
Vestibular Stimulation May Drive Multisensory Processing: Principles for Targeted Sensorimotor Therapy (TSMT)


 ,
, 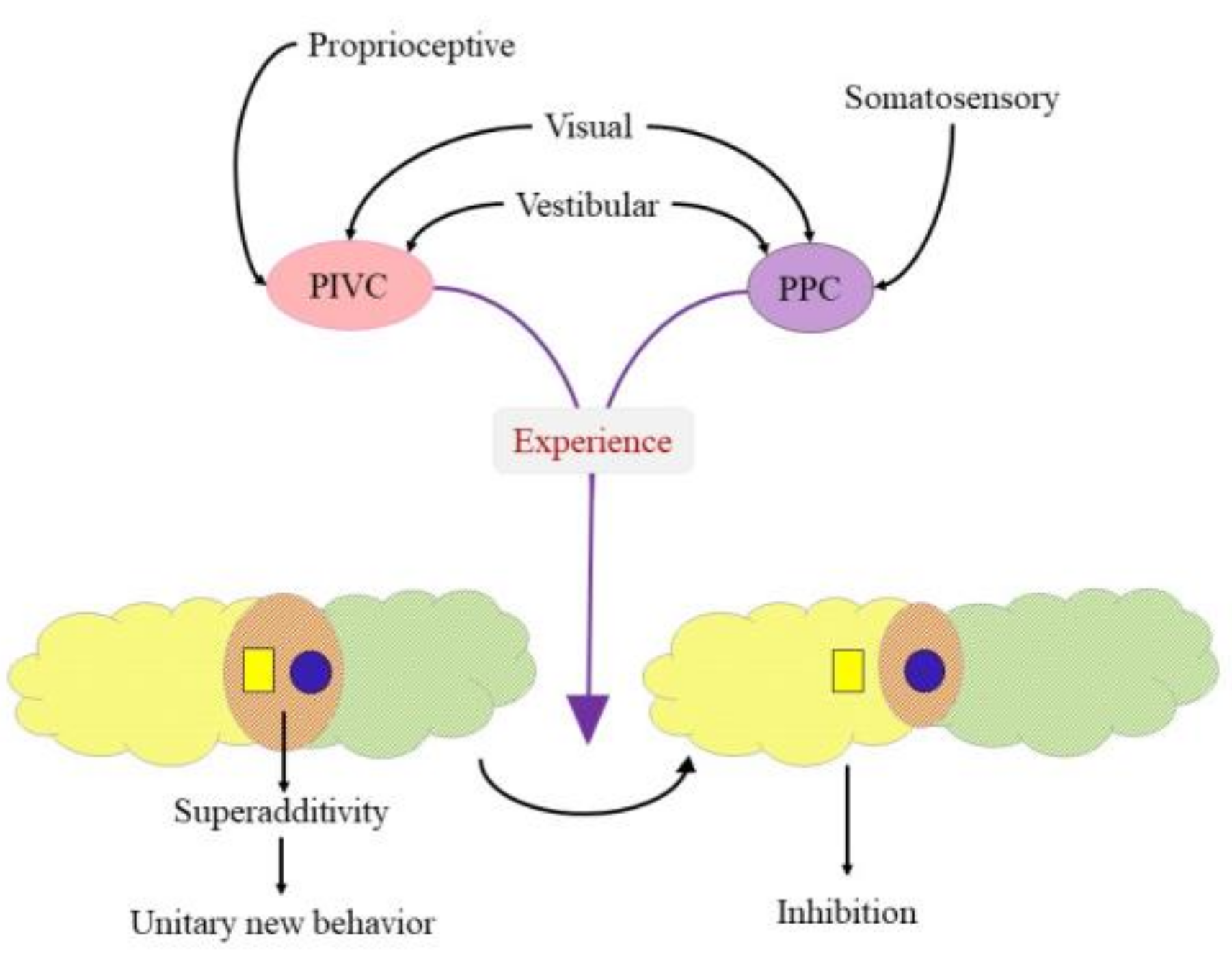{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
Development of Multisensory Integration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Gori, M. Multisensory Integration and Calibration in Children and Adults with and without Sensory and Motor Disabilities. Multisens. Res. 2015, 28, 71–99. [Google Scholar] [CrossRef]
- Chandrasekaran, C. Computational principles and models of multisensory integration. Curr. Opin. Neurobiol. 2017, 43, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purpura, G.; Perazza, S.; Cioni, G.; Tinelli, F. Visuo-haptic transfer for object recognition in children with periventricular leukomalacia and bilateral cerebral palsy. Child Neuropsychol. 2019, 25, 1084–1097. [Google Scholar] [CrossRef]
- Pavao, S.L.; Rocha, N.A.C.F. Sensory processing disorders in children with cerebral palsy. Infant. Behav. Dev. 2017, 46, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Jarvinen, A.; Ng, R.; Crivelli, D.; Arnold, A.J.; Woo-VonHoogenstyn, N.; Bellugi, U. Relations between social-perceptual ability in multi- and unisensory contexts, autonomic reactivity, and social functioning in individuals with Williams syndrome. Neuropsychologia 2015, 73, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.Y.; Cai, X.L.; Weigl, M.; Bang, P.; Cheung, E.F.C.; Chan, R.C.K. Multisensory temporal binding window in autism spectrum disorders and schizophrenia spectrum disorders: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2018, 86, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, R.A.; Baum, S.H.; Segers, M.; Ferber, S.; Barense, M.D.; Wallace, M.T. Multisensory speech perception in autism spectrum disorder: From phoneme to whole-word perception. Autism Res. 2017, 10, 1280–1290. [Google Scholar] [CrossRef]
- Feldman, J.I.; Dunham, K.; Cassidy, M.; Wallace, M.T.; Liu, Y.; Woynaroski, T.G. Audiovisual multisensory integration in individuals with autism spectrum disorder: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2018, 95, 220–234. [Google Scholar] [CrossRef]
- Lane, S.J.; Reynolds, S. Sensory Over-Responsivity as an Added Dimension in ADHD. Front. Integr. Neurosci. 2019, 13, 40. [Google Scholar] [CrossRef]
- Myers, M.H.; Iannaccone, A.; Bidelman, G.M. A pilot investigation of audiovisual processing and multisensory integration in patients with inherited retinal dystrophies. BMC Ophthalmol. 2017, 17, 240. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.S.; Owen, J.P.; Desai, S.S.; Hill, S.S.; Arnett, A.B.; Harris, J.; Marco, E.J.; Mukherjee, P. Autism and sensory processing disorders: Shared white matter disruption in sensory pathways but divergent connectivity in social-emotional pathways. PLoS ONE 2014, 9, e103038. [Google Scholar]
- Kumar, G.V.; Halder, T.; Jaiswal, A.K.; Mukherjee, A.; Roy, D.; Banerjee, A. Large Scale Functional Brain Networks Underlying Temporal Integration of Audio-Visual Speech Perception: An EEG Study. Front. Psychol. 2016, 7, 1558. [Google Scholar] [CrossRef] [PubMed]
- Tiippana, K.; Möttönen, R.; Schwartz, J.L. Multisensory and sensorimotor interactions in speech perception. Front. Psychol. 2015, 6, 458. [Google Scholar] [CrossRef] [Green Version]
- Hahn, N.; Foxe, J.J.; Molholm, S. Impairments of multisensory integration and cross-sensory learning as pathways to dyslexia. Neurosci. Biobehav. Rev. 2014, 47, 384–392. [Google Scholar] [CrossRef] [Green Version]
- Goddard, S. The role of primitive survival reflexes in the development of the visual system. Mind Moves Inst. 1995, 6, 31–35. [Google Scholar]
- Le Gall, A.; Hilber, P.; Chesneau, C.; Bulla, J.; Toulouse, J.; Machado, M.L.; Philoxene, B.; Smith, P.F.; Besnard, S. The critical role of vestibular graviception during cognitivo-motor development. Behav. Brain Res. 2019, 372, 112040. [Google Scholar] [CrossRef]
- Wiener-Vacher, S.R.; Hamilton, D.A.; Wiener, S.I. Vestibular activity and cognitive development in children: Perspectives. Front. Integr. Neurosci. 2013, 7, 92. [Google Scholar] [CrossRef] [Green Version]
- Cullen, K.E. The vestibular system: Multimodal integration and encoding of self-motion for motor control. Trends Neurosci. 2012, 35, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Assaiante, C. Development of locomotor balance control in healthy children. Neurosci. Biobehav. Rev. 1998, 22, 527–532. [Google Scholar] [CrossRef]
- Green, A.M.; Angelaki, D.E. Multisensory integration: Resolving sensory ambiguities to build novel representations. Curr. Opin. Neurobiol. 2010, 20, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Cuturi, L.F.; Gori, M. Biases in the Visual and Haptic Subjective Vertical Reveal the Role of Proprioceptive/Vestibular Priors in Child Development. Front. Neurol. 2019, 9, 1151. [Google Scholar] [CrossRef]
- Gonzalez, S.R.; Ciuffreda, K.J.; Hernández, L.C.; Escalante, J.B. The Correlation between Primitive Reflexes and Saccadic Eye Movements in 5th Grade Children with Teacher-Reported Reading Problems. Optom. Vis. Dev. 2008, 39, 140–145. [Google Scholar]
- Blythe, S.G. Releasing educational potential through movement: A summary of individual studies carried out using the INPP test battery and developmental exercise programme for use in schools with children with special needs. Child Care Pract. 2005, 11, 415–432. [Google Scholar] [CrossRef]
- Rochat, P. Self-Unity as Ground Zero of Learning and Development. Front. Psychol. 2019, 10, 414. [Google Scholar] [CrossRef] [PubMed]
- Deroualle, D.; Lopez, C. Toward a vestibular contribution to social cognition. Front. Integr. Neurosci. 2014, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Zsuga, J.; Biro, K.; Papp, C.; Tajti, G.; Gesztelyi, R. The proactive model of learning: Integrative framework for model-free and model-based reinforcement learning utilizing the associative learning-based proactive brain concept. Behav. Neurosci. 2016, 130, 6. [Google Scholar] [CrossRef] [PubMed]
- Zsuga, J.; Biro, K.; Tajti, G.; Szilasi, M.E.; Papp, C.; Juhasz, B.; Gesztelyi, R. ‘Proactive’ use of cue-context congruence for building reinforcement learning’s reward function. BMC Neurosci. 2016, 17, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, M.T.; Stein, B.E. Early experience determines how the senses will interact. J. Neurophysiol. 2007, 97, 921–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shams, L.; Seitz, A.R. Benefits of multisensory learning. Trends Cogn. Sci. 2008, 12, 411–417. [Google Scholar] [CrossRef]
- Bair, W.N.; Kiemel, T.; Jeka, J.J.; Clark, J.E. Development of multisensory reweighting is impaired for quiet stance control in children with developmental coordination disorder (DCD). PLoS ONE 2012, 7, e40932. [Google Scholar] [CrossRef] [Green Version]
- Gutfreund, Y.; Zheng, W.; Knudsen, E.I. Gated visual input to the central auditory system. Science 2002, 297, 1556–1559. [Google Scholar] [CrossRef]
- Sabes, P.N. Sensory integration for reaching: Models of optimality in the context of behavior and the underlying neural circuits. Prog. Brain Res. 2011, 191, 195–209. [Google Scholar]
- Peterka, R.J.; Loughlin, P.J. Dynamic regulation of sensorimotor integration in human postural control. J. Neurophysiol. 2004, 91, 410–423. [Google Scholar] [CrossRef] [Green Version]
- Alais, D.; Burr, D. The ventriloquist effect results from near-optimal bimodal integration. Curr. Biol. 2004, 14, 257–262. [Google Scholar] [CrossRef]
- Kording, K.P.; Wolpert, D.M. Bayesian integration in sensorimotor learning. Nature 2004, 427, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Kording, K.P.; Beierholm, U.; Ma, W.J.; Quartz, S.; Tenenbaum, J.B.; Shams, L. Causal inference in multisensory perception. PLoS ONE 2007, 2, e943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, C.; Sokhey, T.; Gaebler-Spira, D.; Kording, K.P. The development of Bayesian integration in sensorimotor estimation. J. Vis. 2018, 18, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldman, H.; Friston, K.J. Attention, uncertainty, and free-energy. Front. Hum. Neurosci. 2010, 4, 215. [Google Scholar] [CrossRef] [Green Version]
- Friston, K. The free-energy principle: A unified brain theory? Nat. Rev. Neurosci. 2010, 11, 127–138. [Google Scholar] [CrossRef]
- Avillac, M.; Ben Hamed, S.; Duhamel, J.R. Multisensory integration in the ventral intraparietal area of the macaque monkey. J. Neurosci. 2007, 27, 1922–1932. [Google Scholar] [CrossRef] [Green Version]
- Stanford, T.R.; Stein, B.E. Superadditivity in multisensory integration: Putting the computation in context. Neuroreport 2007, 18, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Harrar, V.; Harris, L.R.; Spence, C. Multisensory integration is independent of perceived simultaneity. Exp. Brain Res. 2017, 235, 763–775. [Google Scholar] [CrossRef] [PubMed]
- ten Oever, S.; Schroeder, C.E.; Poeppel, D.; van Atteveldt, N.; Zion-Golumbic, E. Rhythmicity and cross-modal temporal cues facilitate detection. Neuropsychologia 2014, 63, 43–50. [Google Scholar] [CrossRef]
- Bahrick, L.E.; Lickliter, R. Intersensory redundancy guides attentional selectivity and perceptual learning in infancy. Dev. Psychol. 2000, 36, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Brown, V.A.; Strand, J.F. “Paying” attention to audiovisual speech: Do incongruent stimuli incur greater costs? Atten. Percept. Psychophys. 2019, 81, 1743–1756. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.C.; Jeka, J.; Clark, J.E. Development of adaptive sensorimotor control in infant sitting posture. Gait Posture 2016, 45, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Nardini, M.; Jones, P.; Bedford, R.; Braddick, O. Development of cue integration in human navigation. Curr. Biol. 2008, 18, 689–693. [Google Scholar] [CrossRef] [Green Version]
- Gori, M.; Del Viva, M.; Sandini, G.; Burr, D.C. Young children do not integrate visual and haptic form information. Curr. Biol. 2008, 18, 694–698. [Google Scholar] [CrossRef] [Green Version]
- Stein, B.E.; Stanford, T.R. Multisensory integration: Current issues from the perspective of the single neuron. Nat. Rev. Neurosci. 2008, 9, 255–266. [Google Scholar] [CrossRef]
- Stevenson, R.A.; Geoghegan, M.L.; James, T.W. Superadditive BOLD activation in superior temporal sulcus with threshold non-speech objects. Exp. Brain Res. 2007, 179, 85–95. [Google Scholar] [CrossRef]
- Frank, S.M.; Greenlee, M.W. The parieto-insular vestibular cortex in humans: More than a single area? J. Neurophysiol. 2018, 120, 1438–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, L.K.; Meredith, M.A.; Stein, B.E. The role of anterior ectosylvian cortex in cross-modality orientation and approach behavior. Exp. Brain Res. 1996, 112, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rowland, B.A.; Jiang, W.; Stein, B.E. Brief cortical deactivation early in life has long-lasting effects on multisensory behavior. J. Neurosci. 2014, 34, 7198–7202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Xu, J.; Rowland, B.A.; Stein, B.E. Multisensory Plasticity in Superior Colliculus Neurons is Mediated by Association Cortex. Cereb. Cortex 2016, 26, 1130–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Yu, L.; Rowland, B.A.; Stanford, T.R.; Stein, B.E. Incorporating cross-modal statistics in the development and maintenance of multisensory integration. J. Neurosci. 2012, 32, 2287–2298. [Google Scholar] [CrossRef] [Green Version]
- Grusser, O.J.; Pause, M.; Schreiter, U. Localization and responses of neurones in the parieto-insular vestibular cortex of awake monkeys (Macaca fascicularis). J. Physiol. 1990, 430, 537–557. [Google Scholar] [CrossRef] [Green Version]
- Gunny, R.; Yousry, T.A. Imaging anatomy of the vestibular and visual systems. Curr. Opin. Neurol. 2007, 20, 3–11. [Google Scholar] [CrossRef]
- Ventre-Dominey, J. Vestibular function in the temporal and parietal cortex: Distinct velocity and inertial processing pathways. Front. Integr. Neurosci. 2014, 8, 53. [Google Scholar] [CrossRef] [Green Version]
- Klam, F.; Graf, W. Vestibular signals of posterior parietal cortex neurons during active and passive head movements in macaque monkeys. Ann. N. Y. Acad. Sci. 2003, 1004, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Fries, W. Cortical projections to the superior colliculus in the macaque monkey: A retrograde study using horseradish peroxidase. J. Comp. Neurol. 1984, 230, 55–76. [Google Scholar] [CrossRef]
- Ryckman, J.; Hilton, C.; Rogers, C.; Pineda, R. Sensory processing disorder in preterm infants during early childhood and relationships to early neurobehavior. Early Hum. Dev. 2017, 113, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Pekcetin, S.; Akı, E.; Üstünyurt, Z.; Kayıhan, H. The Efficiency of Sensory Integration Interventions in Preterm Infants. Percept. Mot. Ski. 2016, 123, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Rahkonen, P.; Lano, A.; Pesonen, A.K.; Heinonen, K.; Räikkönen, K.; Vanhatalo, S.; Autti, T.; Valanne, L.; Andersson, S.; Metsäranta, M. Atypical sensory processing is common in extremely low gestational age children. Acta Paediatr. 2015, 104, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, M.; Marom, R.; Berger, I.; Ben Bashat, D.; Gross-Tsur, V.; Ben-Sira, L.; Artzi, M.; Uliel, S.; Leitner, Y.; Geva, R. Neonatal neuropsychology: Emerging relations of neonatal sensory-motor responses to white matter integrity. Neuropsychologia 2014, 62, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Groh, J.M.; Sparks, D.L. Saccades to somatosensory targets. III. eye-position-dependent somatosensory activity in primate superior colliculus. J. Neurophysiol. 1996, 75, 439–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartline, P.H.; Vimal, R.L.; King, A.J.; Kurylo, D.D.; Northmore, D.P. Effects of eye position on auditory localization and neural representation of space in superior colliculus of cats. Exp. Brain Res. 1995, 104, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Stricanne, B.; Andersen, R.A.; Mazzoni, P. Eye-centered, head-centered, and intermediate coding of remembered sound locations in area LIP. J. Neurophysiol. 1996, 76, 2071–2076. [Google Scholar] [CrossRef]
- Pouget, A.; Deneve, S.; Duhamel, J.R. A computational perspective on the neural basis of multisensory spatial representations. Nat. Rev. Neurosci. 2002, 3, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Snyder, L.H. Frame-up. Focus on “eye-centered, head-centered, and complex coding of visual and auditory targets in the intraparietal sulcus”. J. Neurophysiol. 2005, 94, 2259–2260. [Google Scholar] [CrossRef]
- Chen, X.; Deangelis, G.C.; Angelaki, D.E. Diverse spatial reference frames of vestibular signals in parietal cortex. Neuron 2013, 80, 1310–1321. [Google Scholar] [CrossRef] [Green Version]
- Brooks, J.X.; Cullen, K.E. Multimodal integration in rostral fastigial nucleus provides an estimate of body movement. J. Neurosci. 2009, 29, 10499–10511. [Google Scholar] [CrossRef] [PubMed]
- A TSMT és HRG terapeuta moduláris képzési rendszeréről. Available online: https://bhrgmodel.org/eng/tsmt_modules (accessed on 14 July 2021).
- Institutional website. Available online: https://bhrg.hu/ (accessed on 14 July 2021).
- Bodor, E. Az Organikus Érési Zavar Felismerése és Kezelése TSMT-I Terápiával (Early Diagnosis and Therapy of Organic Developmental Delay Using TSMT-I Therapy). Bachelor’s Thesis, University of Miskolc, Miskolc, Hungary, 2014. [Google Scholar]
- Lakatos, K. A Mozgásérettség Vizsgálatának Jelentősége a Tanulási Zavarok Korai Felismerésében (Significance of Motor Development in Early Diagnosis of Learning Difficulties). Ph.D. Thesis, Semmelweis University, Budapest, Hungary, 2005. [Google Scholar]
- Lakatos, K. Az állapot és mozgásvizsgáló teszt (Condition and Movement Tests). Budapest, Hungary, 2006; ISBN 9630395479. [Google Scholar]
- Heri, B. Az Idegrendszeri Éretlenség Gyakoriságának Vizsgálata Viselkedés- és Magatartászavaros Gyerekek Körében (Assessment of CNS Maturity in Children with Behavioral Disabilities). MSc. Thesis, University of Debrecen, Debrecen, Hungary, 2017. [Google Scholar]
- Fenyosi, F. Socio-Economic Status and Human Capital Formation. MSc. Thesis, Wageningen University & Research, Wageningen, The Netherlands, 2014. [Google Scholar]
- Gurvich, C.; Maller, J.J.; Lithgow, B.; Haghgooie, S.; Kulkarni, J. Vestibular insights into cognition and psychiatry. Brain Res. 2013, 1537, 244–259. [Google Scholar] [CrossRef] [Green Version]
- Law, M.; Darrah, J.; Pollock, N.; Rosenbaum, P.; Russell, D.; Walter, S.D.; Petrenchik, T.; Wilson, B.; Wright, V. Focus on function - a randomized controlled trial comparing two rehabilitation interventions for young children with cerebral palsy. BMC Pediatr. 2007, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo-Dougovito, A.M. The role of dynamic systems theory in motor development research: How does theory inform practice and what are the potential implications for autism spectrum disorder? Int. J. Disabil. Hum. Dev. 2017, 16, 141–155. [Google Scholar] [CrossRef] [Green Version]
- Golenia, L.; Schoemaker, M.M.; Otten, E.; Mouton, L.J.; Bongers, R.M. What the dynamic systems approach can offer for understanding development: An example of mid-childhood reaching. Front. Psychol. 2017, 8, 1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, M.C.; Darrah, J.; Pollock, N.; Wilson, B.; Russell, D.J.; Walter, S.D.; Rosenbaum, P.; Galuppi, B. Focus on function: A cluster, randomized controlled trial comparing child-versus context-focused intervention for young children with cerebral palsy. Dev. Med. Child Neurol. 2011, 53, 621–629. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tele-Heri, B.; Dobos, K.; Harsanyi, S.; Palinkas, J.; Fenyosi, F.; Gesztelyi, R.; More, C.E.; Zsuga, J. Vestibular Stimulation May Drive Multisensory Processing: Principles for Targeted Sensorimotor Therapy (TSMT). Brain Sci. 2021, 11, 1111. https://doi.org/10.3390/brainsci11081111
Tele-Heri B, Dobos K, Harsanyi S, Palinkas J, Fenyosi F, Gesztelyi R, More CE, Zsuga J. Vestibular Stimulation May Drive Multisensory Processing: Principles for Targeted Sensorimotor Therapy (TSMT). Brain Sciences. 2021; 11(8):1111. https://doi.org/10.3390/brainsci11081111
Chicago/Turabian StyleTele-Heri, Brigitta, Karoly Dobos, Szilvia Harsanyi, Judit Palinkas, Fanni Fenyosi, Rudolf Gesztelyi, Csaba E. More, and Judit Zsuga. 2021. "Vestibular Stimulation May Drive Multisensory Processing: Principles for Targeted Sensorimotor Therapy (TSMT)" Brain Sciences 11, no. 8: 1111. https://doi.org/10.3390/brainsci11081111