The Proteasome Inhibitor Ixazomib Inhibits the Formation and Growth of Pulmonary and Abdominal Osteosarcoma Metastases in Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
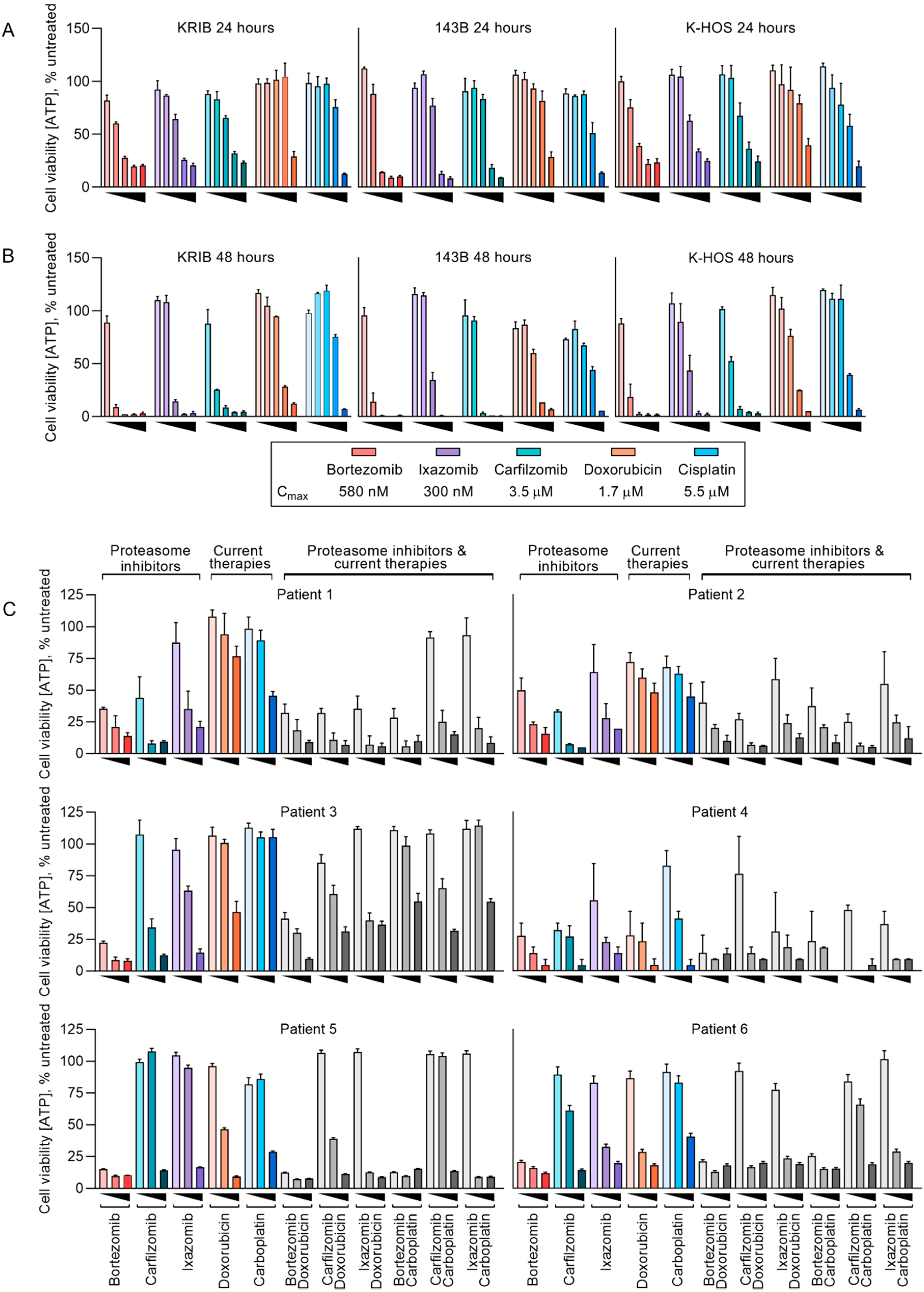
2.1. Established Human Osteosarcoma Cell Lines and Freshly Resected Canine Osteosarcoma Cells Are Sensitive to Proteasome Inhibitors at Physiologically Relevant Concentrations
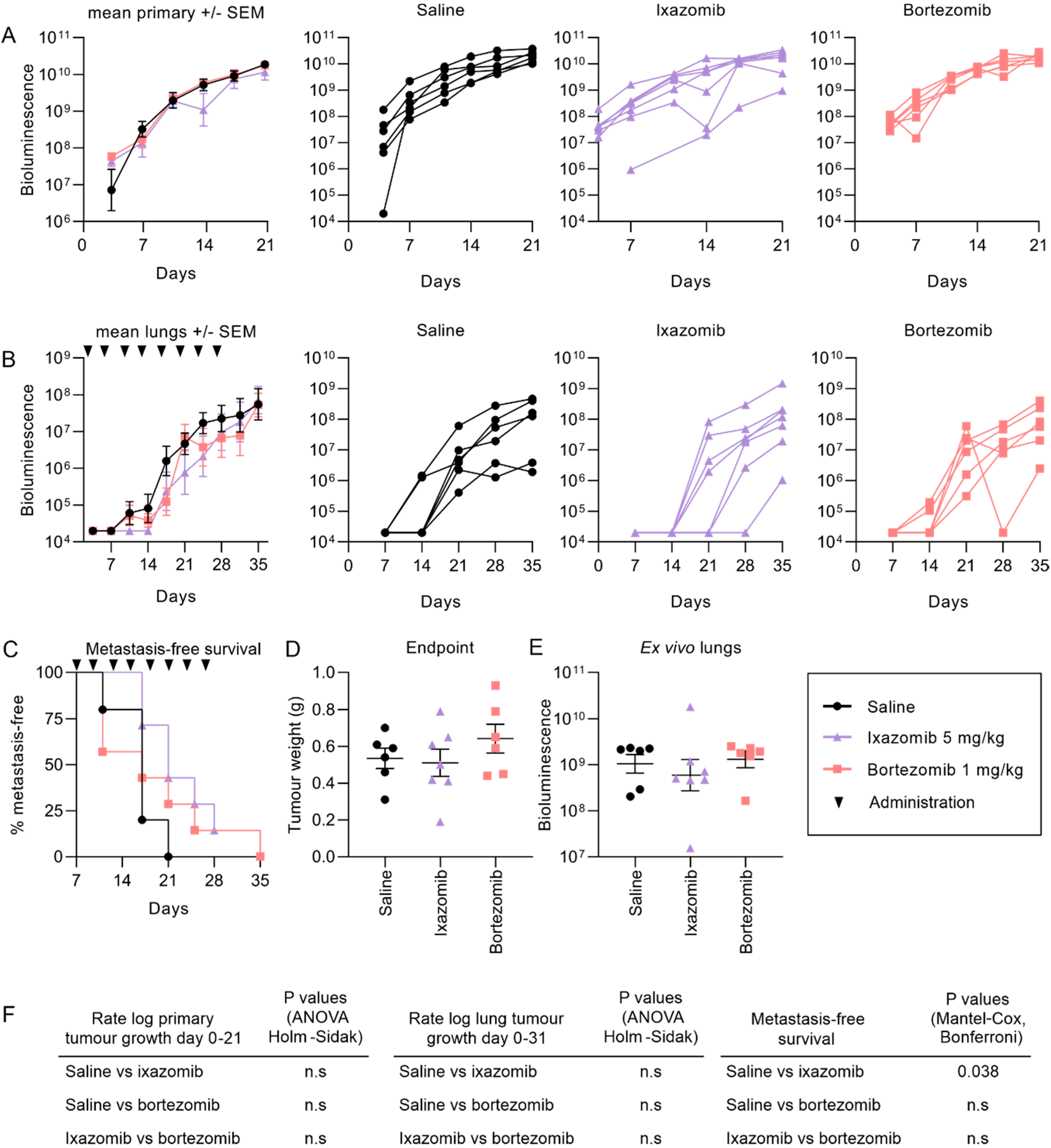
2.2. Ixazomib and Bortezomib Inhibit the Formation and Growth of Pulmonary KRIB Osteosarcoma Metastases But Not Primary Intramuscular Tumors
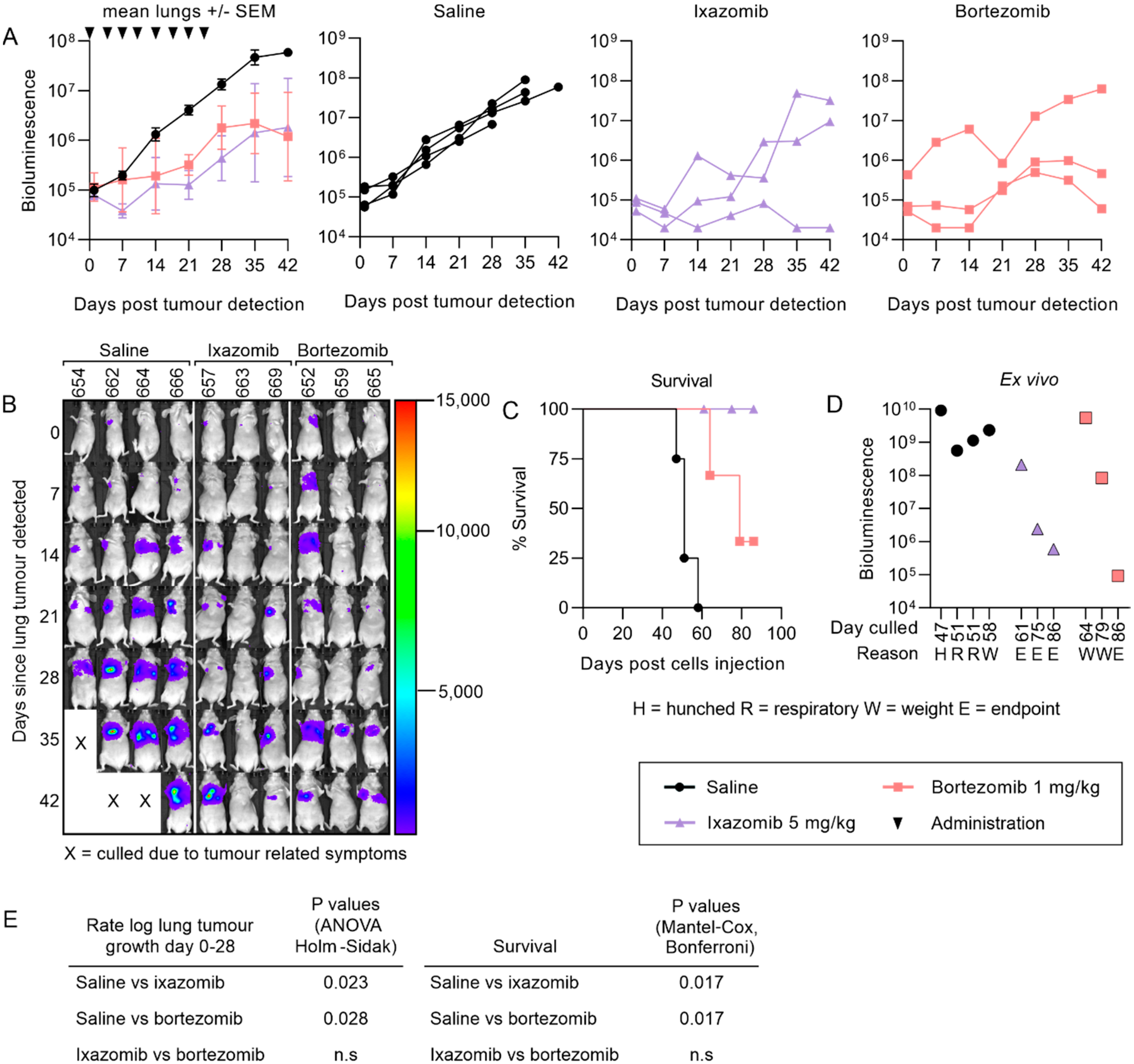
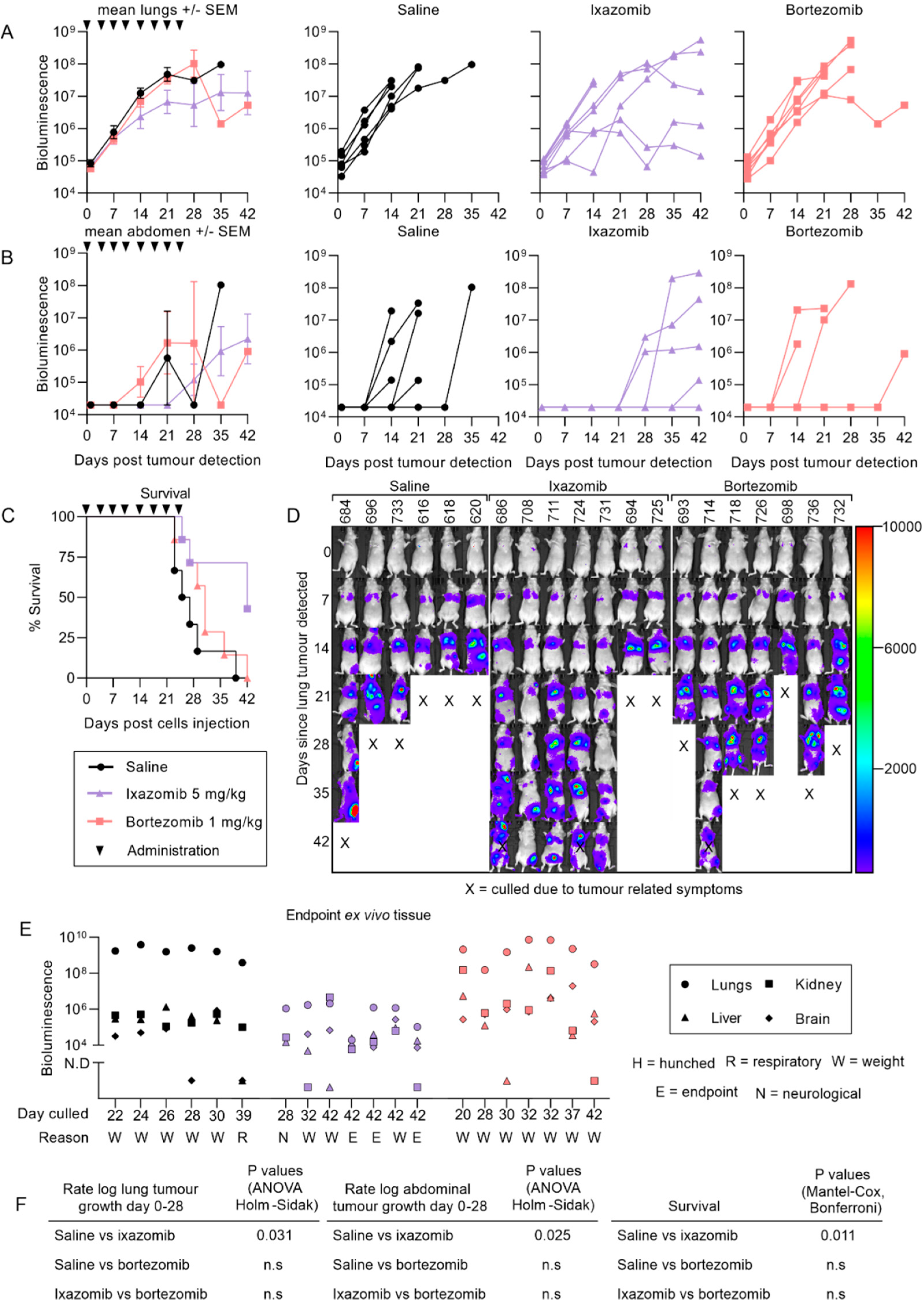
2.3. Ixazomib Inhibits the Growth of 143B Osteosarcoma Metastases and Enhances Survival Compared to Saline Treated Mice
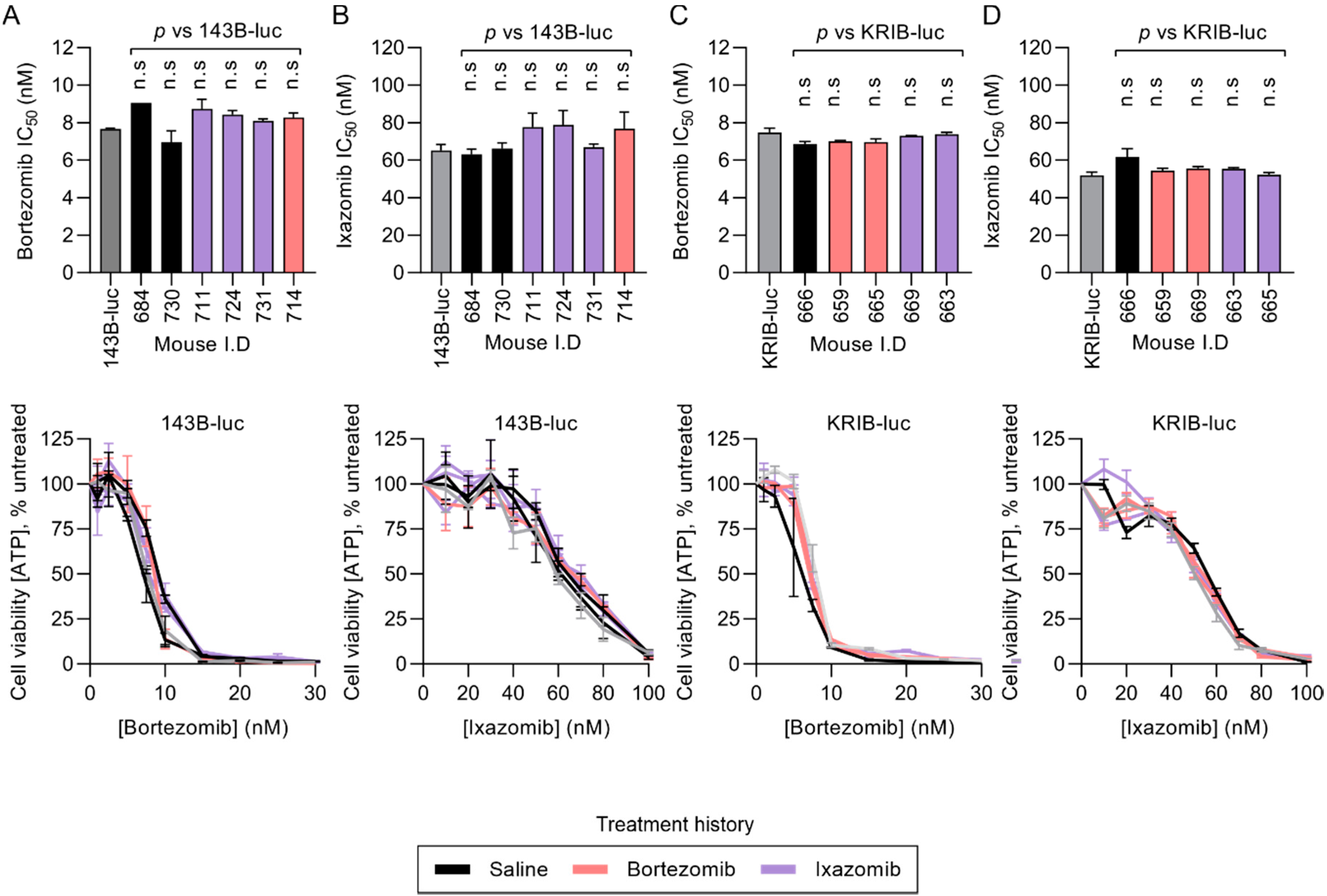
2.4. Resected KRIB-luc and 143B-luc Osteosarcoma Cells Do Not Acquire Resistance During In Vivo Treatment with Proteasome Inhibitors
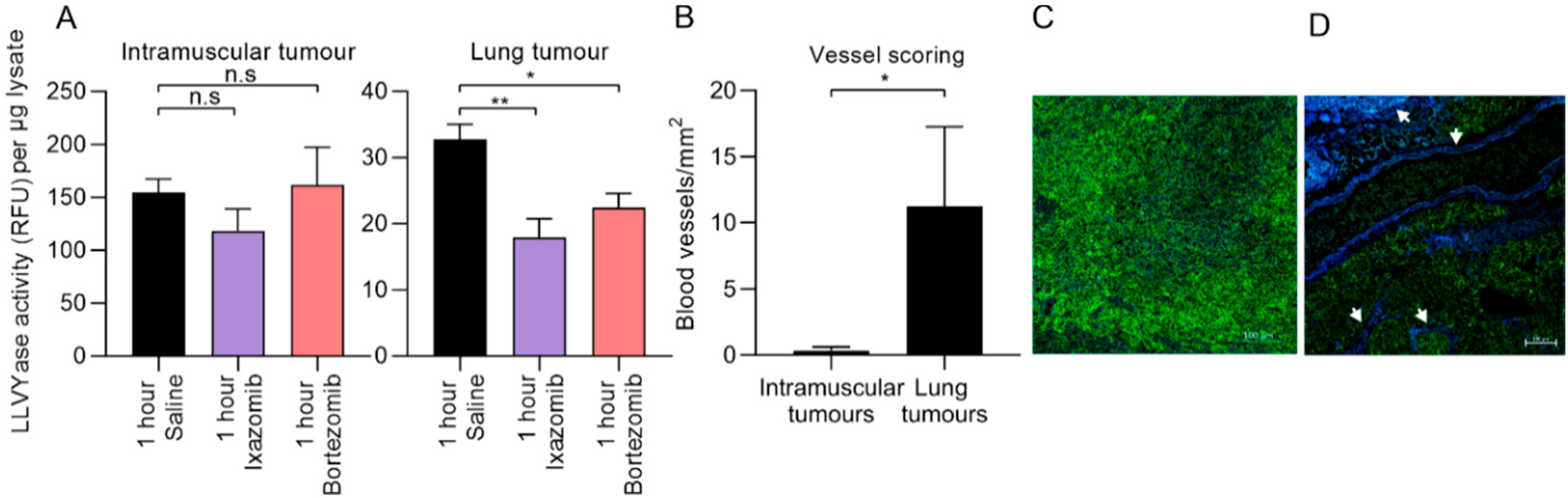
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Drugs
4.3. Animal Studies
4.4. Cell Viability Assays
4.5. Immunofluorescence
4.6. Pharmacodynamics
4.7. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mirabello, L.; Troisi, R.J.; Savage, S.A. International osteosarcoma incidence patterns in children and adolescents, middle ages and elderly persons. Int. J. Cancer 2009, 125, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Gianferante, D.M.; Mirabello, L.; Savage, S.A. Germline and somatic genetics of osteosarcoma - connecting aetiology, biology and therapy. Nat. Rev. Endocrinol. 2017, 13, 480–491. [Google Scholar] [CrossRef]
- McGuire, J.; Utset-Ward, T.; Reed, D.; Lynch, C.C. Re-calculating! Navigating through the osteosarcoma treatment roadblock. Pharmacol. Res. 2017, 117, 54–64. [Google Scholar] [CrossRef]
- Allison, D.C.; Carney, S.C.; Ahlmann, E.R.; Hendifar, A.; Chawla, S.; Fedenko, A.; Angeles, C.; Menendez, L.R. A meta-analysis of osteosarcoma outcomes in the modern medical era. Sarcoma 2012, 2012, 704872. [Google Scholar] [CrossRef] [Green Version]
- Meazza, C.; Scanagatta, P. Metastatic osteosarcoma: A challenging multidisciplinary treatment. Expert Rev. Anticancer Ther. 2016, 16, 543–556. [Google Scholar] [CrossRef]
- Richardson, P.G.; Sonneveld, P.; Schuster, M.W.; Irwin, D.; Stadtmauer, E.A.; Facon, T.; Harousseau, J.-L.; Ben-Yehuda, D.; Lonial, S.; Goldschmidt, H. Bortezomib or high-dose dexamethasone for relapsed multiple myeloma. N. Engl. J. Med. 2005, 352, 2487–2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raedler, L. Velcade (Bortezomib) Receives 2 New FDA Indications: For Retreatment of Patients with Multiple Myeloma and for First-Line Treatment of Patients with Mantle-Cell Lymphoma. Am. Health Drug Benefits 2015, 8, 135–140. [Google Scholar] [PubMed]
- Manasanch, E.E.; Orlowski, R.Z. Proteasome inhibitors in cancer therapy. Nat. Rev. Clin. Oncol. 2017, 14, 417–433. [Google Scholar] [CrossRef]
- Patatsos, K.; Shekhar, T.M.; Hawkins, C.J. Pre-clinical evaluation of proteasome inhibitors for canine and human osteosarcoma. Vet. Comp. Oncol. 2018, 16, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Shapovalov, Y.; Benavidez, D.; Zuch, D.; Eliseev, R.A. Proteasome inhibition with bortezomib suppresses growth and induces apoptosis in osteosarcoma. Int. J. Cancer 2010, 127, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Lou, Z.; Ren, T.; Peng, X.; Sun, Y.; Jiao, G.; Lu, Q.; Zhang, S.; Lu, X.; Guo, W. Bortezomib induces apoptosis and autophagy in osteosarcoma cells through mitogen-activated protein kinase pathway in vitro. J. Int. Med. Res. 2013, 41, 1505–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaney, S.M.; Bernstein, M.; Neville, K.; Ginsberg, J.; Kitchen, B.; Horton, T.; Berg, S.L.; Krailo, M.; Adamson, P.C. Phase I study of the proteasome inhibitor bortezomib in pediatric patients with refractory solid tumors: A Children’s Oncology Group study (ADVL0015). J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2004, 22, 4804–4809. [Google Scholar] [CrossRef] [PubMed]
- Maki, R.G.; Kraft, A.S.; Scheu, K.; Yamada, J.; Wadler, S.; Antonescu, C.R.; Wright, J.J.; Schwartz, G.K. A multicenter Phase II study of bortezomib in recurrent or metastatic sarcomas. Cancer 2005, 103, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Marten, A.; Zeiss, N.; Serba, S.; Mehrle, S.; von Lilienfeld-Toal, M.; Schmidt, J. Bortezomib is ineffective in an orthotopic mouse model of pancreatic adenocarcinoma. Mol. Cancer Ther. 2008, 7, 3624–3631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kupperman, E.; Lee, E.C.; Cao, Y.; Bannerman, B.; Fitzgerald, M.; Berger, A.; Yu, J.; Yang, Y.; Hales, P.; Bruzzese, F.; et al. Evaluation of the proteasome inhibitor MLN9708 in preclinical models of human cancer. Cancer Res. 2010, 70, 1970–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Wu, Y.; Zhou, X.; Xu, J.; Zhu, W.; Shu, Y.; Liu, P. Efficacy of therapy with bortezomib in solid tumors: A review based on 32 clinical trials. Future Oncol. 2014, 10, 1795–1807. [Google Scholar] [CrossRef]
- Roeten, M.S.F.; Cloos, J.; Jansen, G. Positioning of proteasome inhibitors in therapy of solid malignancies. Cancer Chemother. Pharm. 2018, 81, 227–243. [Google Scholar] [CrossRef] [Green Version]
- Flynt, E.; Bisht, K.; Sridharan, V.; Ortiz, M.; Towfic, F.; Thakurta, A. Prognosis, Biology, and Targeting of TP53 Dysregulation in Multiple Myeloma. Cells 2020, 9. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Hu, J.; Li, W.; Song, G.; Shen, J. Combined bortezomib-based chemotherapy and p53 gene therapy using hollow mesoporous silica nanospheres for p53 mutant non-small cell lung cancer treatment. Biomater. Sci. 2016, 5, 77–88. [Google Scholar] [CrossRef]
- Ling, X.; Calinski, D.; Chanan-Khan, A.A.; Zhou, M.; Li, F. Cancer cell sensitivity to bortezomib is associated with survivin expression and p53 status but not cancer cell types. J. Exp. Clin. Cancer Res. 2010, 29, 8. [Google Scholar] [CrossRef] [Green Version]
- Buscail, L.; Bournet, B.; Cordelier, P. Role of oncogenic KRAS in the diagnosis, prognosis and treatment of pancreatic cancer. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.W.; Lin, S.W.; Lin, L.C.; Hsu, Y.H.; Lin, Y.F.; Ho, S.Y.; Wu, Y.H.; Wang, Y.J. Synergistic antitumor effects of radiation and proteasome inhibitor treatment in pancreatic cancer through the induction of autophagy and the downregulation of TRAF6. Cancer Lett. 2015, 365, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Michaelis, M.; Fichtner, I.; Behrens, D.; Haider, W.; Rothweiler, F.; Mack, A.; Cinatl, J.; Doerr, H.W.; Cinatl, J. Anti-cancer effects of bortezomib against chemoresistant neuroblastoma cell lines in vitro and in vivo. Int. J. Oncol. 2006, 28, 439–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bersani, F.; Taulli, R.; Accornero, P.; Morotti, A.; Miretti, S.; Crepaldi, T.; Ponzetto, C. Bortezomib-mediated proteasome inhibition as a potential strategy for the treatment of rhabdomyosarcoma. Eur. J. Cancer 2008, 44, 876–884. [Google Scholar] [CrossRef]
- Brignole, C.; Marimpietri, D.; Pastorino, F.; Nico, B.; Di Paolo, D.; Cioni, M.; Piccardi, F.; Cilli, M.; Pezzolo, A.; Corrias, M.V.; et al. Effect of bortezomib on human neuroblastoma cell growth, apoptosis, and angiogenesis. J. Natl. Cancer Inst. 2006, 98, 1142–1157. [Google Scholar] [CrossRef] [Green Version]
- Bullova, P.; Cougnoux, A.; Marzouca, G.; Kopacek, J.; Pacak, K. Bortezomib Alone and in Combination With Salinosporamid A Induces Apoptosis and Promotes Pheochromocytoma Cell Death In Vitro and in Female Nude Mice. Endocrinology 2017, 158, 3097–3108. [Google Scholar] [CrossRef]
- Williamson, M.J.; Silva, M.D.; Terkelsen, J.; Robertson, R.; Yu, L.; Xia, C.; Hatsis, P.; Bannerman, B.; Babcock, T.; Cao, Y.; et al. The relationship among tumor architecture, pharmacokinetics, pharmacodynamics, and efficacy of bortezomib in mouse xenograft models. Mol. Cancer Ther. 2009, 8, 3234–3243. [Google Scholar] [CrossRef] [Green Version]
- Zuccari, G.; Milelli, A.; Pastorino, F.; Loi, M.; Petretto, A.; Parise, A.; Marchetti, C.; Minarini, A.; Cilli, M.; Emionite, L.; et al. Tumor vascular targeted liposomal-bortezomib minimizes side effects and increases therapeutic activity in human neuroblastoma. J. Control Release 2015, 211, 44–52. [Google Scholar] [CrossRef]
- Richardson, P.G.; Zweegman, S.; O’Donnell, E.K.; Laubach, J.P.; Raje, N.; Voorhees, P.; Ferrari, R.H.; Skacel, T.; Kumar, S.K.; Lonial, S. Ixazomib for the treatment of multiple myeloma. Expert Opin. Pharm. 2018, 19, 1949–1968. [Google Scholar] [CrossRef]
- Okazuka, K.; Ishida, T. Proteasome inhibitors for multiple myeloma. Jpn. J. Clin. Oncol. 2018, 48, 785–793. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Fu, C.; Sun, J.; Wang, X.; Geng, S.; Wang, X.; Zou, J.; Bi, Z.; Yang, C. A New Perspective for Osteosarcoma Therapy: Proteasome Inhibition by MLN9708/2238 Successfully Induces Apoptosis and Cell Cycle Arrest and Attenuates the Invasion Ability of Osteosarcoma Cells in Vitro. Cell Physiol. Biochem. 2017, 41, 451–465. [Google Scholar] [CrossRef]
- Mirabello, L.; Troisi, R.J.; Savage, S.A. Osteosarcoma incidence and survival rates from 1973 to 2004: Data from the Surveillance, Epidemiology, and End Results Program. Cancer 2009, 115, 1531–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelan, J.S.; Bielack, S.S.; Marina, N.; Smeland, S.; Jovic, G.; Hook, J.M.; Krailo, M.; Anninga, J.; Butterfass-Bahloul, T.; Bohling, T.; et al. EURAMOS-1, an international randomised study for osteosarcoma: Results from pre-randomisation treatment. Ann. Oncol. 2015, 26, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Marina, N.M.; Smeland, S.; Bielack, S.S.; Bernstein, M.; Jovic, G.; Krailo, M.D.; Hook, J.M.; Arndt, C.; van den Berg, H.; Brennan, B.; et al. Comparison of MAPIE versus MAP in patients with a poor response to preoperative chemotherapy for newly diagnosed high-grade osteosarcoma (EURAMOS-1): An open-label, international, randomised controlled trial. Lancet Oncol. 2016, 17, 1396–1408. [Google Scholar] [CrossRef] [Green Version]
- Meyers, P.A.; Schwartz, C.L.; Krailo, M.; Kleinerman, E.S.; Betcher, D.; Bernstein, M.L.; Conrad, E.; Ferguson, W.; Gebhardt, M.; Goorin, A.M.; et al. Osteosarcoma: A randomized, prospective trial of the addition of ifosfamide and/or muramyl tripeptide to cisplatin, doxorubicin, and high-dose methotrexate. J. Clin. Oncol. 2005, 23, 2004–2011. [Google Scholar] [CrossRef] [PubMed]
- Rowell, J.L.; McCarthy, D.O.; Alvarez, C.E. Dog models of naturally occurring cancer. Trends Mol. Med. 2011, 17, 380–388. [Google Scholar] [CrossRef] [Green Version]
- Mueller, F.; Fuchs, B.; Kaser-Hotz, B. Comparative biology of human and canine osteosarcoma. Anticancer Res. 2007, 27, 155–164. [Google Scholar]
- Simpson, S.; Dunning, M.D.; de Brot, S.; Grau-Roma, L.; Mongan, N.P.; Rutland, C.S. Comparative review of human and canine osteosarcoma: Morphology, epidemiology, prognosis, treatment and genetics. Acta Vet. Scand. 2017, 59, 71. [Google Scholar] [CrossRef]
- Shekhar, T.M.; Burvenich, I.J.G.; Harris, M.A.; Rigopoulos, A.; Zanker, D.; Spurling, A.; Parker, B.S.; Walkley, C.R.; Scott, A.M.; Hawkins, C.J. Smac mimetics LCL161 and GDC-0152 inhibit osteosarcoma growth and metastasis in mice. BMC Cancer 2019, 19, 924. [Google Scholar] [CrossRef]
- Mori, Y.; Shirai, T.; Terauchi, R.; Tsuchida, S.; Mizoshiri, N.; Hayashi, D.; Arai, Y.; Kishida, T.; Mazda, O.; Kubo, T. Antitumor effects of pristimerin on human osteosarcoma cells in vitro and in vivo. Oncotargets Ther. 2017, 10, 5703–5710. [Google Scholar] [CrossRef] [Green Version]
- Harris, M.A.; Shekhar, T.M.; Coupland, L.A.; Miles, M.A.; Hawkins, C.J. Transient NK cell depletion facilitates pulmonary osteosarcoma metastases after intravenous inoculation in athymic mice. JAYAO 2020, in press. [Google Scholar]
- Moreau, P.; Karamanesht, I.I.; Domnikova, N.; Kyselyova, M.Y.; Vilchevska, K.V.; Doronin, V.A.; Schmidt, A.; Hulin, C.; Leleu, X.; Esseltine, D.L.; et al. Pharmacokinetic, pharmacodynamic and covariate analysis of subcutaneous versus intravenous administration of bortezomib in patients with relapsed multiple myeloma. Clin. Pharmacokinet. 2012, 51, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Richardson, P.G.; Baz, R.; Wang, M.; Jakubowiak, A.J.; Laubach, J.P.; Harvey, R.D.; Talpaz, M.; Berg, D.; Liu, G.; Yu, J.; et al. Phase 1 study of twice-weekly ixazomib, an oral proteasome inhibitor, in relapsed/refractory multiple myeloma patients. Blood 2014, 124, 1038–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quach, H.; White, D.; Spencer, A.; Ho, P.J.; Bhutani, D.; White, M.; Inamdar, S.; Morris, C.; Ou, Y.; Gyger, M. Pharmacokinetics and safety of carfilzomib in patients with relapsed multiple myeloma and end-stage renal disease (ESRD): An open-label, single-arm, phase I study. Cancer Chemother. Pharmacol. 2017, 79, 1067–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rushing, D.A.; Piscitelli, S.C.; Rodvold, K.A.; Tewksbury, D.A. The disposition of doxorubicin on repeated dosing. J. Clin. Pharm. 1993, 33, 698–702. [Google Scholar] [CrossRef] [PubMed]
- van Hennik, M.B.; van der Vijgh, W.J.; Klein, I.; Elferink, F.; Vermorken, J.B.; Winograd, B.; Pinedo, H.M. Comparative pharmacokinetics of cisplatin and three analogues in mice and humans. Cancer Res. 1987, 47, 6297–6301. [Google Scholar]
- Kirpensteijn, J.; Kik, M.; Teske, E.; Rutteman, G.R. TP53 gene mutations in canine osteosarcoma. Vet. Surg. 2008, 37, 454–460. [Google Scholar] [CrossRef]
- Fenger, J.M.; London, C.A.; Kisseberth, W.C. Canine osteosarcoma: A naturally occurring disease to inform pediatric oncology. Inst. Lab. Anim. Res. J. 2014, 55, 69–85. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, J.; Zhao, N.; Wang, C.; Kamar, S.; Zhou, Y.; He, Z.; Yang, J.; Sun, B.; Shi, X.; et al. Progress in the chemotherapeutic treatment of osteosarcoma. Oncol. Lett. 2018, 16, 6228–6237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selmic, L.E.; Burton, J.H.; Thamm, D.H.; Withrow, S.J.; Lana, S.E. Comparison of carboplatin and doxorubicin-based chemotherapy protocols in 470 dogs after amputation for treatment of appendicular osteosarcoma. J. Vet. Intern. Med. 2014, 28, 554–563. [Google Scholar] [CrossRef] [Green Version]
- Gaver, R.C.; George, A.M.; Duncan, G.F.; Morris, A.D.; Deeb, G.; Faulkner, H.C.; Farmen, R.H. The disposition of carboplatin in the beagle dog. Cancer Chemother. Pharmacol. 1988, 21, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-C.; Lo, W.-J.; Huang, Y.-T.; Lin, C.-L.; Feng, C.-C.; Lin, H.-T.; Cheng, H.-C.; Yeh, S.-P. Deferasirox has strong anti-leukemia activity but may antagonize theanti-leukemia effect of doxorubicin. Leuk. Lymphoma 2017, 58, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Uddin, S.; Ahmed, M.; Bavi, P.; El-Sayed, R.; Al-Sanea, N.; AbdulJabbar, A.; Ashari, L.H.; Alhomoud, S.; Al-Dayel, F.; Hussain, A.R.; et al. Bortezomib (Velcade) induces p27Kip1 expression through S-phase kinase protein 2 degradation in colorectal cancer. Cancer Res. 2008, 68, 3379–3388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Park, E.; Jung, C.-K.; Kang, S.-W.; Kim, B.G.; Jung, Y.; Kim, T.H.; Lim, J.-Y.; Lee, S.-E.; Min, C.-K.; et al. Oral proteasome inhibitor with strong preclinical efficacy in myeloma models. BMC Cancer 2016, 16, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augello, G.; Modica, M.; Azzolina, A.; Puleio, R.; Cassata, G.; Emma, M.R.; Di Sano, C.; Cusimano, A.; Montalto, G.; Cervello, M. Preclinical evaluation of antitumor activity of the proteasome inhibitor MLN2238 (ixazomib) in hepatocellular carcinoma cells. Cell Death Dis. 2018, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Kisselev, A.F.; Goldberg, A.L. Monitoring Activity and Inhibition of 26S Proteasomes with Fluorogenic Peptide Substrates. Methods Enzymol. 2005, 398, 364–378. [Google Scholar] [CrossRef]
- Goyal, S.; Julka, P.K. Recurrent osteosarcoma with calcified liver metastases: Uncommon development of a common disease. J. Cancer Res. Ther. 2017, 13, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Jeffree, G.M.; Price, C.H.; Sissons, H.A. The metastatic patterns of osteosarcoma. Br. J. Cancer 1975, 32, 87–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xian, M.; Cao, H.; Cao, J.; Shao, X.; Zhu, D.; Zhang, N.; Huang, P.; Li, W.; Yang, B.; Ying, M.; et al. Bortezomib sensitizes human osteosarcoma cells to adriamycin-induced apoptosis through ROS-dependent activation of p-eIF2α/ATF4/CHOP axis. Int. J. Cancer 2017, 141, 1029–1041. [Google Scholar] [CrossRef]
- Zhao, Y.; Foster, N.R.; Meyers, J.P.; Thomas, S.P.; Northfelt, D.W.; Rowland, K.M., Jr.; Mattar, B.I.; Johnson, D.B.; Molina, J.R.; Mandrekar, S.J.; et al. A phase I/II study of bortezomib in combination with paclitaxel, carboplatin, and concurrent thoracic radiation therapy for non-small-cell lung cancer: North Central Cancer Treatment Group (NCCTG)-N0321. J. Thorac. Oncol. 2015, 10, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Aghajanian, C.; Dizon, D.S.; Sabbatini, P.; Raizer, J.J.; Dupont, J.; Spriggs, D.R. Phase I trial of bortezomib and carboplatin in recurrent ovarian or primary peritoneal cancer. J. Clin. Oncol. 2005, 23, 5943–5949. [Google Scholar] [CrossRef] [PubMed]
- Taromi, S.; Lewens, F.; Arsenic, R.; Sedding, D.; Sänger, J.; Kunze, A.; Möbs, M.; Benecke, J.; Freitag, H.; Christen, F.J.O. Proteasome inhibitor bortezomib enhances the effect of standard chemotherapy in small cell lung cancer. Oncotarget 2017, 8, 97061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, A.M.; Chansky, K.; Lara, P.N., Jr.; Gumerlock, P.H.; Crowley, J.; Albain, K.S.; Vogel, S.J.; Gandara, D.R.; Southwest Oncology, G. Bortezomib plus gemcitabine/carboplatin as first-line treatment of advanced non-small cell lung cancer: A phase II Southwest Oncology Group Study (S0339). J. Thorac. Oncol. Off. Publ. Int. Assoc. Study Lung Cancer 2009, 4, 87–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piperdi, B.; Walsh, W.V.; Bradley, K.; Zhou, Z.; Bathini, V.; Hanrahan-Boshes, M.; Hutchinson, L.; Perez-Soler, R. Phase-I/II study of bortezomib in combination with carboplatin and bevacizumab as first-line therapy in patients with advanced non-small-cell lung cancer. J. Thorac. Oncol. Off. Publ. Int. Assoc. Study Lung Cancer 2012, 7, 1032–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grantab, R.H.; Tannock, I.F. Penetration of anticancer drugs through tumour tissue as a function of cellular packing density and interstitial fluid pressure and its modification by bortezomib. BMC Cancer 2012, 12, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Yu, Y.; Jiang, Z.; Cao, W.M.; Wang, Z.; Dou, J.; Zhao, Y.; Cui, Y.; Zhang, H. Next-generation proteasome inhibitor MLN9708 sensitizes breast cancer cells to doxorubicin-induced apoptosis. Sci. Rep. 2016, 6, 26456. [Google Scholar] [CrossRef]
- Al-Eisawi, Z.; Beale, P.; Chan, C.; Jun, Q.Y.; Huq, F. Carboplatin and oxaliplatin in sequenced combination with bortezomib in ovarian tumour models. J. Ovarian Res. 2013, 6, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, K.G.; Kuhn, D.J.; Kazi, A.; Dou, Q.P. Anti-angiogenic and anti-tumor properties of proteasome inhibitors. Curr. Cancer Drug Targets 2005, 5, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, M.; Bieker, R.; Bielack, S.S.; Auras, T.; Buerger, H.; Gosheger, G.; Jurgens, H.; Berdel, W.E.; Mesters, R.M. Prognostic Relevance of Increased Angiogenesis in Osteosarcoma. Clin. Cancer Res. 2004, 10, 8531–8537. [Google Scholar] [CrossRef] [Green Version]
- Oda, Y.; Yamamoto, H.; Tamiya, S.; Matsuda, S.; Tanaka, K.; Yokoyama, R.; Iwamoto, Y.; Tsuneyoshi, M. CXCR4 and VEGF expression in the primary site and the metastatic site of human osteosarcoma: Analysis within a group of patients, all of whom developed lung metastasis. Mod. Pathol. 2006, 19, 738–745. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, S.; Kumar, R.; Sankineani, S.R.; Marimuthu, K.; Rijal, L.; Prakash, S.; Jalan, D.; Khan, S.A.; Sharma, M.C. Role of vascular endothelial growth factor as a tumour marker in osteosarcoma: A prospective study. Int. Orthop. 2012, 36, 2315–2321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Li, S.; Wang, X.; Wang, M.; Tian, Q.; Yang, J.; Wang, J.; Wang, B.; Liu, P.; Yang, J. Tumor vasculature remolding by thalidomide increases delivery and efficacy of cisplatin. J. Exp. Clin. Cancer Res. 2019, 38, 427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budzynski, W.; Radzikowski, C. Cytotoxic cells in immunodeficient athymic mice. Immunopharmacol. Immunotoxicol. 1994, 16, 319–346. [Google Scholar] [CrossRef] [PubMed]
- Shanker, A.; Pellom, S.T., Jr.; Dudimah, D.F.; Thounaojam, M.C.; de Kluyver, R.L.; Brooks, A.D.; Yagita, H.; McVicar, D.W.; Murphy, W.J.; Longo, D.L.; et al. Bortezomib Improves Adoptive T-cell Therapy by Sensitizing Cancer Cells to FasL Cytotoxicity. Cancer Res. 2015, 75, 5260–5272. [Google Scholar] [CrossRef] [Green Version]
- Pellom, S.T., Jr.; Dudimah, D.F.; Thounaojam, M.C.; Uzhachenko, R.V.; Singhal, A.; Richmond, A.; Shanker, A. Bortezomib augments lymphocyte stimulatory cytokine signaling in the tumor microenvironment to sustain CD8+T cell antitumor function. Oncotarget 2017, 8, 8604–8621. [Google Scholar] [CrossRef] [PubMed]
- Shekhar, T.M.; Miles, M.A.; Gupte, A.; Taylor, S.; Tascone, B.; Walkley, C.R.; Hawkins, C.J. IAP antagonists sensitize murine osteosarcoma cells to killing by TNFα. Oncotarget 2016, 7, 33866–33886. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harris, M.A.; Miles, M.A.; Shekhar, T.M.; Cerra, C.; Georgy, S.R.; Ryan, S.D.; Cannon, C.M.; Hawkins, C.J. The Proteasome Inhibitor Ixazomib Inhibits the Formation and Growth of Pulmonary and Abdominal Osteosarcoma Metastases in Mice. Cancers 2020, 12, 1207. https://doi.org/10.3390/cancers12051207
Harris MA, Miles MA, Shekhar TM, Cerra C, Georgy SR, Ryan SD, Cannon CM, Hawkins CJ. The Proteasome Inhibitor Ixazomib Inhibits the Formation and Growth of Pulmonary and Abdominal Osteosarcoma Metastases in Mice. Cancers. 2020; 12(5):1207. https://doi.org/10.3390/cancers12051207
Chicago/Turabian StyleHarris, Michael A., Mark A. Miles, Tanmay M. Shekhar, Carmelo Cerra, Smitha R. Georgy, Stewart D. Ryan, Claire M. Cannon, and Christine J. Hawkins. 2020. "The Proteasome Inhibitor Ixazomib Inhibits the Formation and Growth of Pulmonary and Abdominal Osteosarcoma Metastases in Mice" Cancers 12, no. 5: 1207. https://doi.org/10.3390/cancers12051207