Preferential Response of Basal-Like Head and Neck Squamous Cell Carcinoma Cell Lines to EGFR-Targeted Therapy Depending on EREG-Driven Oncogenic Addiction

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
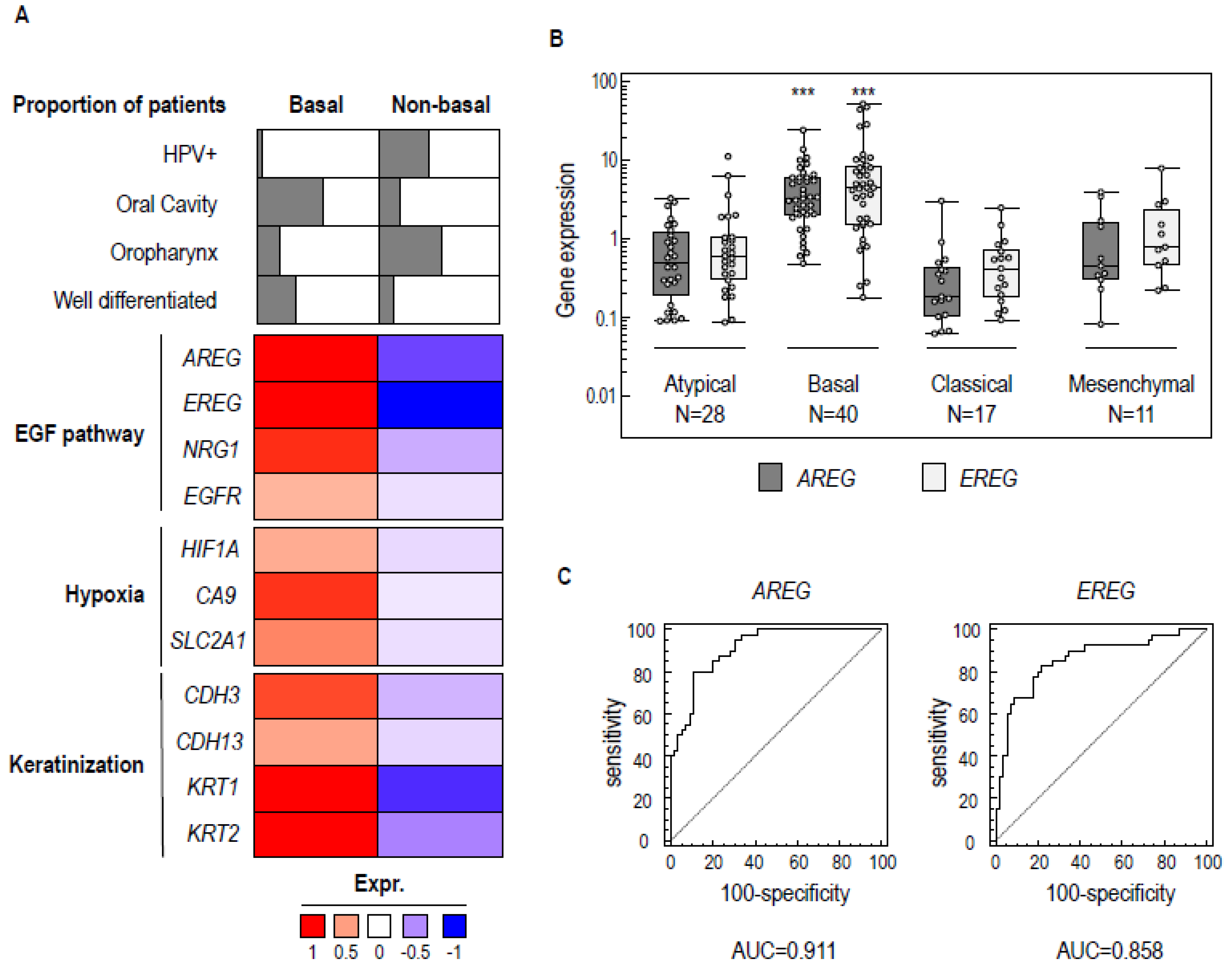
2.1. A Common Molecular Basal-Like Subgroup Can Be Distinguished in Different HNSCC Data Sets
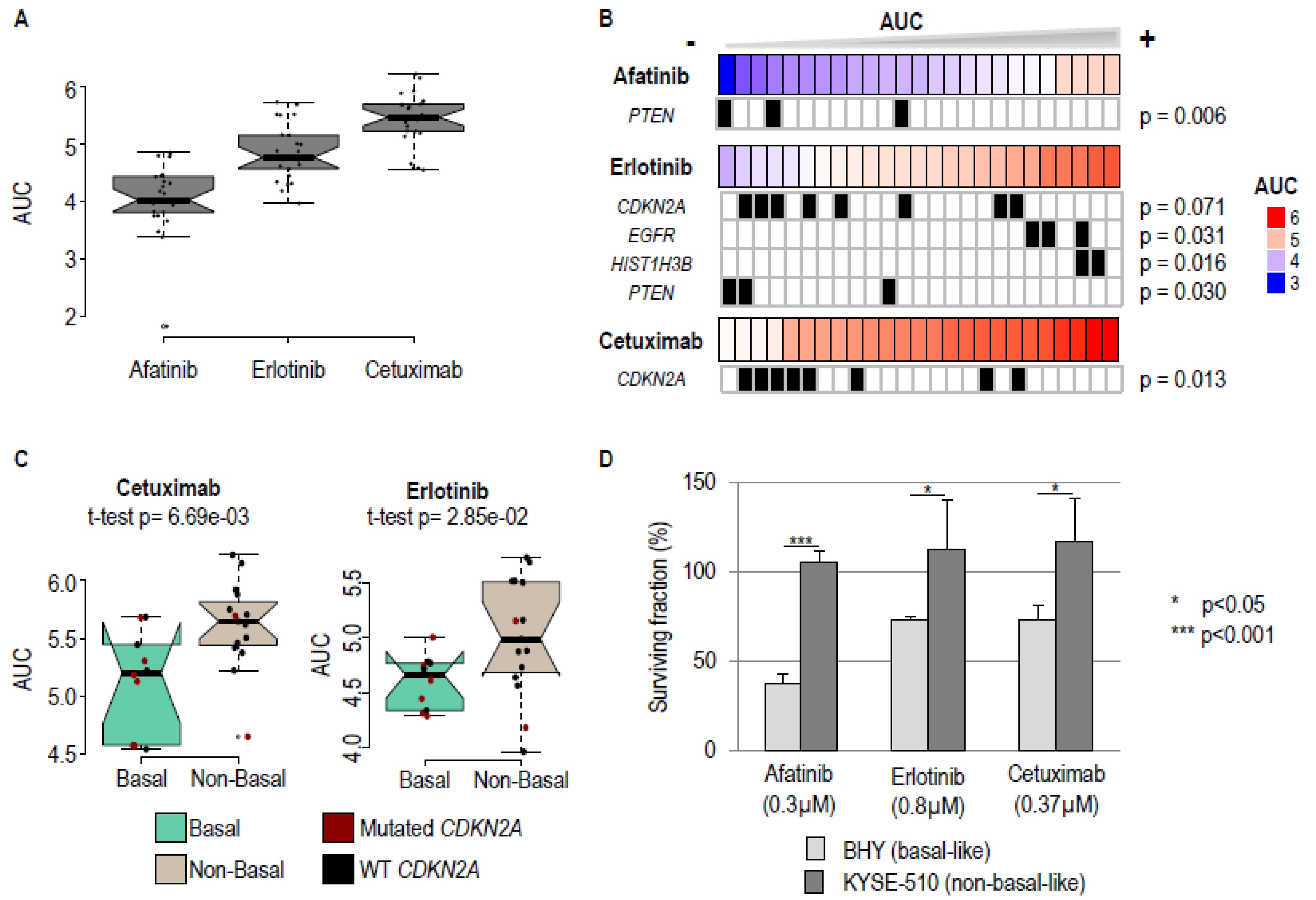
2.2. Basal-Like Cell Lines Are More Sensitive to Pharmacological EGFR Blockade
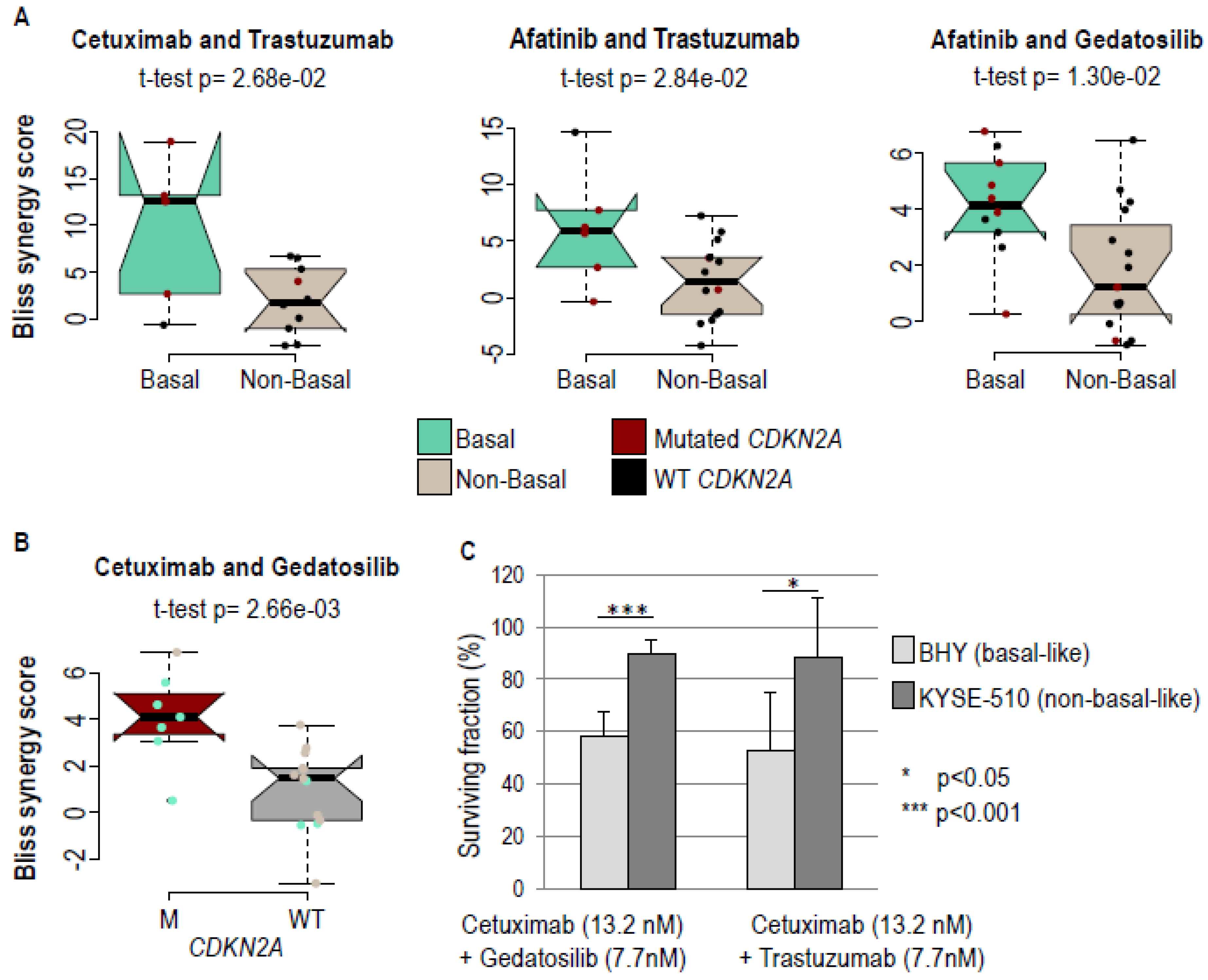
2.3. Greater Response of Basal Cell Lines to Certain Drug Combinations
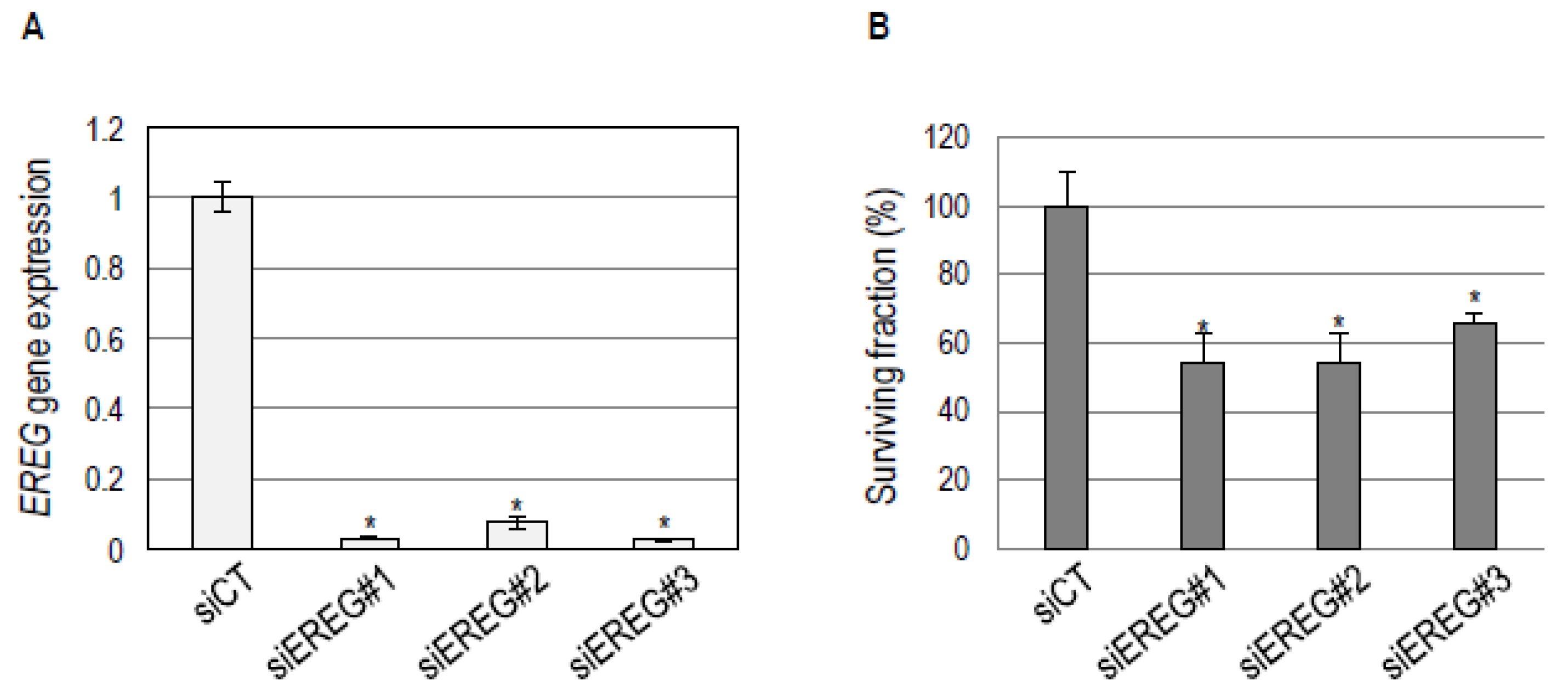
2.4. EREG Downregulation in the Basal-Like BHY Cell Line Inhibits Cell Survival
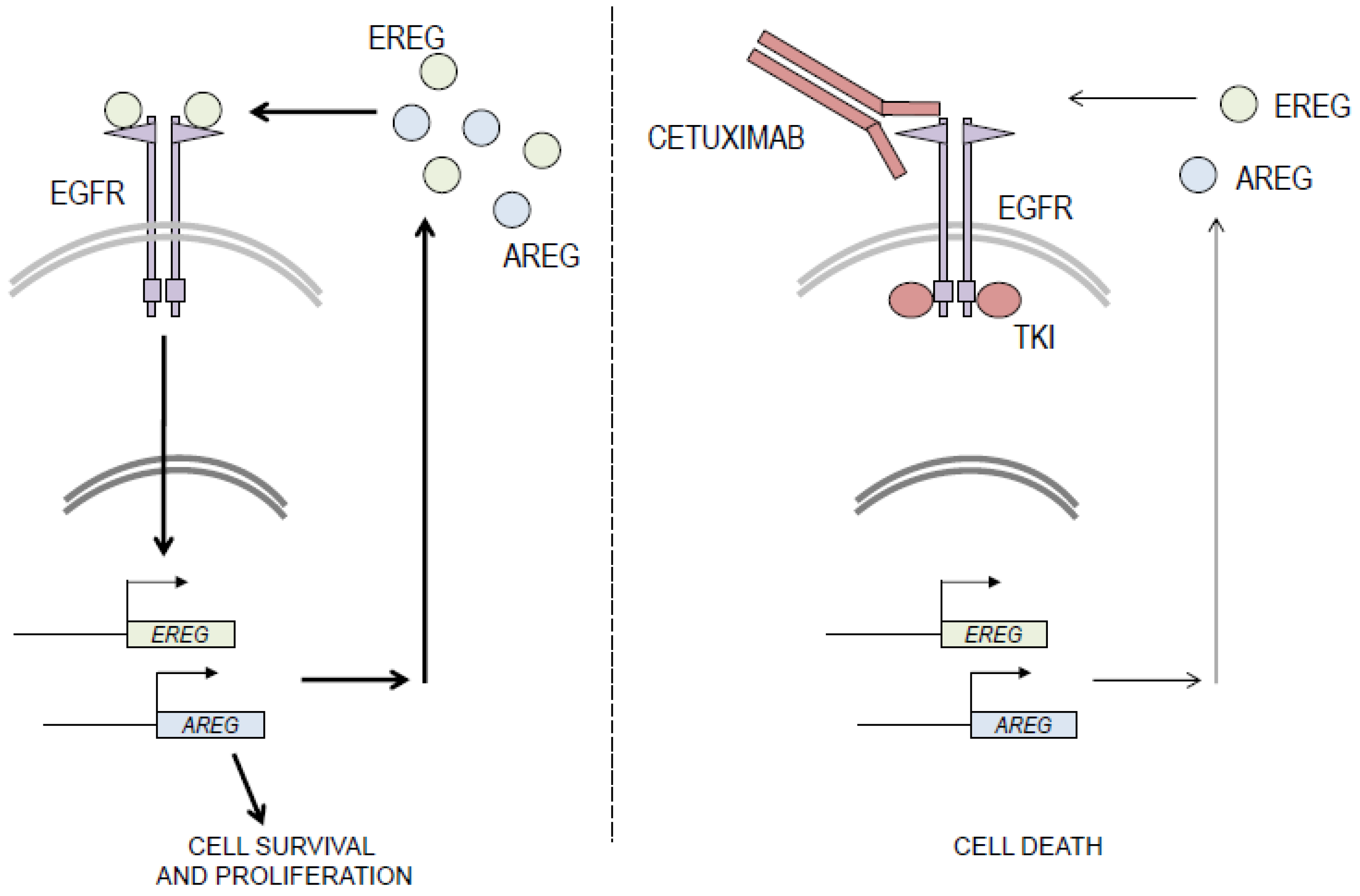
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Reagents
4.3. Small Interfering RNA Transient Transfection
4.4. Single-Drug and Drug-Combination Dose-Response Analyses
4.5. Next Generation Sequencing
4.6. Datasets
4.7. Data Analyses
4.8. Clonogenic Survival Assays
4.9. Gene Expression Assays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Leemans, C.R.; Snijders, P.J.F.; Brakenhoff, R.H. The molecular landscape of head and neck cancer. Nat. Rev. Cancer 2018, 18, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Argiris, A.; Harrington, K.J.; Tahara, M.; Schulten, J.; Chomette, P.; Ferreira Castro, A.; Licitra, L. Evidence-Based Treatment Options in Recurrent and/or Metastatic Squamous Cell Carcinoma of the Head and Neck. Front. Oncol. 2017, 7, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonner, J.A.; Harari, P.M.; Giralt, J.; Azarnia, N.; Shin, D.M.; Cohen, R.B.; Jones, C.U.; Sur, R.; Raben, D.; Jassem, J.; et al. Radiotherapy plus cetuximab for squamous-cell carcinoma of the head and neck. N. Engl. J. Med. 2006, 354, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.H.; Parker, J.S.; Karaca, G.; Wu, J.; Funkhouser, W.K.; Moore, D.; Butterfoss, D.; Xiang, D.; Zanation, A.; Yin, X.; et al. Molecular classification of head and neck squamous cell carcinomas using patterns of gene expression. Cancer Cell 2004, 5, 489–500. [Google Scholar] [CrossRef] [Green Version]
- Walter, V.; Yin, X.; Wilkerson, M.D.; Cabanski, C.R.; Zhao, N.; Du, Y.; Ang, M.K.; Hayward, M.C.; Salazar, A.H.; Hoadley, K.A.; et al. Molecular subtypes in head and neck cancer exhibit distinct patterns of chromosomal gain and loss of canonical cancer genes. PLoS ONE 2013, 8, e56823. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Network. Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature 2015, 517, 576–582. [Google Scholar] [CrossRef] [Green Version]
- Keck, M.K.; Zuo, Z.; Khattri, A.; Stricker, T.P.; Brown, C.D.; Imanguli, M.; Rieke, D.; Endhardt, K.; Fang, P.; Bragelmann, J.; et al. Integrative analysis of head and neck cancer identifies two biologically distinct HPV and three non-HPV subtypes. Clin. Cancer Res. 2015, 21, 870–881. [Google Scholar] [CrossRef]
- De Cecco, L.; Nicolau, M.; Giannoccaro, M.; Daidone, M.G.; Bossi, P.; Locati, L.; Licitra, L.; Canevari, S. Head and neck cancer subtypes with biological and clinical relevance: Meta-analysis of gene-expression data. Oncotarget 2015, 6, 9627–9642. [Google Scholar] [CrossRef] [Green Version]
- Bossi, P.; Bergamini, C.; Siano, M.; Cossu Rocca, M.; Sponghini, A.P.; Favales, F.; Giannoccaro, M.; Marchesi, E.; Cortelazzi, B.; Perrone, F.; et al. Functional Genomics Uncover the Biology behind the Responsiveness of Head and Neck Squamous Cell Cancer Patients to Cetuximab. Clin. Cancer Res. 2016, 22, 3961–3970. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Soares, J.; Greninger, P.; Edelman, E.J.; Lightfoot, H.; Forbes, S.; Bindal, N.; Beare, D.; Smith, J.A.; Thompson, I.R.; et al. Genomics of Drug Sensitivity in Cancer (GDSC): A resource for therapeutic biomarker discovery in cancer cells. Nucleic Acids Res. 2013, 41, D955–D961. [Google Scholar] [CrossRef] [PubMed]
- Boeckx, C.; Baay, M.; Wouters, A.; Specenier, P.; Vermorken, J.B.; Peeters, M.; Lardon, F. Anti-epidermal growth factor receptor therapy in head and neck squamous cell carcinoma: Focus on potential molecular mechanisms of drug resistance. Oncologist 2013, 18, 850–864. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Carpenter, G.; Coffey, R.J. EGF receptor ligands: Recent advances. F1000Research 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Riese, D.J., 2nd; Cullum, R.L. Epiregulin: Roles in normal physiology and cancer. Semin. Cell Dev. Biol. 2014, 28, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunaga, N.; Kaira, K. Epiregulin as a therapeutic target in non-small-cell lung cancer. Lung Cancer 2015, 6, 91–98. [Google Scholar] [CrossRef]
- Yang, J.; Li, S.; Wang, B.; Wu, Y.; Chen, Z.; Lv, M.; Lin, Y.; Yang, J. Potential biomarkers for anti-EGFR therapy in metastatic colorectal cancer. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 11645–11655. [Google Scholar] [CrossRef] [PubMed]
- Tonella, L.; Giannoccaro, M.; Alfieri, S.; Canevari, S.; De Cecco, L. Gene Expression Signatures for Head and Neck Cancer Patient Stratification: Are Results Ready for Clinical Application? Curr. Treat. Options Oncol. 2017, 18, 32. [Google Scholar] [CrossRef] [PubMed]
- Jung, A.C.; Job, S.; Ledrappier, S.; Macabre, C.; Abecassis, J.; de Reynies, A.; Wasylyk, B. A poor prognosis subtype of HNSCC is consistently observed across methylome, transcriptome, and miRNome analysis. Clin. Cancer Res. 2013, 19, 4174–4184. [Google Scholar] [CrossRef] [PubMed]
- Berasain, C.; Avila, M.A. Amphiregulin. Semin. Cell Dev. Biol. 2014, 28, 31–41. [Google Scholar] [CrossRef]
- Garnett, M.J.; Edelman, E.J.; Heidorn, S.J.; Greenman, C.D.; Dastur, A.; Lau, K.W.; Greninger, P.; Thompson, I.R.; Luo, X.; Soares, J.; et al. Systematic identification of genomic markers of drug sensitivity in cancer cells. Nature 2012, 483, 570–575. [Google Scholar] [CrossRef] [Green Version]
- Barretina, J.; Caponigro, G.; Stransky, N.; Venkatesan, K.; Margolin, A.A.; Kim, S.; Wilson, C.J.; Lehar, J.; Kryukov, G.V.; Sonkin, D.; et al. The Cancer Cell Line Encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature 2012, 483, 603–607. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, V.; Rosa, R.; D’Amato, C.; Formisano, L.; Marciano, R.; Nappi, L.; Raimondo, L.; Di Mauro, C.; Servetto, A.; Fusciello, C.; et al. The dual PI3K/mTOR inhibitor PKI-587 enhances sensitivity to cetuximab in EGFR-resistant human head and neck cancer models. Br. J. Cancer 2014, 110, 2887–2895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harari, P.M.; Wheeler, D.L.; Grandis, J.R. Molecular target approaches in head and neck cancer: Epidermal growth factor receptor and beyond. Semin. Radiat. Oncol. 2009, 19, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Soulieres, D.; Senzer, N.N.; Vokes, E.E.; Hidalgo, M.; Agarwala, S.S.; Siu, L.L. Multicenter phase II study of erlotinib, an oral epidermal growth factor receptor tyrosine kinase inhibitor, in patients with recurrent or metastatic squamous cell cancer of the head and neck. J. Clin. Oncol. 2004, 22, 77–85. [Google Scholar] [CrossRef]
- Machiels, J.P.; Haddad, R.I.; Fayette, J.; Licitra, L.F.; Tahara, M.; Vermorken, J.B.; Clement, P.M.; Gauler, T.; Cupissol, D.; Grau, J.J.; et al. Afatinib versus methotrexate as second-line treatment in patients with recurrent or metastatic squamous-cell carcinoma of the head and neck progressing on or after platinum-based therapy (LUX-Head & Neck 1): An open-label, randomised phase 3 trial. Lancet Oncol. 2015, 16, 583–594. [Google Scholar] [CrossRef]
- Martins, R.G.; Parvathaneni, U.; Bauman, J.E.; Sharma, A.K.; Raez, L.E.; Papagikos, M.A.; Yunus, F.; Kurland, B.F.; Eaton, K.D.; Liao, J.J.; et al. Cisplatin and radiotherapy with or without erlotinib in locally advanced squamous cell carcinoma of the head and neck: A randomized phase II trial. J. Clin. Oncol. 2013, 31, 1415–1421. [Google Scholar] [CrossRef]
- Kawaguchi, Y.; Kono, K.; Mimura, K.; Sugai, H.; Akaike, H.; Fujii, H. Cetuximab induce antibody-dependent cellular cytotoxicity against EGFR-expressing esophageal squamous cell carcinoma. Int. J. Cancer 2007, 120, 781–787. [Google Scholar] [CrossRef]
- Lattanzio, L.; Denaro, N.; Vivenza, D.; Varamo, C.; Strola, G.; Fortunato, M.; Chamorey, E.; Comino, A.; Monteverde, M.; Lo Nigro, C.; et al. Elevated basal antibody-dependent cell-mediated cytotoxicity (ADCC) and high epidermal growth factor receptor (EGFR) expression predict favourable outcome in patients with locally advanced head and neck cancer treated with cetuximab and radiotherapy. Cancer Immunol. Immunother. 2017, 66, 573–579. [Google Scholar] [CrossRef]
- Cohen, E.E.W.; Licitra, L.F.; Burtness, B.; Fayette, J.; Gauler, T.; Clement, P.M.; Grau, J.J.; Del Campo, J.M.; Mailliez, A.; Haddad, R.I.; et al. Biomarkers predict enhanced clinical outcomes with afatinib versus methotrexate in patients with second-line recurrent and/or metastatic head and neck cancer. Ann. Oncol. 2017, 28, 2526–2532. [Google Scholar] [CrossRef]
- Sato-Kuwabara, Y.; Neves, J.I.; Fregnani, J.H.; Sallum, R.A.; Soares, F.A. Evaluation of gene amplification and protein expression of HER-2/neu in esophageal squamous cell carcinoma using Fluorescence in situ Hybridization (FISH) and immunohistochemistry. BMC Cancer 2009, 9, 6. [Google Scholar] [CrossRef]
- Xia, W.; Lau, Y.K.; Zhang, H.Z.; Liu, A.R.; Li, L.; Kiyokawa, N.; Clayman, G.L.; Katz, R.L.; Hung, M.C. Strong correlation between c-erbB-2 overexpression and overall survival of patients with oral squamous cell carcinoma. Clin. Cancer Res. 1997, 3, 3–9. [Google Scholar] [PubMed]
- Arkhipov, A.; Shan, Y.; Kim, E.T.; Dror, R.O.; Shaw, D.E. Her2 activation mechanism reflects evolutionary preservation of asymmetric ectodomain dimers in the human EGFR family. Elife 2013, 2, e00708. [Google Scholar] [CrossRef] [PubMed]
- Erjala, K.; Sundvall, M.; Junttila, T.T.; Zhang, N.; Savisalo, M.; Mali, P.; Kulmala, J.; Pulkkinen, J.; Grenman, R.; Elenius, K. Signaling via ErbB2 and ErbB3 associates with resistance and epidermal growth factor receptor (EGFR) amplification with sensitivity to EGFR inhibitor gefitinib in head and neck squamous cell carcinoma cells. Clin. Cancer Res. 2006, 12, 4103–4111. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.L.; Huang, S.; Kruser, T.J.; Nechrebecki, M.M.; Armstrong, E.A.; Benavente, S.; Gondi, V.; Hsu, K.T.; Harari, P.M. Mechanisms of acquired resistance to cetuximab: Role of HER (ErbB) family members. Oncogene 2008, 27, 3944–3956. [Google Scholar] [CrossRef] [PubMed]
- Quesnelle, K.M.; Grandis, J.R. Dual kinase inhibition of EGFR and HER2 overcomes resistance to cetuximab in a novel in vivo model of acquired cetuximab resistance. Clin. Cancer Res. 2011, 17, 5935–5944. [Google Scholar] [CrossRef] [PubMed]
- Kondo, N.; Tsukuda, M.; Sakakibara, A.; Takahashi, H.; Hyakusoku, H.; Komatsu, M.; Niho, T.; Nakazaki, K.; Toth, G. Combined molecular targeted drug therapy for EGFR and HER-2 in head and neck squamous cell carcinoma cell lines. Int. J. Oncol. 2012, 40, 1805–1812. [Google Scholar] [CrossRef] [PubMed]
- Baselga, J.; Bradbury, I.; Eidtmann, H.; Di Cosimo, S.; de Azambuja, E.; Aura, C.; Gomez, H.; Dinh, P.; Fauria, K.; Van Dooren, V.; et al. Lapatinib with trastuzumab for HER2-positive early breast cancer (NeoALTTO): A randomised, open-label, multicentre, phase 3 trial. Lancet 2012, 379, 633–640. [Google Scholar] [CrossRef]
- Hanusch, C.; Schneeweiss, A.; Loibl, S.; Untch, M.; Paepke, S.; Kummel, S.; Jackisch, C.; Huober, J.; Hilfrich, J.; Gerber, B.; et al. Dual Blockade with AFatinib and Trastuzumab as NEoadjuvant Treatment for Patients with Locally Advanced or Operable Breast Cancer Receiving Taxane-Anthracycline Containing Chemotherapy-DAFNE (GBG-70). Clin. Cancer Res. 2015, 21, 2924–2931. [Google Scholar] [CrossRef]
- Collins, D.M.; Gately, K.; Hughes, C.; Edwards, C.; Davies, A.; Madden, S.F.; O’Byrne, K.J.; O’Donovan, N.; Crown, J. Tyrosine kinase inhibitors as modulators of trastuzumab-mediated antibody-dependent cell-mediated cytotoxicity in breast cancer cell lines. Cell. Immunol. 2017, 319, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Okita, R.; Shimizu, K.; Nojima, Y.; Yukawa, T.; Maeda, A.; Saisho, S.; Nakata, M. Lapatinib enhances trastuzumab-mediated antibody-dependent cellular cytotoxicity via upregulation of HER2 in malignant mesothelioma cells. Oncol. Rep. 2015, 34, 2864–2870. [Google Scholar] [CrossRef] [Green Version]
- Bour, B.; (University of Strasbourg, Inserm U1113, Centre Paul Strauss, Strasbourg, France); Wasylyk, W.; (IGBMC, UMR 7104 CNRS-UdS, U.1258 INSERM, Strasbourg, France); Jung, A.C.; (University of Strasbourg, Inserm U1113, Centre Paul Strauss, Strasbourg, France). Personal communication, 2019.
- Oliveras-Ferraros, C.; Cufi, S.; Queralt, B.; Vazquez-Martin, A.; Martin-Castillo, B.; de Llorens, R.; Bosch-Barrera, J.; Brunet, J.; Menendez, J.A. Cross-suppression of EGFR ligands amphiregulin and epiregulin and de-repression of FGFR3 signalling contribute to cetuximab resistance in wild-type KRAS tumour cells. Br. J. Cancer 2012, 106, 1406–1414. [Google Scholar] [CrossRef] [PubMed]
- Oshima, G.; Wennerberg, J.; Yamatodani, T.; Kjellen, E.; Mineta, H.; Johnsson, A.; Ekblad, L. Autocrine epidermal growth factor receptor ligand production and cetuximab response in head and neck squamous cell carcinoma cell lines. J. Cancer Res. Clin. Oncol. 2012, 138, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Jedlinski, A.; Ansell, A.; Johansson, A.C.; Roberg, K. EGFR status and EGFR ligand expression influence the treatment response of head and neck cancer cell lines. J. Oral Pathol. Med. 2013, 42, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Jing, C.; Jin, Y.H.; You, Z.; Qiong, Q.; Jun, Z. Prognostic value of amphiregulin and epiregulin mRNA expression in metastatic colorectal cancer patients. Oncotarget 2016, 7, 55890–55899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonker, D.J.; Karapetis, C.S.; Harbison, C.; O’Callaghan, C.J.; Tu, D.; Simes, R.J.; Malone, D.P.; Langer, C.; Tebbutt, N.; Price, T.J.; et al. Epiregulin gene expression as a biomarker of benefit from cetuximab in the treatment of advanced colorectal cancer. Br. J. Cancer 2014, 110, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Kogashiwa, Y.; Inoue, H.; Kuba, K.; Araki, R.; Yasuda, M.; Nakahira, M.; Sugasawa, M. Prognostic role of epiregulin/amphiregulin expression in recurrent/metastatic head and neck cancer treated with cetuximab. Head Neck 2018, 40, 2424–2431. [Google Scholar] [CrossRef] [PubMed]
- Ianevski, A.; He, L.; Aittokallio, T.; Tang, J. SynergyFinder: A web application for analyzing drug combination dose-response matrix data. Bioinformatics 2017, 33, 2413–2415. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Molecular Subtype | AUC | |||||
|---|---|---|---|---|---|---|---|
| Afatinib | Erlotinib | Cetuximab | Gedatolisib | Cobimetinib | Trastuzumab | ||
| BHY | Basal | 3.467751696 | 4.286063175 | 4.57727436 | 3.302462568 | 4.608746135 | 4.69416933 |
| CAL27 | Basal | 3.814815676 | 4.444098626 | 5.680773203 | 3.184901695 | 3.747852593 | 5.5725818 |
| CAL33 | Basal | 3.744753557 | 4.310451267 | 4.573913673 | 2.406624054 | 4.450866901 | 5.076910559 |
| HSC2 | Basal | 4.3439915 | 4.741863856 | 5.127815501 | 2.566283234 | 4.420333039 | 4.946942339 |
| HSC3 | Basal | 3.957972571 | 4.6095489 | 5.183631235 | 2.952916999 | 3.8894347 | 5.295803542 |
| KYSE150 | Basal | 4.441065136 | 4.780103002 | 5.684081754 | 2.980021031 | 4.687959978 | 5.550264191 |
| SCC15 | Basal | 4.315046458 | 4.766164343 | 4.54432253 | 3.764267942 | 4.6050741 | 5.494692438 |
| SCC25 | Basal | 4.177447191 | 4.336103828 | 5.446338918 | 2.771755456 | 3.798183465 | 5.231781001 |
| SCC4 | Basal | 4.019642792 | 5.002304825 | 5.305982451 | 2.737492906 | 4.32226842 | 5.256117469 |
| SCC9 | Basal | 4.134972926 | 4.713456974 | 5.222760141 | 3.005065893 | 4.873465213 | 4.87701988 |
| KYSE270 | Non-Basal | 3.805157987 | 5.500896683 | 5.647765664 | 3.332916543 | 5.232659439 | 5.565586558 |
| KYSE180 | Non-Basal | 4.01491891 | 4.729217951 | 5.705364107 | 3.147861975 | 4.918800553 | 5.272014082 |
| TE6 | Non-Basal | 4.258968922 | 4.879974238 | 5.750123119 | 3.639308374 | 5.517244329 | 4.979449358 |
| TE5 | Non-Basal | 3.9331781 | 5.159062006 | 5.461107055 | 2.778634257 | 4.731403338 | 5.23314044 |
| TE10 | Non-Basal | 4.006096215 | 5.513704267 | 5.222443794 | 3.497187601 | 4.890993502 | 4.946552101 |
| KYSE140 | Non-Basal | 3.375761348 | 4.562986869 | 5.376787765 | 2.919235293 | 4.740974506 | 5.268327679 |
| KYSE510 | Non-Basal | 4.845901636 | 5.515376689 | 5.879640984 | 3.013249683 | 5.011478461 | 5.476614796 |
| RPMI2650 | Non-Basal | 4.780339567 | 5.726684777 | 6.149644208 | 3.600359879 | 5.476229419 | 5.540633467 |
| LB771HNC | Non-Basal | 4.793902186 | 5.152157915 | 5.694559888 | 3.158211825 | 5.585646234 | 5.618555173 |
| HCE4 | Non-Basal | 4.424539135 | 5.687089202 | 5.91969998 | 3.64854173 | 4.638847027 | 5.069966605 |
| KYSE70 | Non-Basal | 4.46079496 | 4.986291447 | 5.50519843 | 3.657912306 | 4.8211094 | 5.244702124 |
| TE4 | Non-Basal | 1.814915762 | 3.96069768 | 5.420791576 | 2.170670697 | 4.2463357 | 5.08505704 |
| TE14 | Non-Basal | 3.746428006 | 4.638733677 | 5.619299993 | 3.187628054 | 4.881707599 | 4.869384943 |
| TT | Non-Basal | 4.822344476 | 4.872688138 | 6.220243167 | 3.678272613 | 4.728732451 | 5.642166616 |
| HSC4 | Non-Basal | 3.655491187 | 4.180726139 | 4.651136469 | 2.751213387 | 3.871130995 | 5.136060907 |
| Cell Line | Molecular Subtype | Bliss Synergy Score | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Afatinib Cobimetinib | Afatinib Gedatosilib | Afatinib Trastuzumab | Cetuximab Cobimetinib | Cetuximab Gedatosilib | Cetuximab Trastuzumab | Erlotinib Cobimetinib | Erlotinib Gedatosilib | Erlotinib Trastuzumab | ||
| BHY | Basal | 3.777 | 5.658 | −0.348 | 7.32 | 5.569 | 13.195 | 4.33 | 4.807 | 1.701 |
| CAL27 | Basal | 6.881 | 6.787 | 6.243 | 4.252 | 3.649 | 18.943 | −0.354 | 0.38 | −0.958 |
| CAL33 | Basal | 1.89 | 0.267 | 5.684 | 0.134 | 4.621 | 12.537 | −4.981 | 2.219 | −2.872 |
| HSC2 | Basal | −4.527 | 4.403 | NA | NA | 3.066 | NA | −0.745 | 2.266 | 4.648 |
| HSC3 | Basal | −3.278 | 3.889 | 7.754 | 0.688 | 4.096 | NA | 0.517 | 1.721 | −0.263 |
| KYSE150 | Basal | −0.552 | 2.649 | NA | 5.502 | −0.469 | −0.595 | −1.648 | −0.445 | 1.263 |
| SCC4 | Basal | 5.145 | 4.867 | 2.687 | 1.146 | 0.522 | 2.728 | 2.702 | 2.979 | −1.275 |
| SCC9 | Basal | −3.499 | 6.269 | NA | −1.755 | 1.367 | NA | −1.792 | 2.185 | NA |
| SCC15 | Basal | −3.148 | 3.646 | 14.661 | 5.549 | −0.535 | NA | −1.593 | 0.959 | 6.95 |
| SCC25 | Basal | 2.244 | 3.184 | NA | 3.195 | NA | NA | −0.986 | −2.037 | NA |
| HCE4 | Non-Basal | −2.168 | −0.69 | 2.283 | 0.468 | NA | −2.688 | 4.937 | −0.317 | 4.016 |
| HSC4 | Non-Basal | −1.389 | 1.218 | 3.527 | −0.705 | 6.87 | 4.059 | −5.899 | −1.913 | −3.647 |
| KYSE70 | Non-Basal | 0.059 | 3.988 | 0.633 | −2.264 | 1.621 | 1.546 | 0.589 | −1.169 | −1.116 |
| KYSE140 | Non-Basal | 3.049 | 0.646 | 7.26 | 10.795 | NA | 6.541 | 0.466 | −2.501 | 5.82 |
| KYSE180 | Non-Basal | 1.067 | 2.902 | NA | 3.957 | 2.78 | NA | 1.759 | 3.795 | NA |
| KYSE270 | Non-Basal | −4.853 | 0.576 | −1.243 | NA | 1.923 | −2.788 | 1.111 | −1.681 | −3.602 |
| KYSE510 | Non-Basal | −0.962 | 6.481 | −1.99 | 0.86 | −0.325 | 2.142 | 1.738 | 4.474 | 6.968 |
| LB771HNC | Non-Basal | 3.722 | −0.689 | 0.706 | 7.077 | NA | NA | −3.277 | 0.658 | −1.902 |
| RPMI2650 | Non-Basal | −2.515 | 4.699 | −4.25 | NA | NA | NA | NA | NA | NA |
| TT | Non-Basal | 1.248 | 0.669 | 3.563 | 7.21 | −0.107 | 0.137 | 7.854 | 1.774 | −3.544 |
| TE4 | Non-Basal | 6.516 | 4.268 | 3.194 | −4.664 | 1.476 | −0.945 | 3.363 | 1.593 | 7.991 |
| TE5 | Non-Basal | 1.231 | 1.934 | −2.293 | −2.822 | 3.753 | 6.738 | −1.631 | 4.127 | 7.413 |
| TE6 | Non-Basal | NA | −0.834 | 5.833 | 0.528 | 2.57 | NA | −1.479 | −0.38 | 4.62 |
| TE10 | Non-Basal | 4.294 | −0.084 | −1.468 | −6.371 | −3.032 | NA | 0.085 | 0.77 | −1.671 |
| TE14 | Non-Basal | 5.072 | 2.447 | 5.155 | −5.61 | 1.86 | 5.379 | −2.549 | −1.334 | 10.887 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Job, S.; de Reyniès, A.; Heller, B.; Weiss, A.; Guérin, E.; Macabre, C.; Ledrappier, S.; Bour, C.; Wasylyk, C.; Etienne-Selloum, N.; et al. Preferential Response of Basal-Like Head and Neck Squamous Cell Carcinoma Cell Lines to EGFR-Targeted Therapy Depending on EREG-Driven Oncogenic Addiction. Cancers 2019, 11, 795. https://doi.org/10.3390/cancers11060795
Job S, de Reyniès A, Heller B, Weiss A, Guérin E, Macabre C, Ledrappier S, Bour C, Wasylyk C, Etienne-Selloum N, et al. Preferential Response of Basal-Like Head and Neck Squamous Cell Carcinoma Cell Lines to EGFR-Targeted Therapy Depending on EREG-Driven Oncogenic Addiction. Cancers. 2019; 11(6):795. https://doi.org/10.3390/cancers11060795
Chicago/Turabian StyleJob, Sylvie, Aurélien de Reyniès, Betty Heller, Amélie Weiss, Eric Guérin, Christine Macabre, Sonia Ledrappier, Cyril Bour, Christine Wasylyk, Nelly Etienne-Selloum, and et al. 2019. "Preferential Response of Basal-Like Head and Neck Squamous Cell Carcinoma Cell Lines to EGFR-Targeted Therapy Depending on EREG-Driven Oncogenic Addiction" Cancers 11, no. 6: 795. https://doi.org/10.3390/cancers11060795