
Functional Connectivity at Rest between the Human Medial Posterior Parietal Cortex and the Primary Motor Cortex Detected by Paired-Pulse Transcranial Magnetic Stimulation

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Localization of Brain Sites
2.3. Transcranial Magnetic Stimulation
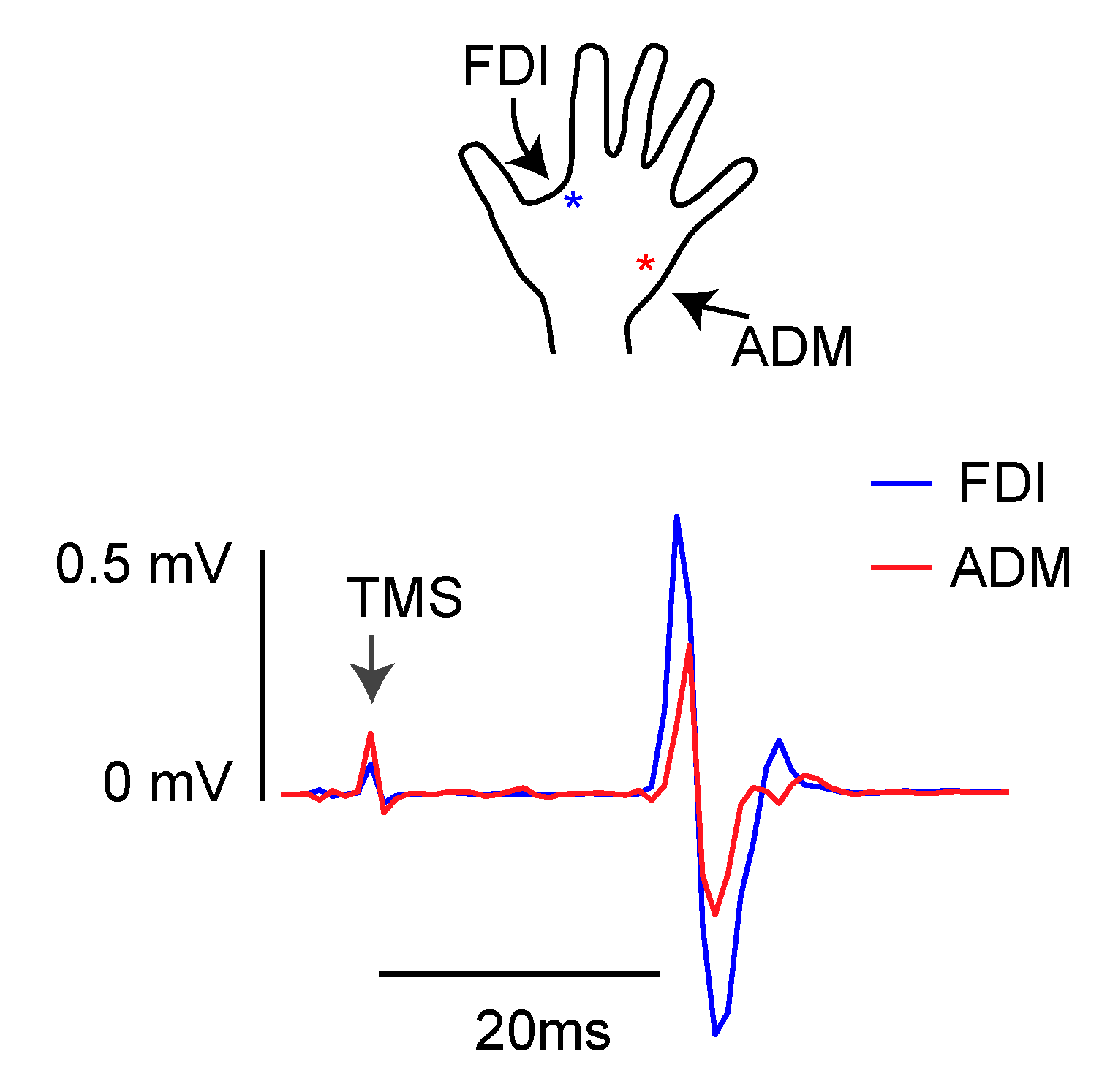
2.4. Electromyographic Recordings
2.5. Data Analysis
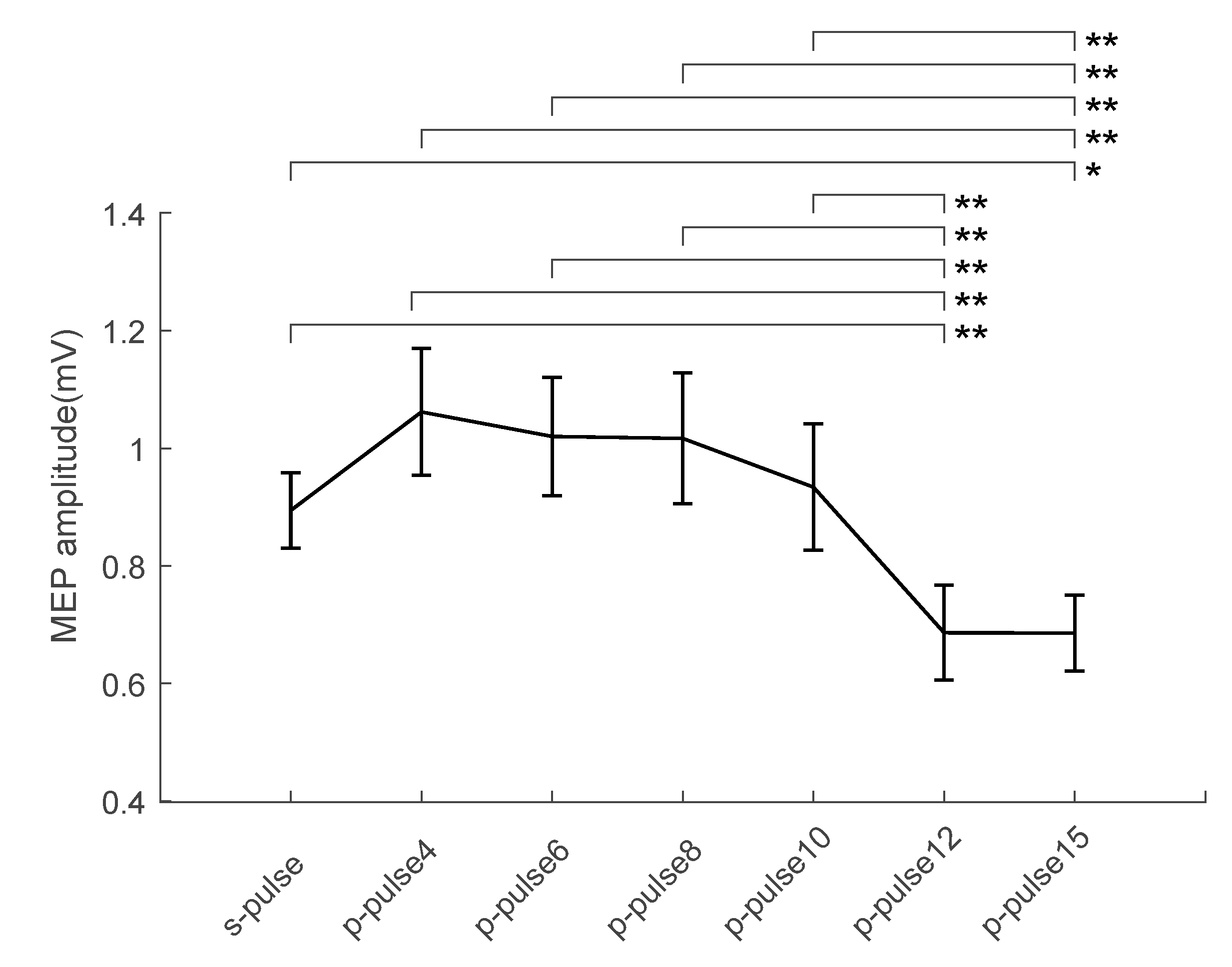
3. Results
4. Discussion
Comparisons with Other Studies about Parietal Resting Connectivity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galletti, C.; Kutz, D.F.; Gamberini, M.; Breveglieri, R.; Fattori, P. Role of the medial parieto-occipital cortex in the control of reaching and grasping movements. Exp. Brain Res. 2003, 153, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Fattori, P.; Breveglieri, R.; Bosco, A.; Gamberini, M.; Galletti, C. Vision for Prehension in the Medial Parietal Cortex. Cereb. Cortex 2017, 27, 1149–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, P.; Scherberger, H. Visual Guidance in Control of Grasping. Annu. Rev. Neurosci. 2015, 38, 69–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonini, L.; Serventi, F.U.; Bruni, S.; Maranesi, M.; Bimbi, M.; Simone, L.; Rozzi, S.; Ferrari, P.F.; Fogassil, L. Selectivity for grip type and action goal in macaque inferior parietal and ventral premotor grasping neurons. J. Neurophysiol. 2012, 108, 1607–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzolatti, G.; Matelli, M. Two different streams form the dorsal visual system: Anatomy and functions. Exp. Brain Res. 2003, 153, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Pesaran, B.; Nelson, M.J.; Andersen, R.A. Dorsal Premotor Neurons Encode the Relative Position of the Hand, Eye, and Goal during Reach Planning. Neuron 2006, 51, 125–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyder, L.H.; Batista, A.P.; Andersen, R.A. Intention-related activity in the posterior parietal cortex: A review. Vis. Res. 2000, 40, 1433–1441. [Google Scholar] [CrossRef] [Green Version]
- Tosoni, A.; Pitzalis, S.; Committeri, G.; Fattori, P.; Galletti, C.; Galati, G. Resting-state connectivity and functional specialization in human medial parieto-occipital cortex. Brain Struct. Funct. 2015, 220, 3307–3321. [Google Scholar] [CrossRef]
- Culham, J.C.; Gallivan, J.; Cavina-Pratesi, C.; Quinlan, D.J. fMRI investigations of reaching and ego space in human superior parieto-occipital cortex. In Embodiment, Ego-Space and Action; Klatzky, R., McWhinney, B., Behrmann, M., Eds.; Press Taylor & Francis: New York, NY, USA, 2008; pp. 247–274. [Google Scholar]
- Cavina-Pratesi, C.; Monaco, S.; Fattori, P.; Galletti, C.; McAdam, T.D.; Quinlan, D.J.; Goodale, M.A.; Culham, J.C. Functional magnetic resonance imaging reveals the neural substrates of arm transport and grip formation in reach-to-grasp actions in humans. J. Neurosci. 2010, 30, 10306–10323. [Google Scholar] [CrossRef] [Green Version]
- Vesia, M.; Bolton, D.A.; Mochizuki, G.; Staines, W.R. Human parietal and primary motor cortical interactions are selectively modulated during the transport and grip formation of goal-directed hand actions. Neuropsychologia 2013, 51, 410–417. [Google Scholar] [CrossRef]
- Vesia, M.; Barnett-Cowan, M.; Elahi, B.; Jegatheeswaran, G.; Isayama, R.; Neva, J.L.; Davare, M.; Staines, W.R.; Culham, J.C.; Chen, R. Human dorsomedial parieto-motor circuit specifies grasp during the planning of goal-directed hand actions. Cortex 2017, 92, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.; Del Olmo, M.F.; Cheeran, B.; Ruge, D.; Schippling, S.; Caltagirone, C.; Rothwell, J.C. Focal stimulation of the posterior parietal cortex increases the excitability of the ipsilateral motor cortex. J. Neurosci. 2007, 27, 6815–6822. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.; Del Olmo, M.F.; Cheeran, B.; Ruge, D.; Schippling, S.; Caltagirone, C.; Rothwell, J.C. Functional interplay between posterior parietal and ipsilateral motor cortex revealed by twin-coil transcranial magnetic stimulation during reach planning toward contralateral space. J. Neurosci. 2008, 28, 5944–5953. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.; Cercignani, M.; Pecchioli, C.; Versace, V.; Oliveri, M.; Caltagirone, C.; Rothwell, J.; Bozzali, M. In vivo definition of parieto-motor connections involved in planning of grasping movements. Neuroimage 2010, 51, 300–312. [Google Scholar] [CrossRef]
- Davare, M.; Rothwell, J.C.; Lemon, R.N. Causal Connectivity between the Human Anterior Intraparietal Area and Premotor Cortex during Grasp. Curr. Biol. 2010, 20, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Hadjidimitrakis, K.; Bertozzi, F.; Breveglieri, R.; Bosco, A.; Galletti, C.; Fattori, P. Common neural substrate for processing depth and direction signals for reaching in the monkey medial posterior parietal cortex. Cereb. Cortex 2014, 24, 1645–1657. [Google Scholar] [CrossRef] [Green Version]
- Bosco, A.; Breveglieri, R.; Chinellato, E.; Galletti, C.; Fattori, P. Reaching activity in the medial posterior parietal cortex of monkeys is modulated by visual feedback. J. Neurosci. 2010, 30, 14773–14785. [Google Scholar] [CrossRef]
- Fattori, P.; Breveglieri, R.; Marzocchi, N.; Filippini, D.; Bosco, A.; Galletti, C. Hand orientation during reach-to-grasp movements modulates neuronal activity in the medial posterior parietal area V6A. J. Neurosci. 2009, 29, 1928–1936. [Google Scholar] [CrossRef]
- Breveglieri, R.; Bosco, A.; Galletti, C.; Passarelli, L.; Fattori, P. Neural activity in the medial parietal area V6A while grasping with or without visual feedback. Sci. Rep. 2016, 6, 28893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breveglieri, R.; de Vitis, M.; Bosco, A.; Galletti, C.; Fattori, P. Interplay between Grip and Vision in the Monkey Medial Parietal Lobe. Cereb. Cortex 2018, 28, 2028–2042. [Google Scholar] [CrossRef] [PubMed]
- Fattori, P.; Raos, V.; Breveglieri, R.; Bosco, A.; Marzocchi, N.; Galletti, C. The dorsomedial pathway is not just for reaching: Grasping neurons in the medial parieto-occipital cortex of the macaque monkey. J. Neurosci. 2010, 30, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Gamberini, M.; Passarelli, L.; Fattori, P.; Zucchelli, M.; Bakola, S.; Luppino, G.; Galletti, C. Cortical connections of the visuomotor parietooccipital area V6Ad of the macaque monkey. J. Comp. Neurol. 2009, 513, 622–642. [Google Scholar] [CrossRef]
- Passarelli, L.; Rosa, M.G.; Gamberini, M.; Bakola, S.; Burman, K.J.; Fattori, P.; Galletti, C. Cortical Connections of Area V6Av in the Macaque: A Visual-Input Node to the Eye/Hand Coordination System. J. Neurosci. 2011, 31, 1790–1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passarelli, L.; Rosa, M.G.; Bakola, S.; Gamberini, M.; Worthy, K.H.; Fattori, P.; Galletti, C. Uniformity and diversity of cortical projections to precuneate areas in the macaque monkey: What defines area PGm? Cereb. Cortex 2018, 28, 1700–1717. [Google Scholar] [CrossRef] [Green Version]
- Galletti, C.; Fattori, P. The dorsal visual stream revisited: Stable circuits or dynamic pathways? Cortex 2018, 98, 203–217. [Google Scholar] [CrossRef]
- Passarelli, L.; Gamberini, M.; Fattori, P. The superior parietal lobule of primates: A sensory-motor hub for interaction with the environment. J. Integr. Neurosci. 2021, 20, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Pitzalis, S.; Fattori, P.; Galletti, C. The human cortical areas V6 and V6A. Vis. Neurosci. 2015, 32, E007. [Google Scholar] [CrossRef] [PubMed]
- Makris, N.; Kennedy, D.N.; McInerney, S.; Sorensen, A.G.; Wang, R.; Caviness, V.S., Jr.; Pandya, D.N. Segmentation of subcomponents within the superior longitudinal fascicle in humans: A quantitative, in vivo, DT-MRI study. Cereb. Cortex 2005, 15, 854–869. [Google Scholar] [CrossRef] [PubMed]
- Parlatini, V.; Radua, J.; Dell’Acqua, F.; Leslie, A.; Simmons, A.; Murphy, D.G.; Catani, M.; de Schotten, M.T. Functional segregation and integration within fronto-parietal networks. Neuroimage 2017, 146, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Espay, A.J.; Gunraj, C.A.; Pal, P.K.; Cunic, D.I.; Lang, A.E.; Chen, R. Interhemispheric and ipsilateral connections in Parkinson’s disease: Relation to mirror movements. Mov. Disord. 2007, 22, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Sattler, V.; Dickler, M.; Michaud, M.; Meunier, S.; Simonetta-Moreau, M. Does abnormal interhemispheric inhibition play a role in mirror dystonia? Mov. Disord. 2014, 29, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Chiappini, E.; Borgomaneri, S.; Marangon, M.; Turrini, S.; Romei, V.; Avenanti, A. Driving associative plasticity in premotor-motor connections through a novel paired associative stimulation based on long-latency cortico-cortical interactions. Brain Stimul. 2020, 13, 1461–1463. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, L.; Barchiesi, G. Transcranial magnetic mapping of the short-latency modulations of corticospinal activity from the ipsilateral hemisphere during rest. Front. Neural Circuits 2011, 5, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Civardi, C.; Cantello, R.; Asselman, P.; Rothwell, J.C. Transcranial magnetic stimulation can be used to test connections to primary motor areas from frontal and medial cortex in humans. Neuroimage 2001, 14, 1444–1453. [Google Scholar] [CrossRef]
- di Lazzaro, V.V.; Oliviero, A.; Profice, P.; Insola, A.; Mazzone, P.; Tonali, P.; Rothwell, J.C. Direct demonstration of interhemispheric inhibition of the human motor cortex produced by transcranial magnetic stimulation. Exp. Brain Res. 1999, 124, 520–524. [Google Scholar] [CrossRef]
- Ferbert, A.; Priori, A.; Rothwell, J.C.; Day, B.L.; Colebatch, J.G.; Marsden, C.D. Interhemispheric inhibition of the human motor cortex. J. Physiol. 1992, 453, 525–546. [Google Scholar] [CrossRef]
- Rothwell, J.C. Using transcranial magnetic stimulation methods to probe connectivity between motor areas of the brain. Hum. Mov. Sci. 2011, 30, 906–915. [Google Scholar] [CrossRef] [PubMed]
- Fiori, F.; Chiappini, E.; Candidi, M.; Romei, V.; Borgomaneri, S.; Avenanti, A. Long-latency interhemispheric interactions between motor-related areas and the primary motor cortex: A dual site TMS study. Sci. Rep. 2017, 7, 14936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiori, F.; Chiappini, E.; Soriano, M.; Paracampo, R.; Romei, V.; Borgomaneri, S.; Avenanti, A. Long-latency modulation of motor cortex excitability by ipsilateral posterior inferior frontal gyrus and pre-supplementary motor area. Sci. Rep. 2016, 6, 38396. [Google Scholar] [CrossRef] [Green Version]
- Karabanov, A.N.; Chao, C.C.; Paine, R.; Hallett, M. Mapping different intra-hemispheric parietal-motor networks using twin coil TMS. Brain Stimul. 2013, 6, 384–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattaneo, L.; Giampiccolo, D.; Meneghelli, P.; Tramontano, V.; Sala, F. Cortico-cortical connectivity between the superior and inferior parietal lobules and the motor cortex assessed by intraoperative dual cortical stimulation. Brain Stimul. 2020, 13, 819–831. [Google Scholar] [CrossRef] [Green Version]
- Ziluk, A.; Premji, A.; Nelson, A.J. Functional connectivity from area 5 to primary motor cortex via paired-pulse transcranial magnetic stimulation. Neurosci. Lett. 2010, 484, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Lang, A.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Briggs, G.G.; Nebes, R.D. Patterns of Hand Preference in a Student Population. Cortex 1975, 11, 230–238. [Google Scholar] [CrossRef] [Green Version]
- Avenanti, A.; Paracampo, R.; Annella, L.; Tidoni, E.; Aglioti, S.M. Boosting and decreasing action prediction abilities through excitatory and inhibitory tDCS of inferior frontal cortex. Cereb. Cortex 2018, 28, 1282–1296. [Google Scholar] [CrossRef] [Green Version]
- Paracampo, R.; Pirruccio, M.; Costa, M.; Borgomaneri, S.; Avenanti, A. Visual, sensorimotor and cognitive routes to understanding others’ enjoyment: An individual differences rTMS approach to empathic accuracy. Neuropsychologia 2018, 116, 86–98. [Google Scholar] [CrossRef]
- Carducci, F.; Brusco, R. Accuracy of an individualized MR-based head model for navigated brain stimulation. Psychiatry Res. Neuroimaging 2012, 203, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Talairach, J.; Tournoux, P. Co-Planar Stereotaxic Atlas of the Human Brain: 3-Dimensional Proportional System: An Approach to Cerebral Imaging; Thieme Medical Publishers: Stuttgart, Germany; New York, NY, USA, 1988. [Google Scholar]
- Ciavarro, M.; Ambrosini, E.; Tosoni, A.; Committeri, G.; Fattori, P.; Galletti, C. rTMS of Medial Parieto-occipital Cortex Interferes with Attentional Reorienting during Attention and Reaching Tasks. J. Cogn. Neurosci. 2013, 25, 1453–1462. [Google Scholar] [CrossRef]
- Breveglieri, R.; Bosco, A.; Borgomaneri, S.; Tessari, A.; Galletti, C.; Avenanti, A.; Fattori, P. Transcranial Magnetic Stimulation Over the Human Medial Posterior Parietal Cortex Disrupts Depth Encoding During Reach Planning. Cereb. Cortex 2021, 31, 267–280. [Google Scholar] [CrossRef] [PubMed]
- Vesia, M.; Prime, S.L.; Yan, X.; Sergio, L.E.; Crawford, J.D. Specificity of human parietal saccade and reach regions during transcranial magnetic stimulation. J. Neurosci. 2010, 30, 13053–13065. [Google Scholar] [CrossRef] [Green Version]
- Gallivan, J.P.; McLean, A.; Culham, J.C. Neuroimaging reveals enhanced activation in a reach-selective brain area for objects located within participants’ typical hand workspaces. Neuropsychologia 2011, 49, 3710–3721. [Google Scholar] [CrossRef] [PubMed]
- Pitzalis, S.; Sereno, M.I.; Committeri, G.; Fattori, P.; Galati, G.; Tosoni, A.; Galletti, C. The human homologue of macaque area V6A. Neuroimage 2013, 82, 517–530. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A.; Safety of TMS Consensus Group. Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [Green Version]
- Rossini, P.M.; Burke, D.; Chen, R.; Cohen, L.G.; Daskalakis, Z.; di Iorio, R.; di Lazzaro, V.; Ferreri, F.; Fitzgerald, P.B.; George, M.S.; et al. Non-invasive electrical and magnetic stimulation of the brain, spinal cord, roots and peripheral nerves: Basic principles and procedures for routine clinical and research application. An updated report from an I.F.C.N. Committee. Clin. Neurophysiol. 2015, 126, 1071–1107. [Google Scholar] [CrossRef] [PubMed]
- Cardellicchio, P.; Dolfini, E.; Fadiga, L.; D’Ausilio, A. Parallel fast and slow motor inhibition processes in Joint Action coordination. Cortex 2020, 133, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Tomassini, V.; Jbabdi, S.; Klein, J.C.; Behrens, T.E.J.; Pozzilli, C.; Matthews, P.M.; Rushworth, M.F.S.; Johansen-Berg, H. Diffusion-weighted imaging tractography-based parcellation of the human lateral premotor cortex identifies dorsal and ventral subregions with anatomical and functional specializations. J. Neurosci. 2007, 27, 10259–10269. [Google Scholar] [CrossRef] [PubMed]
- Strick, P.L.; Kim, C.C. Input to primate motor cortex from posterior parietal cortex (area 5). I. Demonstration by retrograde transport. Brain Res. 1978, 157, 325–330. [Google Scholar] [CrossRef]
- Zarzecki, P.; Strick, P.L. Input to primate motor cortex from posterior parietal cortex (area 5). II. Identification by antidromic activation. Brain Res. 1978, 157, 331–335. [Google Scholar] [CrossRef]
- Bruni, S.; Gerbella, M.; Bonini, L.; Borra, E.; Coudé, G.; Ferrari, P.F.; Fogassi, L.; Maranesi, M.; Rodà, F.; Simone, L.; et al. Cortical and subcortical connections of parietal and premotor nodes of the monkey hand mirror neuron network. Brain Struct. Funct. 2018, 223, 1713–1729. [Google Scholar] [CrossRef] [PubMed]
- Rathelot, J.A.; Dum, R.P.; Strick, P.L. Posterior parietal cortex contains a command apparatus for hand movements. Proc. Natl. Acad. Sci. USA 2017, 114, 4255–4260. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; Lisanby, S.; Peterchev, A. Electric field depth–focality tradeoff in transcranial magnetic stimulation: Simulation comparison of 50 coil designs. Brain Stimul. 2013, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheperjans, F.; Eickhoff, S.B.; Hömke, L.; Mohlberg, H.; Hermann, K.; Amunts, K.; Zilles, K. Probabilistic maps, morphometry, and variability of cytoarchitectonic areas in the human superior parietal cortex. Cereb. Cortex 2008, 18, 2141–2157. [Google Scholar] [CrossRef] [PubMed]
- Karabanov, A.; Jin, S.H.; Joutsen, A.; Poston, B.; Aizen, J.; Ellenstein, A.; Hallett, M. Timing-dependent modulation of the posterior parietal cortex-primary motor cortex pathway by sensorimotor training. J. Neurophysiol. 2012, 107, 3190–3199. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| TMS | Mean | Standard Error |
|---|---|---|
| s-pulse | 0.89 | 0.06 |
| p-pulse4 | 1.06 | 0.10 |
| p-pulse6 | 1.02 | 0.10 |
| p-pulse8 | 1.02 | 0.11 |
| p-pulse10 | 0.93 | 0.11 |
| p-pulse12 | 0.69 | 0.08 |
| p-pulse15 | 0.69 | 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Breveglieri, R.; Borgomaneri, S.; Filippini, M.; De Vitis, M.; Tessari, A.; Fattori, P. Functional Connectivity at Rest between the Human Medial Posterior Parietal Cortex and the Primary Motor Cortex Detected by Paired-Pulse Transcranial Magnetic Stimulation. Brain Sci. 2021, 11, 1357. https://doi.org/10.3390/brainsci11101357
Breveglieri R, Borgomaneri S, Filippini M, De Vitis M, Tessari A, Fattori P. Functional Connectivity at Rest between the Human Medial Posterior Parietal Cortex and the Primary Motor Cortex Detected by Paired-Pulse Transcranial Magnetic Stimulation. Brain Sciences. 2021; 11(10):1357. https://doi.org/10.3390/brainsci11101357
Chicago/Turabian StyleBreveglieri, Rossella, Sara Borgomaneri, Matteo Filippini, Marina De Vitis, Alessia Tessari, and Patrizia Fattori. 2021. "Functional Connectivity at Rest between the Human Medial Posterior Parietal Cortex and the Primary Motor Cortex Detected by Paired-Pulse Transcranial Magnetic Stimulation" Brain Sciences 11, no. 10: 1357. https://doi.org/10.3390/brainsci11101357