
Increased Vocalization of Rats in Response to Ultrasonic Playback as a Sign of Hypervigilance Following Fear Conditioning

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Surgical Procedures
2.3. Fear Conditioning
2.4. Playback Experiment
2.5. Ultrasonic Playback Presentation
2.6. Analysis of USV and Locomotor Activity
2.7. Statistical Analysis
3. Results
3.1. Rats Showed Freezing after Fear Conditioning
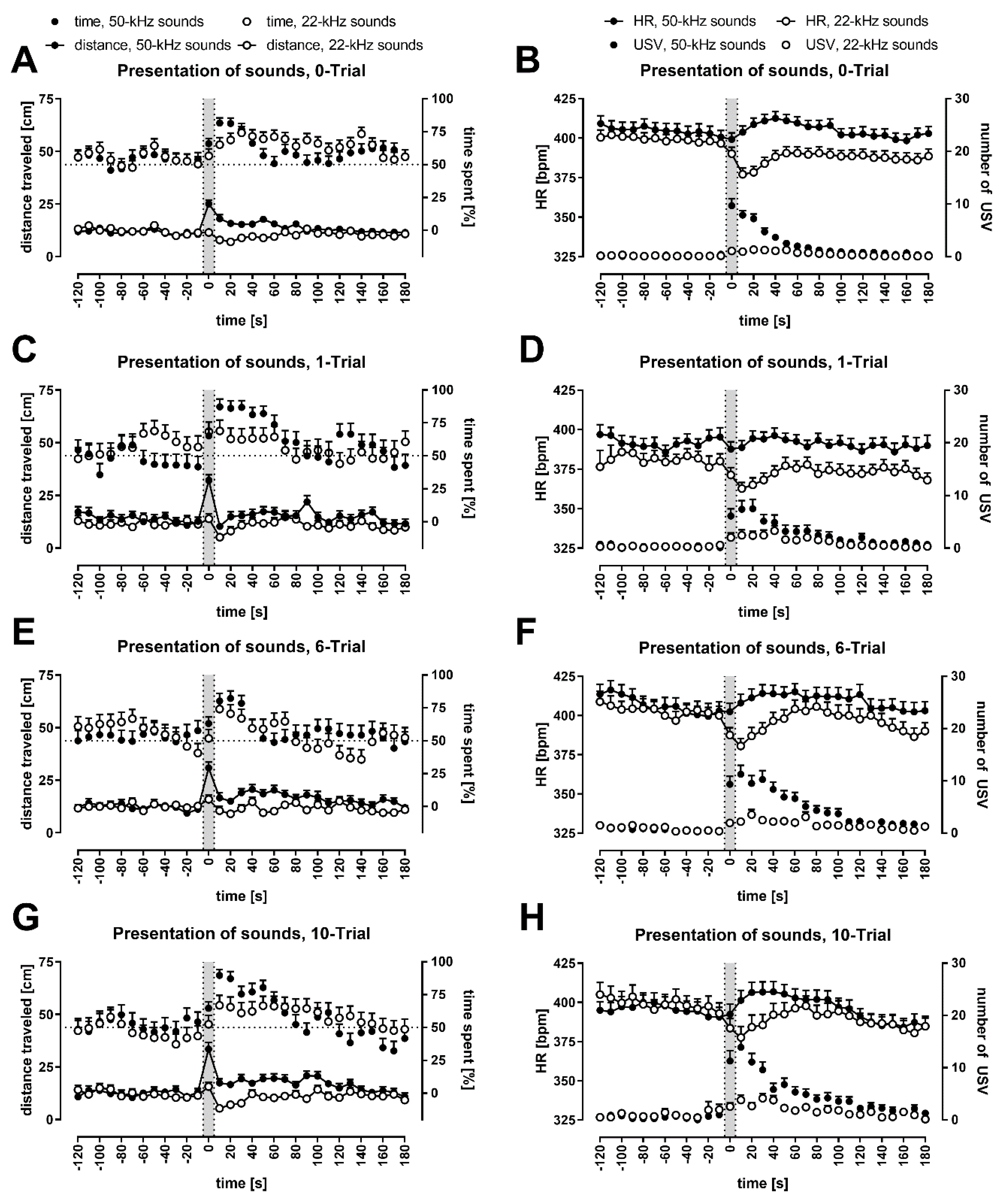
3.2. Except for the Periods of Ultrasonic Playback, Rats’ Behavior Remained Relatively Constant
3.3. Animals Moved Faster during 50-kHz Ultrasonic Presentations and Slowed Down after 22-kHz Ultrasonic Presentations
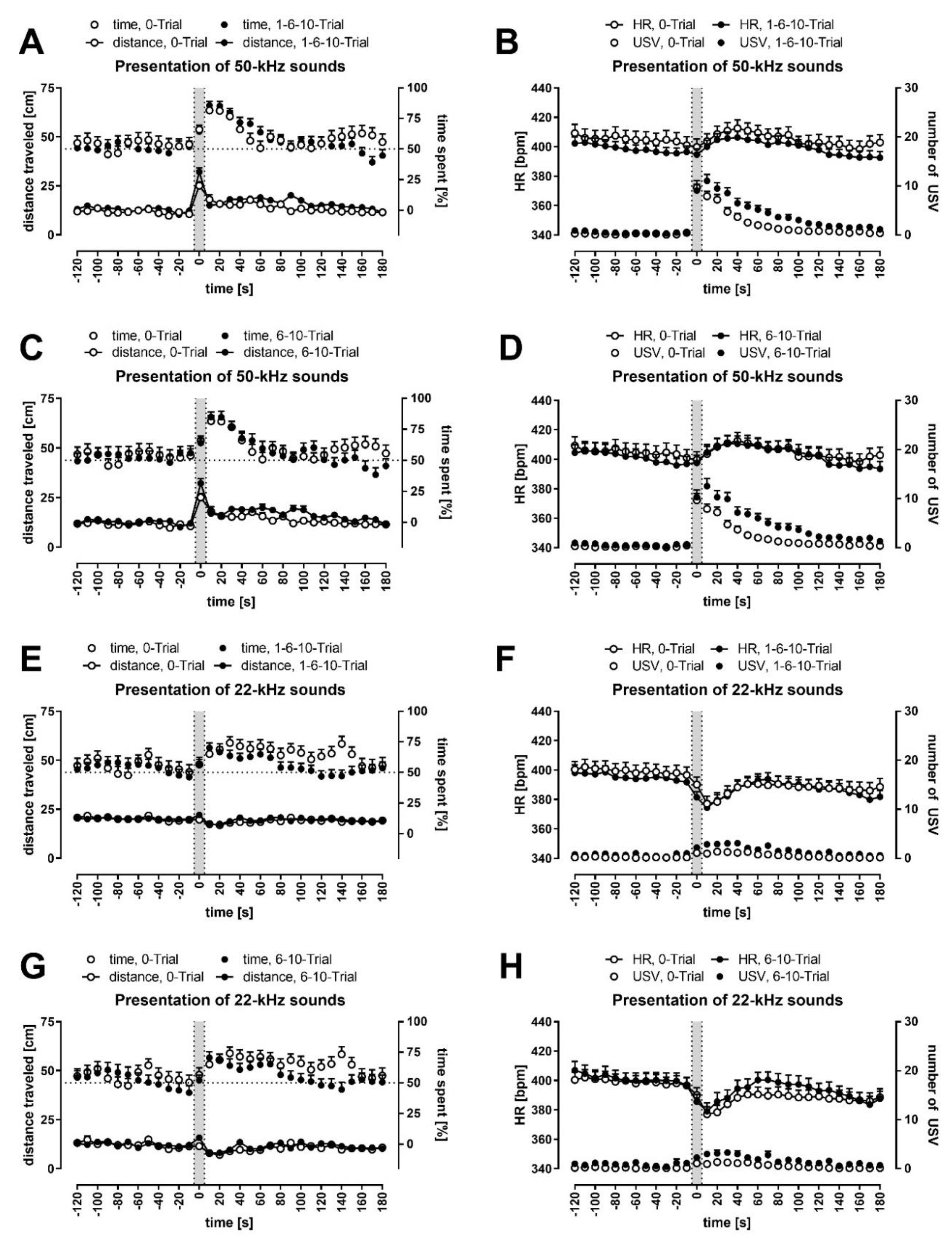
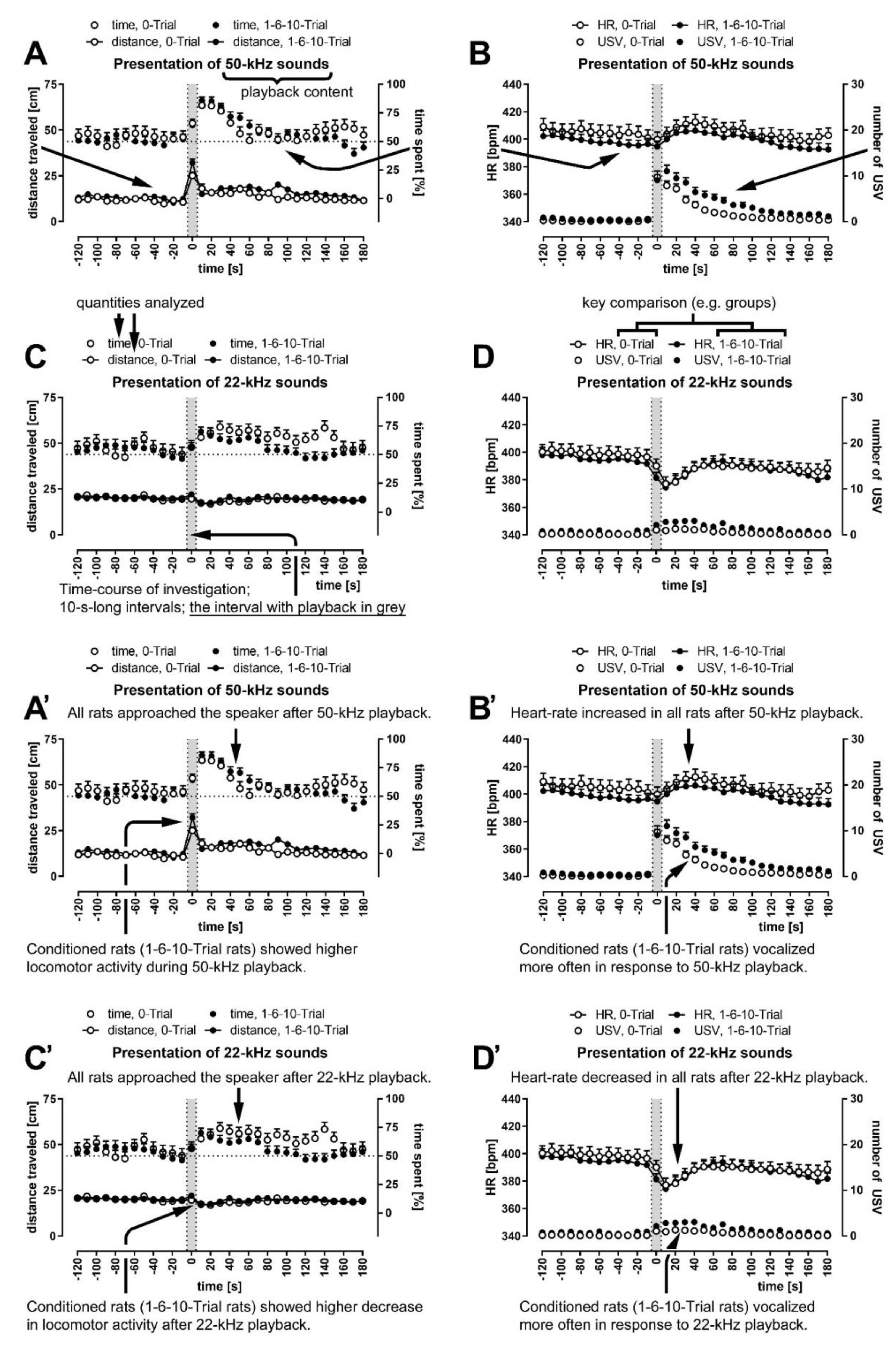
3.4. Conditioned Rats Showed Higher Locomotor Activity during 50-kHz Playback
3.5. Conditioned Rats Showed a More Significant Decrease in Activity Following 22-kHz Playback
3.6. All Rats Approached the Speaker; It Was More Pronounced during and Following 50-kHz Playback
3.7. HR Levels Declined during the Whole Experimental Session
3.8. 50-kHz Sounds Caused HR to Increase; 22-kHz Sounds Caused HR to Decrease
3.9. One-Trial Rats Showed Lower HR Levels and a Smaller Response to 50-kHz Playback
3.10. Rats Vocalized More Often during and Following 50-kHz Playback Than 22-kHz Playback
3.11. Natural and Artificial Ultrasounds Produced Similar Results, but Still, Some Differences Stood Out
3.12. Previously-Shocked Rats Vocalized More Often, with Longer and Higher Frequency USV
4. Discussion
- −
- Rats’ overt behavior remained relatively constant except during ultrasonic playback;
- −
- Rats moved faster during 50-kHz ultrasonic presentations;
- −
- Rats slowed down right after 22-kHz ultrasonic presentations;
- −
- Rats approached the speaker during and following 50-kHz and 22-kHz playbacks;
- −
- The approach was more pronounced during and following 50-kHz playback;
- −
- HR levels declined during the whole experimental session;
- −
- Rats’ HR increase after exposure to 50-kHz playback;
- −
- Rats’ HR decrease when exposed to 22-kHz playback;
- −
- The difference in HR following 50-kHz vs. 22-kHz playback lasts for at least 3 min;
- −
- Both 50- and 22-kHz sounds evoked an ultrasonic response, mainly in the 50-kHz range;
- −
- 50-kHz USV emitted in response to 50-kHz playback were longer and of higher frequency;
- −
- Rats vocalized more often during and following 50-kHz playback than 22-kHz playback;
- −
- In general, rats reacted in a similar way to both natural and artificial ultrasonic playback.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bali, A.; Jaggi, A.S. Electric foot shock stress: A useful tool in neuropsychiatric studies. Rev. Neurosci. 2015, 26, 655–677. [Google Scholar] [CrossRef]
- Careaga, M.B.L.; Girardi, C.E.N.; Suchecki, D. Understanding posttraumatic stress disorder through fear conditioning, extinction and reconsolidation. Neurosci. Biobehav. Rev. 2016, 71, 48–57. [Google Scholar] [CrossRef]
- Aliczki, M.; Haller, J. Electric shock as model of post-traumatic stress disorder in rodents. In Comprehensive Guide to Post-Traumatic Stress Disorder; Martin, C.R., Preedy, V.R., Patel, V.B., Eds.; Springer International Publishing: Basel, Switzerland, 2014. [Google Scholar]
- Verbitsky, A.; Dopfel, D.; Zhang, N. Rodent models of post-traumatic stress disorder: Behavioral assessment. Transl. Psychiatry 2020, 10, 132. [Google Scholar] [CrossRef]
- Asok, A.; Kandel, E.R.; Rayman, J.B. The Neurobiology of Fear Generalization. Front. Behav. Neurosci. 2018, 12, 329. [Google Scholar] [CrossRef]
- Fanselow, M.S. Conditioned and unconditional components of post-shock freezing. Pavlov. J. Biol. Sci. 1980, 15, 177–182. [Google Scholar] [CrossRef]
- Frankland, P.W.; Cestari, V.; Filipkowski, R.K.; McDonald, R.J.; Silva, A.J. The dorsal hippocampus is essential for context discrimination but not for contextual conditioning. Behav. Neurosci. 1998, 112, 863–874. [Google Scholar] [CrossRef]
- Blanchard, R.J.; Blanchard, D.C. Passive and active reactions to fear-eliciting stimuli. J. Comp. Physiol. Psychol. 1969, 68, 129–135. [Google Scholar] [CrossRef]
- Katz, R.J.; Roth, K.A.; Carroll, B.J. Acute and chronic stress effects on open field activity in the rat: Implications for a model of depression. Neurosci. Biobehav. Rev. 1981, 5, 247–251. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, R.; Rodrigues, H.; Novaes, F.; Arbol, J.; Volchan, E.; Coutinho, E.S.; Figueira, I.; Ventura, P. Listening to the heart: A meta-analysis of cognitive behavior therapy impact on the heart rate of patients with anxiety disorders. J. Affect. Disord. 2015, 172, 231–240. [Google Scholar] [CrossRef]
- Pittig, A.; Arch, J.J.; Lam, C.W.; Craske, M.G. Heart rate and heart rate variability in panic, social anxiety, obsessive-compulsive, and generalized anxiety disorders at baseline and in response to relaxation and hyperventilation. Int. J. Psychophysiol. 2013, 87, 19–27. [Google Scholar] [CrossRef]
- Nijsen, M.J.; Croiset, G.; Diamant, M.; Stam, R.; Delsing, D.; de Wied, D.; Wiegant, V.M. Conditioned fear-induced tachycardia in the rat: Vagal involvement. Eur. J. Pharmacol. 1998, 350, 211–222. [Google Scholar] [CrossRef]
- Carnevali, L.; Bondarenko, E.; Sgoifo, A.; Walker, F.R.; Head, G.A.; Lukoshkova, E.V.; Day, T.A.; Nalivaiko, E. Metyrapone and fluoxetine suppress enduring behavioral but not cardiac effects of subchronic stress in rats. Am. J. Physiol. Regul. Integr. Comp Physiol. 2011, 301, R1123–R1131. [Google Scholar] [CrossRef]
- Resstel, L.B.; Alves, F.H.; Reis, D.G.; Crestani, C.C.; Correa, F.M.; Guimaraes, F.S. Anxiolytic-like effects induced by acute reversible inactivation of the bed nucleus of stria terminalis. Neuroscience 2008, 154, 869–876. [Google Scholar] [CrossRef]
- Resstel, L.B.; Correa, F.M.; Guimaraes, F.S. The expression of contextual fear conditioning involves activation of an NMDA receptor-nitric oxide pathway in the medial prefrontal cortex. Cereb. Cortex 2008, 18, 2027–2035. [Google Scholar] [CrossRef] [Green Version]
- Antoniadis, E.A.; McDonald, R.J. Discriminative fear conditioning to context expressed by multiple measures of fear in the rat. Behav. Brain Res. 1999, 101, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Carrive, P. Conditioned fear to environmental context: Cardiovascular and behavioral components in the rat. Brain Res. 2000, 858, 440–445. [Google Scholar] [CrossRef]
- Zhang, W.N.; Murphy, C.A.; Feldon, J. Behavioural and cardiovascular responses during latent inhibition of conditioned fear: Measurement by telemetry and conditioned freezing. Behav. Brain Res. 2004, 154, 199–209. [Google Scholar] [CrossRef]
- Meerlo, P.; Sgoifo, A.; De Boer, S.F.; Koolhaas, J.M. Long-lasting consequences of a social conflict in rats: Behavior during the interaction predicts subsequent changes in daily rhythms of heart rate, temperature, and activity. Behav. Neurosci. 1999, 113, 1283–1290. [Google Scholar] [CrossRef]
- Burgdorf, J.S.; Brudzynski, S.M.; Moskal, J.R. Using rat ultrasonic vocalization to study the neurobiology of emotion: From basic science to the development of novel therapeutics for affective disorders. Curr. Opin. Neurobiol. 2020, 60, 192–200. [Google Scholar] [CrossRef]
- Simola, N.; Granon, S. Ultrasonic vocalizations as a tool in studying emotional states in rodent models of social behavior and brain disease. Neuropharmacology 2019, 159, 107420. [Google Scholar] [CrossRef]
- Wohr, M.; Schwarting, R.K. Affective communication in rodents: Ultrasonic vocalizations as a tool for research on emotion and motivation. Cell Tissue Res. 2013, 354, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Jelen, P.; Soltysik, S.; Zagrodzka, J. 22-kHz ultrasonic vocalization in rats as an index of anxiety but not fear: Behavioral and pharmacological modulation of affective state. Behav. Brain Res. 2003, 141, 63–72. [Google Scholar] [CrossRef]
- Wohr, M.; Borta, A.; Schwarting, R.K. Overt behavior and ultrasonic vocalization in a fear conditioning paradigm: A dose-response study in the rat. Neurobiol. Learn Mem. 2005, 84, 228–240. [Google Scholar] [CrossRef]
- Brudzynski, S.M. Ethotransmission: Communication of emotional states through ultrasonic vocalization in rats. Curr. Opin. Neurobiol. 2013, 23, 310–317. [Google Scholar] [CrossRef]
- Olszynski, K.H.; Polowy, R.; Malz, M.; Boguszewski, P.M.; Filipkowski, R.K. Playback of Alarm and Appetitive Calls Differentially Impacts Vocal, Heart-Rate, and Motor Response in Rats. iScience 2020, 23, 101577. [Google Scholar] [CrossRef]
- Pestana-Oliveira, N.; Nahey, D.B.; Johnson, T.; Collister, J.P. Development of the Deoxycorticosterone Acetate (DOCA)-salt Hypertensive Rat Model. Bio Protoc. 2020, 10, e3708. [Google Scholar] [CrossRef]
- Lindquist, D.H.; Jarrard, L.E.; Brown, T.H. Perirhinal cortex supports delay fear conditioning to rat ultrasonic social signals. J. Neurosci. 2004, 24, 3610–3617. [Google Scholar] [CrossRef] [Green Version]
- Sadananda, M.; Wohr, M.; Schwarting, R.K. Playback of 22-kHz and 50-kHz ultrasonic vocalizations induces differential c-fos expression in rat brain. Neurosci. Lett. 2008, 435, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Berg, E.L.; Copping, N.A.; Rivera, J.K.; Pride, M.C.; Careaga, M.; Bauman, M.D.; Berman, R.F.; Lein, P.J.; Harony-Nicolas, H.; Buxbaum, J.D.; et al. Developmental social communication deficits in the Shank3 rat model of phelan-mcdermid syndrome and autism spectrum disorder. Autism Res. 2018, 11, 587–601. [Google Scholar] [CrossRef]
- Seffer, D.; Rippberger, H.; Schwarting, R.K.; Wohr, M. Pro-social 50-kHz ultrasonic communication in rats: Post-weaning but not post-adolescent social isolation leads to social impairments-phenotypic rescue by re-socialization. Front. Behav. Neurosci. 2015, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Wohr, M.; Schwarting, R.K. Ultrasonic communication in rats: Effects of morphine and naloxone on vocal and behavioral responses to playback of 50-kHz vocalizations. Pharmacol. Biochem. Behav. 2009, 94, 285–295. [Google Scholar] [CrossRef]
- Wohr, M.; Schwarting, R.K. Ultrasonic communication in rats: Can playback of 50-kHz calls induce approach behavior? PLoS ONE 2007, 2, e1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burman, O.; Owen, D.; Abouismail, U.; Mendl, M. Removing individual rats affects indicators of welfare in the remaining group members. Physiol. Behav. 2008, 93, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Seffer, D.; Schwarting, R.K.; Wohr, M. Pro-social ultrasonic communication in rats: Insights from playback studies. J. Neurosci. Methods 2014, 234, 73–81. [Google Scholar] [CrossRef]
- Snoeren, E.M.; Agmo, A. The incentive value of males’ 50-kHz ultrasonic vocalizations for female rats (Rattus norvegicus). J. Comp. Psychol. 2014, 128, 40–55. [Google Scholar] [CrossRef] [PubMed]
- Davidson, N.B.; Hurst, J.L. Testing the potential of 50 kHz rat calls as a species-specific rat attractant. PLoS ONE 2019, 14, e0211601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schonfeld, L.M.; Zech, M.P.; Schable, S.; Wohr, M.; Kalenscher, T. Lesions of the rat basolateral amygdala reduce the behavioral response to ultrasonic vocalizations. Behav. Brain Res. 2020, 378, 112274. [Google Scholar] [CrossRef]
- Brudzynski, S.M.; Chiu, E.M. Behavioural responses of laboratory rats to playback of 22 kHz ultrasonic calls. Physiol. Behav. 1995, 57, 1039–1044. [Google Scholar] [CrossRef]
- Wohr, M.; Seffer, D.; Schwarting, R.K. Studying Socio-Affective Communication in Rats through Playback of Ultrasonic Vocalizations. Curr. Protoc. Neurosci. 2016, 75, 8–35. [Google Scholar] [CrossRef]
- Inagaki, H.; Ushida, T. The effect of playback of 22-kHz and 50-kHz ultrasonic vocalizations on rat behaviors assessed with a modified open-field test. Physiol. Behav. 2021, 229, 113251. [Google Scholar] [CrossRef] [PubMed]
- Demaestri, C.; Brenhouse, H.C.; Honeycutt, J.A. 22 kHz and 55 kHz ultrasonic vocalizations differentially influence neural and behavioral outcomes: Implications for modeling anxiety via auditory stimuli in the rat. Behav. Brain Res. 2019, 360, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Willuhn, I.; Tose, A.; Wanat, M.J.; Hart, A.S.; Hollon, N.G.; Phillips, P.E.; Schwarting, R.K.; Wohr, M. Phasic dopamine release in the nucleus accumbens in response to pro-social 50 kHz ultrasonic vocalizations in rats. J. Neurosci. 2014, 34, 10616–10623. [Google Scholar] [CrossRef]
- Vianna, D.M.; Brandao, M.L. Anatomical connections of the periaqueductal gray: Specific neural substrates for different kinds of fear. Braz. J. Med. Biol. Res. 2003, 36, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Koba, S.; Inoue, R.; Watanabe, T. Role played by periaqueductal gray neurons in parasympathetically mediated fear bradycardia in conscious rats. Physiol. Rep. 2016, 4. [Google Scholar] [CrossRef]
- Porges, S.W. Orienting in a defensive world: Mammalian modifications of our evolutionary heritage. A Polyvagal Theory. Psychophysiology 1995, 32, 301–318. [Google Scholar] [CrossRef]
- Porges, S.W. The polyvagal perspective. Biol. Psychol. 2007, 74, 116–143. [Google Scholar] [CrossRef] [PubMed]
- Porges, S.W. The polyvagal theory: New insights into adaptive reactions of the autonomic nervous system. Cleve Clin. J. Med. 2009, 76, S86–S90. [Google Scholar] [CrossRef]
- Daviu, N.; Fuentes, S.; Nadal, R.; Armario, A. A single footshock causes long-lasting hypoactivity in unknown environments that is dependent on the development of contextual fear conditioning. Neurobiol. Learn Mem. 2010, 94, 183–190. [Google Scholar] [CrossRef]
- Sawamura, T.; Shimizu, K.; Nibuya, M.; Wakizono, T.; Suzuki, G.; Tsunoda, T.; Takahashi, Y.; Nomura, S. Effect of paroxetine on a model of posttraumatic stress disorder in rats. Neurosci. Lett. 2004, 357, 37–40. [Google Scholar] [CrossRef]
- Mikics, E.; Baranyi, J.; Haller, J. Rats exposed to traumatic stress bury unfamiliar objects—A novel measure of hyper-vigilance in PTSD models? Physiol. Behav. 2008, 94, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Armario, A.; Escorihuela, R.M.; Nadal, R. Long-term neuroendocrine and behavioural effects of a single exposure to stress in adult animals. Neurosci. Biobehav. Rev. 2008, 32, 1121–1135. [Google Scholar] [CrossRef]
- Grillon, C.; Ameli, R.; Foot, M.; Davis, M. Fear-potentiated startle: Relationship to the level of state/trait anxiety in healthy subjects. Biol. Psychiatry 1993, 33, 566–574. [Google Scholar] [CrossRef]
- Djordjevic, J.; Djordjevic, A.; Adzic, M.; Radojcic, M.B. Effects of chronic social isolation on Wistar rat behavior and brain plasticity markers. Neuropsychobiology 2012, 66, 112–119. [Google Scholar] [CrossRef]
- Robbins, T.W. Neurobehavioural sequelae of social deprivation in rodents revisited: Modelling social adversity for developmental neuropsychiatric disorders. J. Psychopharmacol. 2016, 30, 1082–1089. [Google Scholar] [CrossRef]
- Brudzynski, S.M. Principles of rat communication: Quantitative parameters of ultrasonic calls in rats. Behav. Genet. 2005, 35, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Karwicka, W.; Wiatrowska, M.; Kondrakiewicz, K.; Knapska, E.; Kursa, M.B.; Hamed, A. Relaying Aversive Ultrasonic Alarm Calls Depends on Previous Experience. Empathy, Social Buffering, or Panic? Brain Sci. 2021, 11, 759. [Google Scholar] [CrossRef]
- Pezze, M.A.; Feldon, J. Mesolimbic dopaminergic pathways in fear conditioning. Prog. Neurobiol. 2004, 74, 301–320. [Google Scholar] [CrossRef]
- Ringel, L.E.; Basken, J.N.; Grant, L.M.; Ciucci, M.R. Dopamine D1 and D2 receptor antagonism effects on rat ultrasonic vocalizations. Behav. Brain Res. 2013, 252, 252–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carnevali, L.; Sgoifo, A. Vagal modulation of resting heart rate in rats: The role of stress, psychosocial factors, and physical exercise. Front. Physiol. 2014, 5, 118. [Google Scholar] [CrossRef] [Green Version]
- Armario, A.; Labad, J.; Nadal, R. Focusing attention on biological markers of acute stressor intensity: Empirical evidence and limitations. Neurosci. Biobehav. Rev. 2020, 111, 95–103. [Google Scholar] [CrossRef]
- Buckley, T.C.; Kaloupek, D.G. A meta-analytic examination of basal cardiovascular activity in posttraumatic stress disorder. Psychosom. Med. 2001, 63, 585–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pole, N. The psychophysiology of posttraumatic stress disorder: A meta-analysis. Psychol. Bull. 2007, 133, 725–746. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Arlington, VA, USA, 2013. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Groups | Number of USV | Parameters of 50-kHz USV | ||||
|---|---|---|---|---|---|---|
| Total USV | 50-kHz USV | Short 22-kHz | Long 22-kHz | Duration [ms] | MPF [kHz] | |
| USV emitted during the whole experiment | ||||||
| 0-Trial | 152.7 ± 24.2 | 141.6 ± 23.8 | 10.7 ± 2.6 | 0.4 ± 0.4 | 25.8 ± 1.2 | 58.0 ± 0.9 |
| 1-Trial | 230.0 ± 68.0 | 205.7 ± 66.7 | 22.2 ± 8.3 | 2.0 ± 2.0 | 26.3 ± 2.0 | 58.3 ± 1.5 |
| 6-Trial | 366.1 ± 84.9 | 346.7 ± 85.2 | 9.1 ± 1.9 | 10.4 ± 7.1 | 25.3 ± 2.1 | 61.7 ± 1.1 |
| 10-Trial | 354.2 ± 84.7 ** | 336.1 ± 84.2 ** | 10.7 ± 2.2 | 7.3 ± 7.3 | 29.0 ± 1.8 | 60.1 ± 0.4 |
| all FC’ed | 322.4 ± 46.8 * | 302.0 ± 46.7 | 13.5 ± 2.7 | 6.9 ± 3.6 | 26.9 ± 1.1 | 60.2 ± 0.6 |
| USV emitted during the first 10 min of silence | ||||||
| 0-Trial | 23.9 ± 7f.1 | 19.4 ± 7.0 | 4.5 ± 1.0 | 0.0 ± 0.0 | 15.7 ± 1.7 | 52.1 ± 1.9 |
| 1-Trial | 53.6 ± 18.0 * | 38.4 ± 15.6 | 13.1 ± 6.6 | 2.0 ± 2.0 | 22.2 ± 3.4 | 52.7 ± 2.9 |
| 6-Trial | 72.8 ± 26.6 | 57.8 ± 26.6 | 4.6 ± 1.2 | 10.4 ± 7.1 | 21.7 ± 2.1 * | 55.5 ± 1.1 |
| 10-Trial | 49.7 ± 23.1 | 38.1 ± 22.3 | 4.4 ± 1.3 | 7.3 ± 7.3 | 21.0 ± 2.0 * | 56.6 ± 1.7 |
| all FC’ed | 59.2 ± 13.4 | 45.3 ± 13.0 | 7.0 ± 2.0 | 6.9 ± 3.6 | 21.6 ± 1.4 ** | 55.0 ± 1.2 |
| USV emitted to 50-kHz USV playback (0–120 s time-intervals) | ||||||
| 0-Trial | 41.2 ± 7.1 | 39.9 ± 6.9 | 0.9 ± 0.2 | 0.3 ± 0.3 | 27.6 ± 1.7 | 60.5 ± 1.5 |
| 1-Trial | 59.1 ± 19.5 | 57.8 ± 19.3 | 1.3 ± 0.6 | 0.0 ± 0.0 | 24.8 ± 2.2 | 60.4 ± 1.1 |
| 6-Trial | 86.4 ± 16.5 | 85.7 ± 16.5 | 0.8 ± 0.3 | 0.0 ± 0.0 | 27.3 ± 2.6 | 62.8 ± 1.4 |
| 10-Trial | 97.5 ± 17.5 ** | 95.8 ± 17.2 ** | 1.7 ± 0.7 | 0.0 ± 0.0 | 30.6 ± 1.8 | 60.9 ± 0.7 |
| all FC’ed | 82.3 ± 10.3 * | 81.0 ± 10.2 * | 1.2 ± 0.3 | 0.0 ± 0.0 | 27.8 ± 1.3 | 61.4 ± 0.6 |
| USV emitted to 22-kHz USV playback (0–120 s time-intervals) | ||||||
| 0-Trial | 13.5 ± 3.6 | 12.9 ± 3.6 | 0.6 ± 0.3 | 0.0 ± 0.0 | 21.5 ± 1.9 | 58.7 ± 1.8 |
| 1-Trial | 14.6 ± 5.4 | 13.4 ± 5.3 | 1.1 ± 0.6 | 0.0 ± 0.0 | 31.3 ± 13.1 | 55.9 ± 3.3 |
| 6-Trial | 32.2 ± 7.6 * | 31.9 ± 7.6 | 0.3 ± 0.1 | 0.0 ± 0.0 | 29.4 ± 2.4 * | 60.1 ± 0.7 |
| 10-Trial | 30.9 ± 9.8 | 29.9 ± 9.8 | 1.1 ± 0.5 | 0.0 ± 0.0 | 23.8 ± 2.4 | 58.9 ± 1.8 |
| all FC’ed | 26.6 ± 4.7 | 25.8 ± 4.7 | 0.8 ± 0.3 | 0.0 ± 0.0 | 27.7 ± 3.5 | 58.5 ± 1.1 |
| USV emitted to 50-kHz tone playback (0–120 s time-intervals) | ||||||
| 0-Trial | 41.4 ± 6.6 | 40.4 ± 6.5 | 1.0 ± 0.4 | 0.0 ± 0.0 | 27.5 ± 1.3 | 59.1 ± 0.7 |
| 1-Trial | 49.3 ± 16.2 | 48.6 ± 16.1 | 0.8 ± 0.2 | 0.0 ± 0.0 | 25.4 ± 2.2 | 60.6 ± 1.4 |
| 6-Trial | 76.0 ± 18.5 | 74.6 ± 18.4 | 1.4 ± 0.5 | 0.0 ± 0.0 | 30.1 ± 2.9 | 60.1 ± 1.1 |
| 10-Trial | 76.3 ± 15.3 | 75.4 ± 15.2 | 0.9 ± 0.4 | 0.0 ± 0.0 | 31.7 ± 2.2 | 60.6 ± 0.6 |
| all FC’ed | 68.3 ± 9.7 | 67.3 ± 9.7 | 1.0 ± 0.2 | 0.0 ± 0.0 | 29.3 ± 1.4 | 60.5 ± 0.6 * |
| USV emitted to 22-kHz tone playback (0–120 s time-intervals) | ||||||
| 0-Trial | 6.6 ± 2.1 | 6.1 ± 2.0 | 0.5 ± 0.3 | 0.0 ± 0.0 | 17.5 ± 2.1 | 55.5 ± 1.9 |
| 1-Trial | 16.0 ± 6.9 | 15.3 ± 6.9 | 0.7 ± 0.5 | 0.0 ± 0.0 | 27.8 ± 7.1 | 54.9 ± 2.7 |
| 6-Trial | 18.1 ± 5.5 * | 17.8 ± 5.5 | 0.4 ± 0.2 | 0.0 ± 0.0 | 26.9 ± 3.3 * | 60.1 ± 0.9 |
| 10-Trial | 23.3 ± 8.1 | 23.2 ± 8.1 * | 0.1 ± 0.1 | 0.0 ± 0.0 | 25.1 ± 2.2 * | 58.3 ± 1.4 |
| all FC’ed | 19.3 ± 3.9 * | 18.9 ± 3.9 * | 0.4 ± 0.2 | 0.0 ± 0.0 | 26.5 ± 2.4 ** | 57.9 ± 1.0 |
| USV emitted to 50-kHz sound playback (0–120 s time-intervals) | ||||||
| 0-Trial | 41.3 ± 6.0 | 40.1 ± 5.9 | 1.0 ± 0.3 | 0.1 ± 0.1 | 28.0 ± 1.4 | 59.9 ± 0.8 |
| 1-Trial | 54.2 ± 17.4 | 53.2 ± 17.2 | 1.0 ± 0.4 | 0.0 ± 0.0 | 25.1 ± 1.8 | 60.5 ± 1.1 |
| 6-Trial | 81.2 ± 16.4 | 80.1 ± 16.3 | 1.1 ± 0.3 | 0.0 ± 0.0 | 27.6 ± 2.7 | 62.4 ± 1.4 |
| 10-Trial | 86.9 ± 15.1 * | 85.6 ± 15.0 ** | 1.3 ± 0.4 | 0.0 ± 0.0 | 31.2 ± 1.9 # | 60.8 ± 0.6 |
| all FC’ed | 75.3 ± 9.4 | 74.2 ± 9.3 | 1.1 ± 0.2 | 0.0 ± 0.0 | 28.2 ± 1.3 | 61.2 ± 0.6 |
| USV emitted to 22-kHz sound playback (0–120 s time-intervals) | ||||||
| 0-Trial | 10.1 ± 2.7 | 9.5 ± 2.6 | 0.6 ± 0.3 | 0.0 ± 0.0 | 18.4 ± 1.6 | 58.3 ± 1.6 |
| 1-Trial | 15.3 ± 4.7 | 14.4 ± 4.7 | 0.9 ± 0.4 | 0.0 ± 0.0 | 36.5 ± 11.6 | 53.7 ± 3.1 |
| 6-Trial | 25.2 ± 5.9 * | 24.8 ± 5.9 * | 0.3 ± 0.1 | 0.0 ± 0.0 | 26.8 ± 2.4 ** | 60.4 ± 0.7 |
| 10-Trial | 27.1 ± 8.3 * | 26.6 ± 8.3 * | 0.6 ± 0.3 | 0.0 ± 0.0 | 23.2 ± 2.2 | 58.0 ± 1.6 |
| all FC’ed | 23.0 ± 3.8 * | 22.4 ± 3.8 * | 0.6 ± 0.2 | 0.0 ± 0.0 | 28.2 ± 3.4 ** | 57.6 ± 1.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olszyński, K.H.; Polowy, R.; Wardak, A.D.; Grymanowska, A.W.; Filipkowski, R.K. Increased Vocalization of Rats in Response to Ultrasonic Playback as a Sign of Hypervigilance Following Fear Conditioning. Brain Sci. 2021, 11, 970. https://doi.org/10.3390/brainsci11080970
Olszyński KH, Polowy R, Wardak AD, Grymanowska AW, Filipkowski RK. Increased Vocalization of Rats in Response to Ultrasonic Playback as a Sign of Hypervigilance Following Fear Conditioning. Brain Sciences. 2021; 11(8):970. https://doi.org/10.3390/brainsci11080970
Chicago/Turabian StyleOlszyński, Krzysztof H., Rafał Polowy, Agnieszka D. Wardak, Aneta W. Grymanowska, and Robert K. Filipkowski. 2021. "Increased Vocalization of Rats in Response to Ultrasonic Playback as a Sign of Hypervigilance Following Fear Conditioning" Brain Sciences 11, no. 8: 970. https://doi.org/10.3390/brainsci11080970