The Healing Effect of Human Milk Fat Globule-EGF Factor 8 Protein (MFG-E8) in A Rat Model of Parkinson’s Disease


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animal Care
2.3. Preparation of hADSCs
2.4. Cell Differentiation
2.5. Preparation of hADSC-CMs for the Analysis of the Protein Expression by LC–MS/MS
2.6. Preparation of the Mouse Model of PD
2.7. Detection of the TH Expression of Dopaminergic Neurons by Immunohistochemistry
2.8. Protein Identification by a Nano LC–MS/MS Analysis
2.9. Data Analyses
2.9.1. Database Searching
2.9.2. Criteria for Protein Identification
2.10. RefEx Analysis
2.11. The BioGSP Analysis
3. Results
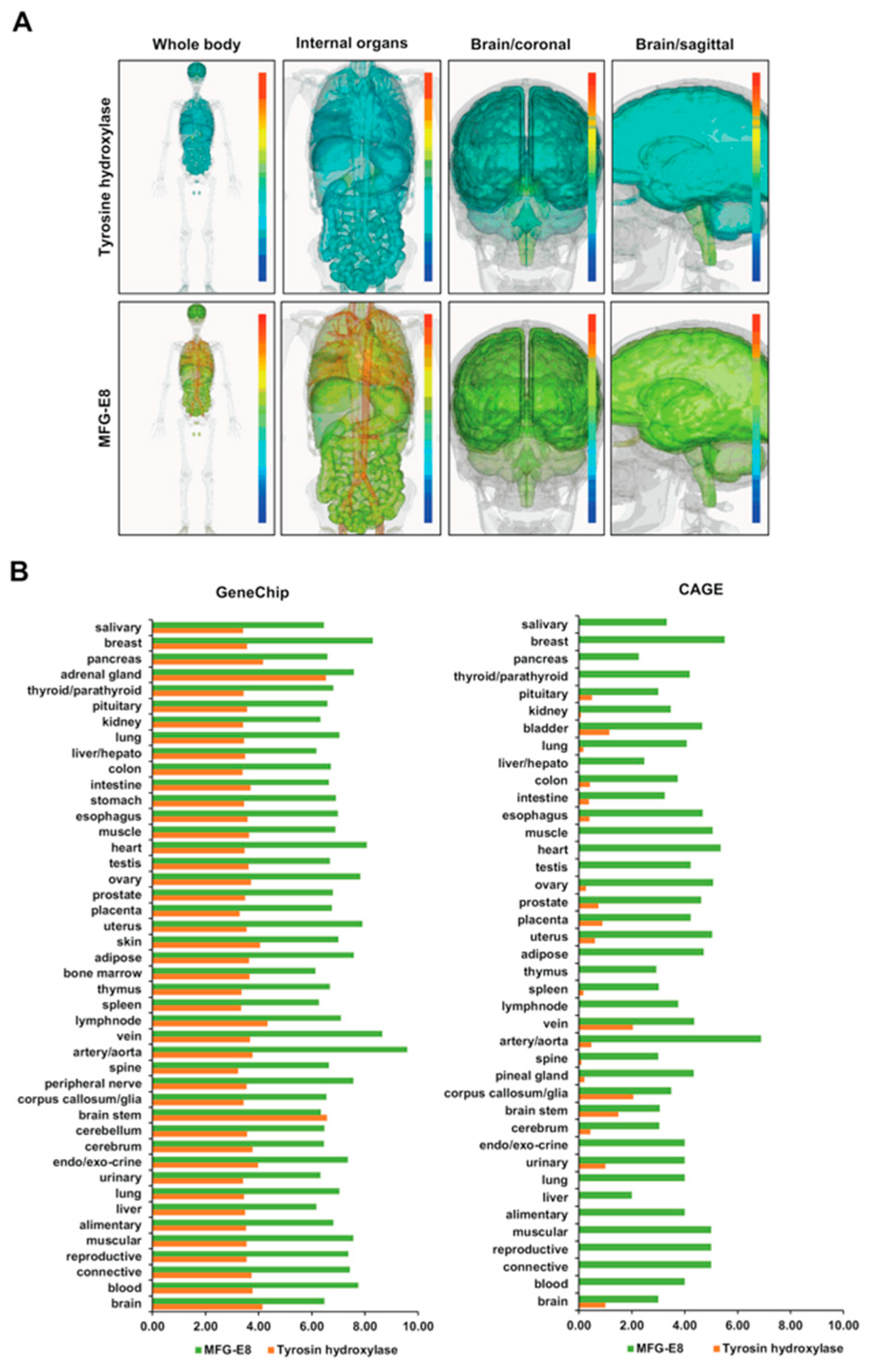
3.1. Distribution of Systemic DNA and the mRNA Expression of TH
3.2. Distribution of Systemic DNA and the mRNA Expression of MFG-E8
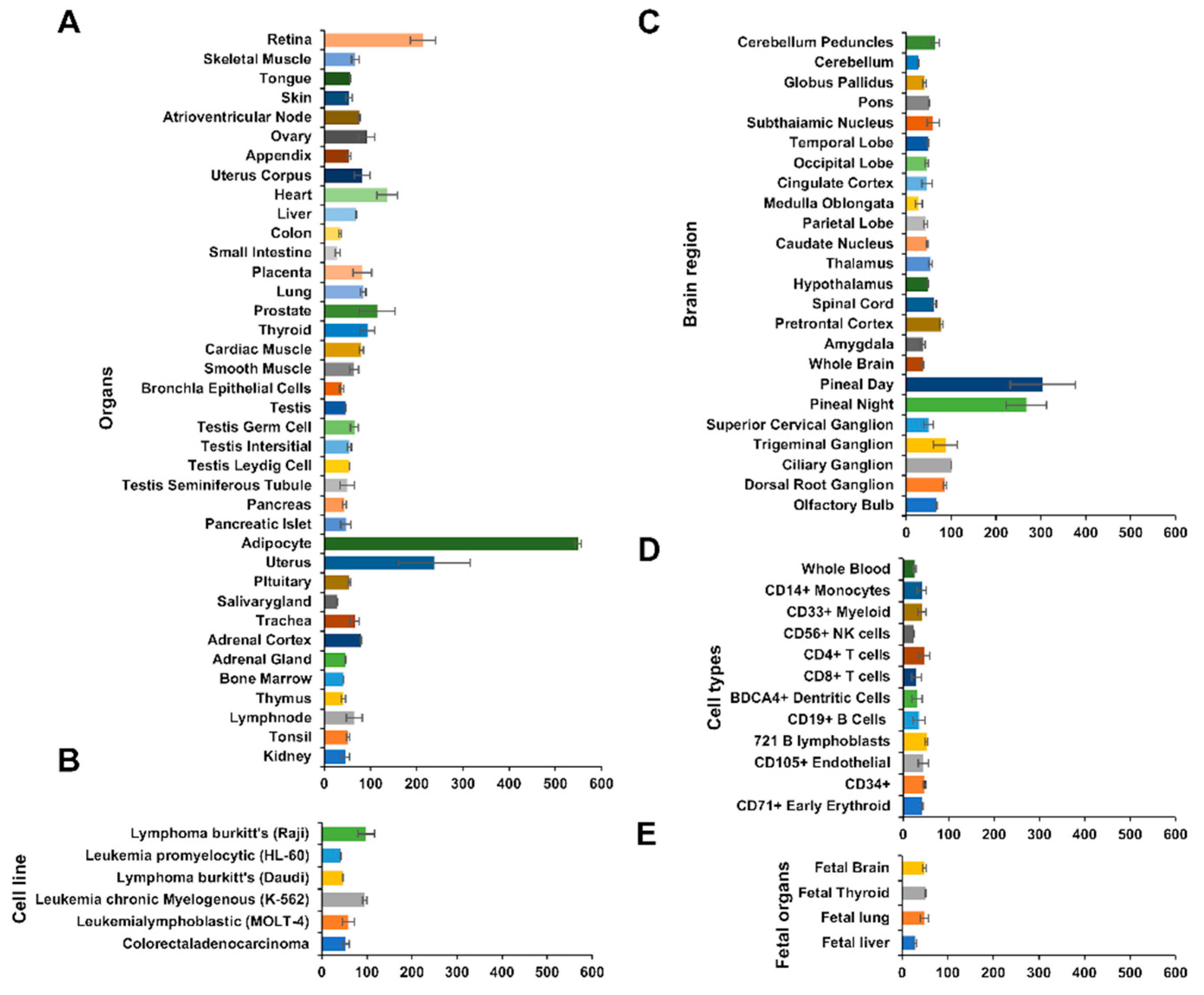
3.3. Comparison of the Human MFG-E8 mRNA Expression Levels in the Organs, Brain, and Different Cell Types
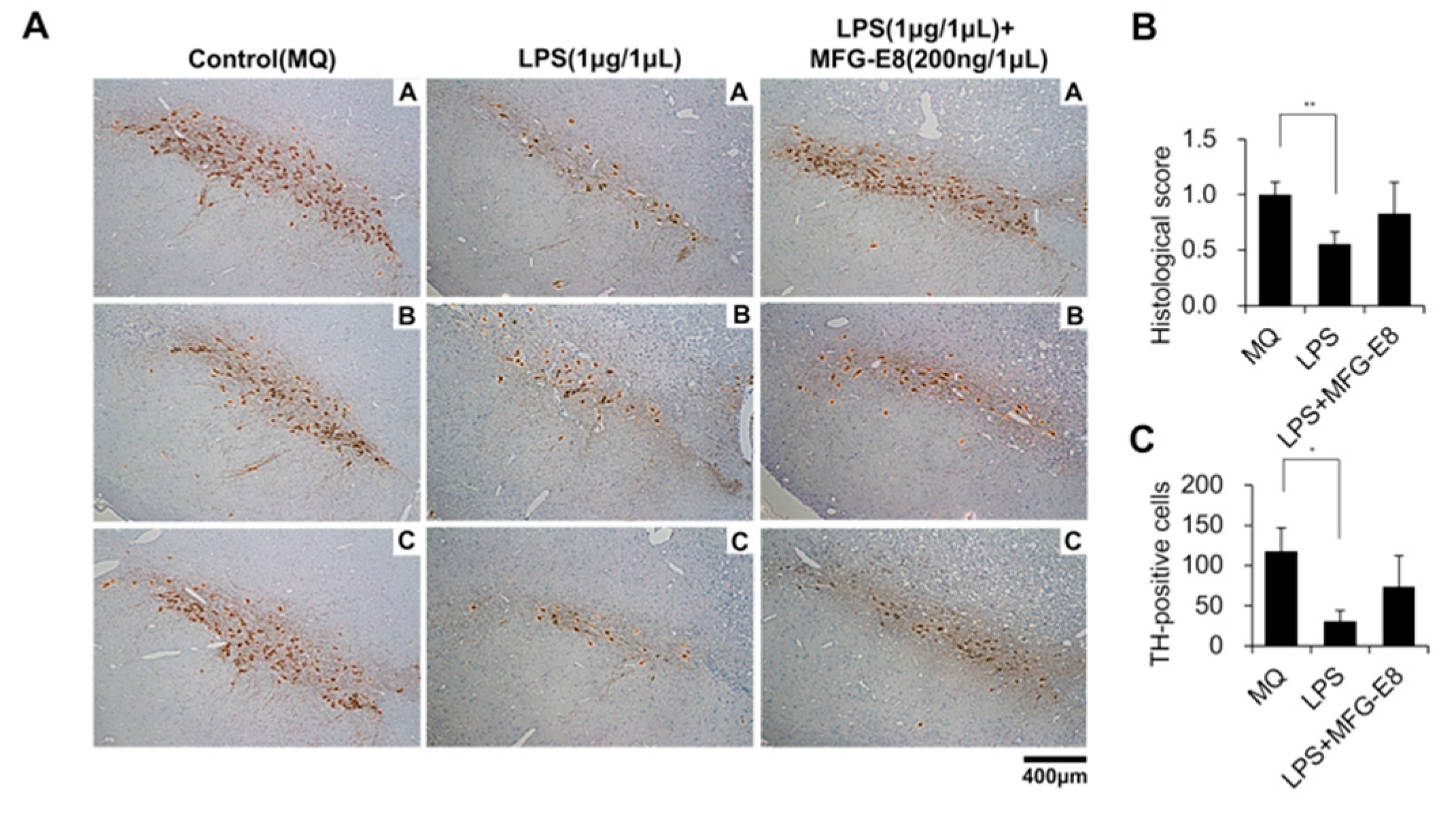
3.4. Human MFG-E8 Prevents the LPS-Induced Reduction of Dopamine Neurons in the Rat Nigra
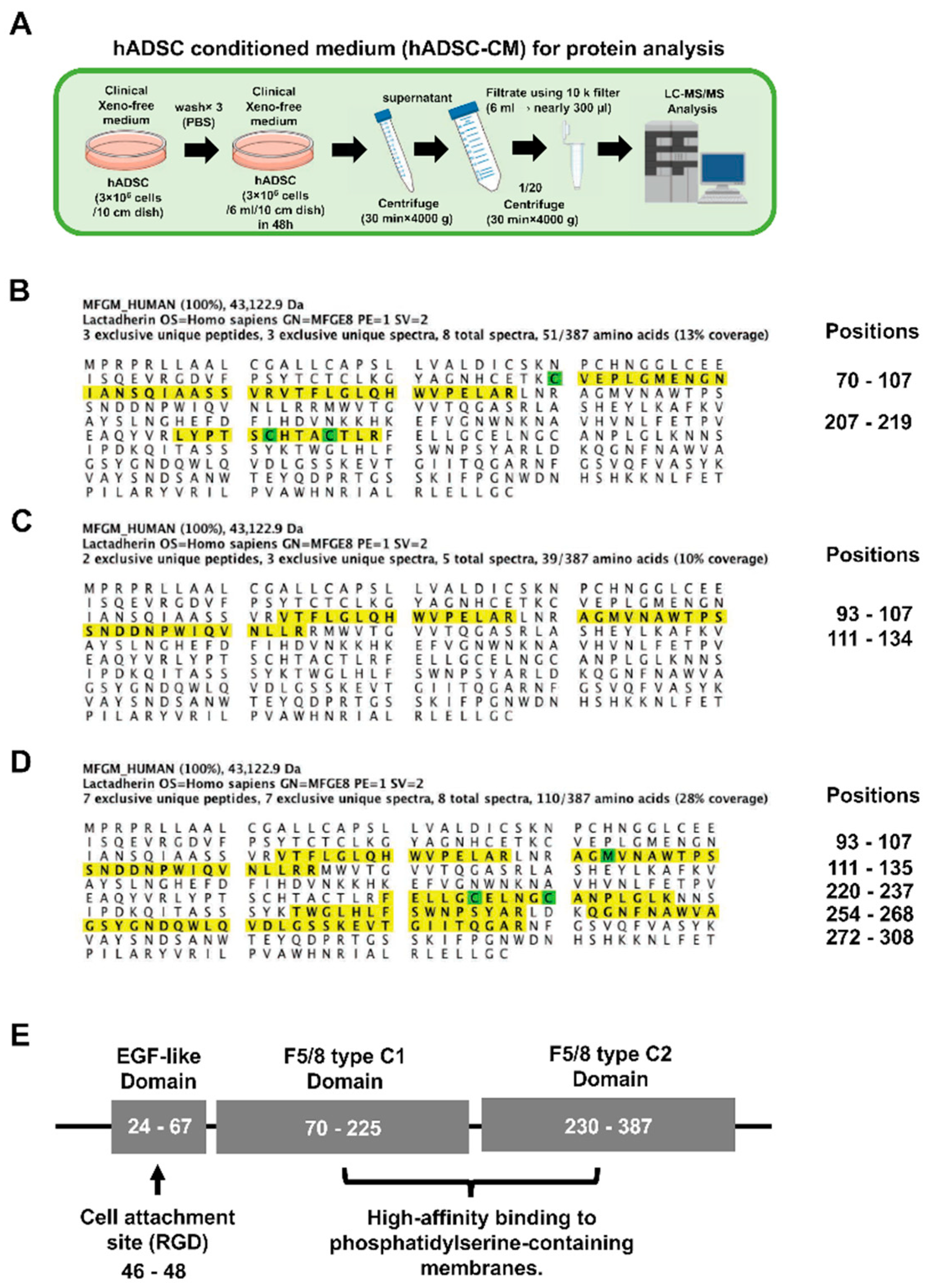
3.5. Preparations of hADSC-CMs for the Analysis of the Protein Expression by LC–MS/MS
3.6. The Peptide Sequence of MFG-E8 Secreted or Expressed by hADSCs
3.7. The Biological Processes, Cellular Components, and Molecular Function of Proteins Identified from hADSC-CM, Cell Lysate of hADSC Cultured in CDM or DMEM-10% FBS
3.7.1. Biological Processes
3.7.2. Cellular Components
3.7.3. Molecular Function
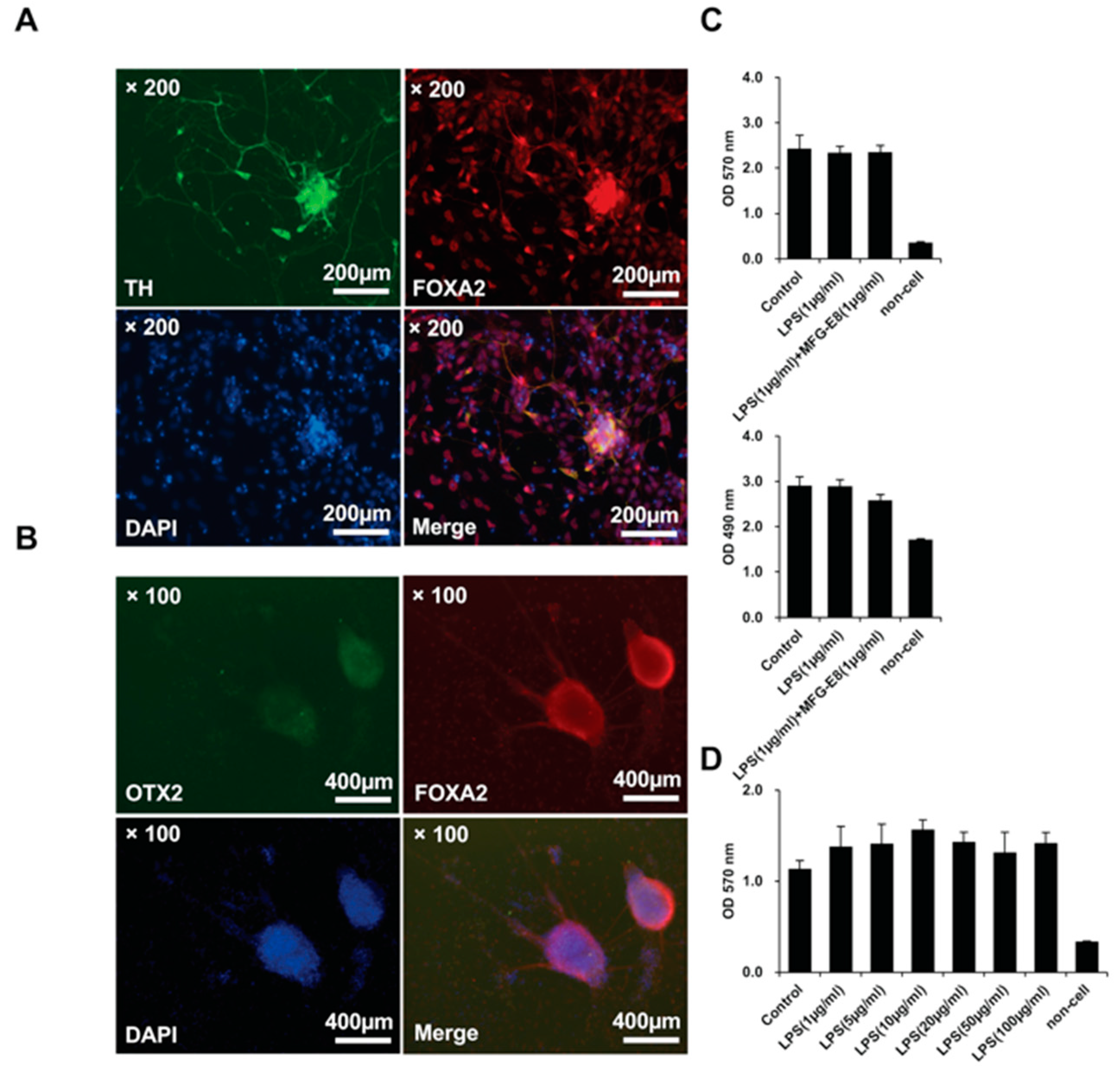
3.8. The Effects of LPS and Human MFG-E8 on iPS Cell-Derived Dopamine Neurons In Vitro
3.9. The Effects of LPS and Human MFG-E8 on iPS Cell-Derived Neurons In Vitro
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rao, S.S.; Hofmann, L.A.; Shakil, A. Parkinson’s disease: Diagnosis and treatment. Am. Fam. Phys. 2006, 74, 2046–2054. [Google Scholar]
- Zhang, W.; Wang, T.; Pei, Z.; Miller, D.S.; Wu, X.; Block, M.L.; Wilson, B.; Zhang, W.; Zhou, Y.; Hong, J.S.; et al. Aggregated alpha-synuclein activates microglia: A process leading to disease progression in parkinson’s disease. FASEB J. 2005, 19, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.L.; Dragicevic, N.; Seifert, K.; Choi, D.Y.; Liu, M.; Kim, H.C.; Cass, W.A.; Sullivan, P.G.; Bing, G. Inflammation induces mitochondrial dysfunction and dopaminergic neurodegeneration in the nigrostriatal system. J. Neurochem. 2007, 100, 1375–1386. [Google Scholar] [CrossRef] [PubMed]
- Terada, T.; Yokokura, M.; Yoshikawa, E.; Futatsubashi, M.; Kono, S.; Konishi, T.; Miyajima, H.; Hashizume, T.; Ouchi, Y. Extrastriatal spreading of microglial activation in parkinson’s disease: A positron emission tomography study. Ann. Nucl. Med. 2016, 30, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, Y.; Yoshikawa, E.; Sekine, Y.; Futatsubashi, M.; Kanno, T.; Ogusu, T.; Torizuka, T. Microglial activation and dopamine terminal loss in early parkinson’s disease. Ann. Neurol. 2005, 57, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, C.C.; Tarelli, R. Parkinson’s disease and systemic inflammation. Parkinsons Dis. 2011, 2011, 436813. [Google Scholar] [CrossRef] [PubMed]
- Leow-Dyke, S.; Allen, C.; Denes, A.; Nilsson, O.; Maysami, S.; Bowie, A.G.; Rothwell, N.J.; Pinteaux, E. Neuronal toll-like receptor 4 signaling induces brain endothelial activation and neutrophil transmigration in vitro. J. Neuroinflamm. 2012, 9, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.; Lee, H.J.; Masliah, E.; Lee, S.J. Non-cell-autonomous neurotoxicity of alpha-synuclein through microglial toll-like receptor 2. Exp. Neurobiol. 2016, 25, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Okita, K.; Nakagawa, M.; Yamanaka, S. Induction of pluripotent stem cells from fibroblast cultures. Nat. Protoc. 2007, 2, 3081–3089. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Morizane, A.; Doi, D.; Magotani, H.; Onoe, H.; Hayashi, T.; Mizuma, H.; Takara, S.; Takahashi, R.; Inoue, H.; et al. Human ips cell-derived dopaminergic neurons function in a primate parkinson’s disease model. Nature 2017, 548, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Bing, G. Lipopolysaccharide animal models for parkinson’s disease. Parkinsons Dis. 2011, 2011, 327089. [Google Scholar] [CrossRef] [PubMed]
- Schintu, N.; Zhang, X.; Svenningsson, P. Studies of depression-related states in animal models of parkinsonism. J. Parkinsons Dis. 2012, 2, 87–106. [Google Scholar] [PubMed]
- Hoban, D.B.; Connaughton, E.; Connaughton, C.; Hogan, G.; Thornton, C.; Mulcahy, P.; Moloney, T.C.; Dowd, E. Further characterisation of the lps model of parkinson’s disease: A comparison of intra-nigral and intra-striatal lipopolysaccharide administration on motor function, microgliosis and nigrostriatal neurodegeneration in the rat. Brain Behav. Immun. 2013, 27, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.; Herrera, A.J.; Venero, J.L.; Santiago, M.; de Pablos, R.M.; Villaran, R.F.; Espinosa-Oliva, A.M.; Arguelles, S.; Sarmiento, M.; Delgado-Cortes, M.J.; et al. Inflammatory animal model for parkinson’s disease: The intranigral injection of lps induced the inflammatory process along with the selective degeneration of nigrostriatal dopaminergic neurons. ISRN Neurol. 2011, 2011, 476158. [Google Scholar] [CrossRef] [PubMed]
- Dutta, G.; Zhang, P.; Liu, B. The lipopolysaccharide parkinson’s disease animal model: Mechanistic studies and drug discovery. Fundam. Clin. Pharmacol. 2008, 22, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.K.; Jou, I.; Joe, E.H. Systemic lps administration induces brain inflammation but not dopaminergic neuronal death in the substantia nigra. Exp. Mol. Med. 2010, 42, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, Y.; Miyagi-Shiohira, C.; Kobayashi, N.; Saitoh, I.; Watanabe, M.; Noguchi, H. A proteome analysis of pig pancreatic islets and exocrine tissue by liquid chromatography with tandem mass spectrometry. Islets 2017, 9, 159–176. [Google Scholar] [CrossRef] [PubMed]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef] [PubMed]
- Ono, H.; Ogasawara, O.; Okubo, K.; Bono, H. Refex, a reference gene expression dataset as a web tool for the functional analysis of genes. Sci. Data 2017, 4, 170105. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Orozco, C.; Boyer, J.; Leglise, M.; Goodale, J.; Batalov, S.; Hodge, C.L.; Haase, J.; Janes, J.; Huss, J.W.; et al. Biogps: An extensible and customizable portal for querying and organizing gene annotation resources. Genome Biol. 2009, 10, R130. [Google Scholar] [CrossRef] [PubMed]
- Su, A.I.; Wiltshire, T.; Batalov, S.; Lapp, H.; Ching, K.A.; Block, D.; Zhang, J.; Soden, R.; Hayakawa, M.; Kreiman, G.; et al. A gene atlas of the mouse and human protein-encoding transcriptomes. Proc. Natl. Acad. Sci. USA 2004, 101, 6062–6067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, P.; Randall, S.M.; Muddiman, D.C.; Rao, B.M. Targeted proteomics of the secretory pathway reveals the secretome of mouse embryonic fibroblasts and human embryonic stem cells. Mol. Cell. Proteom. 2012, 11, 1829–1839. [Google Scholar] [CrossRef] [PubMed]
- Castellanos, E.R.; Ciferri, C.; Phung, W.; Sandoval, W.; Matsumoto, M.L. Expression, purification, and characterization of recombinant human and murine milk fat globule-epidermal growth factor-factor 8. Protein Expr. Purif. 2016, 124, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.R.; Couto, J.R.; Scallan, C.D.; Ceriani, R.L.; Peterson, J.A. Lactadherin (formerly ba46), a membrane-associated glycoprotein expressed in human milk and breast carcinomas, promotes arg-gly-asp (rgd)-dependent cell adhesion. DNA Cell Biol. 1997, 16, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Totsuka, R.; Miyatani, S.; Kurata, S.; Sato, S.; Katoh, I.; Kobayashi, S.; Ikawa, Y. Production of the long and short forms of mfg-e8 by epidermal keratinocytes. Cell Tissue Res. 2005, 321, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, T.; Kurata, S.; Tomimori, Y.; Fukunishi, N.; Sato, S.; Osada, M.; Tsukinoki, K.; Jin, H.F.; Yamashita, A.; Ito, M.; et al. P63(tp63) elicits strong trans-activation of the mfg-e8/lactadherin/ba46 gene through interactions between the ta and delta n isoforms. Oncogene 2008, 27, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Oshima, K.; Aoki, N.; Kato, T.; Kitajima, K.; Matsuda, T. Secretion of a peripheral membrane protein, mfg-e8, as a complex with membrane vesicles–A possible role in membrane secretion. Eur. J. Biochem. 2002, 269, 1209–1218. [Google Scholar] [CrossRef] [PubMed]
- Hanayama, R.; Tanaka, M.; Miwa, K.; Shinohara, A.; Iwamatsu, A.; Nagata, S. Identification of a factor that links apoptotic cells to phagocytes. Nature 2002, 417, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Hanayama, R.; Nagata, S. Impaired involution of mammary glands in the absence of milk fat globule egf factor 8. Proc. Natl. Acad. Sci. USA 2005, 102, 16886–16891. [Google Scholar] [CrossRef] [PubMed]
- Hanayama, R.; Tanaka, M.; Miyasaka, K.; Aozasa, K.; Koike, M.; Uchiyama, Y.; Nagata, S. Autoimmune disease and impaired uptake of apoptotic cells in mfg-e8-deficient mice. Science 2004, 304, 1147–1150. [Google Scholar] [CrossRef] [PubMed]
- Fricker, M.; Neher, J.J.; Zhao, J.W.; Thery, C.; Tolkovsky, A.M.; Brown, G.C. Mfg-e8 mediates primary phagocytosis of viable neurons during neuroinflammation. J. Neurosci. 2012, 32, 2657–2666. [Google Scholar] [CrossRef] [PubMed]
- Bu, H.F.; Zuo, X.L.; Wang, X.; Ensslin, M.A.; Koti, V.; Hsueh, W.; Raymond, A.S.; Shur, B.D.; Tan, X.D. Milk fat globule-egf factor 8/lactadherin plays a crucial role in maintenance and repair of murine intestinal epithelium. J. Clin. Investig. 2007, 117, 3673–3683. [Google Scholar] [CrossRef] [PubMed]
- Nandrot, E.F.; Anand, M.; Almeida, D.; Atabai, K.; Sheppard, D.; Finnemann, S.C. Essential role for mfg-e8 as ligand for alpha v beta 5 integrin in diurnal retinal phagocytosis. Proc. Natl. Acad. Sci. USA 2007, 104, 12005–12010. [Google Scholar] [CrossRef] [PubMed]
- Motegi, S.; Leitner, W.W.; Lu, M.; Tada, Y.; Sardy, M.; Wu, C.J.; Chavakis, T.; Udey, M.C. Pericyte-derived mfg-e8 regulates pathologic angiogenesis. Arterioscl. Throm. Vas. 2011, 31, 2024–2034. [Google Scholar] [CrossRef] [PubMed]
- Silvestre, J.S.; Thery, C.; Hamard, G.; Boddaert, J.; Aguilar, B.; Delcayre, A.; Houbron, C.; Tamarat, R.; Blanc-Brude, O.; Heeneman, S.; et al. Lactadherin promotes vegf-dependent neovascularization. Nat. Med. 2005, 11, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Neutzner, M.; Lopez, T.; Feng, X.; Bergmann-Leitner, E.S.; Leitner, W.W.; Udey, M.C. Mfg-e8/lactadherin promotes tumor growth in an angiogenesis-dependent transgenic mouse model of multistage carcinogenesis. Cancer Res. 2007, 67, 6777–6785. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.Y.; Fu, Z.M.; Wu, J.; Zhang, J.; Jiang, L.Q.; Khazan, B.; Telljohann, R.; Zhao, M.M.; Krug, A.W.; Pikilidou, M.; et al. Mfg-e8 activates proliferation of vascular smooth muscle cells via integrin signaling. Aging Cell 2012, 11, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Joh, T.H. Microglia, major player in the brain inflammation: Their roles in the pathogenesis of parkinson’s disease. Exp. Mol. Med. 2006, 38, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Ishii, A.; Ohtaki, H.; Shioda, S.; Yoshida, T.; Numazawa, S. Activation of microglia induces symptoms of parkinson’s disease in wild-type, but not in il-1 knockout mice. J. Neuroinflamm. 2013, 10, 143. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.M.; Jiang, J.; Wilson, B.; Zhang, W.; Hong, J.S.; Liu, B. Microglial activation-mediated delayed and progressive degeneration of rat nigral dopaminergic neurons: Relevance to parkinson’s disease. J. Neurochem. 2002, 81, 1285–1297. [Google Scholar] [CrossRef] [PubMed]
- Neniskyte, U.; Brown, G.C. Lactadherin/mfg-e8 is essential for microglia-mediated neuronal loss and phagoptosis induced by amyloid beta. J. Neurochem. 2013, 126, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Fuller, A.D.; Van Eldik, L.J. Mfg-e8 regulates microglial phagocytosis of apoptotic neurons. J. Neuroimmune Pharm. 2008, 3, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, X.S.; Guo, C.Y.; Nie, P.; Liu, Y.; Ma, J. Essential role of mfg-e8 for phagocytic properties of microglial cells. PLoS ONE 2013, 8, e55754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raoul, W.; Poupel, L.; Tregouet, D.A.; Lavalette, S.; Camelo, S.; Keller, N.; Krumeich, S.; Calippe, B.; Guillonneau, X.; Behar-Cohen, F.; et al. Mfge8 does not influence chorio-retinal homeostasis or choroidal neovascularization in vivo. PLoS ONE 2012, 7, e33244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, G.W.; Chen, X.D.; Dong, F.P.; Li, W.Z.; Ren, X.H.; Zhang, Y.Y.; Shi, Y.F. Concise review: Mesenchymal stem cells and translational medicine: Emerging issues. Stem Cell Transl. Med. 2012, 1, 51–58. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakashima, Y.; Miyagi-Shiohira, C.; Noguchi, H.; Omasa, T. The Healing Effect of Human Milk Fat Globule-EGF Factor 8 Protein (MFG-E8) in A Rat Model of Parkinson’s Disease. Brain Sci. 2018, 8, 167. https://doi.org/10.3390/brainsci8090167
Nakashima Y, Miyagi-Shiohira C, Noguchi H, Omasa T. The Healing Effect of Human Milk Fat Globule-EGF Factor 8 Protein (MFG-E8) in A Rat Model of Parkinson’s Disease. Brain Sciences. 2018; 8(9):167. https://doi.org/10.3390/brainsci8090167
Chicago/Turabian StyleNakashima, Yoshiki, Chika Miyagi-Shiohira, Hirofumi Noguchi, and Takeshi Omasa. 2018. "The Healing Effect of Human Milk Fat Globule-EGF Factor 8 Protein (MFG-E8) in A Rat Model of Parkinson’s Disease" Brain Sciences 8, no. 9: 167. https://doi.org/10.3390/brainsci8090167