The Impact of Mesothelin in the Ovarian Cancer Tumor Microenvironment

Department of Chemistry and Biochemistry, Harper Cancer Research Institute, University of Notre Dame, Notre Dame, IN 46617, USA
Cancers 2018, 10(9), 277; https://doi.org/10.3390/cancers10090277
Submission received: 8 June 2018
/
Revised: 17 August 2018
/
Accepted: 18 August 2018
/
Published: 21 August 2018
(This article belongs to the Special Issue The Tumor Microenvironment of High Grade Serous Ovarian Cancer)
{kind=link}
{kind=link}
Abstract
:Ovarian cancer is the deadliest gynecological disease among U.S. women. Poor 5-year survival rates (<30%) are due to presentation of most women at diagnosis with advanced stage disease with widely disseminated intraperitoneal metastasis. However, when diagnosed before metastatic propagation the overall 5-year survival rate is >90%. Metastasizing tumor cells grow rapidly and aggressively attach to the mesothelium of all organs within the peritoneal cavity, including the parietal peritoneum and the omentum, producing secondary lesions. In this review, the involvement of mesothelin (MSLN) in the tumor microenvironment is discussed. MSLN, a 40kDa glycoprotein that is overexpressed in many cancers including ovarian and mesotheliomas is suggested to play a role in cell survival, proliferation, tumor progression, and adherence. However, the biological function of MSLN is not fully understood as MSLN knockout mice do not present with an abnormal phenotype. Conversely, MSLN has been shown to bind to the ovarian cancer antigen, CA-125, and thought to play a role in the peritoneal diffusion of ovarian tumor cells. Although the cancer-specific expression of MSLN makes it a potential therapeutic target, more studies are needed to validate the role of MSLN in tumor metastasis.
1. Introduction
Ovarian cancer is the fifth leading cause of cancer death in U.S. women, making it the most lethal gynecological malignancy. The American Cancer Society estimates that about 22,240 new cases of ovarian cancer will be diagnosed in the United States in 2018, of which 14,070 (>60%) women will die of the disease [1]. The overall 5-year survival rate of women diagnosed with ovarian cancer is 47% and for women diagnosed with advanced stage disease, presenting with intraperitoneal metastasis, the 5-year survival rate is only 29% [1,2]. Ovarian cancer is a heterogeneous disease composed of seven histological subtypes: high-grade serous, low-grade serous, mucinous, endometrioid, clear cell, carcinosarcoma, and Brenner tumors [3]. Approximately 90% of ovarian cancers are classified as malignant epithelial ovarian carcinomas (EOCs), of which high-grade serous carcinomas (HGSC) account for 70% of tumor types [4,5,6,7]. Early signs or symptoms of ovarian cancer are often subtle and nonspecific which are frequently ignored or treated with medicine to relieve discomfort. In 50–80% of high-grade serous carcinomas, the most frequent genetic change is a p53 mutation found in tumors of all stages [8,9,10]. Mutations in BRCA1 and BRCA2, tumor suppressor genes, are found in about 50% and 70% of ovarian cancer patients with a family history of ovarian cancer, but 95% of ovarian cancer cases are sporadic [11,12,13].
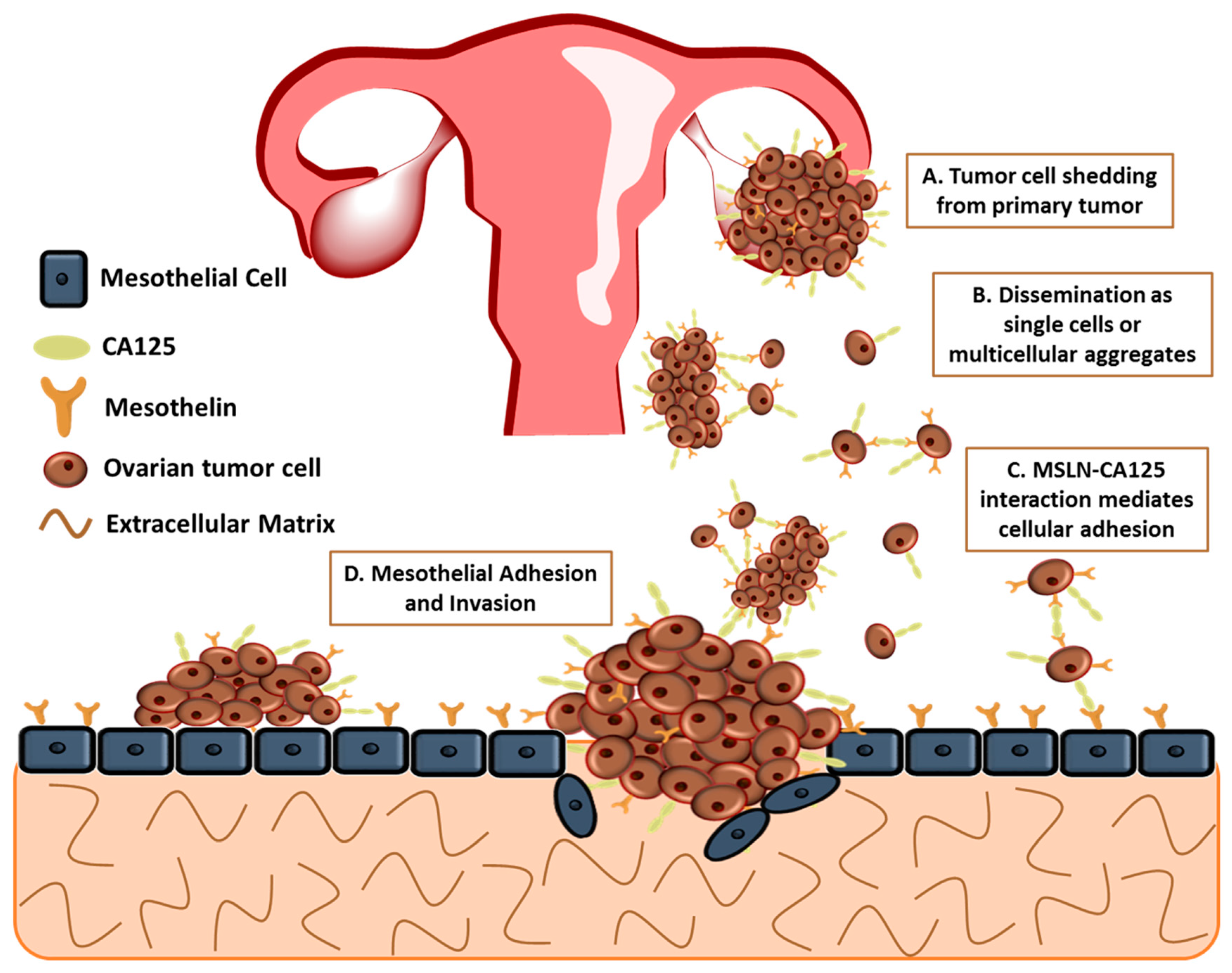
The major cause of death is due to therapy-resistant metastasis from the primary tumor to the peritoneum [14,15,16,17,18]. The lack of successful eradication of the disease can be owing to the various complex overlapping signaling networks, together with the peritoneal tumor microenvironment composed of mesothelial cells, the submesothelial matrix, and adipose. Unlike other cancers, ovarian cancer uniquely metastasizes by the detachment of tumor cells, either single or multicellular aggregates, from the primary ovarian/fallopian tube tumor instead of the classically studied pattern of hematogenous metastasis (Figure 1A,B) [15,16,18,19]. Recent studies have challenged this mode of metastasis, suggesting that hematogenous spread of ovarian cancer may play a larger role in ovarian cancer cell metastasis; however, for the purpose of this review, ovarian cancer metastasis will be discussed as direct shedding of tumor cells [20,21]. This distinctive process bypasses several steps of intra- and extravasation before metastasis to other organs [19]. These detached cells undergo epithelial to mesenchymal transition before detaching, resulting in the loss of E-cadherin, a glycoprotein located at cellular junctions, and an invasive phenotype [22]. The metastatic cells disseminate throughout the peritoneal cavity, facilitated by natural fluid flow and preferentially attach to the mesothelium that covers all the organs in the peritoneal cavity including the omentum, abdominal peritoneum and the contralateral ovary (Figure 1C,D) [14,23,24]. Proliferation of disseminated tumor cells on the omentum eventually results in the obstruction of the bowel and stomach [25,26]. It is unknown if the primary tumor prepares secondary metastatic sites, including the omentum and peritoneum, for colonization, a process that has been implicated in other cancers [19].
Currently, there are no simple screening tests available to detect ovarian cancer. However, available diagnostic testing includes pelvic examinations, transvaginal ultrasonography and serum measurements of cancer antigen-125 (CA125) [27]. Identification of additional screening strategies to accurately diagnose patients in early stages are of great need. Moreover, mesothelin, a glycoprotein expressed in mesothelial cells and overexpressed in EOCs, could be useful as both a screening biomarker as well as a therapeutic target [28]. Understanding the interaction of the tumor and mesothelium and regulating the molecules that modify the metastatic tumor microenvironment is of great importance for the development of future therapeutics.
2. CA125
CA125, a repeating peptide epitope of the mucin 16 (MUC16), is a large membrane-bound cell surface mucin, discovered in 1981 by a monoclonal antibody OC125 developed from mice immunized with human ovarian cancer cells [29]. CA125 is a heavily glycosylated type I transmembrane protein belonging to the family of tethered mucins containing both O-linked and N-linked oligosaccharides [30]. CA125 is overexpressed in many tumors of epithelial origin suggesting that it plays an important role in tumorigenesis [30,31]. CA125 is commonly used as a biomarker to monitor ovarian cancer disease progression and relapse as it is highly expressed in ovarian carcinomas yet minimally expressed in normal ovarian tissues [32,33,34]. CA125/MUC16 has been shown to inhibit cytolytic responses of human natural killer cells in ovarian cancer, therefore acting as a suppressor of the immune response directed against the ovarian tumors [35,36]. CA125 has been shown to promote cancer cell proliferation [37]. Although the role of CA125 is mainly studied in ovarian cancer, recent studies have shown that CA125 is also highly expressed in other cancers including peritoneal mesotheliomas, pancreatic, and colorectal cancer, implicating a mesothelial cell interaction [38,39,40].
3. Mesothelial Cells
All organs of the abdominal cavity are covered by the mesothelium, a monolayer of mesothelial cells covering a basement membrane composed of fibronectin, collagen I and IV and laminin [41,42]. Mesothelial cells are flattened squamous-like cells derived from the mesoderm and possess both epithelial and mesenchymal characteristics [43,44]. Mesothelial cells have well-developed cell–cell junction complexes, including tight junctions, that are critical for cell surface polarity and the formation and maintenance of a semi-permeable diffusion barrier. The mesothelium functions to provide a protective barrier as well as a frictionless interface for the free movement of organs and tissues [45]. The mesothelium also plays an important role in contributing to the homeostasis of the peritoneal cavity, fluid and cell transport, tissue repair, initiation and resolution of inflammation and possibly tumor dissemination [46,47]. In the tumor microenvironment, mesothelial cells are preconditioned by the cancer cell secretome to induce the expression of multiple pro-inflammatory factors [48]. Mesothelial cells are implicated in both epithelial-to-mesenchymal transition (EMT) and mesothelial-to-mesenchymal transition (MMT), an EMT-like process [49,50,51]. EMT is the biological process by which epithelial cells lose cell–cell adhesion and gain migratory properties and MMT is a biologic process in which mesothelial cells of the peritoneal cavity acquire a fibroblast-like phenotype, with increased migratory capabilities [50,52]. Mesothelial cells, expressing mesothelin, line the peritoneal wall and all the organs of the peritoneal cavity that is susceptible to ovarian cancer metastasis.
4. Mesothelin
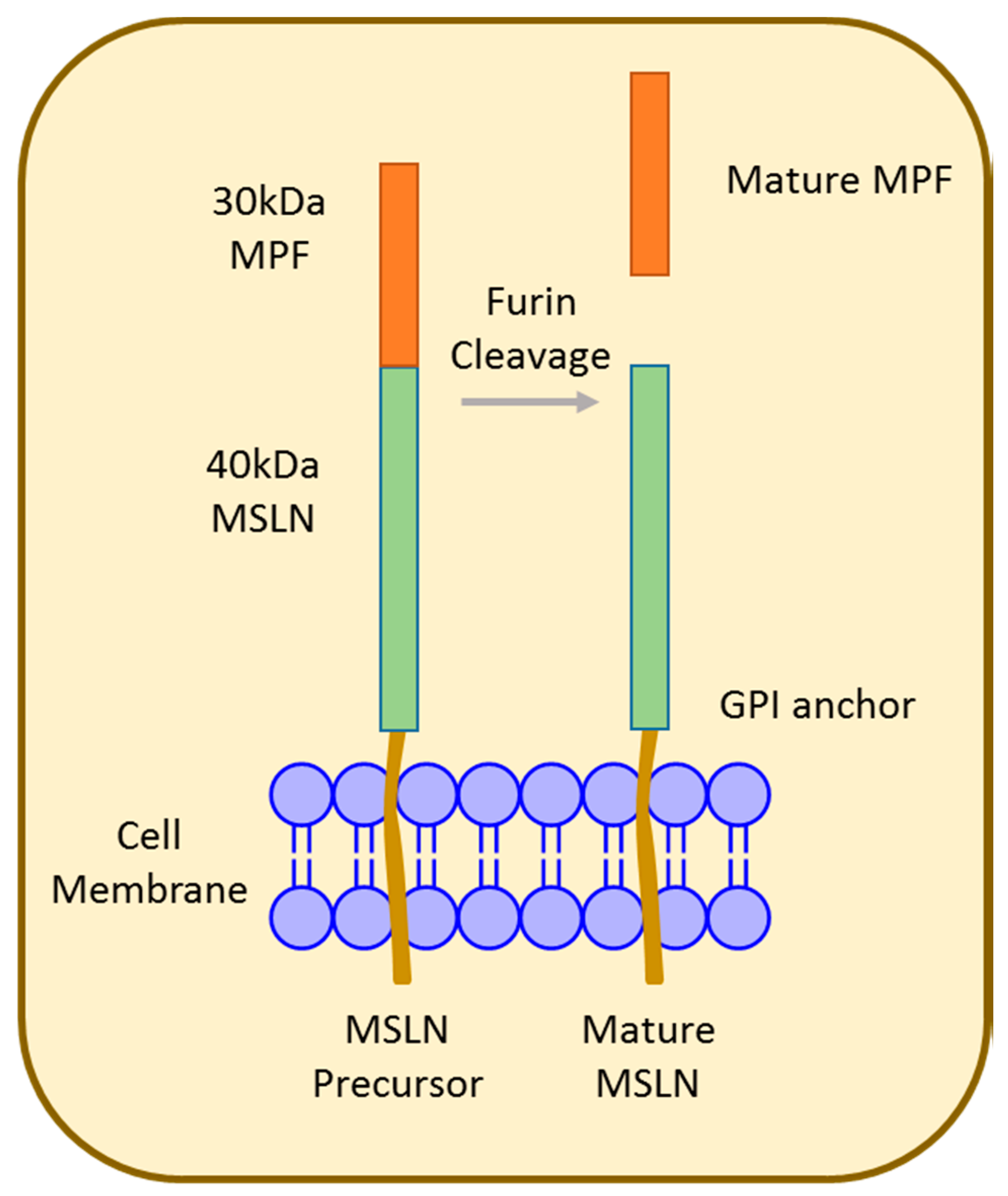
Mesothelin (MSLN), first identified in 1992 [53], is synthesized as a 70 kDa precursor that is proteolytically cleaved at Arg295, resulting in an approximately 30 kDa fragment called megakaryocyte potentiating factor (MPF) and the 40 kDa MSLN membrane-bound fragment (Figure 2) [54,55]. Both MSLN and MPF are biologically active; however, the exact function remains unknown [56]. MSLN is a glycosylphosphatidylinositol (GPI)-anchored membrane glycoprotein that is physiologically expressed at the cell surface of mesothelial cells lining the pleura, pericardium, and peritoneum [57,58]. Composed of 16 exons spanning 7733 bp, the human MSLN gene occupies approximately 8 kb located at chromosome 16 p 13.3. Alternative splicing results in the predominant variant 1 encoded by MSLN1, variant 2 (24 bp insert), and variant 3 (82 bp insert) [55,57,59,60].
Although many prediction programs have attempted to predict the three-dimensional structure of the MSLN precursor and mature MSLN, the structure still remains unknown [61]. MSLN1 was found by Hellstrom et al. to be primarily expressed at the cell surface and was also released into body fluids of patients of several tumor types. Soluble MSLN results from a cleavage of variants 1 at the C-terminal domain [60]. An 18-bp enhancer sequence, CanScript, located −65 to −46 bp 5′ of one of three transcriptional start sites in the promoter region of the MSLN gene, was identified in cancer cell lines with aberrant overexpression of MSLN. The CanScript sequence enhancer consists of two functionally putative binding motifs: the conventional MCAT element and a SP1-like element [62]. All eight nucleotides in the MCAT element were shown to be essential for its function; conversely, the SP1-like element was shown to have two mutations suggesting, that the cancer-specific expression of MSLN is thought to occur through the binding of an unknown transcription factor. Transcription factors such as KLF6 and YAP1 have been investigated but binding of these factors are not adequate for MSLN overexpression in certain cancer types [63]. Nonetheless, the essential transcriptional factor that regulates the MSLN overexpression in human cancers has not been identified.
MSLN is normally expressed in mesothelial cells in trace amounts. In contrast, MSLN is highly expressed in human cancers including 70% of ovarian cancers [54,64,65,66], mesotheliomas [54], and pancreatic adenocarcinoma [67,68] and therefore identified as a tumor-associated marker. The biological function of MSLN is not fully understood as MSLN knockout mice do not present with an abnormal phenotype, suggesting that MSLN is a non-essential protein [58]. Furthermore, MSLN is reported to play a role in cell adhesion [69], tumor progression [65,70,71,72,73], and chemoresistance [73,74,75,76]. Specifically, MSLN has been shown to have oncogenic properties by increasing ovarian cancer invasion by inducing MMP-7 through MAPK/ERK and JNK pathways and by inducing drug resistance through PI3K/AKT and MAPK/ERK signaling pathways [65,74]. Albeit, mechanisms that regulate MSLN cell-surface expression are not well understood.
5. MSLN and CA125
CA125, the ovarian cancer antigen/biomarker, has been identified as a MSLN ligand and could potentially mediate cell adhesion [69]. Rump et al. demonstrated MSLN–CA125 interaction mediates heterotypic cellular adhesion (Figure 1C) of the human ovarian cancer cell line, OVCAR3, expressing CA125 to a MSLN expressing endothelial-like cell line [69]. Additionally, Gubbels et al. established that MSLN binds to CA125 in a specific and N-linked glycan-dependent manner, thus CA125-expressing ovarian tumor cells could bind specifically to the mesothelin-expressing peritoneal lining (Figure 1D) [77]. The N-linked oligosaccharides of CA125 are necessary for the binding to MSLN with MSLN having a strong affinity to CA125 with an apparent dissociation constant (Kd) of 5 nM [77,78,79]. Consequently, MSLN:CA125-dependent cell attachment may play an important role in the peritoneal implantation of ovarian tumor cells [54,80]. The MSLN:CA125 role in cell attachment is supported by work from Bruney et al., demonstrating the overexpression of membrane type 1 matrix metalloproteinase (MT1-MMP) in human ovarian cancer cells (OVCA433-MT)-decreased cell surface expression of CA125/MUC16, subsequently increasing CA125/MUC16 ectodomain shedding, resulting in the release of CA125 from the cell surface. Additionally, there was decreased adhesion of OVCA433-MT to human mesothelial cells (LP9) and to intact peritoneal explants, suggesting the importance of MSLN:CA125 initial adhesion of [81]. After initial attachment of ovarian cancer cells to the peritoneal mesothelium, the co-overexpression of both MSLN and CA125 can lead to recruitment of other ovarian cancer cells being sloughed off from the primary site (Figure 1B,C) [82]. Therefore, the tumor load at secondary sites could be a combination of excessive proliferation and adhesion of circulating single or multicellular aggregates in peritoneal ascites fluid [77,83]. Conversely, the exact function of MSLN in tumor progression remains unclear [84]; however, understanding the importance of CA125:mesothelin binding may lead to novel therapies to control ovarian peritoneal metastasis.
6. Targeting MSLN
Clarifying the function of MSLN will enhance its clinical application in ovarian cancer, including early detection, chemo-response, prognosis and therapeutic targeting. Several features of MSLN make it a useful candidate for cancer therapy, including that it is well-internalized, enabling it to be a good target for immunotoxins [85]. Additionally, MSLN is actively shed from the cell surface generating a pool of antigens in ascites or blood circulation allowing for the quantification of circulating serum MSLN levels potentially used for diagnosis of ovarian cancer patients [28,86,87,88]. The use of MSLN as a plasma biomarker has been investigated by several groups using blood ELISA tests and demonstrated that serum MSLN levels decrease after surgical therapy and, therefore, may be useful in monitoring treatment response in MSLN expression cancers [86,89]. Pools of antigens, from shed MSLN, in the tumor interstitial space will unavoidably interact with a targeting agent during tumor dissemination [60,85,87]. The first identified sheddase, TNF-α converting enzyme (TACE) was shown to mediate MSLN shedding. TACE is a transmembrane glycoprotein, known for its role in releasing EGFR ligands from the cell surface, therefore regulating the activation of the EGFR pathway [60,90]. Tumor targeting is a complex process and, furthermore, modulation of MSLN shedding could have an influence on drug kinetics in both circulation and tumor tissue. However, shedding is not the only way MSLN could be modulated. The expression levels of MSLN could potentially be regulated similarly to other antigens by trogocytosis [91] or antigen masking [92]; however, the role of these antigens remains to be elucidated. Furthermore, MSLN is expressed in dispensable mesothelial cells so the risk of non-specific toxicity is decreased.
6.1. Molecular Imaging for the Detection of MSLN
Mesothelin has recently been investigated as a target for molecular imaging probes. These probes are designed to guide antibody-based treatments that can be used to assess tumor uptake, response to treatment and the distribution in primary tumors and secondary sites. Prantner et al. identified and characterized an antimesothelin nanobody (NbG3a) used for in vitro diagnostic applications [93]. Further studies from the same group established the potential use of NbG3a for a novel molecular imaging probe with promising results for human imaging and therapeutic applications [94]. Terwisscha van Scheltinga et al. investigated the use of an antibody–drug conjugate (anti-mesothelin antibody-monomethyl auristatin E) coupled to molecular imaging with 89Zr immuno-positive emission tomography (PET). Using this technique, quantitative immuno-PET measurement of relative antibody uptake was determined to correlate with tumor growth inhibition [95]. Furthermore, non-antibody protein scaffolds have successfully been engineered to bind to mesothelin with high affinity [96]. Unlike antibodies that are large in size and have slow clearance from circulation, non-antibody protein scaffolds have demonstrated specific binding to identify tumors expressing the molecular target in murine models [97,98,99] and have demonstrated promising results in both preclinical and clinical evaluations [100]. The use of these techniques demonstrates the translational potential of MSLN.
6.2. Clinical Trials
There are many clinical trials testing MSLN-targeting agents using strategies such as antibody-based immunotoxins such as SS1P, consisting of an anti-MLSN Fv obtained from a phage display library of immunized mice with recombinant MSLN fused to a truncated form of the Pseudomonas Exotoxin PE38 that mediates cell death. The mechanism of action of an immunotoxin is threefold. First, the immunotoxin binds to cell-bound MSLN; second, this complex is internalized by endocytosis, undergoes retrograde transport to the endoplasmic reticulum and the PE portion is translocated to the cytosol; and third, the PE catalyzes ADP-ribosylation of the elongation factor-2, halting protein synthesis and activating apoptosis [85,101]. There have been two Phase I clinical trials with different modes of administration using either continuous infusion or as bolus intravenous infusions in mesotheliomas, ovarian, and pancreatic cancers. Continuous infusion was well tolerated and showed modest clinical activity; however, there was advantage seen over bolus dosing [102,103]. Additionally, there is a high affinity chimeric antibody, amatuximab (MORAb-009), with high affinity and specificity for mesothelin that is under investigation in clinical trials. Amatuximab works by inducing antibody-dependent cellular cytoxicity [104]. It was observed that upon treatment with amatuximab, patients had an increase in CA125 levels suggesting that amatuximab interferes with the MSLN:CA125 interaction [105]. A tumor vaccine CRS-207 utilizing a live attenuated strain of bacterium Listeria monocytogenes (Lm) expressing human MSLN has shown good tolerance and MSLN-specific T-cell response in a phase I study of safety clinical trial. This phase I study not only demonstrated that vaccines are safe and tolerable but also showed that a tumor antigen-modified Lm can induce tumor antigen-specific T-cell responses in patients with advanced cancer, suggesting that further evaluation of Lm vaccine as a candidate biomarker of improved clinical outcomes is needed [106]. A two-part phase I/II trial is underway using combination therapy with CRS-207, epacadostat, and pembrolizumab (keytruda) in patients with platinum-resistant ovarian, fallopian tube, and peritoneal cancers using different combinations of the three treatments (ClinicalTrials.gov Identifier NCT02575807). Antibody–drug conjugates is another strategy used to target MSLN. An ongoing phase I clinical trial with anetumab ravtansine (BAY94-9343) to determine the safety and maximum tolerated dose for patients with advanced solid tumors including ovarian carcinoma and mesothelioma opened in 2011 (ClinicalTrials.gov Identifier NCT01439152). Anetumab ravtansine consists of the fully human anti-MSLN IgG1 linked to a potent tubulin-binding drug, DM4. In preclinical trials, anetumab ravtansine inhibited both subcutaneous and orthotopic tumor growth in xenograft models of ovarian, pancreatic, and mesothelioma cancers [107]. In patients with recurrent MSLN-expressing platinum-resistant recurrent ovarian, fallopian tube or primary peritoneal cancer, a phase Ib clinical trial to determine the maximum tolerated dose of anetumab ravtansine that could be safely combined with pegylated liposomal doxorubicin is underway (ClinicalTrials.gov Identifier NCT02751918). Several ongoing clinical trials are utilizing anti-MSLN CAR-modified T cells as MSLN targeting agent. The T cells are obtained by apheresis and introduced to a temporary gene which will cause them to make a new type of antibody that will attach to MSLN. Once attached, the cells will become activated and stimulate the host immune system to attack the MSLN-expressing cells [108]. The above clinical trials have confirmed that targeting MSLN could be beneficial in improving existing therapeutic options for patients diagnosed with a MSLN-expressing cancer, including ovarian cancer.
7. Conclusions
Ovarian cancer is the deadliest gynecological malignancy among U.S. women and is often diagnosed at a late stage when the disease has metastasized into the peritoneal cavity. Mesothelin, a glycoprotein normally expressed in mesothelial cells, is highly expressed in several cancers including ovarian, pancreatic, and mesotheliomas. It has been shown that MSLN binds to the ovarian cancer biomarker CA125 and this interaction plays a role in the peritoneal metastasis of ovarian cancer. The differential expression of mesothelin in normal and cancer tissues makes it a promising candidate for targeted therapeutics. Several candidate immunotherapies targeting MSLN are in ongoing clinical trials. New strategies to disrupt the MSLN:CA125 interaction are emerging. Although MSLN is implicated in many cancers, the role of MSLN is still poorly understood warranting further investigation and clinical trial studies. Future advances in ovarian cancer therapy depend on novel treatment mechanisms in combination with current chemotherapeutic approaches that will result in cytotoxicity, inhibition of metastasis and angiogenesis, and increasing the immunological detection of tumors. Further mechanistic studies on MSLN are needed to validate the potential role of MSLN in tumor metastasis that possibly will provide insight for effective MSLN-targeting therapies for several cancers.
Funding
This work was supported by grant KO1CA218305 from the National Institutes of Health/National Cancer Institute.
Acknowledgments
I would like to thank members of the Stack lab at the University of Notre Dame for help with editing this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Peres, L.C.; Cushing-Haugen, K.L.; Kobel, M.; Harris, H.R.; Berchuck, A.; Rossing, M.A.; Schildkraut, J.M.; Doherty, J.A. Invasive epithelial ovarian cancer survival by histotype and disease stage. J. Natl. Cancer Inst. 2018. [Google Scholar] [CrossRef] [PubMed]
- Meinhold-Heerlein, I.; Fotopoulou, C.; Harter, P.; Kurzeder, C.; Mustea, A.; Wimberger, P.; Hauptmann, S.; Sehouli, J. The new who classification of ovarian, fallopian tube, and primary peritoneal cancer and its clinical implications. Arch. Gynecol. Obstet. 2016, 293, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Auersperg, N.; Wong, A.S.; Choi, K.C.; Kang, S.K.; Leung, P.C. Ovarian surface epithelium: Biology, endocrinology, and pathology. Endocr. Rev. 2001, 22, 255–288. [Google Scholar] [CrossRef] [PubMed]
- Choi, J. Ovarian epithelial cancer: Etiology and pathogenesis. Biowave 2008, 10, 32. [Google Scholar]
- Auersperg, N. The origin of ovarian cancers–hypotheses and controversies. Front. Biosci. (Sch. Ed.) 2013, 5, 709–719. [Google Scholar] [CrossRef]
- Mutch, D.G.; Prat, J. 2014 FIGO staging for ovarian, fallopian tube and peritoneal cancer. Gynecol. Oncol. 2014, 133, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Kohler, M.F.; Marks, J.R.; Wiseman, R.W.; Jacobs, I.J.; Davidoff, A.M.; Clarke-Pearson, D.L.; Soper, J.T.; Bast, R.C., Jr.; Berchuck, A. Spectrum of mutation and frequency of allelic deletion of the p53 gene in ovarian cancer. J. Natl. Cancer Inst. 1993, 85, 1513–1519. [Google Scholar] [CrossRef] [PubMed]
- Singer, G.; Stohr, R.; Cope, L.; Dehari, R.; Hartmann, A.; Cao, D.F.; Wang, T.L.; Kurman, R.J.; Shih, I.M. Patterns of p53 mutations separate ovarian serous borderline tumors and low- and high-grade carcinomas and provide support for a new model of ovarian carcinogenesis: A mutational analysis with immunohistochemical correlation. Am. J. Surg. Pathol. 2005, 29, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Milner, B.J.; Allan, L.A.; Eccles, D.M.; Kitchener, H.C.; Leonard, R.C.; Kelly, K.F.; Parkin, D.E.; Haites, N.E. P53 mutation is a common genetic event in ovarian carcinoma. Cancer Res. 1993, 53, 2128–2132. [Google Scholar] [PubMed]
- Struewing, J.P.; Hartge, P.; Wacholder, S.; Baker, S.M.; Berlin, M.; McAdams, M.; Timmerman, M.M.; Brody, L.C.; Tucker, M.A. The risk of cancer associated with specific mutations of brca1 and brca2 among ashkenazi jews. N. Engl. J. Med. 1997, 336, 1401–1408. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, A.; Pharoah, P.D.; Narod, S.; Risch, H.A.; Eyfjord, J.E.; Hopper, J.L.; Loman, N.; Olsson, H.; Johannsson, O.; Borg, A.; et al. Average risks of breast and ovarian cancer associated with BRCA1 or BRCA2 mutations detected in case series unselected for family history: A combined analysis of 22 studies. Am. J. Hum. Genet. 2003, 72, 1117–1130. [Google Scholar] [CrossRef] [PubMed]
- Mavaddat, N.; Peock, S.; Frost, D.; Ellis, S.; Platte, R.; Fineberg, E.; Evans, D.G.; Izatt, L.; Eeles, R.A.; Adlard, J.; et al. Cancer risks for BRCA1 and BRCA2 mutation carriers: Results from prospective analysis of embrace. J. Natl. Cancer Inst. 2013, 105, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.S.; Agarwal, R.; Kaye, S.B. Mechanisms of transcoelomic metastasis in ovarian cancer. Lancet Oncol. 2006, 7, 925–934. [Google Scholar] [CrossRef]
- Burleson, K.M.; Casey, R.C.; Skubitz, K.M.; Pambuccian, S.E.; Oegema, T.R., Jr.; Skubitz, A.P. Ovarian carcinoma ascites spheroids adhere to extracellular matrix components and mesothelial cell monolayers. Gynecol. Oncol. 2004, 93, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Burleson, K.M.; Hansen, L.K.; Skubitz, A.P. Ovarian carcinoma spheroids disaggregate on type i collagen and invade live human mesothelial cell monolayers. Clin. Exp. Metastasis 2004, 21, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Carmignani, C.P.; Sugarbaker, T.A.; Bromley, C.M.; Sugarbaker, P.H. Intraperitoneal cancer dissemination: Mechanisms of the patterns of spread. Cancer Metastasis Rev. 2003, 22, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Buy, J.N.; Moss, A.A.; Ghossain, M.A.; Sciot, C.; Malbec, L.; Vadrot, D.; Paniel, B.J.; Decroix, Y. Peritoneal implants from ovarian tumors: Ct findings. Radiology 1988, 169, 691–694. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.P.; Massague, J. Cancer metastasis: Building a framework. Cell 2006, 127, 679–695. [Google Scholar] [CrossRef] [PubMed]
- Pradeep, S.; Kim, S.W.; Wu, S.Y.; Nishimura, M.; Chaluvally-Raghavan, P.; Miyake, T.; Pecot, C.V.; Kim, S.J.; Choi, H.J.; Bischoff, F.Z.; et al. Hematogenous metastasis of ovarian cancer: Rethinking mode of spread. Cancer Cell 2014, 26, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Coffman, L.G.; Burgos-Ojeda, D.; Wu, R.; Cho, K.; Bai, S.; Buckanovich, R.J. New models of hematogenous ovarian cancer metastasis demonstrate preferential spread to the ovary and a requirement for the ovary for abdominal dissemination. Transl. Res. 2016, 175, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyers, M.A.; Oliphant, M.; Berne, A.S.; Feldberg, M.A. The peritoneal ligaments and mesenteries: Pathways of intraabdominal spread of disease. Radiology 1987, 163, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Coakley, F.V.; Hricak, H. Imaging of peritoneal and mesenteric disease: Key concepts for the clinical radiologist. Clin. Radiol. 1999, 54, 563–574. [Google Scholar] [CrossRef]
- White, E.A.; Kenny, H.A.; Lengyel, E. Three-dimensional modeling of ovarian cancer. Adv. Drug Deliv. Rev. 2014, 79–80, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Lengyel, E. Ovarian cancer development and metastasis. Am. J. Pathol. 2010, 177, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Cannistra, S.A. Cancer of the ovary. N. Engl. J. Med. 2004, 351, 2519–2529. [Google Scholar] [CrossRef] [PubMed]
- Scholler, N.; Fu, N.; Yang, Y.; Ye, Z.; Goodman, G.E.; Hellstrom, K.E.; Hellstrom, I. Soluble member(s) of the mesothelin/megakaryocyte potentiating factor family are detectable in sera from patients with ovarian carcinoma. Proc. Natl. Acad. Sci. USA 1999, 96, 11531–11536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bast, R.C., Jr.; Feeney, M.; Lazarus, H.; Nadler, L.M.; Colvin, R.B.; Knapp, R.C. Reactivity of a monoclonal antibody with human ovarian carcinoma. J. Clin. Investig. 1981, 68, 1331–1337. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, T.J.; Beard, J.B.; Underwood, L.J.; Shigemasa, K. The ca 125 gene: A newly discovered extension of the glycosylated N-terminal domain doubles the size of this extracellular superstructure. Tumour Biol. 2002, 23, 154–169. [Google Scholar] [PubMed]
- Yin, B.W.; Lloyd, K.O. Molecular cloning of the ca125 ovarian cancer antigen: Identification as a new mucin, muc16. J. Biol. Chem. 2001, 276, 27371–27375. [Google Scholar] [CrossRef] [PubMed]
- Gubbels, J.A.; Claussen, N.; Kapur, A.K.; Connor, J.P.; Patankar, M.S. The detection, treatment, and biology of epithelial ovarian cancer. J. Ovarian Res. 2010, 3, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bast, R.C., Jr.; Klug, T.L.; St John, E.; Jenison, E.; Niloff, J.M.; Lazarus, H.; Berkowitz, R.S.; Leavitt, T.; Griffiths, C.T.; Parker, L.; et al. A radioimmunoassay using a monoclonal antibody to monitor the course of epithelial ovarian cancer. N. Engl. J. Med. 1983, 309, 883–887. [Google Scholar] [CrossRef] [PubMed]
- Yin, B.W.; Dnistrian, A.; Lloyd, K.O. Ovarian cancer antigen ca125 is encoded by the muc16 mucin gene. Int. J. Cancer 2002, 98, 737–740. [Google Scholar] [CrossRef] [PubMed]
- Patankar, M.S.; Jing, Y.; Morrison, J.C.; Belisle, J.A.; Lattanzio, F.A.; Deng, Y.; Wong, N.K.; Morris, H.R.; Dell, A.; Clark, G.F. Potent suppression of natural killer cell response mediated by the ovarian tumor marker ca125. Gynecol. Oncol. 2005, 99, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Senapati, S.; Das, S.; Batra, S.K. Mucin-interacting proteins: From function to therapeutics. Trends Biochem. Sci. 2010, 35, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Reinartz, S.; Failer, S.; Schuell, T.; Wagner, U. Ca125 (muc16) gene silencing suppresses growth properties of ovarian and breast cancer cells. Eur. J. Cancer 2012, 48, 1558–1569. [Google Scholar] [CrossRef] [PubMed]
- Streppel, M.M.; Vincent, A.; Mukherjee, R.; Campbell, N.R.; Chen, S.H.; Konstantopoulos, K.; Goggins, M.G.; Van Seuningen, I.; Maitra, A.; Montgomery, E.A. Mucin 16 (cancer antigen 125) expression in human tissues and cell lines and correlation with clinical outcome in adenocarcinomas of the pancreas, esophagus, stomach, and colon. Hum. Pathol. 2012, 43, 1755–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, A.; Hirono, S.; Tani, M.; Kawai, M.; Okada, K.; Miyazawa, M.; Kitahata, Y.; Nakamura, Y.; Noda, T.; Yokoyama, S.; et al. Coexpression of muc16 and mesothelin is related to the invasion process in pancreatic ductal adenocarcinoma. Cancer Sci. 2012, 103, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Baratti, D.; Kusamura, S.; Martinetti, A.; Seregni, E.; Oliva, D.G.; Laterza, B.; Deraco, M. Circulating ca125 in patients with peritoneal mesothelioma treated with cytoreductive surgery and intraperitoneal hyperthermic perfusion. Ann. Surg. Oncol. 2007, 14, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Yeung, T.L.; Leung, C.S.; Yip, K.P.; Au Yeung, C.L.; Wong, S.T.; Mok, S.C. Cellular and molecular processes in ovarian cancer metastasis. Am. J. Physiol. Cell Physiol. 2015, 309, C444–C456. [Google Scholar] [CrossRef] [PubMed]
- Kenny, H.A.; Chiang, C.Y.; White, E.A.; Schryver, E.M.; Habis, M.; Romero, I.L.; Ladanyi, A.; Penicka, C.V.; George, J.; Matlin, K.; et al. Mesothelial cells promote early ovarian cancer metastasis through fibronectin secretion. J. Clin. Investig. 2014, 124, 4614–4628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Baal, J.O.; Van de Vijver, K.K.; Nieuwland, R.; van Noorden, C.J.; van Driel, W.J.; Sturk, A.; Kenter, G.G.; Rikkert, L.G.; Lok, C.A. The histophysiology and pathophysiology of the peritoneum. Tissue Cell 2017, 49, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Ferrandez-Izquierdo, A.; Navarro-Fos, S.; Gonzalez-Devesa, M.; Gil-Benso, R.; Llombart-Bosch, A. Immunocytochemical typification of mesothelial cells in effusions: In vivo and in vitro models. Diagn. Cytopathol. 1994, 10, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Mutsaers, S.E. The mesothelial cell. Int. J. Biochem. Cell Biol. 2004, 36, 9–16. [Google Scholar] [CrossRef]
- Roth, J. Ultrahistochemical demonstration of saccharide components of complex carbohydrates at the alveolar cell surface and at the mesothelial cell surface of the pleura visceralis of mice by means of concanavalin A. Exp. Pathol. (Jena) 1973, 8, 157–167. [Google Scholar] [PubMed]
- Yung, S.; Chan, T.M. Pathophysiology of the peritoneal membrane during peritoneal dialysis: The role of hyaluronan. J. Biomed. Biotechnol. 2011, 2011, 180594. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Xiao, Y.J.; Singh, L.S.; Zhao, X.; Zhao, Z.; Feng, L.; Rose, T.M.; Prestwich, G.D.; Xu, Y. Lysophosphatidic acid is constitutively produced by human peritoneal mesothelial cells and enhances adhesion, migration, and invasion of ovarian cancer cells. Cancer Res. 2006, 66, 3006–3014. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Sun, B.; Zhao, X.; Gu, Q.; Dong, X.; Mo, J.; Sun, T.; Wang, J.; Sun, R.; Liu, Y. Hypoxia promotes vasculogenic mimicry formation by inducing epithelial-mesenchymal transition in ovarian carcinoma. Gynecol. Oncol. 2014, 133, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Yanez-Mo, M.; Lara-Pezzi, E.; Selgas, R.; Ramirez-Huesca, M.; Dominguez-Jimenez, C.; Jimenez-Heffernan, J.A.; Aguilera, A.; Sanchez-Tomero, J.A.; Bajo, M.A.; Alvarez, V.; et al. Peritoneal dialysis and epithelial-to-mesenchymal transition of mesothelial cells. N. Engl. J. Med. 2003, 348, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, P.; Jimenez-Heffernan, J.A.; Rynne-Vidal, A.; Perez-Lozano, M.L.; Gilsanz, A.; Ruiz-Carpio, V.; Reyes, R.; Garcia-Bordas, J.; Stamatakis, K.; Dotor, J.; et al. Carcinoma-associated fibroblasts derive from mesothelial cells via mesothelial-to-mesenchymal transition in peritoneal metastasis. J. Pathol. 2013, 231, 517–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rynne-Vidal, A.; Au-Yeung, C.L.; Jimenez-Heffernan, J.A.; Perez-Lozano, M.L.; Cremades-Jimeno, L.; Barcena, C.; Cristobal-Garcia, I.; Fernandez-Chacon, C.; Yeung, T.L.; Mok, S.C.; et al. Mesothelial-to-mesenchymal transition as a possible therapeutic target in peritoneal metastasis of ovarian cancer. J. Pathol. 2017, 242, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.; Pastan, I.; Willingham, M.C. Isolation and characterization of a monoclonal antibody, k1, reactive with ovarian cancers and normal mesothelium. Int. J. Cancer 1992, 50, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.; Pastan, I. Molecular cloning of mesothelin, a differentiation antigen present on mesothelium, mesotheliomas, and ovarian cancers. Proc. Natl. Acad. Sci. USA 1996, 93, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Bera, T.; Pastan, I. Mesothelin: A new target for immunotherapy. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2004, 10, 3937–3942. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Yamamura, Y.; Konishi, E.; Ueda, K.; Kojima, T.; Hattori, K.; Oheda, M.; Imai, N.; Taniguchi, Y.; Tamura, M.; et al. Characterization, molecular cloning and expression of megakaryocyte potentiating factor. Stem Cells 1996, 14, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Scholler, N. Mesothelin. In Encyclopedia of Cancer; Schwab, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 2241–2245. [Google Scholar]
- Bera, T.K.; Pastan, I. Mesothelin is not required for normal mouse development or reproduction. Mol. Cell. Biol. 2000, 20, 2902–2906. [Google Scholar] [CrossRef] [PubMed]
- Muminova, Z.E.; Strong, T.V.; Shaw, D.R. Characterization of human mesothelin transcripts in ovarian and pancreatic cancer. BMC Cancer 2004, 4, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellstrom, I.; Raycraft, J.; Kanan, S.; Sardesai, N.Y.; Verch, T.; Yang, Y.; Hellstrom, K.E. Mesothelin variant 1 is released from tumor cells as a diagnostic marker. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1014–1020. [Google Scholar] [CrossRef] [PubMed]
- Sathyanarayana, B.K.; Hahn, Y.; Patankar, M.S.; Pastan, I.; Lee, B. Mesothelin, stereocilin, and otoancorin are predicted to have superhelical structures with arm-type repeats. BMC Struct. Biol. 2009, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Hucl, T.; Brody, J.R.; Gallmeier, E.; Iacobuzio-Donahue, C.A.; Farrance, I.K.; Kern, S.E. High cancer-specific expression of mesothelin (MSLN) is attributable to an upstream enhancer containing a transcription enhancer factor dependent mcat motif. Cancer Res. 2007, 67, 9055–9065. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.R.; Patel, K.; Paun, B.C.; Kern, S.E. Structural analysis of the cancer-specific promoter in mesothelin and in other genes overexpressed in cancers. J. Biol. Chem. 2011, 286, 11960–11969. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Kreitman, R.J.; Pastan, I.; Willingham, M.C. Localization of mesothelin in epithelial ovarian cancer. Appl. Immunohistochem. Mol. Morphol. 2005, 13, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Chen, C.A.; Chen, P.J.; Chiang, Y.C.; Chen, Y.L.; Mao, T.L.; Lin, H.W.; Lin Chiang, W.H.; Cheng, W.F. Mesothelin enhances invasion of ovarian cancer by inducing mmp-7 through Mapk/Erk and Jnk pathways. Biochem. J. 2012, 442, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Yen, M.J.; Hsu, C.Y.; Mao, T.L.; Wu, T.C.; Roden, R.; Wang, T.L.; Shih Ie, M. Diffuse mesothelin expression correlates with prolonged patient survival in ovarian serous carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2006, 12, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Bodempudi, V.; Liu, Z.; Borrego-Diaz, E.; Yamoutpoor, F.; Meyer, A.; Woo, R.A.; Pan, W.; Dudek, A.Z.; Olyaee, M.S.; et al. Inhibition of mesothelin as a novel strategy for targeting cancer cells. PLoS ONE 2012, 7, e33214. [Google Scholar] [CrossRef] [PubMed]
- Argani, P.; Iacobuzio-Donahue, C.; Ryu, B.; Rosty, C.; Goggins, M.; Wilentz, R.E.; Murugesan, S.R.; Leach, S.D.; Jaffee, E.; Yeo, C.J.; et al. Mesothelin is overexpressed in the vast majority of ductal adenocarcinomas of the pancreas: Identification of a new pancreatic cancer marker by serial analysis of gene expression (sage). Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2001, 7, 3862–3868. [Google Scholar]
- Rump, A.; Morikawa, Y.; Tanaka, M.; Minami, S.; Umesaki, N.; Takeuchi, M.; Miyajima, A. Binding of ovarian cancer antigen ca125/muc16 to mesothelin mediates cell adhesion. J. Biol. Chem. 2004, 279, 9190–9198. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Bharadwaj, U.; Zhang, R.; Zhang, S.; Mu, H.; Fisher, W.E.; Brunicardi, F.C.; Chen, C.; Yao, Q. Mesothelin is a malignant factor and therapeutic vaccine target for pancreatic cancer. Mol. Cancer Ther. 2008, 7, 286–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharadwaj, U.; Li, M.; Chen, C.; Yao, Q. Mesothelin-induced pancreatic cancer cell proliferation involves alteration of cyclin e via activation of signal transducer and activator of transcription protein 3. Mol. Cancer Res. 2008, 6, 1755–1765. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, U.; Marin-Muller, C.; Li, M.; Chen, C.; Yao, Q. Mesothelin overexpression promotes autocrine il-6/sil-6r trans-signaling to stimulate pancreatic cancer cell proliferation. Carcinogenesis 2011, 32, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, U.; Marin-Muller, C.; Li, M.; Chen, C.; Yao, Q. Mesothelin confers pancreatic cancer cell resistance to tnf-alpha-induced apoptosis through akt/pi3k/nf-kappab activation and il-6/mcl-1 overexpression. Mol. Cancer 2011, 10, 106. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Chen, C.A.; Hsieh, C.Y.; Lee, C.N.; Su, Y.N.; Hu, Y.H.; Cheng, W.F. Mesothelin inhibits paclitaxel-induced apoptosis through the pi3k pathway. Biochem. J. 2009, 424, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.F.; Huang, C.Y.; Chang, M.C.; Hu, Y.H.; Chiang, Y.C.; Chen, Y.L.; Hsieh, C.Y.; Chen, C.A. High mesothelin correlates with chemoresistance and poor survival in epithelial ovarian carcinoma. Br. J. Cancer 2009, 100, 1144–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uehara, N.; Matsuoka, Y.; Tsubura, A. Mesothelin promotes anchorage-independent growth and prevents anoikis via extracellular signal-regulated kinase signaling pathway in human breast cancer cells. Mol. Cancer Res. 2008, 6, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Gubbels, J.A.; Belisle, J.; Onda, M.; Rancourt, C.; Migneault, M.; Ho, M.; Bera, T.K.; Connor, J.; Sathyanarayana, B.K.; Lee, B.; et al. Mesothelin-muc16 binding is a high affinity, n-glycan dependent interaction that facilitates peritoneal metastasis of ovarian tumors. Mol. Cancer 2006, 5, 50. [Google Scholar] [CrossRef] [PubMed]
- Kui Wong, N.; Easton, R.L.; Panico, M.; Sutton-Smith, M.; Morrison, J.C.; Lattanzio, F.A.; Morris, H.R.; Clark, G.F.; Dell, A.; Patankar, M.S. Characterization of the oligosaccharides associated with the human ovarian tumor marker ca125. J. Biol. Chem. 2003, 278, 28619–28634. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, O.; Gong, L.; Zhang, J.; Hansen, J.K.; Hassan, R.; Lee, B.; Ho, M. A binding domain on mesothelin for ca125/muc16. J. Biol. Chem. 2009, 284, 3739–3749. [Google Scholar] [CrossRef] [PubMed]
- Scholler, N.; Garvik, B.; Hayden-Ledbetter, M.; Kline, T.; Urban, N. Development of a ca125-mesothelin cell adhesion assay as a screening tool for biologics discovery. Cancer Lett. 2007, 247, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Bruney, L.; Conley, K.C.; Moss, N.M.; Liu, Y.; Stack, M.S. Membrane-type i matrix metalloproteinase-dependent ectodomain shedding of mucin16/ca-125 on ovarian cancer cells modulates adhesion and invasion of peritoneal mesothelium. Biol. Chem. 2014, 395, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Qian, M.; Ho, M. The role of mesothelin in tumor progression and targeted therapy. Anticancer Agents Med. Chem. 2013, 13, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Bast, R.C., Jr.; Badgwell, D.; Lu, Z.; Marquez, R.; Rosen, D.; Liu, J.; Baggerly, K.A.; Atkinson, E.N.; Skates, S.; Zhang, Z.; et al. New tumor markers: Ca125 and beyond. Int. J. Gynecol. Cancer 2005, 15, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Hilliard, T.; Iwamoto, K.; Loughran, E.; Liu, Y.; Yang, J.; Asem, M.; Tarwater, L.; Klymenko, Y.; Johnson, J.; Shi, Z.; et al. Mesothelin expression increases ovarian cancer metastasis in the peritoneal microenvironment. Cancer Lett. 2018, in press. [Google Scholar]
- Pastan, I.; Zhang, Y. Modulating mesothelin shedding to improve therapy. Oncotarget 2012, 3, 114–115. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Remaley, A.T.; Sampson, M.L.; Zhang, J.; Cox, D.D.; Pingpank, J.; Alexander, R.; Willingham, M.; Pastan, I.; Onda, M. Detection and quantitation of serum mesothelin, a tumor marker for patients with mesothelioma and ovarian cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2006, 12, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.; Onda, M.; Wang, Q.C.; Hassan, R.; Pastan, I.; Lively, M.O. Mesothelin is shed from tumor cells. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1751. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Cheng, W.F.; Lee, C.N.; Su, Y.N.; Chien, S.C.; Tzeng, Y.L.; Hsieh, C.Y.; Chen, C.A. Serum mesothelin in epithelial ovarian carcinoma: A new screening marker and prognostic factor. Anticancer Res. 2006, 26, 4721–4728. [Google Scholar] [PubMed]
- McIntosh, M.W.; Drescher, C.; Karlan, B.; Scholler, N.; Urban, N.; Hellstrom, K.E.; Hellstrom, I. Combining ca 125 and smr serum markers for diagnosis and early detection of ovarian carcinoma. Gynecol. Oncol. 2004, 95, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chertov, O.; Zhang, J.; Hassan, R.; Pastan, I. Cytotoxic activity of immunotoxin ss1p is modulated by tace-dependent mesothelin shedding. Cancer Res. 2011, 71, 5915–5922. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.; Mero, P.; Booth, J.W. Dynamics of macrophage trogocytosis of rituximab-coated b cells. PLoS ONE 2011, 6, e14498. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.; Friedlander, E.; Tanner, M.; Kapanen, A.I.; Carraway, K.L.; Isola, J.; Jovin, T.M. Decreased accessibility and lack of activation of erbb2 in jimt-1, a herceptin-resistant, muc4-expressing breast cancer cell line. Cancer Res. 2005, 65, 473–482. [Google Scholar] [PubMed]
- Prantner, A.M.; Turini, M.; Kerfelec, B.; Joshi, S.; Baty, D.; Chames, P.; Scholler, N. Anti-mesothelin nanobodies for both conventional and nanoparticle-based biomedical applications. J. Biomed. Nanotechnol. 2015, 11, 1201–1212. [Google Scholar] [CrossRef] [PubMed]
- Prantner, A.M.; Yin, C.; Kamat, K.; Sharma, K.; Lowenthal, A.C.; Madrid, P.B.; Scholler, N. Molecular imaging of mesothelin-expressing ovarian cancer with a human and mouse cross-reactive nanobody. Mol. Pharm. 2018, 15, 1403–1411. [Google Scholar] [CrossRef] [PubMed]
- Terwisscha van Scheltinga, A.G.; Ogasawara, A.; Pacheco, G.; Vanderbilt, A.N.; Tinianow, J.N.; Gupta, N.; Li, D.; Firestein, R.; Marik, J.; Scales, S.J.; et al. Preclinical efficacy of an antibody-drug conjugate targeting mesothelin correlates with quantitative 89zr-immunopet. Mol. Cancer Ther. 2017, 16, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Sirois, A.R.; Deny, D.A.; Baierl, S.R.; George, K.S.; Moore, S.J. Fn3 proteins engineered to recognize tumor biomarker mesothelin internalize upon binding. PLoS ONE 2018, 13, e0197029. [Google Scholar] [CrossRef] [PubMed]
- Hackel, B.J.; Kimura, R.H.; Gambhir, S.S. Use of (64)cu-labeled fibronectin domain with egfr-overexpressing tumor xenograft: Molecular imaging. Radiology 2012, 263, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Abou-Elkacem, L.; Wilson, K.E.; Johnson, S.M.; Chowdhury, S.M.; Bachawal, S.; Hackel, B.J.; Tian, L.; Willmann, J.K. Ultrasound molecular imaging of the breast cancer neovasculature using engineered fibronectin scaffold ligands: A novel class of targeted contrast ultrasound agent. Theranostics 2016, 6, 1740–1752. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Park, S.; Kim, D.Y.; Pyo, A.; Kimura, R.H.; Sathirachinda, A.; Choy, H.E.; Min, J.J.; Gambhir, S.S.; Hong, Y. Isolation and characterization of a monobody with a fibronectin domain iii scaffold that specifically binds epha2. PLoS ONE 2015, 10, e0132976. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Levi, J.; Cheng, Z. Protein scaffold-based molecular probes for cancer molecular imaging. Amino Acids 2011, 41, 1037–1047. [Google Scholar] [CrossRef] [PubMed]
- Alewine, C.; Hassan, R.; Pastan, I. Advances in anticancer immunotoxin therapy. Oncologist 2015, 20, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Kreitman, R.J.; Hassan, R.; Fitzgerald, D.J.; Pastan, I. Phase i trial of continuous infusion anti-mesothelin recombinant immunotoxin ss1p. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2009, 15, 5274–5279. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Bullock, S.; Premkumar, A.; Kreitman, R.J.; Kindler, H.; Willingham, M.C.; Pastan, I. Phase i study of ss1p, a recombinant anti-mesothelin immunotoxin given as a bolus i.V. Infusion to patients with mesothelin-expressing mesothelioma, ovarian, and pancreatic cancers. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2007, 13, 5144–5149. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Schweizer, C.; Lu, K.F.; Schuler, B.; Remaley, A.T.; Weil, S.C.; Pastan, I. Inhibition of mesothelin-ca-125 interaction in patients with mesothelioma by the anti-mesothelin monoclonal antibody morab-009: Implications for cancer therapy. Lung Cancer 2010, 68, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Cohen, S.J.; Phillips, M.; Pastan, I.; Sharon, E.; Kelly, R.J.; Schweizer, C.; Weil, S.; Laheru, D. Phase i clinical trial of the chimeric anti-mesothelin monoclonal antibody morab-009 in patients with mesothelin-expressing cancers. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2010, 16, 6132–6138. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Brockstedt, D.G.; Nir-Paz, R.; Hampl, J.; Mathur, S.; Nemunaitis, J.; Sterman, D.H.; Hassan, R.; Lutz, E.; Moyer, B.; et al. A live-attenuated listeria vaccine (anz-100) and a live-attenuated listeria vaccine expressing mesothelin (crs-207) for advanced cancers: Phase i studies of safety and immune induction. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Golfier, S.; Kopitz, C.; Kahnert, A.; Heisler, I.; Schatz, C.A.; Stelte-Ludwig, B.; Mayer-Bartschmid, A.; Unterschemmann, K.; Bruder, S.; Linden, L.; et al. Anetumab ravtansine: A novel mesothelin-targeting antibody-drug conjugate cures tumors with heterogeneous target expression favored by bystander effect. Mol. Cancer Ther. 2014, 13, 1537–1548. [Google Scholar] [CrossRef] [PubMed]
- Beatty, G.L.; Haas, A.R.; Maus, M.V.; Torigian, D.A.; Soulen, M.C.; Plesa, G.; Chew, A.; Zhao, Y.; Levine, B.L.; Albelda, S.M.; et al. Mesothelin-specific chimeric antigen receptor mrna-engineered t cells induce anti-tumor activity in solid malignancies. Cancer Immunol. Res. 2014, 2, 112–120. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Model for peritoneal metastasis of ovarian tumors. Ovarian cancer metastasis is unique as tumor cells shed from the primary tumor and spread throughout the peritoneal cavity. MSLN:CA125 interaction mediates heterotypic and homotypic cellular adhesion.
Figure 1.
Model for peritoneal metastasis of ovarian tumors. Ovarian cancer metastasis is unique as tumor cells shed from the primary tumor and spread throughout the peritoneal cavity. MSLN:CA125 interaction mediates heterotypic and homotypic cellular adhesion.

Figure 2.
Structure of mesothelin (MSLN). The 70 kDa MSLN precursor protein is proteolytically cleaved to release the 30 kDa N-terminal megakaryocyte potentiating factor (MPF) and is displayed as mature MSLN on the cell surface.
Figure 2.
Structure of mesothelin (MSLN). The 70 kDa MSLN precursor protein is proteolytically cleaved to release the 30 kDa N-terminal megakaryocyte potentiating factor (MPF) and is displayed as mature MSLN on the cell surface.

© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hilliard, T.S. The Impact of Mesothelin in the Ovarian Cancer Tumor Microenvironment. Cancers 2018, 10, 277. https://doi.org/10.3390/cancers10090277
AMA Style
Hilliard TS. The Impact of Mesothelin in the Ovarian Cancer Tumor Microenvironment. Cancers. 2018; 10(9):277. https://doi.org/10.3390/cancers10090277
Chicago/Turabian StyleHilliard, Tyvette S. 2018. "The Impact of Mesothelin in the Ovarian Cancer Tumor Microenvironment" Cancers 10, no. 9: 277. https://doi.org/10.3390/cancers10090277
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.