
Acute Sleep Deprivation Impairs Motor Inhibition in Table Tennis Athletes: An ERP Study

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
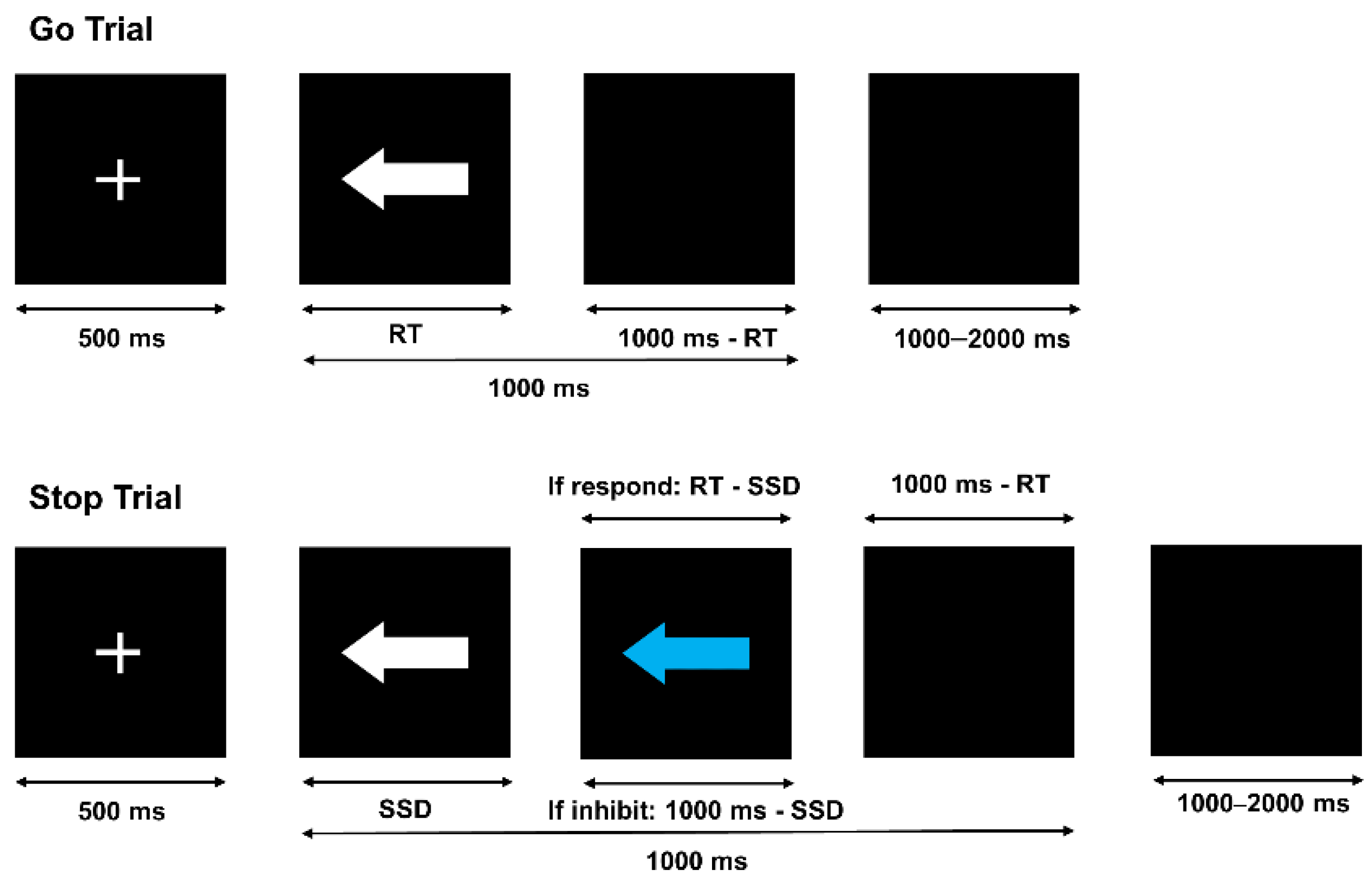
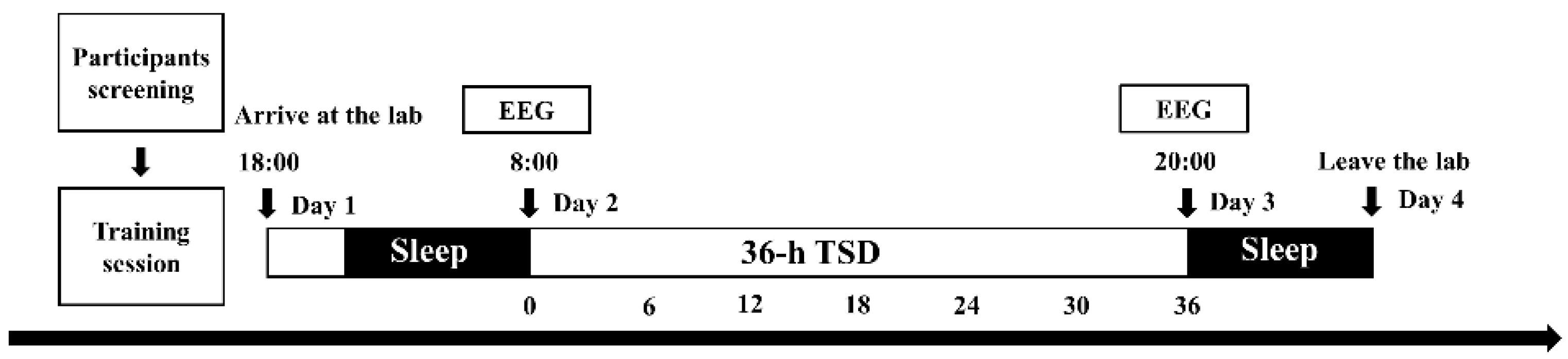
2.2. Experimental Design
2.3. Experimental Procedures
2.4. Data Acquisition and Analysis
2.4.1. Behavioral Data Acquisition
- (1)
- Go RT: Time to correct response to the go trials.
- (2)
- Go Accuracy: % correct responses in go trials.
- (3)
- Mean SSD: The average SSD of all stop trials.
- (4)
- Quantile RT (QRT): The response times of all the correct responses of the go trials were sorted in ascending order, and the go RT at the percentage of inhibition failure was taken as the QRT.
- (5)
- SSRT: QRT-mean SSD.
- (6)
- Stop Accuracy: % correct inhibited responses in stop trials.
2.4.2. ERP Data Acquisition
Analysis of N2 and P3 Amplitudes
Analysis of ERN Amplitude
2.4.3. Data Analysis
3. Results
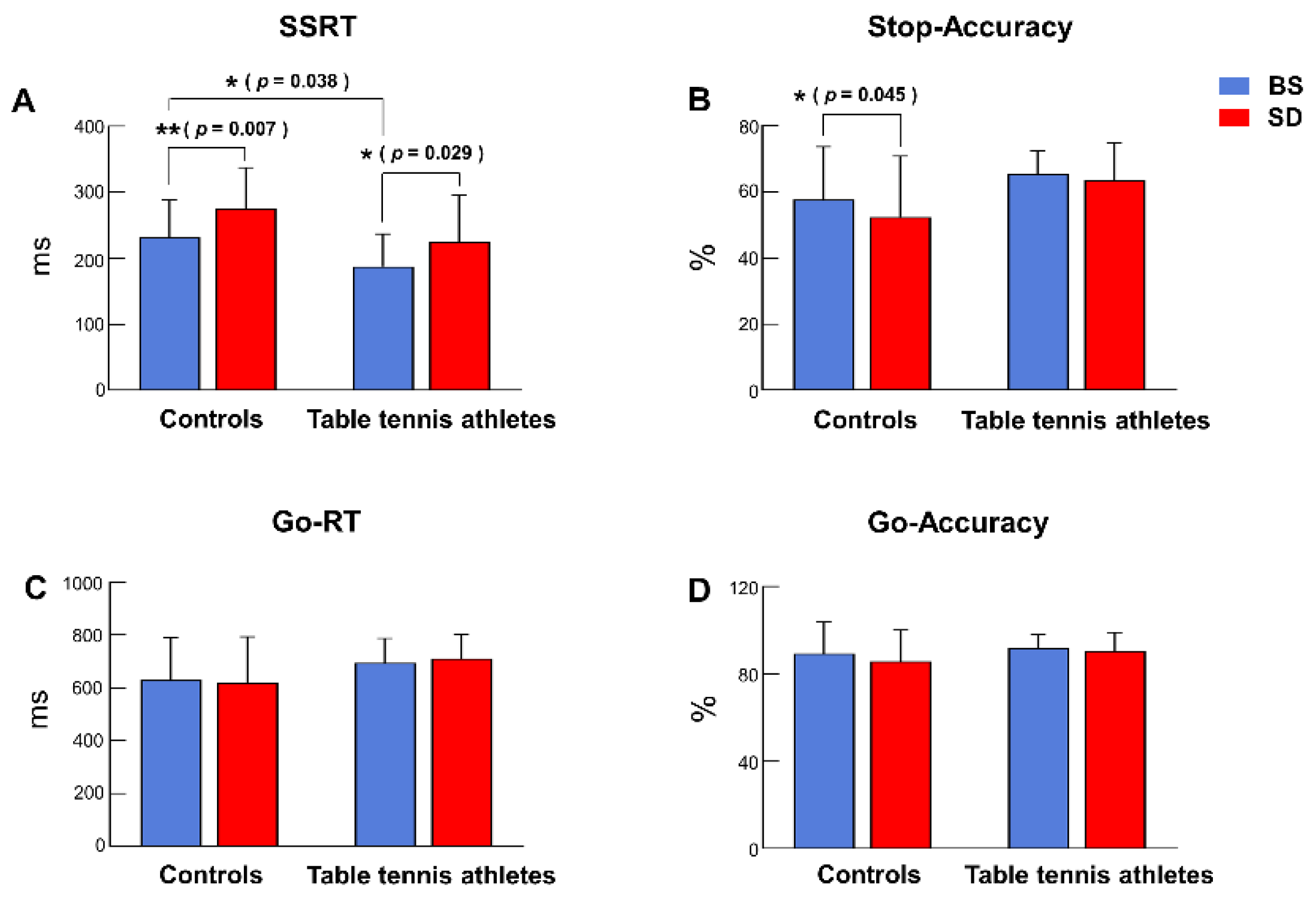
3.1. Behavioral Performance
3.1.1. SSRT
3.1.2. Stop Accuracy
3.1.3. Go-RT
3.1.4. Go Accuracy
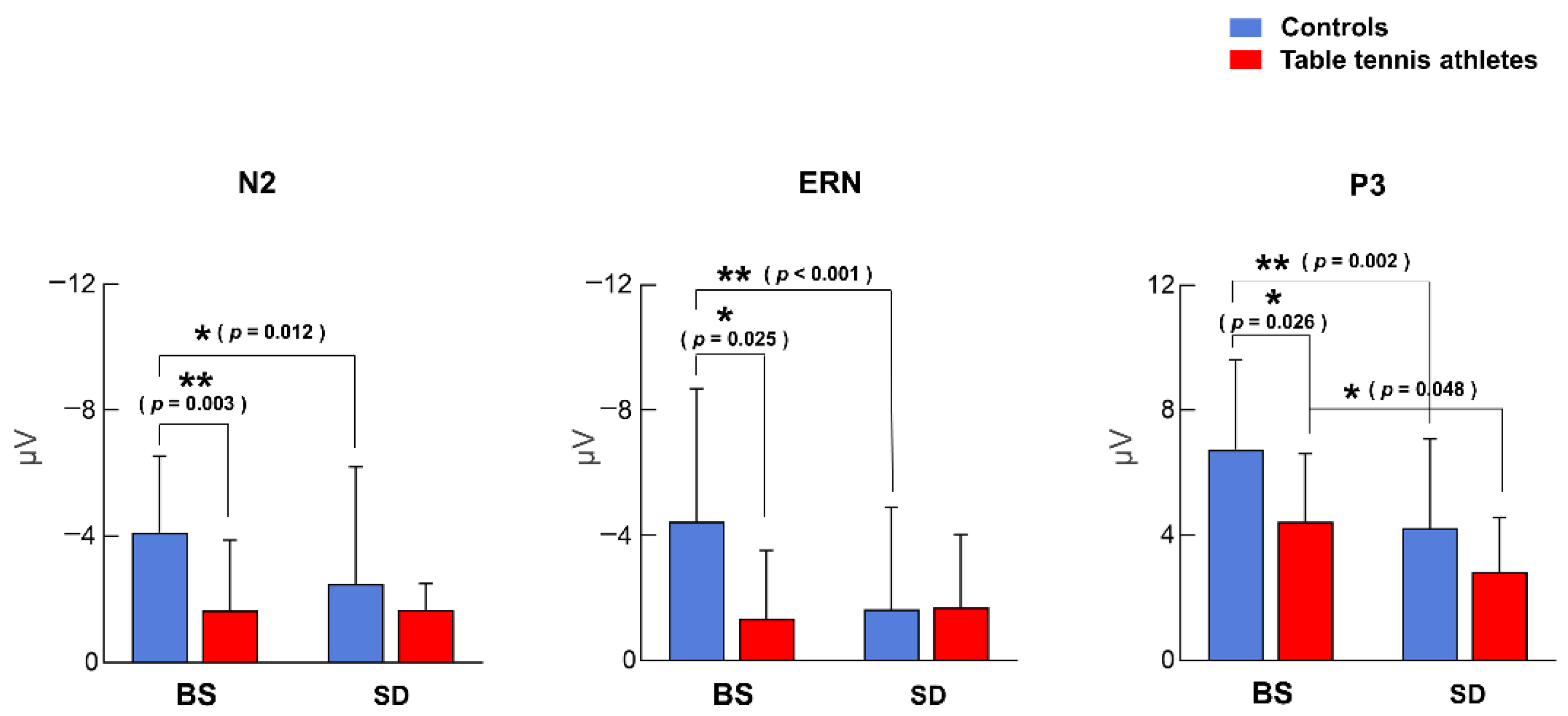
3.2. ERP Component Amplitude
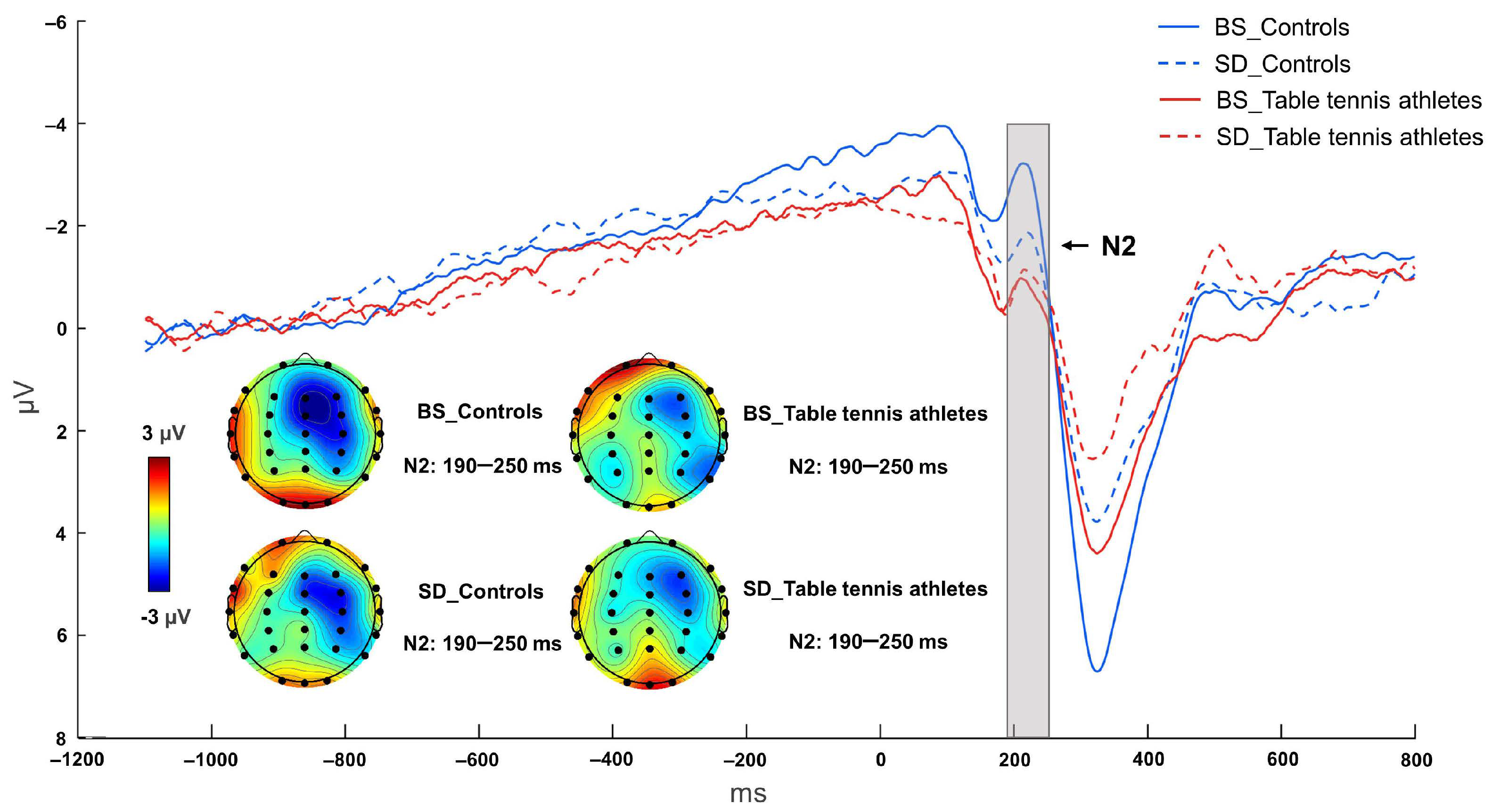
3.2.1. N2 Component
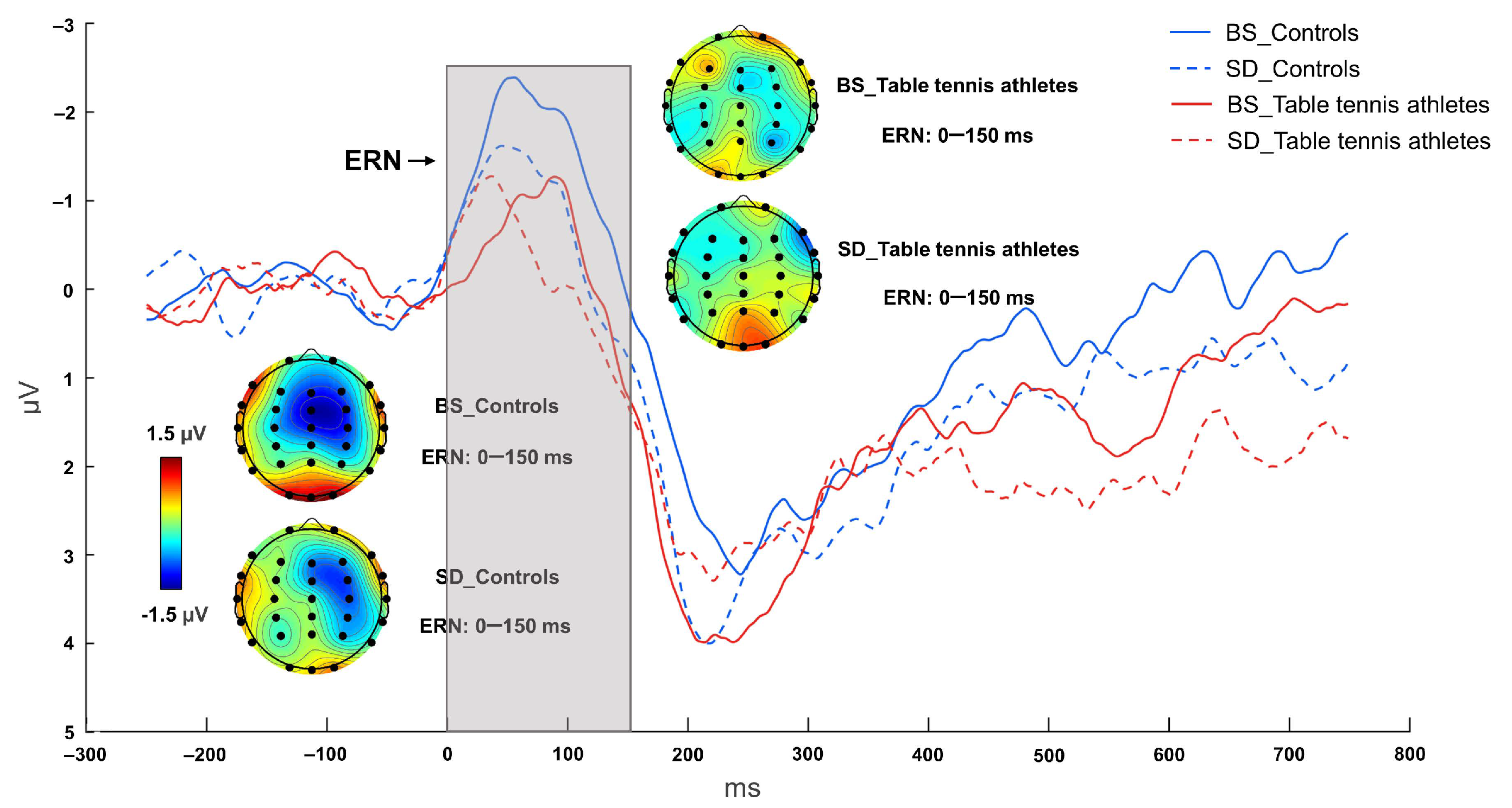
3.2.2. ERN Component
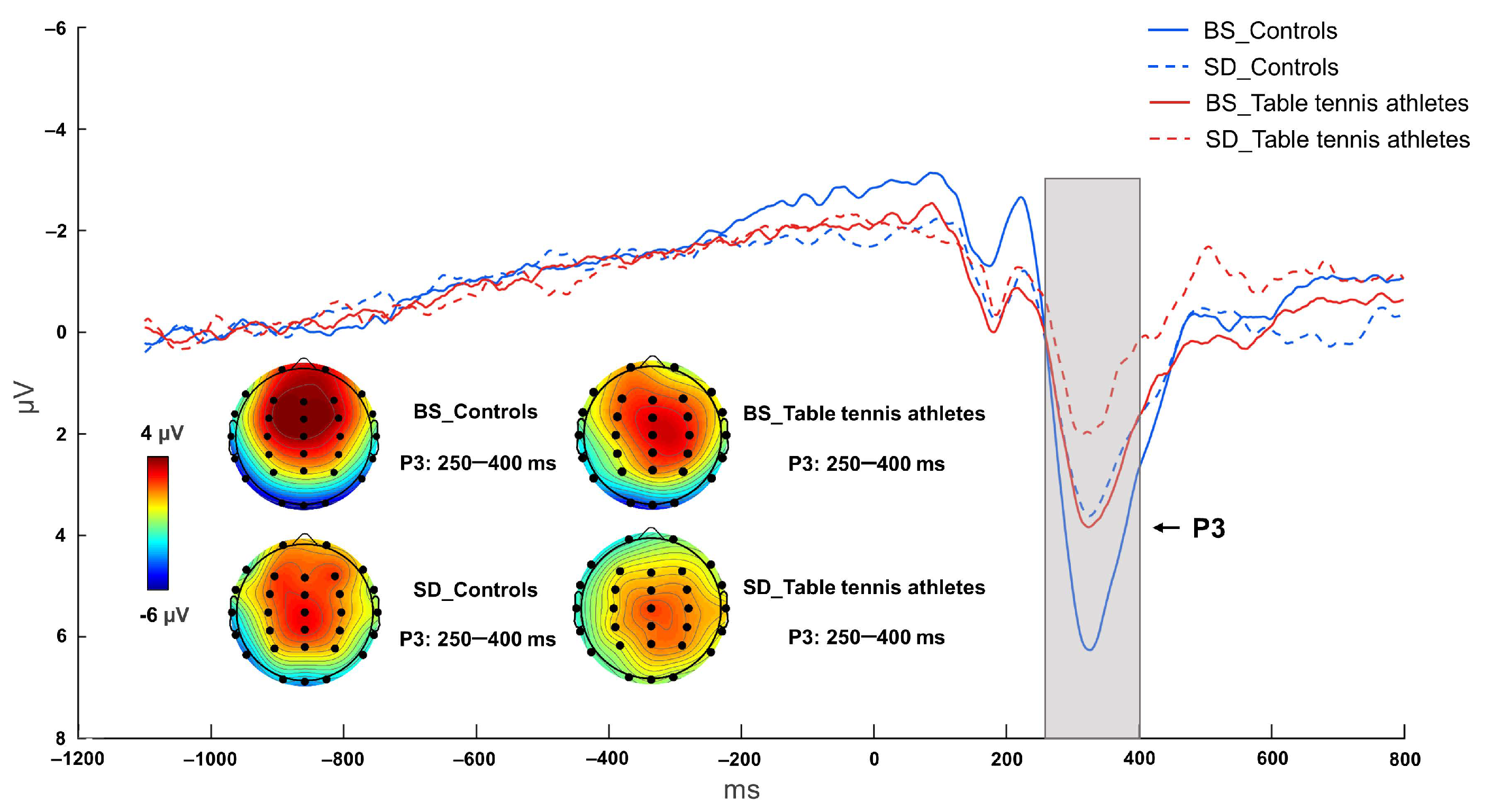
3.2.3. P3 Component
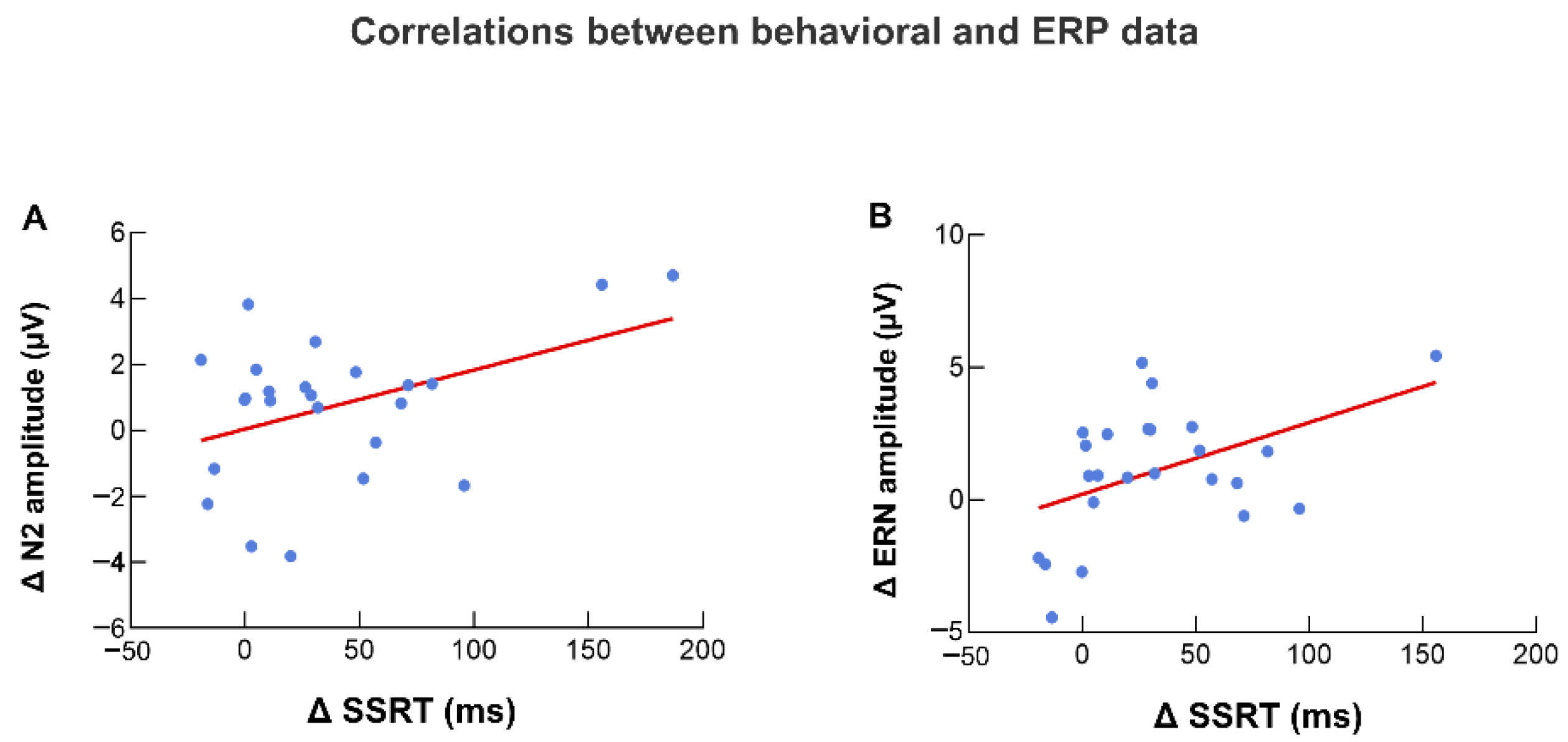
3.3. Correlation between EEG and Behavioral Data
3.3.1. N2 Amplitude and SSRT
3.3.2. ERN Amplitude and SSRT
3.3.3. P3 Amplitude and SSRT
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barnes, S. “The Best Recovery You Could Possibly Get”: Sleep, Rest, and the National Basketball Association. Sociol. Sport J. 2021, 38, 16–25. [Google Scholar] [CrossRef]
- Andrews, J.L.; Zhang, X.; McCarthy, J.J.; McDearmon, E.L.; Hornberger, T.A.; Russell, B.; Campbell, K.S.; Arbogast, S.; Reid, M.B.; Walker, J.R.; et al. CLOCK and BMAL1 regulate MyoD and are necessary for maintenance of skeletal muscle phenotype and function. Proc. Natl. Acad. Sci. USA 2010, 107, 19090–19095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, J.P.P.; Rodrigues, D.F.; Silva, A.; Simim, M.A.D.M.; Costa, V.T.; Noce, F.; de Mello, M.T. 2016 Rio Olympic Games: Can the schedule of events compromise athletes’ performance? Chronobiol. Int. 2016, 33, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Erlacher, D.; Ehrlenspiel, F.; Adegbesan, O.A.; El-Din, H.G. Sleep habits in German athletes before important competitions or games. J. Sports Sci. 2011, 29, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Juliff, L.E.; Halson, S.L.; Peiffer, J.J. Understanding sleep disturbance in athletes prior to important competitions. J. Sci. Med. Sport 2015, 18, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Leeder, J.; Glaister, M.; Pizzoferro, K.; Dawson, J.; Pedlar, C. Sleep duration and quality in elite athletes measured using wristwatch actigraphy. J. Sports Sci. 2012, 30, 541–545. [Google Scholar] [CrossRef]
- Nédélec, M.; Halson, S.; Abaidia, A.-E.; Ahmaidi, S.; Dupont, G. Stress, Sleep and Recovery in Elite Soccer: A Critical Review of the Literature. Sports Med. 2015, 45, 1387–1400. [Google Scholar] [CrossRef]
- Venter, R. Perceptions of team athletes on the importance of recovery modalities. Eur. J. Sport Sci. 2011, 14, S69–S76. [Google Scholar] [CrossRef]
- Charest, J.; Grandner, M.A. Sleep and Athletic Performance: Impacts on Physical Performance, Mental Performance, Injury Risk and Recovery, and Mental Health. Sleep Med. Clin. 2020, 15, 41–57. [Google Scholar] [CrossRef]
- Fullagar, H.H.K.; Skorski, S.; Duffield, R.; Hammes, D.; Coutts, A.J.; Meyer, T. Sleep and Athletic Performance: The Effects of Sleep Loss on Exercise Performance, and Physiological and Cognitive Responses to Exercise. Sports Med. 2014, 45, 161–186. [Google Scholar] [CrossRef]
- Jarraya, S.; Jarraya, M.; Chtourou, H.; Souissi, N. Effect of time of day and partial sleep deprivation on the reaction time and the attentional capacities of the handball goalkeeper. Biol. Rhythm Res. 2013, 45, 183–191. [Google Scholar] [CrossRef]
- Minkel, J.D.; Banks, S.; Htaik, O.; Moreta, M.C.; Jones, C.W.; McGlinchey, E.L.; Simpson, N.S.; Dinges, D.F. Sleep deprivation and stressors: Evidence for elevated negative affect in response to mild stressors when sleep deprived. Emotion 2012, 12, 1015–1020. [Google Scholar] [CrossRef] [Green Version]
- Reyner, L.; Horne, J. Sleep restriction and serving accuracy in performance tennis players, and effects of caffeine. Physiol. Behav. 2013, 120, 93–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riederer, M.F. How Sleep Impacts Performance in Youth Athletes. Curr. Sports Med. Rep. 2020, 19, 463–467. [Google Scholar] [CrossRef]
- Skein, M.; Duffield, R.; Edge, J.; Short, M.J.; Mündel, T. Intermittent-Sprint Performance and Muscle Glycogen after 30 h of Sleep Deprivation. Med. Sci. Sports Exerc. 2011, 43, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- Taheri, M.; Arabameri, E. The Effect of Sleep Deprivation on Choice Reaction Time and Anaerobic Power of College Student Athletes. Asian J. Sports Med. 2012, 3, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, A.M. Sleep and Athletic Performance. Curr. Sports Med. Rep. 2017, 16, 413–418. [Google Scholar] [CrossRef]
- Diamond, A. Executive Functions. Annu. Rev. Psychol. 2013, 64, 135–168. [Google Scholar] [CrossRef] [Green Version]
- Verbruggen, F.; Logan, G.D. Models of response inhibition in the stop-signal and stop-change paradigms. Neurosci. Biobehav. Rev. 2009, 33, 647–661. [Google Scholar] [CrossRef] [Green Version]
- Logan, G.D.; Cowan, W.B.; Davis, K.A. On the ability to inhibit simple and choice reaction time responses: A model and a method. J. Exp. Psychol. Hum. Percept. Perform. 1984, 10, 276–291. [Google Scholar] [CrossRef]
- Kusztor, A.; Raud, L.; Juel, B.E.; Nilsen, A.S.; Storm, J.F.; Huster, R.J. Sleep deprivation differentially affects subcomponents of cognitive control. Sleep 2019, 42, zsz016. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.; Platten, C.R. Sleep deprivation lowers inhibition and enhances impulsivity to negative stimuli. Behav. Brain Res. 2011, 217, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Drummond, S.P.A.; Paulus, M.P.; Tapert, S.F. Effects of two nights sleep deprivation and two nights recovery sleep on response inhibition. J. Sleep Res. 2006, 15, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Killgore, W.D. Effects of sleep deprivation on cognition. Prog. Brain Res. 2010, 185, 105–129. [Google Scholar] [CrossRef] [PubMed]
- Chuah, Y.M.L.; Venkatraman, V.; Dinges, D.F.; Chee, M.W.L. The Neural Basis of Interindividual Variability in Inhibitory Efficiency after Sleep Deprivation. J. Neurosci. 2006, 26, 7156–7162. [Google Scholar] [CrossRef]
- Jin, X.; Ye, E.; Qi, J.; Wang, L.; Lei, Y.; Chen, P.; Mi, G.; Zou, F.; Shao, Y.; Yang, Z. Recovery Sleep Reverses Impaired Response Inhibition due to Sleep Restriction: Evidence from a Visual Event Related Potentials Study. PLoS ONE 2015, 10, e0142361. [Google Scholar] [CrossRef]
- Kelly, C.; Garavan, H. Human Functional Neuroimaging of Brain Changes Associated with Practice. Cereb. Cortex 2004, 15, 1089–1102. [Google Scholar] [CrossRef] [Green Version]
- Poldrack, R. Imaging Brain Plasticity: Conceptual and Methodological Issues—A Theoretical Review. NeuroImage 2000, 12, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Picton, T.W.; Lins, O.G.; Scherg, M. The recording and analysis of event related potentials. In Handbook of Neuropsychology; Boller, F., Grafman, J., Eds.; Elsevier: Amsterdam, The Netherlands, 1995. [Google Scholar]
- Groom, M.J.; Cragg, L. Differential modulation of the N2 and P3 event-related potentials by response conflict and inhibition. Brain Cogn. 2015, 97, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ramautar, J.R.; Kok, A.; Ridderinkhof, K.R. Effects of stop-signal modality on the N2/P3 complex elicited in the stop-signal paradigm. Biol. Psychol. 2006, 72, 96–109. [Google Scholar] [CrossRef]
- Wessel, J.R.; Aron, A.R. It’s not too late: The onset of the frontocentral P3 indexes successful response inhibition in the stop-signal paradigm. Psychophysiology 2015, 52, 472–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donkers, F.C.; van Boxtel, G. The N2 in go/no-go tasks reflects conflict monitoring not response inhibition. Brain Cogn. 2004, 56, 165–176. [Google Scholar] [CrossRef]
- Dong, G.; Yang, L.; Hu, Y.; Jiang, Y. Is N2 associated with successful suppression of behavior responses in impulse control processes? NeuroReport 2009, 20, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Folstein, J.R.; Van Petten, C. Influence of cognitive control and mismatch on the N2 component of the ERP: A review. Psychophysiology 2007, 45, 152–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enriquez-Geppert, S.; Konrad, C.; Pantev, C.; Huster, R.J. Conflict and inhibition differentially affect the N200/P300 complex in a combined go/nogo and stop-signal task. NeuroImage 2010, 51, 877–887. [Google Scholar] [CrossRef]
- Albert, J.; Lopez-Martin, S.; Hinojosa, J.; Carretié, L. Spatiotemporal characterization of response inhibition. NeuroImage 2013, 76, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Wessel, J.R. Prepotent motor activity and inhibitory control demands in different variants of the go/no-go paradigm. Psychophysiology 2017, 55, e12871. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-S.R.; Huang, C.; Constable, R.T.; Sinha, R. Imaging Response Inhibition in a Stop-Signal Task: Neural Correlates Independent of Signal Monitoring and Post-Response Processing. J. Neurosci. 2006, 26, 186–192. [Google Scholar] [CrossRef]
- Gehring, W.J.; Goss, B.; Coles, M.G.H.; Meyer, D.E.; Donchin, E. The Error-Related Negativity. Perspect. Psychol. Sci. 2018, 13, 200–204. [Google Scholar] [CrossRef]
- Riesel, A. The erring brain: Error-related negativity as an endophenotype for OCD-A review and meta-analysis. Psychophysiology 2019, 56, e13348. [Google Scholar] [CrossRef]
- Chang, W.-P.; Davies, P.L.; Gavin, W.J. Individual differences in error monitoring in healthy adults: Psychological symptoms and antisocial personality characteristics. Eur. J. Neurosci. 2010, 32, 1388–1396. [Google Scholar] [CrossRef] [PubMed]
- Logan, G.D.; Schachar, R.J.; Tannock, R. Impulsivity and Inhibitory Control. Psychol. Sci. 1997, 8, 60–64. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, M.-T.; Lu, H.; Lin, C.-I.; Sun, T.-H.; Chen, Y.-R.; Cheng, C.-H. Effects of Trait Anxiety on Error Processing and Post-error Adjustments: An Event-Related Potential Study with Stop-Signal Task. Front. Hum. Neurosci. 2021, 15, 650838. [Google Scholar] [CrossRef]
- Dienes, Z. Using Bayes to get the most out of non-significant results. Front. Psychol. 2014, 5, 781. [Google Scholar] [CrossRef] [Green Version]
- Wagenmakers, E.-J.; Love, J.; Marsman, M.; Jamil, T.; Ly, A.; Verhagen, J.; Selker, R.; Gronau, Q.F.; Dropmann, D.; Boutin, B.; et al. Bayesian inference for psychology. Part II: Example applications with JASP. Psychon. Bull. Rev. 2018, 25, 58–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffreys, H. Theory of Probability, 3rd ed.; Oxford University Press: Oxford, UK, 1961. [Google Scholar]
- Boucher, L.; Palmeri, T.J.; Logan, G.D.; Schall, J.D. Inhibitory control in mind and brain: An interactive race model of countermanding saccades. Psychol. Rev. 2007, 114, 376–397. [Google Scholar] [CrossRef] [PubMed]
- Logan, G.D.; Van Zandt, T.; Verbruggen, F.; Wagenmakers, E.-J. On the ability to inhibit thought and action: General and special theories of an act of control. Psychol. Rev. 2014, 121, 66–95. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-H.; Chang, C.-C.; Liang, Y.-M.; Shih, C.-M.; Chiu, W.-S.; Tseng, P.; Hung, D.L.; Tzeng, O.J.L.; Muggleton, N.G.; Juan, C.-H. Open vs. Closed Skill Sports and the Modulation of Inhibitory Control. PLoS ONE 2013, 8, e55773. [Google Scholar] [CrossRef] [Green Version]
- Nakamoto, H.; Mori, S. Sport-Specific Decision-Making in a Go/Nogo Reaction Task: Difference among Nonathletes and Baseball and Basketball Players. Percept. Mot. Skills 2008, 106, 163–170. [Google Scholar] [CrossRef]
- Nakamoto, H.; Mori, S. Effects of stimulus–response compatibility in mediating expert performance in baseball players. Brain Res. 2008, 1189, 179–188. [Google Scholar] [CrossRef]
- Vestberg, T.; Gustafson, R.; Maurex, L.; Ingvar, M.; Petrovic, P. Executive Functions Predict the Success of Top-Soccer Players. PLoS ONE 2012, 7, e34731. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, M.E.; VanMeter, J.; Janelle, C.M.; Braun, A.; Miller, M.W.; Oldham, J.; Russell, B.A.H.; Hatfield, B.D. Neural Efficiency in Expert Cognitive-Motor Performers during Affective Challenge. J. Mot. Behav. 2016, 48, 573–588. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, H.D.; Popp, C.; Reith, W.; Krick, C. Gains of item-specific training in visual working memory and their neural correlates. Brain Res. 2012, 1466, 44–55. [Google Scholar] [CrossRef]
- Neubauer, A.C.; Fink, A. Intelligence and neural efficiency. Neurosci. Biobehav. Rev. 2009, 33, 1004–1023. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Fang, C.; Li, L.; Song, Y. Impact of sport training on adaptations in neural functioning and behavioral performance: A scoping review with meta-analysis on EEG research. J. Exerc. Sci. Fit. 2022, 20, 206–215. [Google Scholar] [CrossRef]
- Smith, J.; Smith, E.A.; Provost, A.; Heathcote, A. Sequence effects support the conflict theory of N2 and P3 in the Go/NoGo task. Int. J. Psychophysiol. 2010, 75, 217–226. [Google Scholar] [CrossRef]
- Yuan, J.; Meng, X.; Yang, J.; Yao, G.; Hu, L.; Yuan, H. The valence strength of unpleasant emotion modulates brain processing of behavioral inhibitory control: Neural correlates. Biol. Psychol. 2012, 89, 240–251. [Google Scholar] [CrossRef]
- Di Gregorio, F.; Steinhauser, M.; Maier, M.E. Error-related brain activity and error awareness in an error classification paradigm. NeuroImage 2016, 139, 202–210. [Google Scholar] [CrossRef]
- Schulreich, S. Altered Performance Monitoring in Psychopathy: A Review of Studies on Action Selection, Error, and Feedback Processing. Curr. Behav. Neurosci. Rep. 2016, 3, 19–27. [Google Scholar] [CrossRef]
- Guo, Z.; Li, A.; Yu, L. “Neural Efficiency” of Athletes’ Brain during Visuo-Spatial Task: An fMRI Study on Table Tennis Players. Front. Behav. Neurosci. 2017, 11, 72. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Smith, D.M. Neural Efficiency in Athletes: A Systematic Review. Front. Behav. Neurosci. 2021, 15, 698555. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, K.; Ogyu, S.; Arakomo, Y.; Suzuki, K.; Mafune, K.; Hiro, H.; Nagata, S. Recovery of Cognitive Performance and Fatigue after One Night of Sleep Deprivation. J. Occup. Health 2009, 51, 412–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skurvydas, A.; Kazlauskaite, D.; Zlibinaite, L.; Cekanauskaite, A.; Valanciene, D.; Karanauskiene, D.; Zuoziene, I.J.; Majauskiene, D.; Mickeviciene, D.; Satas, A. Effects of two nights of sleep deprivation on executive function and central and peripheral fatigue during maximal voluntary contraction lasting 60s. Physiol. Behav. 2020, 229, 113226. [Google Scholar] [CrossRef] [PubMed]
- Wesensten, N.J.; Belenky, G.; Thorne, D.R.; Kautz, A.M.; Balkin, T.J. Modafinil vs. caffeine: Effects on fatigue during sleep deprivation. Aviat. Space Environ. Med. 2004, 75, 520–525. [Google Scholar]
- Cote, K.A.; Milner, C.E.; Smith, B.A.; Aubin, A.J.; Greason, T.A.; Cuthbert, B.P.; Wiebe, S.; Duffus, S.E.G. CNS arousal and neurobehavioral performance in a short-term sleep restriction paradigm. J. Sleep Res. 2009, 18, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Kropotov, J.D.; Ponomarev, V.A.; Hollup, S.; Mueller, A. Dissociating action inhibition, conflict monitoring and sensory mismatch into independent components of event related potentials in GO/NOGO task. NeuroImage 2011, 57, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Johnstone, S.; Barry, R. Movement-related potentials in the Go/NoGo task: The P3 reflects both cognitive and motor inhibition. Clin. Neurophysiol. 2008, 119, 704–714. [Google Scholar] [CrossRef]
- Qi, J.-L.; Shao, Y.-C.; Miao, D.; Fan, M.; Bi, G.-H.; Yang, Z. The Effects of 43 Hours of Sleep Deprivation on Executive Control Functions: Event-Related Potentials in a Visual Go/No Go Task. Soc. Behav. Personal. Int. J. 2010, 38, 29–42. [Google Scholar] [CrossRef]
- Liu, Q.; Zhou, R.; Liu, L.; Zhao, X. Effects of 72 hours total sleep deprivation on male astronauts’ executive functions and emotion. Compr. Psychiatry 2015, 61, 28–35. [Google Scholar] [CrossRef]
- Miyazaki, S.; Liu, C.-Y.; Hayashi, Y. Sleep in vertebrate and invertebrate animals, and insights into the function and evolution of sleep. Neurosci. Res. 2017, 118, 3–12. [Google Scholar] [CrossRef]
- Simpson, N.S.; Gibbs, E.L.; Matheson, G.O. Optimizing sleep to maximize performance: Implications and recommendations for elite athletes. Scand. J. Med. Sci. Sports 2016, 27, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, R.K. Sleep, Recovery, and Performance in Sports. Neurol. Clin. 2017, 35, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.; Reddy, V.; Sharma, S. Physiology, Circadian Rhythm; StatPearls Publishing: Tampa, FL, USA, 2021.
- Clark, I.; Landolt, H.-P. Coffee, caffeine, and sleep: A systematic review of epidemiological studies and randomized controlled trials. Sleep Med. Rev. 2017, 31, 70–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murillo-Rodriguez, E.; Arias-Carrion, O.; Zavala-Garcia, A.; Sarro-Ramirez, A.; Huitron-Resendiz, S.; Arankowsky-Sandoval, G. Basic sleep mechanisms: An integrative review. Cent. Nerv. Syst. Agents Med. Chem. 2012, 12, 38–54. [Google Scholar] [CrossRef]
- Tosini, G.; Ferguson, I.; Tsubota, K. Effects of blue light on the circadian system and eye physiology. Mol. Vis. 2016, 22, 61–72. [Google Scholar]
- Wesensten, N.J.; Belenky, G.; Kautz, M.A.; Thorne, D.R.; Reichardt, R.M.; Balkin, T.J. Maintaining alertness and performance during sleep deprivation: Modafinil versus caffeine. Psychopharmacology 2001, 159, 238–247. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Controls | Athletes | p-Value |
|---|---|---|---|
| Number | 20 | 16 | |
| Gender | Male | Male | |
| Age | 21.25 ± 2.15 | 22.44 ± 2.13 | 0.107 |
| Education | 14.90 ± 1.77 | 15.88 ± 1.89 | 0.121 |
| Habitual sleep time | 7.27 ± 0.74 | 7.03 ± 1.02 | 0.421 |
| Physical activity | <200 min of physical activity per week | >10 h training time per week |
| BS | SD | |||
|---|---|---|---|---|
| Controls | Athletes | Controls | Athletes | |
| SSRT (ms) | 231.806 ± 56.433 | 187.717 ± 49.138 | 275.732 ± 61.761 | 224.755 ± 70.910 |
| Stop-ACC (%) | 57.672 ± 16.027 | 65.452 ± 7.073 | 52.349 ± 18.465 | 63.531 ± 11.468 |
| Go-RT (ms) | 628.795 ± 160.636 | 694.308 ± 92.807 | 619.496 ± 173.676 | 711.154 ± 91.855 |
| Go-ACC (%) | 89.387 ± 14.360 | 91.639 ± 6.329 | 85.797 ± 14.247 | 90.543 ± 8.399 |
| BS | SD | |||
|---|---|---|---|---|
| Controls | Athletes | Controls | Athletes | |
| N2 (μV) | −4.353 ± 2.119 | −1.648 ± 2.235 | −2.481 ± 3.729 | −1.668 ± 0.834 |
| ERN (μV) | −4.447 ± 4.238 | −1.332 ± 2.184 | −1.634 ± 3.262 | −1.698 ± 2.328 |
| P3 (μV) | 6.745 ± 2.868 | 4.448 ± 2.164 | 4.231 ± 2.859 | 2.822 ± 1.758 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Song, T.; Peng, Z.; Dai, C.; Wang, L.; Shao, Y.; Wang, L.; Weng, X.; Han, M. Acute Sleep Deprivation Impairs Motor Inhibition in Table Tennis Athletes: An ERP Study. Brain Sci. 2022, 12, 746. https://doi.org/10.3390/brainsci12060746
Xu L, Song T, Peng Z, Dai C, Wang L, Shao Y, Wang L, Weng X, Han M. Acute Sleep Deprivation Impairs Motor Inhibition in Table Tennis Athletes: An ERP Study. Brain Sciences. 2022; 12(6):746. https://doi.org/10.3390/brainsci12060746
Chicago/Turabian StyleXu, Lin, Tao Song, Ziyi Peng, Cimin Dai, Letong Wang, Yongcong Shao, Lanxiang Wang, Xiechuan Weng, and Mengfei Han. 2022. "Acute Sleep Deprivation Impairs Motor Inhibition in Table Tennis Athletes: An ERP Study" Brain Sciences 12, no. 6: 746. https://doi.org/10.3390/brainsci12060746