sohet, F.M.; Dewulf, E.M.; Pachikian, B.D.; Neyrinck, A.M.; Thissen, J.P.; et al. Restoring specific lactobacilli levels decreases inflammation and muscle atrophy markers in an acute leukemia mouse model. PLoS ONE 2012, 7, e37971. [Google Scholar] [CrossRef] [Green Version]
- Bindels, L.B.; Neyrinck, A.M.; Loumaye, A.; Catry, E.; Walgrave, H.; Cherbuy, C.; Leclercq, S.; Van Hul, M.; Plovier, H.; Pachikian, B.; et al. Increased gut permeability in cancer cachexia: Mechanisms and clinical relevance. Oncotarget 2018, 9, 18224–18238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, J.G.; Albrecht, U.; Haussinger, D.; Heinrich, P.C.; Schaper, F. Hepatic acute phase proteins--regulation by IL-6- and IL-1-type cytokines involving STAT3 and its crosstalk with NF-kappaB-dependent signaling. Eur J. Cell Biol. 2012, 91, 496–505. [Google Scholar] [CrossRef]
- Narsale, A.A.; Carson, J.A. Role of interleukin-6 in cachexia: Therapeutic implications. Curr. Opin. Support. Palliat. Care 2014, 8, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Gabay, C.; Kushner, I. Acute-phase proteins and other systemic responses to inflammation. N. Engl. J. Med. 1999, 340, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Shum, A.M.; Fung, D.C.; Corley, S.M.; McGill, M.C.; Bentley, N.L.; Tan, T.C.; Wilkins, M.R.; Polly, P. Cardiac and skeletal muscles show molecularly distinct responses to cancer cachexia. Physiol Genom. 2015, 47, 588–599. [Google Scholar] [CrossRef] [PubMed]
- Bonetto, A.; Aydogdu, T.; Kunzevitzky, N.; Guttridge, D.C.; Khuri, S.; Koniaris, L.G.; Zimmers, T.A. STAT3 activation in skeletal muscle links muscle wasting and the acute phase response in cancer cachexia. PLoS ONE 2011, 6, e22538. [Google Scholar] [CrossRef] [Green Version]
- Cornwell, E.W.; Mirbod, A.; Wu, C.L.; Kandarian, S.C.; Jackman, R.W. C26 cancer-induced muscle wasting is IKKbeta-dependent and NF-kappaB-independent. PLoS ONE 2014, 9, e87776. [Google Scholar] [CrossRef]
- Judge, S.M.; Wu, C.L.; Beharry, A.W.; Roberts, B.M.; Ferreira, L.F.; Kandarian, S.C.; Judge, A.R. Genome-wide identification of FoxO-dependent gene networks in skeletal muscle during C26 cancer cachexia. BMC Cancer 2014, 14, 997. [Google Scholar] [CrossRef] [Green Version]
- Talbert, E.E.; Cuitino, M.C.; Ladner, K.J.; Rajasekerea, P.V.; Siebert, M.; Shakya, R.; Leone, G.W.; Ostrowski, M.C.; Paleo, B.; Weisleder, N.; et al. Modeling Human Cancer-induced Cachexia. Cell Rep. 2019, 28, 1612–1622.e4. [Google Scholar] [CrossRef] [Green Version]
- Barreto, R.; Mandili, G.; Witzmann, F.A.; Novelli, F.; Zimmers, T.A.; Bonetto, A. Cancer and Chemotherapy Contribute to Muscle Loss by Activating Common Signaling Pathways. Front. Physiol. 2016, 7, 472. [Google Scholar] [CrossRef] [Green Version]
- Braun, T.P.; Grossberg, A.J.; Krasnow, S.M.; Levasseur, P.R.; Szumowski, M.; Zhu, X.X.; Maxson, J.E.; Gabriel Knoll, J.; Barnes, A.P.; Marks, D.L. Cancer- and endotoxin-induced cachexia require intact glucocorticoid signaling in skeletal muscle. FASEB J. 2013, 27, 3572–3582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa-Caldwell, M.E.; Fix, D.K.; Washington, T.A.; Greene, N.P. Muscle alterations in the development and progression of cancer-induced muscle atrophy: A review. J. Appl. Physiol. 2020, 128, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Argilés, J.M.; López-Soriano, F.J.; Busquets, S. Mediators of cachexia in cancer patients. Nutrition 2019, 66, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.; Dowling, P.; Zweyer, M.; Henry, M.; Meleady, P.; Mundegar, R.R.; Swandulla, D.; Ohlendieck, K. Proteomic profiling of mdx-4cv serum reveals highly elevated levels of the inflammation-induced plasma marker haptoglobin in muscular dystrophy. Int. J. Mol. Med. 2017, 39, 1357–1370. [Google Scholar] [CrossRef] [Green Version]
- Gueugneau, M.; d’Hose, D.; Barbe, C.; de Barsy, M.; Lause, P.; Maiter, D.; Bindels, L.B.; Delzenne, N.M.; Schaeffer, L.; Gangloff, Y.-G.; et al. Increased Serpina3n release into circulation during glucocorticoid-mediated muscle atrophy. J. Cachexia Sarcopenia Muscle 2018, 9, 929–946. [Google Scholar] [CrossRef] [Green Version]
- Langhans, C.; Weber-Carstens, S.; Schmidt, F.; Hamati, J.; Kny, M.; Zhu, X.; Wollersheim, T.; Koch, S.; Krebs, M.; Schulz, H.; et al. Inflammation-induced acute phase response in skeletal muscle and critical illness myopathy. PLoS ONE 2014, 9, e92048. [Google Scholar] [CrossRef] [Green Version]
- Ebhardt, H.A.; Degen, S.; Tadini, V.; Schilb, A.; Johns, N.; Greig, C.A.; Fearon, K.C.H.; Aebersold, R.; Jacobi, C. Comprehensive proteome analysis of human skeletal muscle in cachexia and sarcopenia: A pilot study. J. Cachexia Sarcopenia Muscle 2017, 8, 567–582. [Google Scholar] [CrossRef] [Green Version]
- Hahn, A.; Kny, M.; Pablo-Tortola, C.; Todiras, M.; Willenbrock, M.; Schmidt, S.; Schmoeckel, K.; Jorde, I.; Nowak, M.; Jarosch, E.; et al. Serum amyloid A1 mediates myotube atrophy via Toll-like receptors. J. Cachexia Sarcopenia Muscle 2019. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, C.; Li, Y.; Miwa, T.; Liu, C.; Cui, W.; Song, W.C.; Du, J. Complement C3a signaling facilitates skeletal muscle regeneration by regulating monocyte function and trafficking. Nat. Commun. 2017, 8, 2078. [Google Scholar] [CrossRef] [Green Version]
- Lautaoja, J.H.; Lalowski, M.; Nissinen, T.A.; Hentila, J.; Shi, Y.; Ritvos, O.; Cheng, S.; Hulmi, J.J. Muscle and serum metabolomes are dysregulated in colon-26 tumor-bearing mice despite amelioration of cachexia with activin receptor type 2B ligand blockade. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E852–E865. [Google Scholar] [CrossRef]
- Neyroud, D.; Nosacka, R.L.; Judge, A.R.; Hepple, R.T. Colon 26 adenocarcinoma (C26)-induced cancer cachexia impairs skeletal muscle mitochondrial function and content. J. Muscle Res. Cell Motil. 2019, 40, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Shum, A.M.; Mahendradatta, T.; Taylor, R.J.; Painter, A.B.; Moore, M.M.; Tsoli, M.; Tan, T.C.; Clarke, S.J.; Robertson, G.R.; Polly, P. Disruption of MEF2C signaling and loss of sarcomeric and mitochondrial integrity in cancer-induced skeletal muscle wasting. Aging 2012, 4, 133–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballarò, R.; Beltrà, M.; Lucia, S.D.; Pin, F.; Ranjbar, K.; Hulmi, J.J.; Costelli, P.; Penna, F. Moderate exercise in mice improves cancer plus chemotherapy-induced muscle wasting and mitochondrial alterations. FASEB J. 2019, 33, 5482–5494. [Google Scholar] [CrossRef]
- Pin, F.; Barretol, R.; Couch, M.E.; Bonetto, A.; O’Connell, T.M. Cachexia induced by cancer and chemotherapy yield distinct perturbations to energy metabolism. J. Cachexia Sarcopenia Muscle 2019, 10, 140–154. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L.; Rosa-Caldwell, M.E.; Lee, D.E.; Blackwell, T.A.; Brown, L.A.; Perry, R.A.; Haynie, W.S.; Hardee, J.P.; Carson, J.A.; Wiggs, M.P.; et al. Mitochondrial degeneration precedes the development of muscle atrophy in progression of cancer cachexia in tumour-bearing mice. J. Cachexia Sarcopenia Muscle 2017, 8, 926–938. [Google Scholar] [CrossRef]
- de Castro, G.S.; Simoes, E.; Lima, J.; Ortiz-Silva, M.; Festuccia, W.T.; Tokeshi, F.; Alcântara, P.S.; Otoch, J.P.; Coletti, D.; Seelaender, M. Human Cachexia Induces Changes in Mitochondria, Autophagy and Apoptosis in the Skeletal Muscle. Cancers 2019, 11, 1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nissinen, T.A.; Hentila, J.; Penna, F.; Lampinen, A.; Lautaoja, J.H.; Fachada, V.; Holopainen, T.; Ritvos, O.; Kivelä, R.; Hulmi, J.J. Treating cachexia using soluble ACVR2B improves survival, alters mTOR localization, and attenuates liver and spleen responses. J. Cachexia Sarcopenia Muscle 2018, 9, 514–529. [Google Scholar] [CrossRef]
- Deshmukh, A.S.; Murgia, M.; Nagaraj, N.; Treebak, J.T.; Cox, J.; Mann, M. Deep proteomics of mouse skeletal muscle enables quantitation of protein isoforms, metabolic pathways, and transcription factors. Mol. Cell Proteom. 2015, 14, 841–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shum, A.M.Y.; Poljak, A.; Bentley, N.L.; Turner, N.; Tan, T.C.; Polly, P. Proteomic profiling of skeletal and cardiac muscle in cancer cachexia: Alterations in sarcomeric and mitochondrial protein expression. Oncotarget 2018, 9, 22001–22022. [Google Scholar] [CrossRef] [Green Version]
- Sciorati, C.; Touvier, T.; Buono, R.; Pessina, P.; François, S.; Perrotta, C.; Meneveri, R.; Clementi, E.; Brunelli, S. Necdin is expressed in cachectic skeletal muscle to protect fibers from tumor-induced wasting. J. Cell Sci. 2009, 122, 1119–1125. [Google Scholar] [CrossRef] [Green Version]
- Pierre, N.; Salée, C.; Massot, C.; Blétard, N.; Mazzucchelli, G.; Smargiasso, N.; Morsa, D.; Baiwir, D.; De Pauw, E.; Reenaers, C.; et al. Proteomics Highlights Common and Distinct Pathophysiological Processes Associated with Ileal and Colonic Ulcers in Crohn’s Disease. J. Crohns Colitis 2020, 14, 205–215. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [Green Version]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- MacLean, B.; Tomazela, D.M.; Shulman, N.; Chambers, M.; Finney, G.L.; Frewen, B.; Kern, R.; Tabb, D.L.; Liebler, D.C.; MacCoss, M.J. Skyline: An open source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 2010, 26, 966–968. [Google Scholar] [CrossRef] [Green Version]
- Loumaye, A.; de Barsy, M.; Nachit, M.; Lause, P.; Frateur, L.; van Maanen, A.; Trefois, P.; Gruson, D.; Thissen, J.P. Role of Activin A and myostatin in human cancer cachexia. J. Clin. Endocrinol. Metab. 2015, 100, 2030–2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balan, E.; Schwalm, C.; Naslain, D.; Nielens, H.; Francaux, M.; Deldicque, L. Regular Endurance Exercise Promotes Fission, Mitophagy, and Oxidative Phosphorylation in Human Skeletal Muscle Independently of Age. Front. Physiol. 2019, 10, 1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | Number of Proteins Submitted | Category | Term | Genes | Count | % | p Value | FDR |
|---|---|---|---|---|---|---|---|---|
| SF | Downregulated (n = 120) | KEGG Pathway | Oxidative phosphorylation | Atp5f1a, Atp5f1b, Atp5f1c, Atp5f1d, Atp5me, Atp5mf, Atp5pb, Atp5pd, Atp5pf, Atp5po, Cox4i1, Cox5a, Cox5b, Cox6b1, Cyc1, Mtco2, Ndufa10, Ndufa11, Ndufa6, Ndufa8, Ndufa9, Ndufb10, Ndufb5, Ndufb8, Ndufs1, Ndufs2, Ndufs4, Ndufs7, Ndufv1, Ndufv2, Sdha, Sdhb, Uqcrb, Uqcrc1, Uqcrc2, Uqcrfs1, Uqcrh | 37 | 30.8 | 2.9 × 10−23 | 1.4 × 10−21 |
| Upregulated (n = 117) | KEGG Pathway | Complement and coagulation cascades | A2m, F2, C3, C4b, Fga, Fgb, Fgg, Kng1, Serpinf2, Serping1 | 10 | 8.5 | 2.6 × 10−6 | 4.0 × 10−4 | |
| UP Keywords | Acute phase | F2, Hp, Itih4, Orm1, Orm2, Serpina3n, Serpinf2, Saa1 | 8 | 6.8 | 9.6 × 10−6 | 5.1 × 10−4 | ||
| MF | Downregulated (n = 116) | KEGG Pathway | Ribosome | Rpl10a, Rpl11, Rpl12, Rpl13, Rpl13a, Rpl14, Rpl18, Rpl19, Rpl30, Rpl4, Rpl6, Rpl7, Rpl7a, Rplp0, Rplp1, Rplp2, Rps10, Rps16, Rps17, Rps18, Rps19, Rps2, Rps21, Rps 24, Rps26, Rps28, Rps3, Rps3a1, Rps5, Rps6, Rps7, Rps8, Rps9, Rpsa | 34 | 29.3 | 5.3 × 10−24 | 3.4 × 10−22 |
| Upregulated (n = 88) | KEGG Pathway | Complement and coagulation cascades | F13a1, C3, Cfh, Fga, Fgb, Fgg, Kng1 | 7 | 8.0 | 5.9 × 10−6 | 8.4 × 10−4 | |
| UP Keywords | Acute phase | Fn1, Hp, Itih4, Serpina3n, Saa1, Stat3 | 6 | 6.8 | 3.7 × 10−8 | 1.7 × 10−6 |
| Gene Name | Majority Protein ID | Protein Name | Classical Secretion (C) or Not (NC) | SF | MF | ||
|---|---|---|---|---|---|---|---|
| p Value | Ratio C26/CT | p Value | Ratio C26/CT | ||||
| Apoa4 | P06728 | Apolipoprotein A-IV | C | 5281 × 10−7 | 0.500 | ||
| Ces1c | P23953 | Carboxylesterase 1C | C | 1779 × 10−3 | 0.426 | ||
| Mup2 | P11589 | Major urinary protein 2 | C | 6556 × 10−3 | 0.484 | ||
| Mup17 | B5X0G2 | Major urinary protein 17 | C | 6094 × 10−7 | 0.174 | ||
| Pcolce | Q61398 | Procollagen C-endopeptidase enhancer 1 | C | Only CT | |||
| Rps2 | P25444 | 40S ribosomal protein S2 | NC | 3339 × 10−5 | 0.448 | ||
| Rps5 | P97461 | 40S ribosomal protein S5 | NC | Only CT | |||
| Serpinf1 | P97298 | Pigment epithelium-derived factor | C | 6618 × 10−3 | 0.446 | ||
| Ttr | P07309 | Transthyretin * | C | 2578 × 10−9 | 0.083 | ||
| Ambp | Q07456 | Protein AMBP * | C | 2548 × 10−4 | 4797 | ||
| Apcs | P12246 | Serum amyloid P-component | C | Only C26 | |||
| Apoe | P08226 | Apolipoprotein E * | C | Only C26 | 5437 × 10−5 | 3505 | |
| A2m | Q61838 | Alpha-2-macroglobulin * | C | 1415 × 10−9 | 2916 | ||
| A2mp | Q6GQT1 | Alpha-2-macroglobulin-P * | C | Only C26 | |||
| Cfh | P06909 | Complement factor H | C | 1394 × 10−5 | 9641 | ||
| C3 | P01027 | Complement C3 * | C | 3517 × 10−9 | 4018 | 6829 × 10−8 | 4402 |
| C4b | P01029 | Complement C4-B | C | 5570 × 10−3 | 2275 | ||
| Fga | E9PV24 | Fibrinogen alpha chain * | C | 8127 × 10−7 | 5936 | 2736 × 10−9 | 6110 |
| Fgb | Q8K0E8 | Fibrinogen beta chain * | C | 2127 × 10−5 | 6712 | 1536 × 10−7 | 12.839 |
| Fgg | Q8VCM7 | Fibrinogen gamma chain * | C | 1298 × 10−6 | 5306 | 3777 × 10−6 | 10.175 |
| Fn1 | P11276 | Fibronectin | C | 4368 × 10−5 | 2340 | ||
| Hp | Q61646 | Haptoglobin * | C | 1507 × 10−8 | 463.598 | Only C26 | |
| Hpx | Q91X72 | Hemopexin * | C | 2891 × 10−9 | 6647 | 2979 × 10−6 | 8883 |
| Itih3 | Q61704 | Inter-alpha-trypsin inhibitor heavy chain H3 * | C | 9150 × 10−8 | 14.356 | ||
| Itih4 | A6X935 | Inter alpha-trypsin inhibitor, heavy chain 4 * | C | 9107 × 10−7 | 62.174 | Only C26 | |
| Kng1 | O08677 | Kininogen-1 * | C | 1164 × 10−8 | 2588 | ||
| Orm1 | Q60590 | Alpha-1-acid glycoprotein 1 * | C | 2897 × 10−9 | 14.330 | ||
| Orm2 | P07361 | Alpha-1-acid glycoprotein 2 * | C | Only C26 | |||
| Pdk4 | O70571 | [Pyruvate dehydrogenase (acetyl-transferring)] kinase isozyme 4, mitochondrial * | NC | 6087 × 10−6 | 29.186 | 1593 × 10−7 | 22.703 |
| SAA1 | P05366 | Serum amyloid A-1 protein * | C | Only C26 | Only C26 | ||
| Serpina3n | Q91WP6 | Serine protease inhibitor A3N * | C | Only C26 | Only C26 | ||
| Serpina10 | Q8R121 | Protein Z-dependent protease inhibitor * | C | only C26 | |||
| Sod3 | O09164 | Extracellular superoxide dismutase [Cu-Zn] | C | Only C26 | |||
| Tcap | O70548 | Telethonin * | NC | 5359 × 10−7 | 2053 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Massart, I.S.; Paulissen, G.; Loumaye, A.; Lause, P.; Pötgens, S.A.; Thibaut, M.M.; Balan, E.; Deldicque, L.; Atfi, A.; Louis, E.; et al. Marked Increased Production of Acute Phase Reactants by Skeletal Muscle during Cancer Cachexia. Cancers 2020, 12, 3221. https://doi.org/10.3390/cancers12113221
Massart IS, Paulissen G, Loumaye A, Lause P, Pötgens SA, Thibaut MM, Balan E, Deldicque L, Atfi A, Louis E, et al. Marked Increased Production of Acute Phase Reactants by Skeletal Muscle during Cancer Cachexia. Cancers. 2020; 12(11):3221. https://doi.org/10.3390/cancers12113221
Chicago/Turabian StyleMassart, Isabelle S., Geneviève Paulissen, Audrey Loumaye, Pascale Lause, Sarah A. Pötgens, Morgane M. Thibaut, Estelle Balan, Louise Deldicque, Azeddine Atfi, Edouard Louis, and et al. 2020. "Marked Increased Production of Acute Phase Reactants by Skeletal Muscle during Cancer Cachexia" Cancers 12, no. 11: 3221. https://doi.org/10.3390/cancers12113221