
Clozapine Suppresses the Gene Expression and the Production of Cytokines and Up-Regulates Cyclooxygenase 2 mRNA in Human Astroglial Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
2.3. Cytokine Array
2.4. Real-Time Polymerase Chain Reaction (RT-PCR)
2.5. Cytokine Determination
2.6. Western Blot
2.7. Statistical Analysis
3. Results
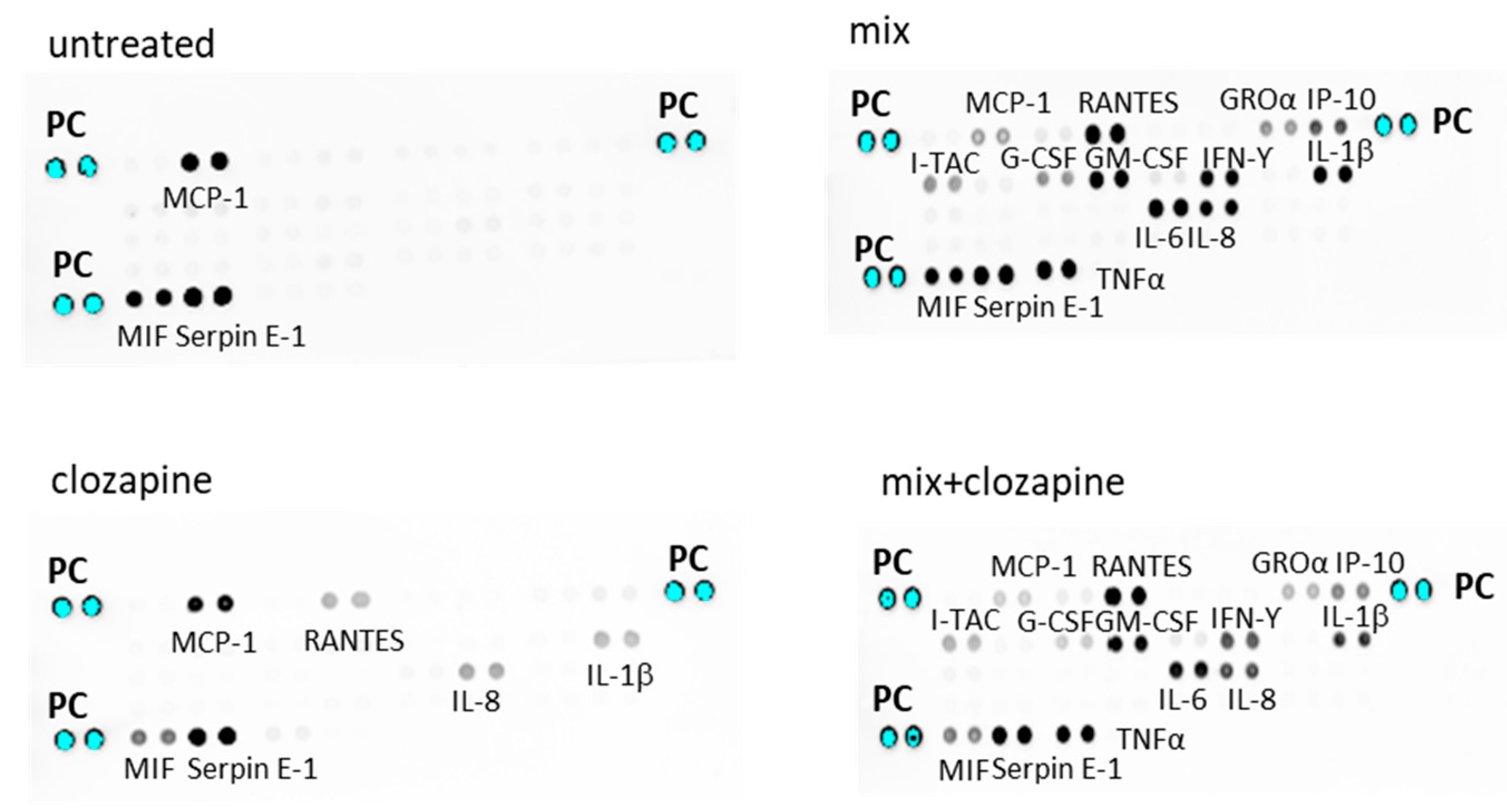
3.1. Effects of Clozapine on Inflammatory Mediators: Cytokine Array Analysis
3.1.1. Non-Inflammatory Condition
3.1.2. Inflammatory Condition
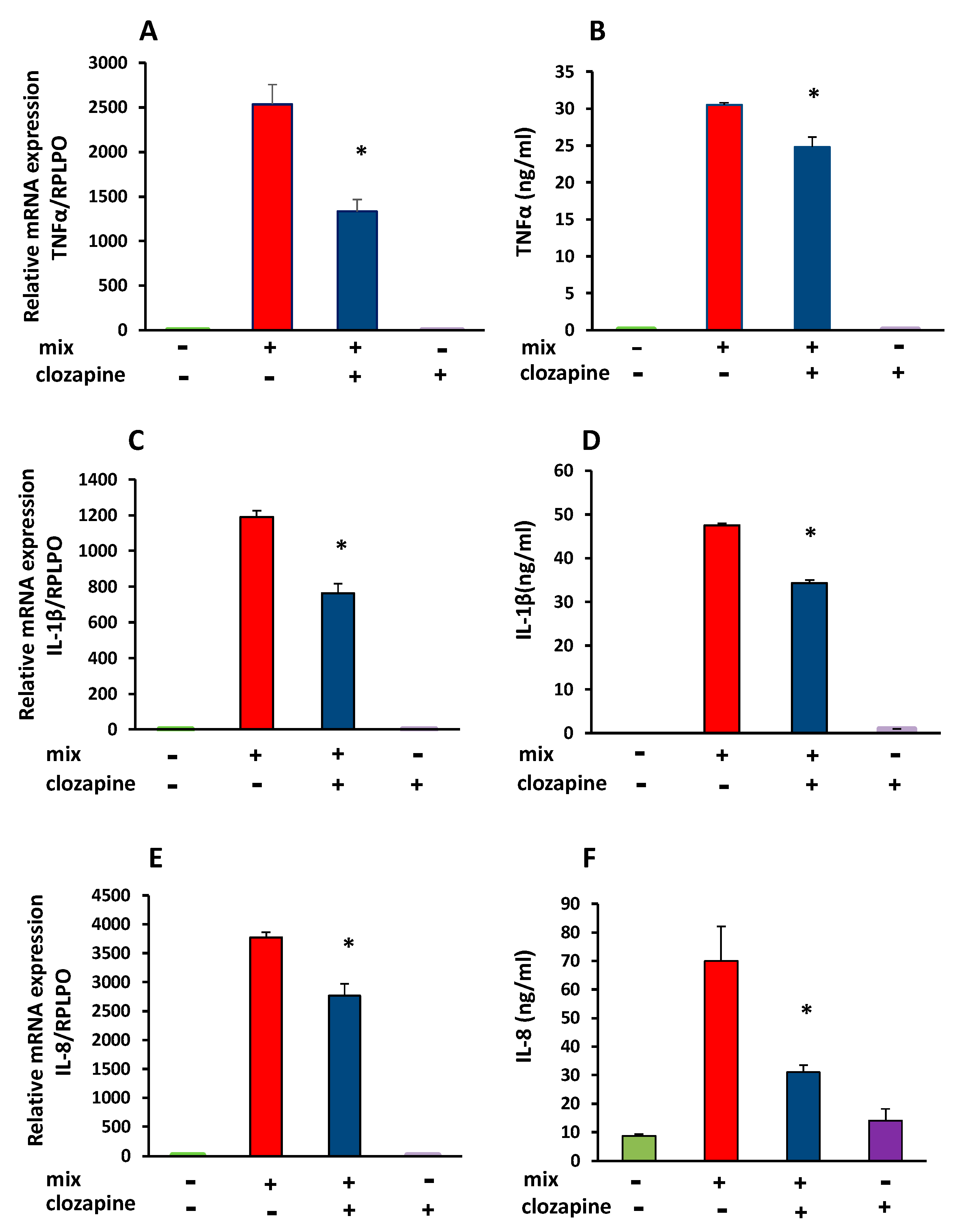
3.2. Inhibitory Effect of Clozapine on TNFα, IL-1β, and IL-8 Gene Expression and Protein Production
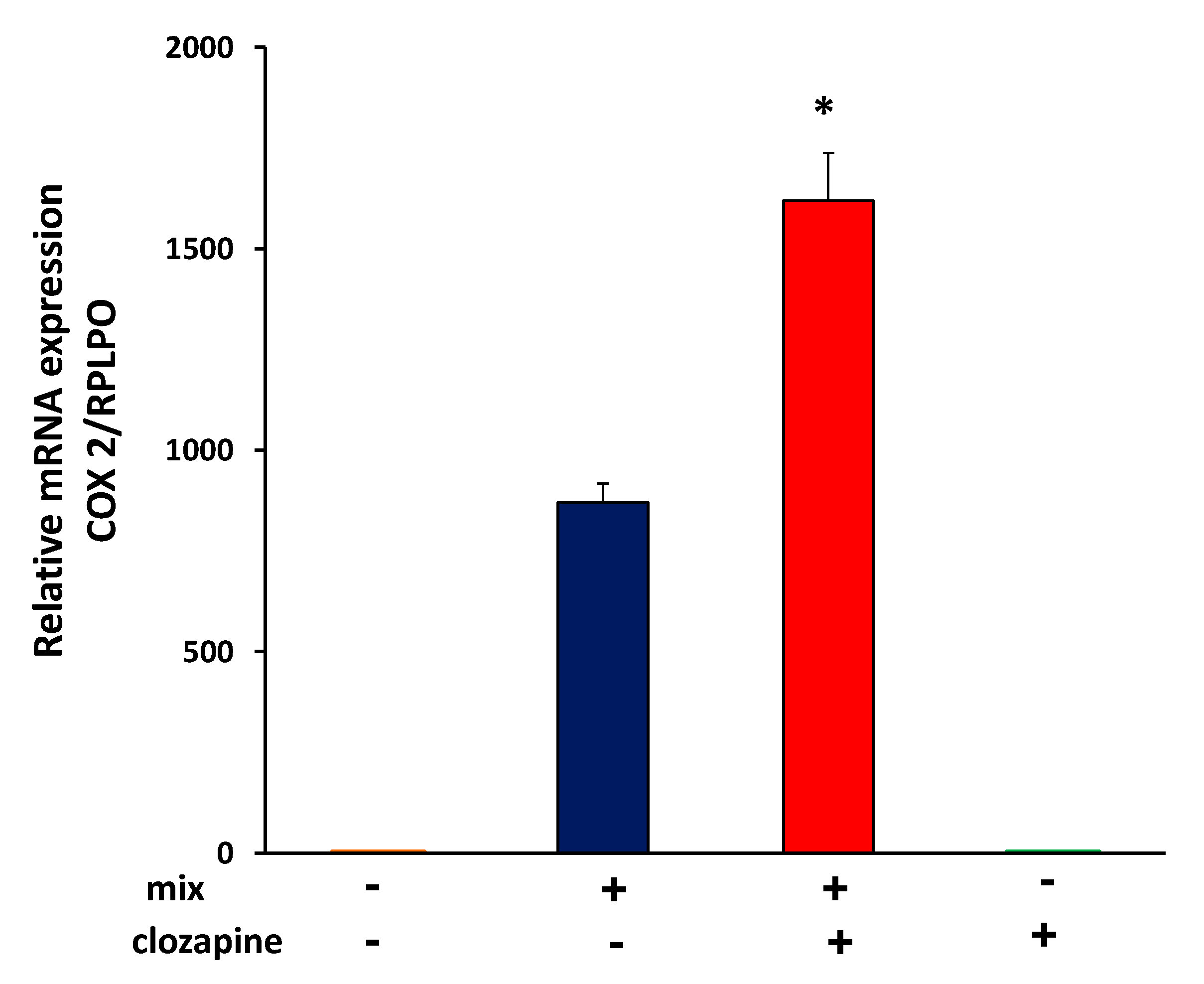
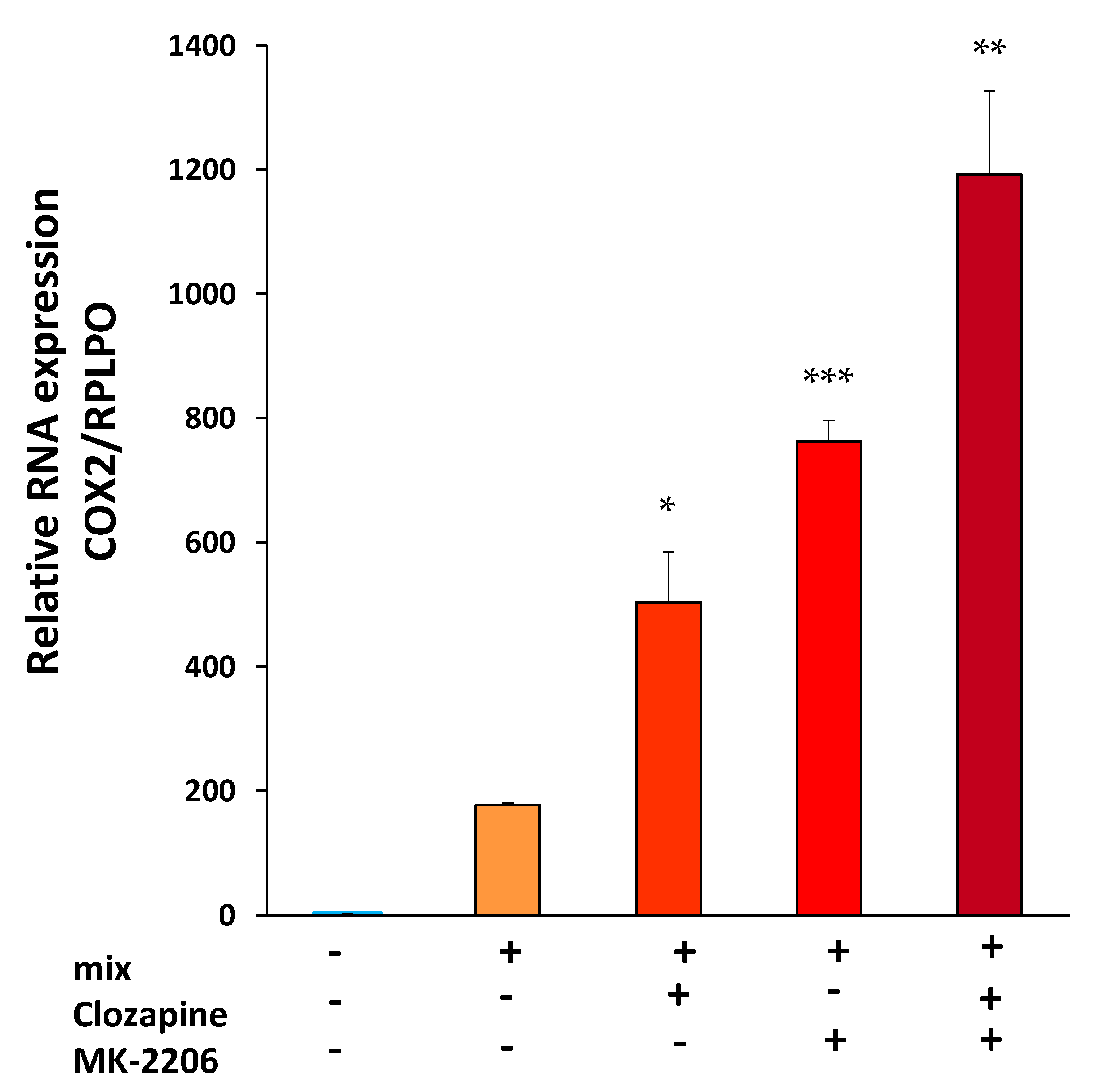
3.3. Clozapine Up-Regulates Cyclooxygenase 2 mRNA Expression
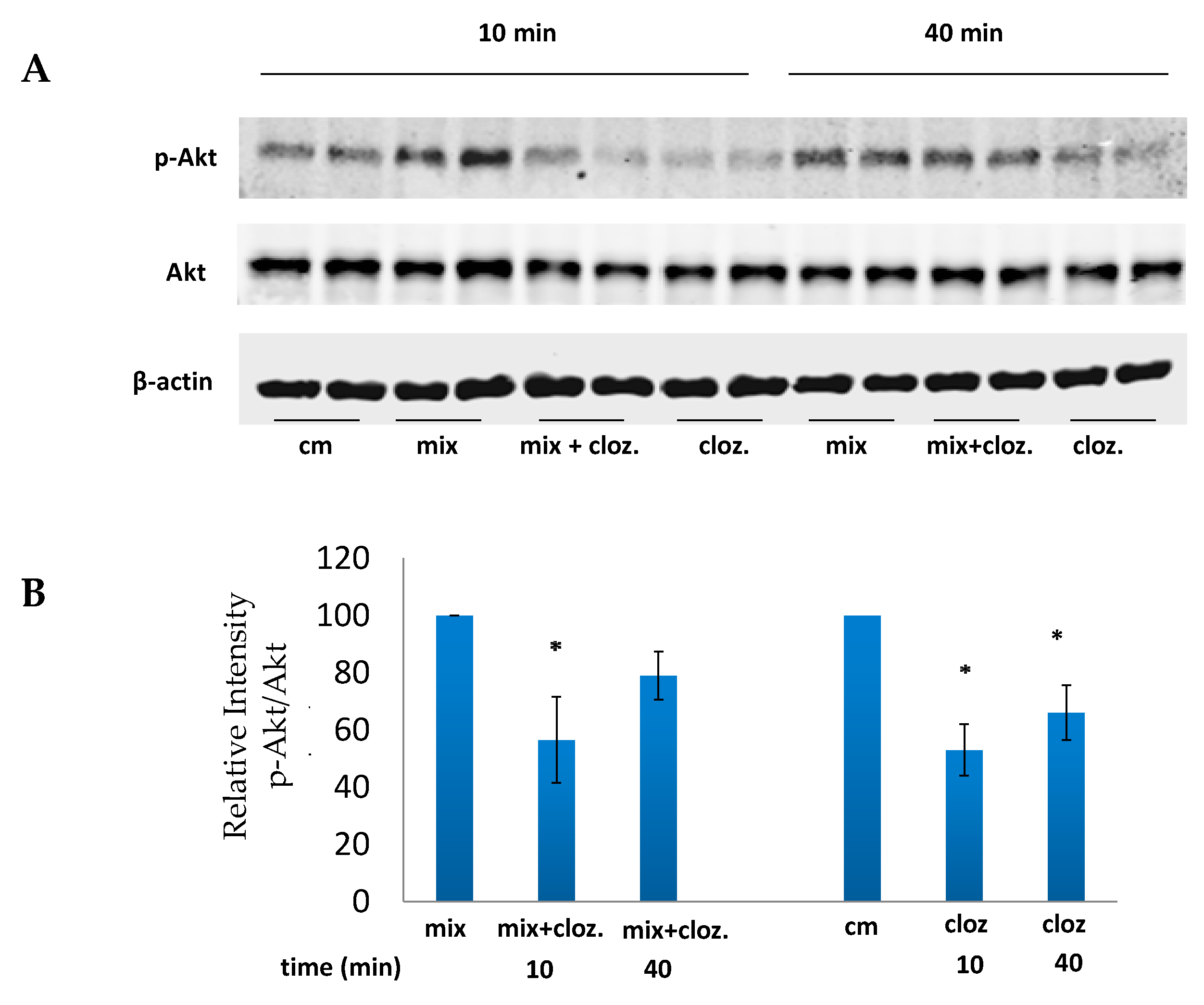
3.4. Clozapine Inhibits Akt Phosphorylation
4. Discussion
Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Correll, C.U.; Agid, O.; Crespo-Facorro, B.; de Bartolomeis, A.; Fagiolini, A.; Seppälä, N.; Howes, O.D. A Guideline and Checklist for Initiating and Managing Clozapine Treatment in Patients with Treatment-Resistant Schizophrenia. CNS Drugs 2022, 36, 659–679. [Google Scholar] [CrossRef]
- Panov, G.P. Early Markers in Resistant Schizophrenia: Effect of the First Antipsychotic Drug. Diagnostics 2022, 12, 803. [Google Scholar] [CrossRef]
- Fond, G.; Lancon, C.; Korchia, T.; Auquier, P.; Boyer, L. The Role of Inflammation in the Treatment of Schizophrenia. Front. Psychiatry 2020, 11, 160. [Google Scholar] [CrossRef] [Green Version]
- Khandaker, G.M.; Cousins, L.; Deakin, J.; Lennox, B.R.; Yolken, R.; Jones, P.B. Inflammation and immunity in schizophrenia: Implications for pathophysiology and treatment. Lancet Psychiatry 2015, 2, 258–270. [Google Scholar] [CrossRef] [Green Version]
- Reale, M.; Patruno, A.; De Lutiis, M.A.; Pesce, M.; Felaco, M.; Di Giannantonio, M.; Di Nicola, M.; Grilli, A. Dysregulation of chemo-cytokine production in schizophrenic patients versus healthy controls. BMC Neurosci. 2011, 12, 13. [Google Scholar] [CrossRef] [Green Version]
- Muller, N. Inflammation in Schizophrenia: Pathogenetic Aspects and Therapeutic Considerations. Schizophr. Bull. 2018, 44, 973–982. [Google Scholar] [CrossRef] [Green Version]
- Na, K.-S.; Jung, H.-Y.; Kim, Y.-K. The role of pro-inflammatory cytokines in the neuroinflammation and neurogenesis of schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 48, 277–286. [Google Scholar] [CrossRef]
- Goldsmith, D.R.; Massa, N.; Pearce, B.D.; Wommack, E.C.; Alrohaibani, A.; Goel, N.; Cuthbert, B.; Fargotstein, M.; Felger, J.C.; Haroon, E.; et al. Inflammatory markers are associated with psychomotor slowing in patients with schizophrenia compared to healthy controls. NPJ Schizophr. 2020, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Khandaker, G.M.; Dantzer, R.; Jones, P.B. Immunopsychiatry: Important facts. Psychol. Med. 2017, 47, 2229–2237. [Google Scholar] [CrossRef] [Green Version]
- Reale, M.; Costantini, E.; Greig, N.H. Cytokine Imbalance in Schizophrenia. From Research to Clinic: Potential Implications for Treatment. Front. Psychiatry 2021, 12, 536257. [Google Scholar] [CrossRef]
- Comer, A.L.; Carrier, M.; Tremblay, M.E.; Cruz-Martin, A. The Inflamed Brain in Schizophrenia: The Convergence of Genetic and Environmental Risk Factors That Lead to Uncontrolled Neuroinflammation. Front. Cell Neurosci. 2020, 14, 274. [Google Scholar] [CrossRef]
- Miller, B.J.; Goldsmith, D.R. Evaluating the Hypothesis That Schizophrenia Is an Inflammatory Disorder. Focus 2020, 18, 391–401. [Google Scholar] [CrossRef]
- Srinivas, L.; Vellichirammal, N.N.; Alex, A.M.; Nair, C.; Nair, I.V.; Banerjee, M. Pro-inflammatory cytokines and their epistatic interactions in genetic susceptibility to schizophrenia. J. Neuroinflamm. 2016, 13, 105. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Tóth, F.; Polyák, H.; Szabó, Á.; Mándi, Y.; Vécsei, L. Immune Influencers in Action: Metabolites and Enzymes of the Tryptophan-Kynurenine Metabolic Pathway. Biomedicines 2021, 9, 734. [Google Scholar] [CrossRef]
- Nyatega, C.O.; Qiang, L.; Adamu, M.J.; Younis, A.; Kawuwa, H.B. Altered Dynamic Functional Connectivity of Cuneus in Schizophrenia Patients: A Resting-State fMRI Study. Appl. Sci. 2021, 11, 11392. [Google Scholar] [CrossRef]
- Sapienza, J.; Spangaro, M.; Guillemin, G.J.; Comai, S.; Bosia, M. Importance of the dysregulation of the kynurenine pathway on cognition in schizophrenia: A systematic review of clinical studie. Eur. Arch. Psychiatry Clin. Neurosci. 2022; Epub ahead of print. [Google Scholar] [CrossRef]
- de Bartolomeis, A.; Barone, A.; Vellucci, L.; Mazza, B.; Austin, M.C.; Iasevoli, F.; Ciccarelli, M. Linking Inflammation, Aberrant Glutamate-Dopamine Interaction, and Post-synaptic Changes: Translational Relevance for Schizophrenia and Antipsychotic Treatment: A Systematic Review. Mol. Neurobiol. 2022, 59, 6460–6501. [Google Scholar] [CrossRef]
- Xia, M.; Abazyan, S.; Jouroukhin, Y.; Pletnikov, M. Behavioral sequelae of astrocyte dysfunction: Focus on animal models of schizophrenia. Schizophr. Res. 2016, 176, 72–82. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.Y.; Luo, D.Z.; Pei, J.C.; Kuo, M.C.; Hsieh, Y.C.; Lai, W.S. Not Just a Bystander: The Emerging Role of Astrocytes and Research Tools in Studying Cognitive Dysfunctions in Schizophrenia. Int. J. Mol. Sci. 2021, 22, 5343. [Google Scholar] [CrossRef]
- Martins-de-Souza, D.; Lebar, M.; Turck, C.W. Proteome analyses of cultured astrocytes treated with MK-801 and clozapine: Similarities with schizophrenia. Eur. Arch. Psychiatry Clin. Neurosci. 2011, 261, 217–228. [Google Scholar] [CrossRef]
- Tanaka, M.; Szabó, Á.; Vécsei, L. Integrating Armchair, Bench, and Bedside Research for Behavioral Neurology and Neuropsychiatry: Editorial. Biomedicines 2022, 10, 2999. [Google Scholar] [CrossRef]
- Momtazmanesh, S.; Zare-Shahabadi, A.; Rezaei, N. Cytokine Alterations in Schizophrenia: An Updated Review. Front. Psychiatry 2019, 10, 892. [Google Scholar] [CrossRef] [Green Version]
- Miller, B.J.; Buckley, P.; Seabolt, W.; Mellor, A.; Kirkpatrick, B. Meta-analysis of cytokine alterations in schizophrenia: Clinical status and antipsychotic effects. Biol. Psychiatry 2011, 70, 663–671. [Google Scholar] [CrossRef] [Green Version]
- Goldsmith, D.R.; Rapaport, M.H.; Miller, B.J. A meta-analysis of blood cytokine network alterations in psychiatric patients: Comparisons between schizophrenia, bipolar disorder and depression. Mol. Psychiatry 2016, 21, 1696–1709. [Google Scholar] [CrossRef]
- Wang, A.K.; Miller, B.J. Meta-analysis of Cerebrospinal Fluid Cytokine and Tryptophan Catabolite Alterations in Psychiatric Patients: Comparisons Between Schizophrenia, Bipolar Disorder, and Depression. Schizophr. Bull. 2018, 44, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Gallego, J.A.; Blanco, E.A.; Husain-Krautter, S.; Madeline Fagen, E.; Moreno-Merino, P.; Del Ojo-Jimenez, J.A.; Ahmed, A.; Rothstein, T.L.; Lencz, T.; Malhotra, A.K. Cytokines in cerebrospinal fluid of patients with schizophrenia spectrum disorders: New data and an updated meta-analysis. Schizophr. Res. 2018, 202, 64–71. [Google Scholar] [CrossRef]
- Pandurangi, A.K.; Buckley, P.F. Inflammation, Antipsychotic Drugs, and Evidence for Effectiveness of Anti-inflammatory Agents in Schizophrenia. Curr. Top Behav. Neurosci. 2020, 44, 227–244. [Google Scholar] [CrossRef]
- Yuhas, Y.; Ashkenazi, S.; Berent, E.; Weizman, A. Immunomodulatory activity of ketamine in human astroglial A172 cells: Possible relevance to its rapid antidepressant activity. J. Neuroimmunol. 2015, 282, 33–38. [Google Scholar] [CrossRef]
- Baharav, E.; Bar, M.; Taler, M.; Gil-Ad, I.; Karp, L.; Weinberger, A.; Weizman, A. Immunomodulatory effect of sertraline in a rat model of rheumatoid arthritis. Neuroimmunomodulation 2012, 19, 309–318. [Google Scholar] [CrossRef]
- Taler, M.; Gil-Ad, I.; Korob, I.; Weizman, A. The immunomodulatory effect of the antidepressant sertraline in an experimental autoimmune encephalomyelitis mouse model of multiple sclerosis. Neuroimmunomodulation 2011, 18, 117–122. [Google Scholar] [CrossRef]
- Gammon, D.; Cheng, C.; Volkovinskaia, A.; Baker, G.B.; Dursun, S.M. Clozapine: Why Is It So Uniquely Effective in the Treatment of a Range of Neuropsychiatric Disorders? Biomolecules 2021, 11, 1030. [Google Scholar] [CrossRef]
- Fukuyama, K.; Okubo, R.; Murata, M.; Shiroyama, T.; Okada, M. Activation of Astroglial Connexin is Involved in Concentration-Dependent Double-Edged Sword Clinical Action of Clozapine. Cells 2020, 9, 414. [Google Scholar] [CrossRef] [Green Version]
- Khokhar, J.Y.; Henricks, A.M.; Sullivan, E.D.K.; Green, A.I. Unique Effects of Clozapine: A Pharmacological Perspective. Adv. Pharmacol. 2018, 82, 137–162. [Google Scholar] [CrossRef] [PubMed]
- Baumeister, D.; Ciufolini, S.; Mondelli, V. Effects of psychotropic drugs on inflammation: Consequence or mediator of therapeutic effects in psychiatric treatment? Psychopharmacology 2016, 233, 1575–1589. [Google Scholar] [CrossRef]
- Zlatković, J.; Todorović, N.; Tomanović, N.; Bošković, M.; Djordjević, S.; Lazarević-Pašti, T.; Bernardi, R.E.; Djurdjević, A.; Filipović, D. Chronic administration of fluoxetine or clozapine induces oxidative stress in rat liver: A histopathological study. Eur. J. Pharm. Sci. 2014, 59, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Zhou, H.; Zhang, D.; Yang, S.; Qian, L.; Wu, H.M.; Chen, P.S.; Wilson, B.; Gao, H.M.; Lu, R.B.; et al. Clozapine protects dopaminergic neurons from inflammation-induced damage by inhibiting microglial overactivation. J. Neuroimmune Pharmacol. 2012, 7, 187–201. [Google Scholar] [CrossRef] [Green Version]
- Jeon, S.; Kim, S.H.; Shin, S.Y.; Lee, Y.H. Clozapine reduces Toll-like receptor 4/NF-kappaB-mediated inflammatory responses through inhibition of calcium/calmodulin-dependent Akt activation in microglia. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 81, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Giridharan, V.V.; Scaini, G.; Colpo, G.D.; Doifode, T.; Pinjari, O.F.; Teixeira, A.L.; Petronilho, F.; Macêdo, D.; Quevedo, J.; Barichello, T. Clozapine Prevents Poly (I:C) Induced Inflammation by Modulating NLRP3 Pathway in Microglial Cells. Cells 2020, 9, 577. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, H.G.; Steiner, J.; Bogerts, B. Glial cells in schizophrenia: Pathophysiological significance and possible consequences for therapy. Expert. Rev. Neurother. 2009, 9, 1059–1071. [Google Scholar] [CrossRef]
- Minghetti, L. Cyclooxygenase-2 (COX-2) in inflammatory and degenerative brain diseases. J. Neuropathol. Exp. Neurol. 2004, 63, 901–910. [Google Scholar] [CrossRef]
- De Kleijn, K.M.A.; Straasheijm, K.R.; Zuure, W.A.; Martens, G.J.M. Molecular Signature of Neuroinflammation Induced in Cytokine-Stimulated Human Cortical Spheroids. Biomedicines 2022, 10, 25. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, Y.; Iwakura, Y.; Sotoyama, H.; Kitayama, E.; Takei, N.; Someya, T.; Nawa, H. Clozapine-dependent inhibition of EGF/neuregulin receptor (ErbB) kinases. Transl. Psychiatry 2019, 9, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuhas, Y.; Azoulay-Alfaguter, I.; Berent, E.; Ashkenazi, S. Rifampin inhibits prostaglandin E2 production and arachidonic acid release in human alveolar epithelial cells. Antimicrob. Agents Chemother. 2007, 51, 4225–4230. [Google Scholar] [CrossRef] [Green Version]
- Yuhas, Y.; Berent, E.; Ashkenazi, S. Effect of rifampin on production of inflammatory mediators in HepG2 liver epithelial cells. Antimicrob. Agents Chemother. 2011, 55, 5541–5546. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Simon, L.S. Role and regulation of cyclooxygenase-2 during inflammation. Am. J. Med. 1999, 106, 37s–42s. [Google Scholar] [CrossRef]
- Tang, B.; Capitao, C.; Dean, B.; Thomas, E.A. Differential age- and disease-related effects on the expression of genes related to the arachidonic acid signaling pathway in schizophrenia. Psychiatry Res. 2012, 196, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.L.; Tsai, T.C.; Lin, Y.Y.; Tsai, Y.M.; Wang, L.K.; Lee, M.C.; Tsai, F.M. Antipsychotic drugs suppress the AKT/NF-κB pathway and regulate the differentiation of T-cell subsets. Immunol. Lett. 2011, 140, 81–91. [Google Scholar] [CrossRef]
- Shin, S.Y.; Choi, B.H.; Ko, J.; Kim, S.H.; Kim, Y.S.; Lee, Y.H. Clozapine, a neuroleptic agent, inhibits Akt by counteracting Ca2+/calmodulin in PTEN-negative U-87MG human glioblastoma cells. Cell Signal. 2006, 18, 1876–1886. [Google Scholar] [CrossRef]
- Kowalchuk, C.; Kanagasundaram, P.; Belsham, D.D.; Hahn, M.K. Antipsychotics differentially regulate insulin, energy sensing, and inflammation pathways in hypothalamic rat neurons. Psychoneuroendocrinology 2019, 104, 42–48. [Google Scholar] [CrossRef]
- Lu, X.H.; Dwyer, D.S. Second-generation antipsychotic drugs, olanzapine, quetiapine, and clozapine enhance neurite outgrowth in PC12 cells via PI3K/AKT, ERK, and pertussis toxin-sensitive pathways. J. Mol. Neurosci. 2005, 27, 43–64. [Google Scholar] [CrossRef]
- Zeng, Z.; Wang, X.; Bhardwaj, S.K.; Zhou, X.; Little, P.J.; Quirion, R.; Srivastava, L.K.; Zheng, W. The Atypical Antipsychotic Agent, Clozapine, Protects Against Corticosterone-Induced Death of PC12 Cells by Regulating the Akt/FoxO3a Signaling Pathway. Mol. Neurobiol. 2017, 54, 3395–3406. [Google Scholar] [CrossRef]
- Aringhieri, S.; Kolachalam, S.; Gerace, C.; Carli, M.; Verdesca, V.; Brunacci, M.G.; Rossi, C.; Ippolito, C.; Solini, A.; Corsini, G.U.; et al. Clozapine as the most efficacious antipsychotic for activating ERK 1/2 kinases: Role of 5-HT(2A) receptor agonism. Eur. Neuropsychopharmacol. 2017, 27, 383–398. [Google Scholar] [CrossRef]
- Schneemann, M.; Schoeden, G. Macrophage biology and immunology: Man is not a mouse. J. Leukoc. Biol. 2007, 81, 579. [Google Scholar] [CrossRef]
- Young, R.; Bush, S.J.; Lefevre, L.; McCulloch, M.E.B.; Lisowski, Z.M.; Muriuki, C.; Waddell, L.A.; Sauter, K.A.; Pridans, C.; Clark, E.L.; et al. Species-Specific Transcriptional Regulation of Genes Involved in Nitric Oxide Production and Arginine Metabolism in Macrophages. Immunohorizons 2018, 2, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Contestabile, A. Regulation of transcription factors by nitric oxide in neurons and in neural-derived tumor cells. Prog. Neurobiol. 2008, 84, 317–328. [Google Scholar] [CrossRef]
- Yuhas, Y.; Berent, E.; Ashkenazi, S. Effect of nitric oxide on microRNA-155 expression in human hepatic epithelial cells. Inflamm. Res. 2014, 63, 591–596. [Google Scholar] [CrossRef]
- Hewett, S.J.; Bell, S.C.; Hewett, J.A. Contributions of cyclooxygenase-2 to neuroplasticity and neuropathology of the central nervous system. Pharmacol. Ther. 2006, 112, 335–357. [Google Scholar] [CrossRef]
- Horrobin, D.F.; Glen, A.I.; Vaddadi, K. The membrane hypothesis of schizophrenia. Schizophr. Res. 1994, 13, 195–207. [Google Scholar] [CrossRef]
- Yang, X.; Li, M.; Jiang, J.; Hu, X.; Qing, Y.; Sun, L.; Yang, T.; Wang, D.; Cui, G.; Gao, Y.; et al. Dysregulation of phospholipase and cyclooxygenase expression is involved in Schizophrenia. EBioMedicine 2021, 64, 103239. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.K.; van Kammen, D.P.; Welker, J.A. Red blood cell membrane dynamics in schizophrenia. II. Fatty acid composition. Schizophr. Res. 1994, 13, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Maida, M.E.; Hurley, S.D.; Daeschner, J.A.; Moore, A.H.; O’Banion, M.K. Cytosolic prostaglandin E2 synthase (cPGES) expression is decreased in discrete cortical regions in psychiatric disease. Brain Res. 2006, 1103, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Holscher, C. Prostaglandins play a role in memory consolidation in the chick. Eur. J. Pharmacol. 1995, 294, 253–259. [Google Scholar] [CrossRef]
- Nitta, M.; Kishimoto, T.; Muller, N.; Weiser, M.; Davidson, M.; Kane, J.M.; Correll, C.U. Adjunctive use of nonsteroidal anti-inflammatory drugs for schizophrenia: A meta-analytic investigation of randomized controlled trials. Schizophr. Bull. 2013, 39, 1230–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cianciulli, A.; Porro, C.; Calvello, R.; Trotta, T.; Lofrumento, D.D.; Panaro, M.A. Microglia Mediated Neuroinflammation: Focus on PI3K Modulation. Biomolecules 2020, 10, 137. [Google Scholar] [CrossRef] [Green Version]
- Weaver, S.A.; Russo, M.P.; Wright, K.L.; Kolios, G.; Jobin, C.; Robertson, D.A.; Ward, S.G. Regulatory role of phosphatidylinositol 3-kinase on TNF-alpha-induced cyclooxygenase 2 expression in colonic epithelial cells. Gastroenterology 2001, 120, 1117–1127. [Google Scholar] [CrossRef]
- Monick, M.M.; Robeff, P.K.; Butler, N.S.; Flaherty, D.M.; Carter, A.B.; Peterson, M.W.; Hunninghake, G.W. Phosphatidylinositol 3-kinase activity negatively regulates stability of cyclooxygenase 2 mRNA. J. Biol. Chem. 2002, 277, 32992–33000. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, A.C.; Candelario-Jalil, E.; Langbein, J.; Wendeburg, L.; Bhatia, H.S.; Schlachetzki, J.C.; Biber, K.; Fiebich, B.L. Pharmacological inhibition of Akt and downstream pathways modulates the expression of COX-2 and mPGES-1 in activated microglia. J. Neuroinflammation 2012, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.J.; Mbonye, U.R.; DeLong, C.J.; Wada, M.; Smith, W.L. Regulation of intracellular cyclooxygenase levels by gene transcription and protein degradation. Prog. Lipid Res. 2007, 46, 108–125. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal. Transduct. Target Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Al Abadey, A.; Connor, B.; Flamme, A.C.; Robichon, K. Clozapine reduces chemokine-mediated migration of lymphocytes by targeting NF-κB and AKT phosphorylation. Cell Signal. 2022, 99, 110449. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Dai, J.; Tang, L.; Mikhailova, T.; Liang, Q.; Li, M.; Zhou, J.; Kopp, R.F.; Weickert, C.; Chen, C.; et al. Neuroimmune transcriptome changes in patient brains of psychiatric and neurological disorders. Mol. Psychiatry. 2022; Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inflammatory Mediator | Relative mRNA Expression | Relative Protein Production | ||||||
|---|---|---|---|---|---|---|---|---|
| Cytokine Mix | Cytokine Mix + Clozapine | t | p | Cytokine Mix | Cytokine Mix + Clozapine | t | p | |
| % | % ±SEM | % | % ±SEM | |||||
| TNFα | 100 | 63.4 ± 6.5 | 5.68 | <0.001 | 100 | 81 ± 4 | 4.07 | <0.05 |
| IL-1β | 100 | 63.6 ± 5.2 | 4.78 | <0.001 | 100 | 73 ± 1.4 | 8.48 | <0.001 |
| IL-8 | 100 | 76.1 ± 3.6 | 13.46 | <0.0001 | 100 | 35 ± 4.1 | 8.51 | <0.001 |
| COX2 | 100 | 186 ± 7.5 | 11.67 | <0.0001 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuhas, Y.; Ashkenazi, S.; Berent, E.; Weizman, A. Clozapine Suppresses the Gene Expression and the Production of Cytokines and Up-Regulates Cyclooxygenase 2 mRNA in Human Astroglial Cells. Brain Sci. 2022, 12, 1703. https://doi.org/10.3390/brainsci12121703
Yuhas Y, Ashkenazi S, Berent E, Weizman A. Clozapine Suppresses the Gene Expression and the Production of Cytokines and Up-Regulates Cyclooxygenase 2 mRNA in Human Astroglial Cells. Brain Sciences. 2022; 12(12):1703. https://doi.org/10.3390/brainsci12121703
Chicago/Turabian StyleYuhas, Yael, Shai Ashkenazi, Eva Berent, and Abraham Weizman. 2022. "Clozapine Suppresses the Gene Expression and the Production of Cytokines and Up-Regulates Cyclooxygenase 2 mRNA in Human Astroglial Cells" Brain Sciences 12, no. 12: 1703. https://doi.org/10.3390/brainsci12121703