
Competitive Endogenous RNA Network Involving miRNA and lncRNA in Non-Hodgkin Lymphoma: Current Advances and Clinical Perspectives

 ,
,  , and
, and
Abstract
:1. Introduction
2. MiRNAs and lncRNAs Deregulation in Lymphomagenesis
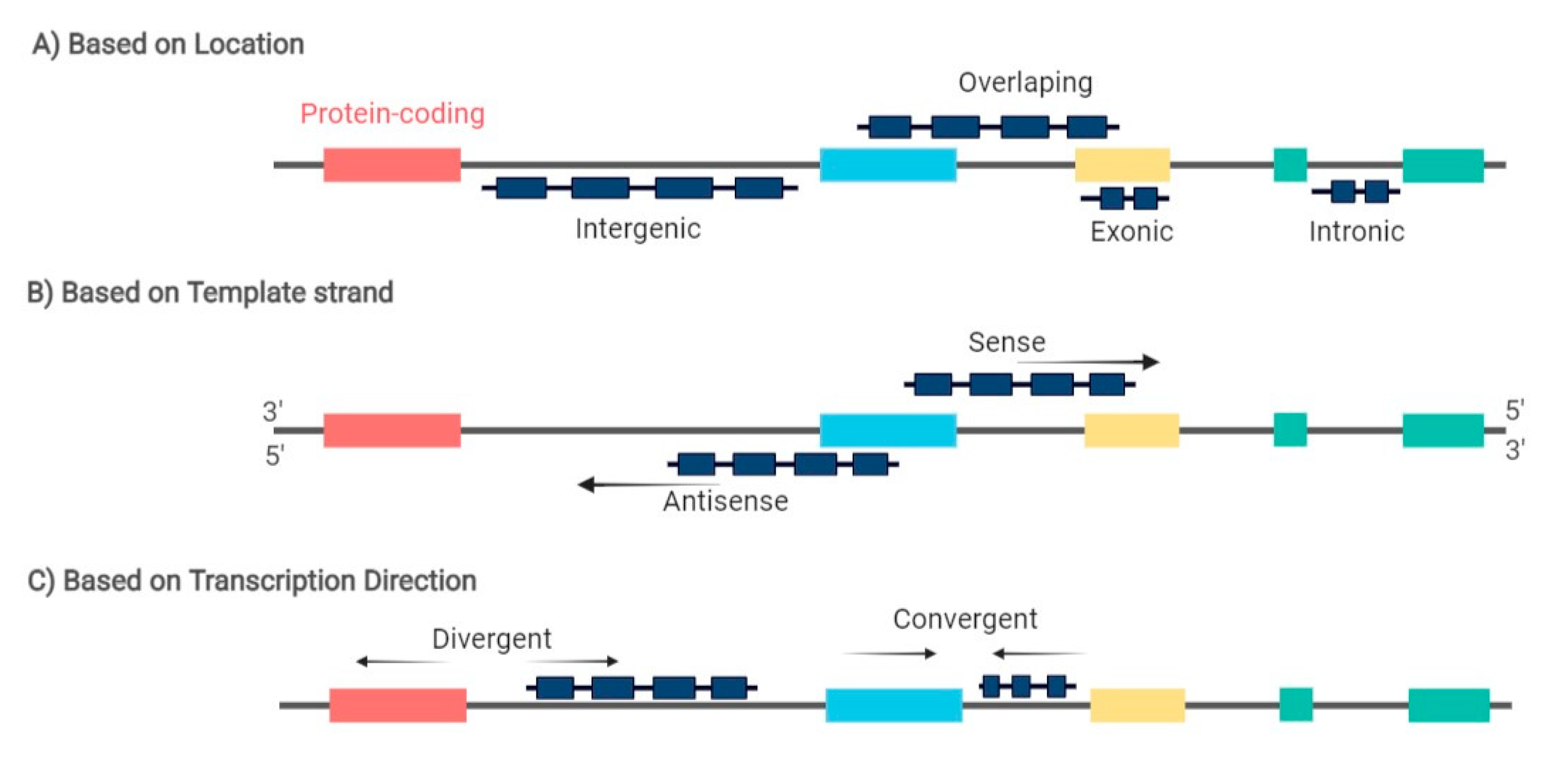
2.1. Evidence Acquisition
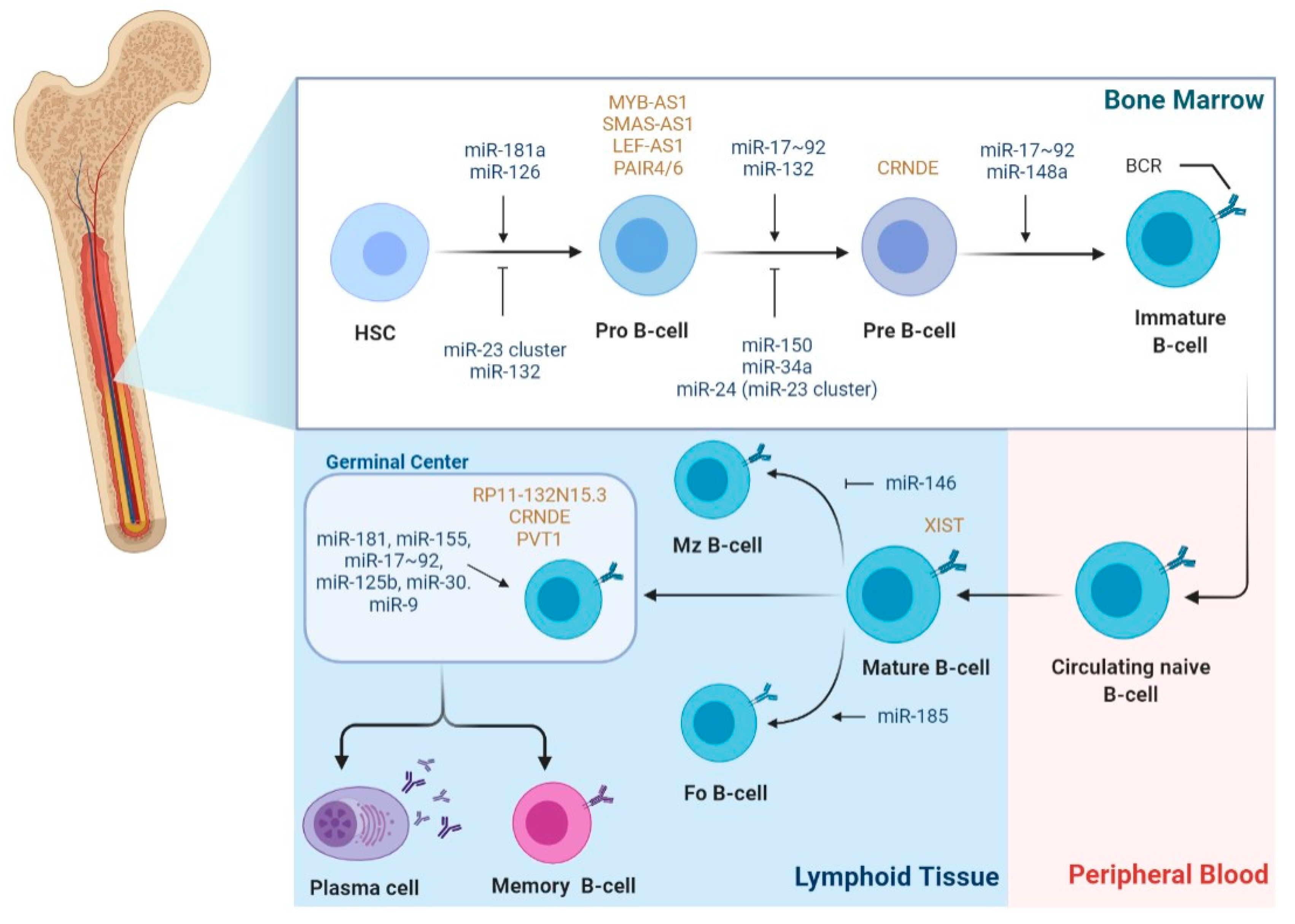
2.2. The Role of miRNAs in B-Cell Lymphomagenesis
2.3. The Role of LncRNAs in B-Cell Lymphomagenesis
2.4. The Role of LncRNAs as ceRNAs in B-Cell Lymphomagenesis
3. MiRNAs and lncRNAs as Potential Biomarkers for NHL
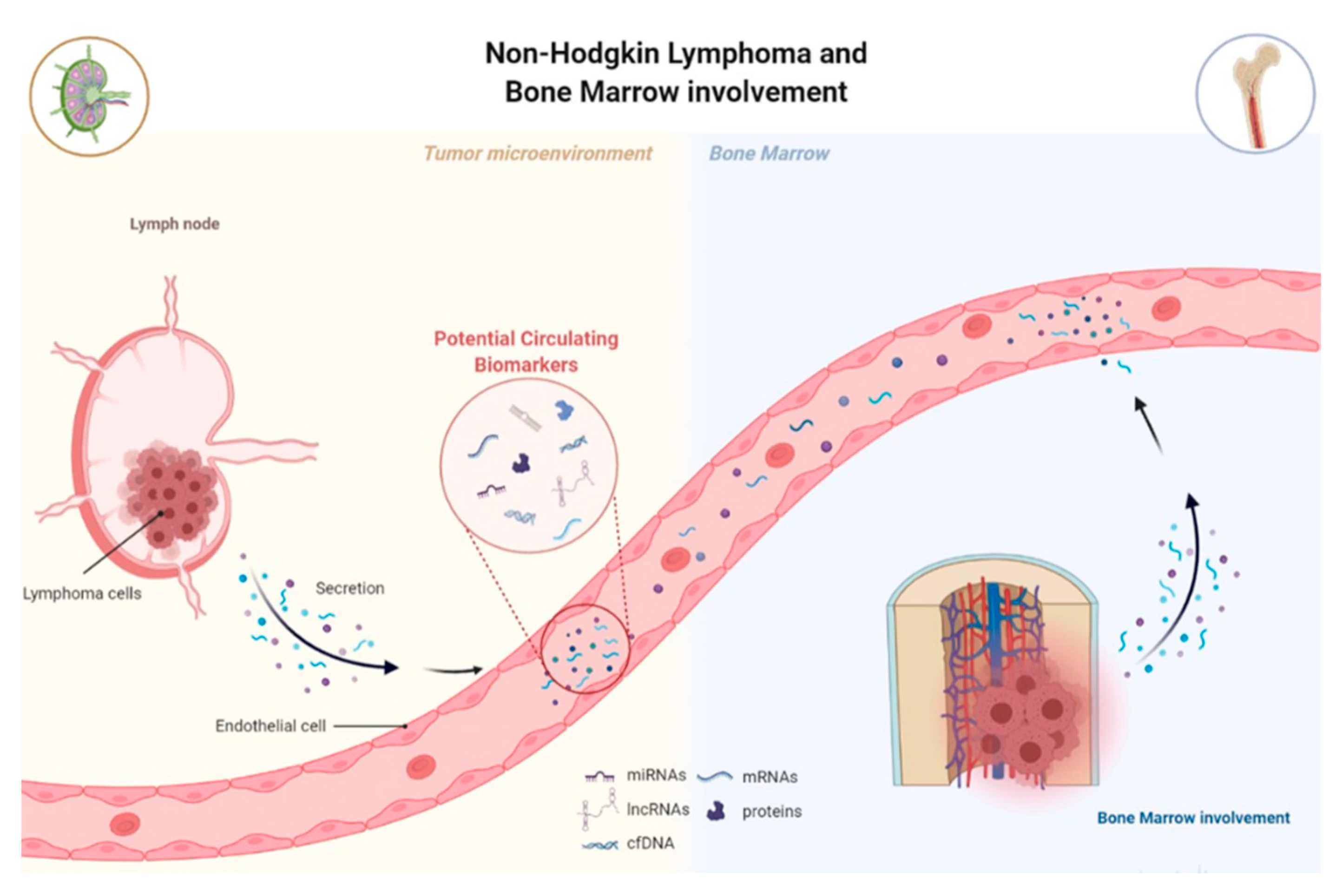
3.1. MiRNAs and lncRNAs as Non-Invasive Diagnostic Biomarkers
3.2. MiRNAs and lncRNAs as Prognostic Biomarkers
3.3. Clinical Trials for Potential miRNA and lncRNA Biomarkers
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Armitage, J.O.; Gascoyne, R.D.; Lunning, M.A.; Cavalli, F. Non-Hodgkin lymphoma. Lancet 2017, 390, 298–310. [Google Scholar] [CrossRef]
- Klener, P.; Klanova, M. Drug Resistance in Non-Hodgkin Lymphomas. Int. J. Mol. Sci. 2020, 21, 2081. [Google Scholar] [CrossRef] [Green Version]
- Rovira, J.; Valera, A.; Colomo, L.; Setoain, X.; Rodríguez, S.; Martínez-Trillos, A.; Giné, E.; Dlouhy, I.; Magnano, L.; Gaya, A. Prognosis of patients with diffuse large B cell lymphoma not reaching complete response or relapsing after frontline chemotherapy or immunochemotherapy. Ann. Hematol. 2015, 94, 803–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crump, M.; Neelapu, S.S.; Farooq, U.; Van Den Neste, E.; Kuruvilla, J.; Westin, J.; Link, B.K.; Hay, A.; Cerhan, J.R.; Zhu, L. Outcomes in refractory diffuse large B-cell lymphoma: Results from the international SCHOLAR-1 study. Blood 2017, 130, 1800–1808. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, M.; Teixeira, A.L.; Medeiros, R. The opportunistic effect of exosomes on Non-Hodgkin Lymphoma microenvironment modulation. Crit. Rev. Oncol. Hematol. 2019, 144, 102825. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.C.R.; Acuña, S.M.; Aoki, J.I.; Floeter-Winter, L.M.; Muxel, S.M. Long Non-Coding RNAs in the Regulation of Gene Expression: Physiology and Disease. Non-Coding RNA 2019, 5, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, A.M.; Chang, H.Y. Long noncoding RNAs in cancer pathways. Cancer Cell 2016, 29, 452–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [Green Version]
- Paraskevopoulou, M.D.; Hatzigeorgiou, A.G. Analyzing MiRNA–LncRNA Interactions. In Long Non-Coding RNAs: Methods and Protocols; Feng, Y., Zhang, L., Eds.; Springer: New York, NY, USA, 2016; pp. 271–286. [Google Scholar]
- Chi, Y.; Wang, D.; Wang, J.; Yu, W.; Yang, J. Long Non-Coding RNA in the Pathogenesis of Cancers. Cells 2019, 8, 1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, Y.; Yang, Z.; Ge, Q.; Yu, L.; Yao, M.; Sun, X.; Ren, Z.; Ding, C. Long non-coding RNA Malat1 activated autophagy, hence promoting cell proliferation and inhibiting apoptosis by sponging miR-101 in colorectal cancer. Cell. Mol. Biol. Lett. 2019, 24, 50. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Luo, W.; Wang, J.; Peng, T.; Sun, G.; Shi, J.; Li, Z.; Zhang, B. Malat1 activates autophagy and promotes cell proliferation by sponging miR-101 and upregulating STMN1, RAB5A and ATG4D expression in glioma. Biochem. Biophys. Res. Commun. 2017, 492, 480–486. [Google Scholar] [CrossRef]
- Xiong, Z.; Wang, L.; Wang, Q.; Yuan, Y. LncRNA MALAT1/miR-129 axis promotes glioma tumorigenesis by targeting SOX2. J. Cell. Mol. Med. 2018, 22, 3929–3940. [Google Scholar] [CrossRef]
- Liao, K.; Lin, Y.; Gao, W.; Xiao, Z.; Medina, R.; Dmitriev, P.; Cui, J.; Zhuang, Z.; Zhao, X.; Qiu, Y.; et al. Blocking lncRNA MALAT1/miR-199a/ZHX1 Axis Inhibits Glioblastoma Proliferation and Progression. Mol. Ther. Nucleic Acids 2019, 18, 388–399. [Google Scholar] [CrossRef] [Green Version]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Greene, J.; Baird, A.-M.; Brady, L.; Lim, M.; Gray, S.G.; McDermott, R.; Finn, S.P. Circular RNAs: Biogenesis, function and role in human diseases. Front. Mol. Biosci. 2017, 4, 38. [Google Scholar] [CrossRef]
- Spierings, D.C.; McGoldrick, D.; Hamilton-Easton, A.M.; Neale, G.; Murchison, E.P.; Hannon, G.J.; Green, D.R.; Withoff, S. Ordered progression of stage-specific miRNA profiles in the mouse B2 B-cell lineage. Blood J. Am. Soc. Hematol. 2011, 117, 5340–5349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benhamou, D.; Labi, V.; Novak, R.; Dai, I.; Shafir-Alon, S.; Weiss, A.; Gaujoux, R.; Arnold, R.; Shen-Orr, S.S.; Rajewsky, K.; et al. A c-Myc/miR17-92/Pten Axis Controls PI3K-Mediated Positive and Negative Selection in B Cell Development and Reconstitutes CD19 Deficiency. Cell Rep. 2016, 16, 419–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neat, M.J.; Foot, N.; Jenner, M.; Goff, L.; Ashcroft, K.; Burford, D.; Dunham, A.; Norton, A.; Lister, T.A.; Fitzgibbon, J. Localisation of a novel region of recurrent amplification in follicular lymphoma to an∼6.8 Mb region of 13q32-33. Genes Chromosomes Cancer 2001, 32, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Monni, O.; Oinonen, R.; Elonen, E.; Franssila, K.; Teerenhovi, L.; Joensuu, H.; Knuutila, S. Gain of 3q and deletion of 11q22 are frequent aberrations in mantle cell lymphoma. Genes Chromosomes Cancer 1998, 21, 298–307. [Google Scholar] [CrossRef]
- Rao, P.H.; Houldsworth, J.; Dyomina, K.; Parsa, N.Z.; Cigudosa, J.C.; Louie, D.C.; Popplewell, L.; Offit, K.; Jhanwar, S.C.; Chaganti, R.S.K. Chromosomal and Gene Amplification in Diffuse Large B-Cell Lymphoma. Blood 1998, 92, 234–240. [Google Scholar] [CrossRef]
- He, L.; Thomson, J.M.; Hemann, M.T.; Hernando-Monge, E.; Mu, D.; Goodson, S.; Powers, S.; Cordon-Cardo, C.; Lowe, S.W.; Hannon, G.J. A microRNA polycistron as a potential human oncogene. Nature 2005, 435, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Srinivasan, L.; Calado, D.P.; Patterson, H.C.; Zhang, B.; Wang, J.; Henderson, J.M.; Kutok, J.L.; Rajewsky, K. Lymphoproliferative disease and autoimmunity in mice with increased miR-17-92 expression in lymphocytes. Nat. Immunol. 2008, 9, 405–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, P.; Han, Y.-C.; Betel, D.; Yao, E.; Squatrito, M.; Ogrodowski, P.; de Stanchina, E.; D’Andrea, A.; Sander, C.; Ventura, A. Genetic dissection of the miR-17∼92 cluster of microRNAs in Myc-induced B-cell lymphomas. Genes Dev. 2009, 23, 2806–2811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.Y.; Oda, H.; Lai, M.; Skalsky, R.L.; Bethel, K.; Shepherd, J.; Kang, S.G.; Liu, W.-H.; Sabouri-Ghomi, M.; Cullen, B.R.; et al. MicroRNA-17∼92 plays a causative role in lymphomagenesis by coordinating multiple oncogenic pathways. EMBO J. 2013, 32, 2377–2391. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Nakagawa, R.; Lazzaro, S.; Staudacher, P.; Abreu-Goodger, C.; Henley, T.; Boiani, S.; Leyland, R.; Galloway, A.; Andrews, S. The miR-155–PU.1 axis acts on Pax5 to enable efficient terminal B cell differentiation. J. Exp. Med. 2014, 211, 2183–2198. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Roccaro, A.M.; Rombaoa, C.; Flores, L.; Obad, S.; Fernandes, S.M.; Sacco, A.; Liu, Y.; Ngo, H.; Quang, P. LNA-mediated anti–miR-155 silencing in low-grade B-cell lymphomas. Blood J. Am. Soc. Hematol. 2012, 120, 1678–1686. [Google Scholar] [CrossRef] [Green Version]
- Vigorito, E.; Perks, K.L.; Abreu-Goodger, C.; Bunting, S.; Xiang, Z.; Kohlhaas, S.; Das, P.P.; Miska, E.A.; Rodriguez, A.; Bradley, A.; et al. microRNA-155 regulates the generation of immunoglobulin class-switched plasma cells. Immunity 2007, 27, 847–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thai, T.-H.; Calado, D.P.; Casola, S.; Ansel, K.M.; Xiao, C.; Xue, Y.; Murphy, A.; Frendewey, D.; Valenzuela, D.; Kutok, J.L.; et al. Regulation of the Germinal Center Response by MicroRNA-155. Science 2007, 316, 604–608. [Google Scholar] [CrossRef]
- Huang, X.; Shen, Y.; Liu, M.; Bi, C.; Jiang, C.; Iqbal, J.; McKeithan, T.W.; Chan, W.C.; Ding, S.-J.; Fu, K. Quantitative proteomics reveals that miR-155 regulates the PI3K-AKT pathway in diffuse large B-cell lymphoma. Am. J. Pathol. 2012, 181, 26–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.-Q.; Zeng, L.; Tang, N.; Tang, Y.-P.; Zhou, B.-P.; Li, F.-F.; Wu, W.-G.; Zeng, X.-B.; Peng, S.-S. MicroRNA-155 downregulation promotes cell cycle arrest and apoptosis in diffuse large B-cell lymphoma. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2016, 24, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Slezak-Prochazka, I.; Kluiver, J.; de Jong, D.; Smigielska-Czepiel, K.; Kortman, G.; Winkle, M.; Rutgers, B.; Koerts, J.; Visser, L.; Diepstra, A. Inhibition of the miR-155 target NIAM phenocopies the growth promoting effect of miR-155 in B-cell lymphoma. Oncotarget 2016, 7, 2391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandhu, S.K.; Volinia, S.; Costinean, S.; Galasso, M.; Neinast, R.; Santhanam, R.; Parthun, M.R.; Perrotti, D.; Marcucci, G.; Garzon, R. miR-155 targets histone deacetylase 4 (HDAC4) and impairs transcriptional activity of B-cell lymphoma 6 (BCL6) in the Eµ-miR-155 transgenic mouse model. Proc. Natl. Acad. Sci. USA 2012, 109, 20047–20052. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, I.M.; Otero, D.; Kao, E.; Miletic, A.V.; Hother, C.; Ralfkiaer, E.; Rickert, R.C.; Gronbaek, K.; David, M. Onco-miR-155 targets SHIP1 to promote TNFα-dependent growth of B cell lymphomas. EMBO Mol. Med. 2009, 1, 288–295. [Google Scholar] [CrossRef]
- Rai, D.; Kim, S.W.; McKeller, M.R.; Dahia, P.L.; Aguiar, R.C. Targeting of SMAD5 links microRNA-155 to the TGF-β pathway and lymphomagenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 3111–3116. [Google Scholar] [CrossRef] [Green Version]
- Gururajan, M.; Haga, C.L.; Das, S.; Leu, C.-M.; Hodson, D.; Josson, S.; Turner, M.; Cooper, M.D. MicroRNA 125b inhibition of B cell differentiation in germinal centers. Int. Immunol. 2010, 22, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; So, A.Y.-L.; Sookram, R.; Wong, S.; Wang, J.K.; Ouyang, Y.; He, P.; Su, Y.; Casellas, R.; Baltimore, D. Epigenetic silencing of miR-125b is required for normal B-cell development. Blood 2018, 131, 1920–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enomoto, Y.; Kitaura, J.; Hatakeyama, K.; Watanuki, J.; Akasaka, T.; Kato, N.; Shimanuki, M.; Nishimura, K.; Takahashi, M.; Taniwaki, M.; et al. Eμ/miR-125b transgenic mice develop lethal B-cell malignancies. Leukemia 2011, 25, 1849–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Jima, D.D.; Jacobs, C.; Fischer, R.; Gottwein, E.; Huang, G.; Lugar, P.L.; Lagoo, A.S.; Rizzieri, D.A.; Friedman, D.R.; et al. Patterns of microRNA expression characterize stages of human B-cell differentiation. Blood 2009, 113, 4586–4594. [Google Scholar] [CrossRef] [Green Version]
- Schell, S.L.; Bricker, K.N.; Fike, A.J.; Chodisetti, S.B.; Domeier, P.P.; Choi, N.M.; Fasnacht, M.J.; Luckenbill, S.A.; Ziegler, S.F.; Rahman, Z.S.M. Context-Dependent miR-21 Regulation of TLR7-Mediated Autoimmune and Foreign Antigen–Driven Antibody-Forming Cell and Germinal Center Responses. J. Immunol. 2021, 206, 2803–2818. [Google Scholar] [CrossRef]
- Medina, P.P.; Nolde, M.; Slack, F.J. OncomiR addiction in an in vivo model of microRNA-21-induced pre-B-cell lymphoma. Nature 2010, 467, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Shao, Q.; Li, C.; Liu, H.; Li, J.; Wang, Y.; Song, W.; Li, L.; Wang, G.; Shao, Z.; et al. Effects of microRNA-21 on apoptosis by regulating the expression of PTEN in diffuse large B-cell lymphoma. Medicine 2017, 96, e7952. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Song, G.; Chen, L.; Nie, Z.; He, B.; Pan, Y.; Xu, Y.; Li, R.; Gao, T.; Cho, W.C.; et al. Inhibition of miR-21 Induces Biological and Behavioral Alterations in Diffuse Large B-Cell Lymphoma. Acta Haematol. 2013, 130, 87–94. [Google Scholar] [CrossRef]
- Bai, H.; Wei, J.; Deng, C.; Yang, X.; Wang, C.; Xu, R. MicroRNA-21 regulates the sensitivity of diffuse large B-cell lymphoma cells to the CHOP chemotherapy regimen. Int. J. Hematol. 2013, 97, 223–231. [Google Scholar] [CrossRef]
- de Yébenes, V.G.; Bartolomé-Izquierdo, N.; Nogales-Cadenas, R.; Pérez-Durán, P.; Mur, S.M.; Martínez, N.; Di Lisio, L.; Robbiani, D.F.; Pascual-Montano, A.; Cañamero, M.; et al. miR-217 is an oncogene that enhances the germinal center reaction. Blood 2014, 124, 229–239. [Google Scholar] [CrossRef]
- Zauli, G.; Voltan, R.; di Iasio, M.G.; Bosco, R.; Melloni, E.; Sana, M.E.; Secchiero, P. miR-34a induces the downregulation of both E2F1 and B-Myb oncogenes in leukemic cells. Clin. Cancer Res. 2011, 17, 2712–2724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, V.J.; Cogliatti, S.B.; Imig, J.; Renner, C.; Neuenschwander, S.; Rehrauer, H.; Schlapbach, R.; Dirnhofer, S.; Tzankov, A.; Müller, A. Myc-mediated repression of microRNA-34a promotes high-grade transformation of B-cell lymphoma by dysregulation of FoxP1. Blood 2011, 117, 6227–6236. [Google Scholar] [CrossRef]
- Rizzo, M.; Mariani, L.; Cavallini, S.; Simili, M.; Rainaldi, G. The over-expression of miR-34a fails to block DoHH2 lymphoma cell proliferation by reducing p53 via c-MYC down-regulation. Nucleic Acid Ther. 2012, 22, 283–288. [Google Scholar] [CrossRef]
- Marques, S.C.; Ranjbar, B.; Laursen, M.B.; Falgreen, S.; Bilgrau, A.E.; Bødker, J.S.; Jørgensen, L.K.; Primo, M.N.; Schmitz, A.; Ettrup, M.S.; et al. High miR-34a expression improves response to doxorubicin in diffuse large B-cell lymphoma. Exp. Hematol. 2016, 44, 238–246.e2. [Google Scholar] [CrossRef] [PubMed]
- Boysen, J.; Sinha, S.; Price-Troska, T.; Warner, S.L.; Bearss, D.J.; Viswanatha, D.; Shanafelt, T.D.; Kay, N.E.; Ghosh, A.K. The tumor suppressor axis p53/miR-34a regulates Axl expression in B-cell chronic lymphocytic leukemia: Implications for therapy in p53-defective CLL patients. Leukemia 2014, 28, 451–455. [Google Scholar] [CrossRef] [Green Version]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc. Natl. Acad. Sci. USA 2005, 102, 13944–13949. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, M.; Bottoni, A.; Shimizu, M.; Spizzo, R.; Nicoloso, M.S.; Rossi, S.; Barbarotto, E.; Cimmino, A.; Adair, B.; Wojcik, S.E.; et al. Association of a microRNA/TP53 feedback circuitry with pathogenesis and outcome of B-cell chronic lymphocytic leukemia. JAMA 2011, 305, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, X.; Lin, J.; Lwin, T.; Wright, G.; Moscinski, L.; Dalton, W.; Seto, E.; Wright, K.; Sotomayor, E. Myc represses miR-15a/miR-16-1 expression through recruitment of HDAC3 in mantle cell and other non-Hodgkin B-cell lymphomas. Oncogene 2012, 31, 3002–3008. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Calado, D.P.; Galler, G.; Thai, T.-H.; Patterson, H.C.; Wang, J.; Rajewsky, N.; Bender, T.P.; Rajewsky, K. MiR-150 Controls B Cell Differentiation by Targeting the Transcription Factor c-Myb. Cell 2007, 131, 146–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Wang, S.; Mayr, C.; Bartel, D.P.; Lodish, H.F. miR-150, a microRNA expressed in mature B and T cells, blocks early B cell development when expressed prematurely. Proc. Natl. Acad. Sci. USA 2007, 104, 7080–7085. [Google Scholar] [CrossRef] [Green Version]
- Musilova, K.; Devan, J.; Cerna, K.; Seda, V.; Pavlasova, G.; Sharma, S.; Oppelt, J.; Pytlik, R.; Prochazka, V.; Prouzova, Z.; et al. miR-150 downregulation contributes to the high-grade transformation of follicular lymphoma by upregulating FOXP1 levels. Blood 2018, 132, 2389–2400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Zou, Y.; Lin, L.; Ma, X.; Zheng, R. miR-101 regulates the cell proliferation and apoptosis in diffuse large B-cell lymphoma by targeting MEK1 via regulation of the ERK/MAPK signaling pathway. Oncol. Rep. 2019, 41, 377–386. [Google Scholar] [CrossRef]
- Tayari, M.M.; Winkle, M.; Kortman, G.; Sietzema, J.; de Jong, D.; Terpstra, M.; Mestdagh, P.; Kroese, F.G.; Visser, L.; Diepstra, A. Long noncoding RNA expression profiling in normal B-cell subsets and Hodgkin lymphoma reveals hodgkin and reed-sternberg cell–specific long noncoding RNAs. Am. J. Pathol. 2016, 186, 2462–2472. [Google Scholar] [CrossRef] [Green Version]
- Brazão, T.F.; Johnson, J.S.; Müller, J.; Heger, A.; Ponting, C.P.; Tybulewicz, V.L. Long noncoding RNAs in B-cell development and activation. Blood J. Am. Soc. Hematol. 2016, 128, e10–e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petri, A.; Dybkaer, K.; Bøgsted, M.; Thrue, C.A.; Hagedorn, P.H.; Schmitz, A.; Bødker, J.S.; Johnsen, H.E.; Kauppinen, S. Long noncoding RNA expression during human B-cell development. PLoS ONE 2015, 10, e0138236. [Google Scholar] [CrossRef] [Green Version]
- Casero, D.; Sandoval, S.; Seet, C.S.; Scholes, J.; Zhu, Y.; Ha, V.L.; Luong, A.; Parekh, C.; Crooks, G.M. Long non-coding RNA profiling of human lymphoid progenitor cells reveals transcriptional divergence of B cell and T cell lineages. Nat. Immunol. 2015, 16, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.B.; He, X.; Huang, Y.F.; Wu, Q.N.; Zhou, Y.C.; Hao, D.J. Long Noncoding RNA CRNDE Promotes Multiple Myeloma Cell Growth by Suppressing miR-451. Oncol. Res. 2017, 25, 1207–1214. [Google Scholar] [CrossRef]
- Ellis, B.C.; Graham, L.D.; Molloy, P.L. CRNDE, a long non-coding RNA responsive to insulin/IGF signaling, regulates genes involved in central metabolism. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 372–386. [Google Scholar] [CrossRef] [Green Version]
- Verma-Gaur, J.; Torkamani, A.; Schaffer, L.; Head, S.R.; Schork, N.J.; Feeney, A.J. Noncoding transcription within the Igh distal VH region at PAIR elements affects the 3D structure of the Igh locus in pro-B cells. Proc. Natl. Acad. Sci. USA 2012, 109, 17004–17009. [Google Scholar] [CrossRef] [Green Version]
- Syrett, C.M.; Sindhava, V.; Hodawadekar, S.; Myles, A.; Liang, G.; Zhang, Y.; Nandi, S.; Cancro, M.; Atchison, M.; Anguera, M.C. Loss of Xist RNA from the inactive X during B cell development is restored in a dynamic YY1-dependent two-step process in activated B cells. PLoS Genet. 2017, 13, e1007050. [Google Scholar] [CrossRef] [Green Version]
- Ebralidze, A.K.; Guibal, F.C.; Steidl, U.; Zhang, P.; Lee, S.; Bartholdy, B.; Jorda, M.A.; Petkova, V.; Rosenbauer, F.; Huang, G. PU.1 expression is modulated by the balance of functional sense and antisense RNAs regulated by a shared cis-regulatory element. Genes Dev. 2008, 22, 2085–2092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeKoter, R.P.; Singh, H. Regulation of B lymphocyte and macrophage development by graded expression of PU. 1. Science 2000, 288, 1439–1441. [Google Scholar] [CrossRef] [PubMed]
- Blume, C.J.; Hotz-Wagenblatt, A.; Hüllein, J.; Sellner, L.; Jethwa, A.; Stolz, T.; Slabicki, M.; Lee, K.; Sharathchandra, A.; Benner, A.; et al. p53-dependent non-coding RNA networks in chronic lymphocytic leukemia. Leukemia 2015, 29, 2015–2023. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, M.; Xu, H.; Wang, Y.; Li, Z.; Chang, Y.; Wang, X.; Fu, X.; Zhou, Z.; Yang, S.; et al. Discovery and validation of the tumor-suppressive function of long noncoding RNA PANDA in human diffuse large B-cell lymphoma through the inactivation of MAPK/ERK signaling pathway. Oncotarget 2017, 8, 72182–72196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, W.; Wu, J.; Feng, J. Long noncoding RNA HULC predicts poor clinical outcome and represents pro-oncogenic activity in diffuse large B-cell lymphoma. Biomed. Pharmacother. 2016, 79, 188–193. [Google Scholar] [CrossRef]
- Shi, X.; Cui, Z.; Liu, X.; Wu, S.; Wu, Y.; Fang, F.; Zhao, H. LncRNA FIRRE is activated by MYC and promotes the development of diffuse large B-cell lymphoma via Wnt/β-catenin signaling pathway. Biochem. Biophys. Res. Commun. 2019, 510, 594–600. [Google Scholar] [CrossRef]
- Song, Y.; Gao, F.; Peng, Y.; Yang, X. Long non-coding RNA DBH-AS1 promotes cancer progression in diffuse large B-cell lymphoma by targeting FN1 via RNA-binding protein BUD13. Cell Biol. Int. 2020, 44, 1331–1340. [Google Scholar] [CrossRef]
- Cheng, H.; Yan, Z.; Wang, X.; Cao, J.; Chen, W.; Qi, K.; Zhou, D.; Xia, J.; Qi, N.; Li, Z.; et al. Downregulation of long non-coding RNA TUG1 suppresses tumor growth by promoting ubiquitination of MET in diffuse large B-cell lymphoma. Mol. Cell. Biochem. 2019, 461, 47–56. [Google Scholar] [CrossRef]
- Yan, Y.; Han, J.; Li, Z.; Yang, H.; Sui, Y.; Wang, M. Elevated RNA expression of long non-coding HOTAIR promotes cell proliferation and predicts a poor prognosis in patients with diffuse large B cell lymphoma. Mol. Med. Rep. 2016, 13, 5125–5131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, E.J.; Kim, S.H.; Yang, W.I.; Ko, Y.H.; Yoon, S.O. Long Non-coding RNA HOTAIR Expression in Diffuse Large B-Cell Lymphoma: In Relation to Polycomb Repressive Complex Pathway Proteins and H3K27 Trimethylation. J. Pathol. Transl. Med. 2016, 50, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, L.; Mathur, R.; Braun, F.K.; Wise, J.F.; Berkova, Z.; Neelapu, S.; Kwak, L.W.; Samaniego, F. FAS-antisense 1 lncRNA and production of soluble versus membrane Fas in B-cell lymphoma. Leukemia 2014, 28, 2376–2387. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sehgal, L.; Jain, N.; Khashab, T.; Mathur, R.; Samaniego, F. LncRNA MALAT1 promotes development of mantle cell lymphoma by associating with EZH2. J. Transl. Med. 2016, 14, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.-J.; Chai, Y.; Guo, X.-J.; Chu, S.-L.; Zhang, L.-S. The effects of the long non-coding RNA MALAT-1 regulated autophagy-related signaling pathway on chemotherapy resistance in diffuse large B-cell lymphoma. Biomed. Pharmacother. 2017, 89, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Trimarchi, T.; Bilal, E.; Ntziachristos, P.; Fabbri, G.; Dalla-Favera, R.; Tsirigos, A.; Aifantis, I. Genome-wide Mapping and Characterization of Notch-Regulated Long Noncoding RNAs in Acute Leukemia. Cell 2014, 158, 593–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, W.; Feng, J. Long noncoding RNA LUNAR1 associates with cell proliferation and predicts a poor prognosis in diffuse large B-cell lymphoma. Biomed. Pharmacother. 2016, 77, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Florian, A.K.; Reschke, M.; Ruocco, A.; Ng, C.; Chapuy, B.; Léopold, V.; Sjoberg, M.; Thomas, M.K.; Verma, A.; Ala, U.; et al. The BRAF Pseudogene Functions as a Competitive Endogenous RNA and Induces Lymphoma In Vivo. Cell 2015, 161, 319–332. [Google Scholar] [CrossRef] [Green Version]
- Qian, C.-S.; Li, L.-J.; Huang, H.-W.; Yang, H.-F.; Wu, D.-P. MYC-regulated lncRNA NEAT1 promotes B cell proliferation and lymphomagenesis via the miR-34b-5p-GLI1 pathway in diffuse large B-cell lymphoma. Cancer Cell Int. 2020, 20, 87. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Su, L.; Jiang, J. Long Non-Coding RNA Paternally Expressed Imprinted Gene 10 (PEG10) Elevates Diffuse Large B-Cell Lymphoma Progression by Regulating Kinesin Family Member 2A (KIF2A) via Targeting MiR-101-3p. Med. Sci. Monit. 2020, 26, e922810. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Li, Y.; Zheng, W.; Liu, Q.Q.; Zhang, X.L.; Liu, J.L.; Zhang, S.; Sheng, Y.X.; Fan, C.B.; Zhang, W.L. LncRNA PCAT1 enhances cell proliferation, migration and invasion by miR-508-3p/NFIB axis in diffuse large B-cell lymphoma. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 2567–2576. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Du, Y.; Shang, J.; Zhang, D.; Dong, X.; Chen, H. Knockdown of UCA1 restrains cell proliferation and metastasis of diffuse large B-cell lymphoma by counteracting miR-331-3p expression. Oncol. Lett. 2021, 21, 39. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.M.; Lian, G.Y.; Song, Y.; Huang, Y.F.; Gong, Y. LncRNA MALAT1 promotes tumorigenesis and immune escape of diffuse large B cell lymphoma by sponging miR-195. Life Sci. 2019, 231, 116335. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.F.; Shen, J.X.; Hou, Z.W.; Chen, S.Y.; Su, Y.Z.; Fang, J.L. lncRNA FOXP4-AS1 predicts poor prognosis and accelerates the progression of mantle cell lymphoma through the miR-423-5p/NACC1 pathway. Oncol. Rep. 2021, 45, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, B.; Lu, C.-L.; Wang, J.-Y.; Gao, M.; Gao, W. LncRNA LINC01857 promotes cell growth and diminishes apoptosis via PI3K/mTOR pathway and EMT process by regulating miR-141-3p/MAP4K4 axis in diffuse large B-cell lymphoma. Cancer Gene Ther. 2020. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lin, Y.; Song, X.; Wu, D. LINC00857 contributes to proliferation and lymphomagenesis by regulating miR-370-3p/CBX3 axis in diffuse large B-cell lymphoma. Carcinogenesis 2021, 42, 733–741. [Google Scholar] [CrossRef]
- Fu, D.-W.; Liu, A.-C. LncRNA SBF2-AS1 Promotes Diffuse Large B-Cell Lymphoma Growth by Regulating FGFR2 via Sponging miR-494-3p. Cancer Manag. Res. 2021, 13, 571–578. [Google Scholar] [CrossRef]
- Xu, H.; Yu, X.; Yang, Z.; Song, Q.; Cheng, S.; He, Z.; Dai, L. PAX5-activated lncRNA ARRDC1-AS1 accelerates the autophagy and progression of DLBCL through sponging miR-2355-5p to regulate ATG5. Life Sci. 2021, 119932. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jia, Z.; Zhao, H.; Liu, X.; Luo, J.; Cui, G.; Kong, X. TUC338 Promotes Diffuse Large B Cell Lymphoma Growth via Regulating EGFR/PI3K/AKT Signaling Pathway. J. Oncol. 2021, 2021, 5593720. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Wei, Y.; Wei, X.; Feng, R. LINC00908 Promotes Diffuse Large B-Cell Lymphoma Development by Down-Regulating miR-671-5p. Cancer Manag. Res. 2021, 13, 3589–3599. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.C.; Jiao, Y.; Zhang, Y.Y.; Ning, J.; Zhang, Y.R.; Xu, J.; Wei, W.; Kang-Sheng, G. Lnc SMAD5-AS1 as ceRNA inhibit proliferation of diffuse large B cell lymphoma via Wnt/β-catenin pathway by sponging miR-135b-5p to elevate expression of APC. Cell Death Dis. 2019, 10, 252. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Xu, H.; Yang, Y.; Zhao, D.; Wen, Y.; Lv, C.; Qiu, H.; Wang, C. The regulation of miR-320a/XBP1 axis through LINC00963 for endoplasmic reticulum stress and autophagy in diffuse large B-cell lymphoma. Cancer Cell Int. 2021, 21, 305. [Google Scholar] [CrossRef]
- Chen, L.Y.; Zhang, X.M.; Han, B.Q.; Dai, H.B. Long Noncoding RNA SNHG12 Indicates the Prognosis and Accelerates Tumorigenesis of Diffuse Large B-Cell Lymphoma Through Sponging microR-195. OncoTargets Ther. 2020, 13, 5563–5574. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, Y.; Zhang, J.; Liu, Y.; Qi, Q. LncRNA SNHG14/miR-5590-3p/ZEB1 positive feedback loop promoted diffuse large B cell lymphoma progression and immune evasion through regulating PD-1/PD-L1 checkpoint. Cell Death Dis. 2019, 10, 731. [Google Scholar] [CrossRef]
- Tian, Y.; Li, L.; Lin, G.; Wang, Y.; Wang, L.; Zhao, Q.; Hu, Y.; Yong, H.; Wan, Y.; Zhang, Y. lncRNA SNHG14 promotes oncogenesis and immune evasion in diffuse large-B-cell lymphoma by sequestering miR-152-3p. Leuk. Lymphoma 2021, 1–15. [Google Scholar] [CrossRef]
- Zhu, Q.; Li, Y.; Guo, Y.; Hu, L.; Xiao, Z.; Liu, X.; Wang, J.; Xu, Q.; Tong, X. Long non-coding RNA SNHG16 promotes proliferation and inhibits apoptosis of diffuse large B-cell lymphoma cells by targeting miR-497-5p/PIM1 axis. J. Cell. Mol. Med. 2019, 23, 7395–7405. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Wang, B.; Wu, Z.; Wu, C.; Ling, J.; Gao, X.; Zeng, H. LncRNA SNHG8 Promotes Proliferation and Inhibits Apoptosis of Diffuse Large B-Cell Lymphoma via Sponging miR-335-5p. Front. Oncol. 2021, 11, 650287. [Google Scholar] [CrossRef] [PubMed]
- De Rubis, G.; Rajeev Krishnan, S.; Bebawy, M. Liquid Biopsies in Cancer Diagnosis, Monitoring, and Prognosis. Trends Pharmacol. Sci. 2019, 40, 172–186. [Google Scholar] [CrossRef]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, T.; Gao, G.; Cao, Y. Long Noncoding RNAs as Novel Biomarkers Have a Promising Future in Cancer Diagnostics. Dis. Markers 2016, 2016, 9085195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrie, C.H.; Gal, S.; Dunlop, H.M.; Pushkaran, B.; Liggins, A.P.; Pulford, K.; Banham, A.H.; Pezzella, F.; Boultwood, J.; Wainscoat, J.S.; et al. Detection of elevated levels of tumour-associated microRNAs in serum of patients with diffuse large B-cell lymphoma. Br. J. Haematol. 2008, 141, 672–675. [Google Scholar] [CrossRef]
- Fang, C.; Zhu, D.-X.; Dong, H.-J.; Zhou, Z.-J.; Wang, Y.-H.; Liu, L.; Fan, L.; Miao, K.-R.; Liu, P.; Xu, W.; et al. Serum microRNAs are promising novel biomarkers for diffuse large B cell lymphoma. Ann. Hematol. 2012, 91, 553–559. [Google Scholar] [CrossRef]
- Bedewy, A.M.L.; Elmaghraby, S.M.; Shehata, A.A.; Kandil, N.S. Prognostic Value of miRNA-155 Expression in B-Cell Non-Hodgkin Lymphoma. Turk. J. Haematol. 2017, 34, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Ahmadvand, M.; Eskandari, M.; Pashaiefar, H.; Yaghmaie, M.; Manoochehrabadi, S.; Khakpour, G.; Sheikhsaran, F.; Montazer Zohour, M. Over expression of circulating miR-155 predicts prognosis in diffuse large B-cell lymphoma. Leuk. Res. 2018, 70, 45–48. [Google Scholar] [CrossRef]
- Sun, R.; Zheng, Z.; Wang, L.; Cheng, S.; Shi, Q.; Qu, B.; Fu, D.; Leboeuf, C.; Zhao, Y.; Ye, J.; et al. A novel prognostic model based on four circulating miRNA in diffuse large B-cell lymphoma: Implications for the roles of MDSC and Th17 cells in lymphoma progression. Mol. Oncol. 2021, 15, 246–261. [Google Scholar] [CrossRef]
- Inada, K.; Okoshi, Y.; Cho, Y.; Saito, H.; Iijima, T.; Hori, M.; Kojima, H. Availability of Circulating microRNAs as a Biomarker for the Early Diagnosis of Diffuse Large B-Cell Lymphoma. Blood 2014, 124, 2988. [Google Scholar] [CrossRef]
- Beheshti, A.; Stevenson, K.; Vanderburg, C.; Ravi, D.; McDonald, J.T.; Christie, A.L.; Shigemori, K.; Jester, H.; Weinstock, D.M.; Evens, A.M. Identification of Circulating Serum Multi-MicroRNA Signatures in Human DLBCL Models. Sci. Rep. 2019, 9, 17161. [Google Scholar] [CrossRef]
- Khare, D.; Goldschmidt, N.; Bardugo, A.; Gur-Wahnon, D.; Ben-Dov, I.Z.; Avni, B. Plasma microRNA profiling: Exploring better biomarkers for lymphoma surveillance. PLoS ONE 2017, 12, e0187722. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Wang, H.; Chen, H.; Liu, S.; Lu, H.; Kong, D.; Huang, X.; Kong, Q.; Lu, Z. Clinical significance and detection of microRNA-21 in serum of patients with diffuse large B-cell lymphoma in Chinese population. Eur. J. Haematol. 2014, 92, 407–412. [Google Scholar] [CrossRef]
- Cui, Q.; Vari, F.; Cristino, A.S.; Salomon, C.; Rice, G.E.; Sabdia, M.B.; Guanzon, D.; Palma, C.; Mathew, M.; Talaulikar, D.; et al. Circulating cell-free miR-494 and miR-21 are disease response biomarkers associated with interim-positron emission tomography response in patients with diffuse large B-cell lymphoma. Oncotarget 2018, 9, 34644–34657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Fu, R.; Yang, L.; Tu, W. miR-21 expression predicts prognosis in diffuse large B-cell lymphoma. Int. J. Clin. Exp. Pathol. 2015, 8, 15019–15024. [Google Scholar]
- Marchesi, F.; Regazzo, G.; Palombi, F.; Terrenato, I.; Sacconi, A.; Spagnuolo, M.; Donzelli, S.; Marino, M.; Ercolani, C.; Di Benedetto, A.; et al. Serum miR-22 as potential non-invasive predictor of poor clinical outcome in newly diagnosed, uniformly treated patients with diffuse large B-cell lymphoma: An explorative pilot study. J. Exp. Clin. Cancer Res. 2018, 37, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, G.; Gu, L.; Li, J.; Tang, Z.; Liu, H.; Chen, B.; Sun, X.; He, B.; Pan, Y.; Wang, S.; et al. Serum microRNA expression profiling predict response to R-CHOP treatment in diffuse large B cell lymphoma patients. Ann. Hematol. 2014, 93, 1735–1743. [Google Scholar] [CrossRef]
- Ohyashiki, K.; Umezu, T.; Yoshizawa, S.-i.; Ito, Y.; Ohyashiki, M.; Kawashima, H.; Tanaka, M.; Kuroda, M.; Ohyashiki, J.H. Clinical Impact of Down-Regulated Plasma miR-92a Levels in Non-Hodgkin’s Lymphoma. PLoS ONE 2011, 6, e16408. [Google Scholar] [CrossRef] [Green Version]
- Yuan, W.X.; Gui, Y.X.; Na, W.N.; Chao, J.; Yang, X. Circulating microRNA-125b and microRNA-130a expression profiles predict chemoresistance to R-CHOP in diffuse large B-cell lymphoma patients. Oncol. Lett. 2016, 11, 423–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.; Quan, L.; Liu, A. Identification of key microRNAs associated with diffuse large B-cell lymphoma by analyzing serum microRNA expressions. Gene 2018, 642, 205–211. [Google Scholar] [CrossRef]
- Jørgensen, S.; Paulsen, I.W.; Hansen, J.W.; Tholstrup, D.; Hother, C.; Sørensen, E.; Petersen, M.S.; Nielsen, K.R.; Rostgaard, K.; Larsen, M.A.H.; et al. The value of circulating microRNAs for early diagnosis of B-cell lymphoma: A case-control study on historical samples. Sci. Rep. 2020, 10, 9637. [Google Scholar] [CrossRef] [PubMed]
- Filip, A.A.; Grenda, A.; Popek, S.; Koczkodaj, D.; Michalak-Wojnowska, M.; Budzyński, M.; Wąsik-Szczepanek, E.; Zmorzyński, S.; Karczmarczyk, A.; Giannopoulos, K. Expression of circulating miRNAs associated with lymphocyte differentiation and activation in CLL—another piece in the puzzle. Ann. Hematol. 2017, 96, 33–50. [Google Scholar] [CrossRef] [Green Version]
- Peng, W.; Fan, H.; Wu, G.; Wu, J.; Feng, J. Upregulation of long noncoding RNA PEG10 associates with poor prognosis in diffuse large B cell lymphoma with facilitating tumorigenicity. Clin. Exp. Med. 2016, 16, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Meng, H.; Zhao, B.; Wang, Y. FOXM1-induced upregulation of lncRNA OR3A4 promotes the progression of diffuse large B-cell lymphoma via Wnt/β-catenin signaling pathway. Exp. Mol. Pathol. 2020, 115, 104451. [Google Scholar] [CrossRef]
- Gao, H.-Y.; Wu, B.; Yan, W.; Gong, Z.-M.; Sun, Q.; Wang, H.-H.; Yang, W. Microarray expression profiles of long non-coding RNAs in germinal center-like diffuse large B-cell lymphoma. Oncol. Rep. 2017, 38, 1363–1372. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Zhao, H.; Xu, W.; Bao, S.; Cheng, L.; Sun, J. Discovery and validation of immune-associated long non-coding RNA biomarkers associated with clinically molecular subtype and prognosis in diffuse large B cell lymphoma. Mol. Cancer 2017, 16, 16. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Fang, S.; Liu, Y.; Li, X.; Liao, S.; Chen, J.; Liu, J.; Zhao, L.; Li, H.; Zhou, W.; et al. The long non-coding RNA NONHSAG026900 predicts prognosis as a favorable biomarker in patients with diffuse large B-cell lymphoma. Oncotarget 2017, 8, 34374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.; Jiang, L.; Tseng, K.F.; Liu, Y.; Zhang, X.; Dong, R.; Lu, Z.; Wang, X. Aberrant NEAT1_1 expression may be a predictive marker of poor prognosis in diffuse large B cell lymphoma. Cancer Biomark. 2018, 23, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Dousti, F.; Shahrisa, A.; Ansari, H.; Hajjari, M.; Tahmasebi Birgani, Y.; Mohammadiasl, J.; Tahmasebi Birgani, M. Long non-coding RNAs expression levels in diffuse large B-cell lymphoma: An in silico analysis. Pathol. Res. Pract. 2018, 214, 1462–1466. [Google Scholar] [CrossRef]
- Senousy, M.A.; El-Abd, A.M.; Abdel-Malek, R.R.; Rizk, S.M. Circulating long non-coding RNAs HOTAIR, Linc-p21, GAS5 and XIST expression profiles in diffuse large B-cell lymphoma: Association with R-CHOP responsiveness. Sci. Rep. 2021, 11, 2095. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Cheng, L.; Shi, H.; Zhang, Z.; Zhao, H.; Wang, Z.; Zhou, M. A potential panel of six-long non-coding RNA signature to improve survival prediction of diffuse large-B-cell lymphoma. Sci. Rep. 2016, 6, 27842. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Li, H.; Guo, Y.; Luo, Y.; Li, H.; Xu, Y.; Deng, J.; Sun, B. A pilot study of long noncoding RNA expression profiling by microarray in follicular lymphoma. Gene 2016, 577, 132–139. [Google Scholar] [CrossRef]
- Roisman, A.; Castellano, G.; Navarro, A.; Gonzalez-Farre, B.; Pérez-Galan, P.; Esteve-Codina, A.; Dabad, M.; Heath, S.; Gut, M.; Bosio, M.; et al. Differential expression of long non-coding RNAs are related to proliferation and histological diversity in follicular lymphomas. Br. J. Haematol. 2019, 184, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lu, P.; Du, H.; Zhang, L. LINK-A lncRNA Promotes Proliferation and Inhibits Apoptosis of Mantle Cell Lymphoma Cell by Upregulating Survivin. Med. Sci. Monit. 2019, 25, 365–370. [Google Scholar] [CrossRef]
- Tang, X.; Long, Y.; Xu, L.; Yan, X. LncRNA MORT Inhibits Cancer Cell Proliferation and Promotes Apoptosis in Mantle Cell Lymphoma by Upregulating miRNA-16. Cancer Manag. Res. 2020, 12, 2119–2125. [Google Scholar] [CrossRef] [Green Version]
- Isin, M.; Ozgur, E.; Cetin, G.; Erten, N.; Aktan, M.; Gezer, U.; Dalay, N. Investigation of circulating lncRNAs in B-cell neoplasms. Clin. Chim. Acta 2014, 431, 255–259. [Google Scholar] [CrossRef]
- Beheshti, A.; Vanderburg, C.; McDonald, J.T.; Ramkumar, C.; Kadungure, T.; Zhang, H.; Gartenhaus, R.B.; Evens, A.M. A Circulating microRNA Signature Predicts Age-Based Development of Lymphoma. PLoS ONE 2017, 12, e0170521. [Google Scholar] [CrossRef]
- Leich, E.; Zamo, A.; Horn, H.; Haralambieva, E.; Puppe, B.; Gascoyne, R.D.; Chan, W.C.; Braziel, R.M.; Rimsza, L.M.; Weisenburger, D.D.; et al. MicroRNA profiles of t(14;18)-negative follicular lymphoma support a late germinal center B-cell phenotype. Blood 2011, 118, 5550–5558. [Google Scholar] [CrossRef]
- Gebauer, N.; Gollub, W.; Stassek, B.; Bernard, V.; Rades, D.; Feller, A.C.; Thorns, C. MicroRNA signatures in subtypes of follicular lymphoma. Anticancer Res. 2014, 34, 2105–2111. [Google Scholar] [PubMed]
- Di Lisio, L.; Sánchez-Beato, M.; Gómez-López, G.; Rodríguez, M.E.; Montes-Moreno, S.; Mollejo, M.; Menárguez, J.; Martínez, M.A.; Alves, F.J.; Pisano, D.G.; et al. MicroRNA signatures in B-cell lymphomas. Blood Cancer J. 2012, 2, e57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, A.; Jiang, Y.; Du, W.; Fairchild, L.; Melnick, A.; Elemento, O. Transcriptome sequencing reveals thousands of novel long non-coding RNAs in B cell lymphoma. Genome Med. 2015, 7, 110. [Google Scholar] [CrossRef] [Green Version]
- Qi, P.; Zhou, X.-y.; Du, X. Circulating long non-coding RNAs in cancer: Current status and future perspectives. Mol. Cancer 2016, 15, 39. [Google Scholar] [CrossRef] [Green Version]
- Diez-Fraile, A.; Ceulaer, J.D.; Derpoorter, C.; Spaas, C.; Backer, T.D.; Lamoral, P.; Abeloos, J.; Lammens, T. Circulating Non-Coding RNAs in Head and Neck Cancer: Roles in Diagnosis, Prognosis, and Therapy Monitoring. Cells 2021, 10, 48. [Google Scholar] [CrossRef]
- Fan, Z.; Wang, X.; Li, P.; Mei, C.; Zhang, M.; Zhao, C. Overexpression of lncRNA GATA6-AS inhibits cancer cell proliferation in mantle cell lymphoma by downregulating GLUT1. Oncol. Lett. 2019, 18, 2443–2447. [Google Scholar] [CrossRef]
- Peng, W.; Wu, J.; Feng, J. LincRNA-p21 predicts favorable clinical outcome and impairs tumorigenesis in diffuse large B cell lymphoma patients treated with R-CHOP chemotherapy. Clin. Exp. Med. 2017, 17, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Adams, H.J.A.; Nievelstein, R.A.J.; Kwee, T.C. Prognostic value of interim and end-of-treatment FDG-PET in follicular lymphoma: A systematic review. Ann. Hematol. 2016, 95, 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouvy, C.; Wannez, A.; George, F.; Graux, C.; Chatelain, C.; Dogné, J.M. Circulating MicroRNAs as Biomarkers in Diffuse Large B-cell Lymphoma: A Pilot Prospective Longitudinal Clinical Study. Biomark. Cancer 2018, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fajardo-Ramirez, O.R.; Villela, L.; Campa-Carranza, J.N.; Perez-Maya, A.A.; Borrego-Soto, G.; Wah-Suarez, M.I.; Rodríguez-Sánchez, I.P.; Zapata-Morin, P.A.; Ortiz-Lopez, R.; Treviño, V.M.; et al. miRNA signature associated with R–CHOP refractoriness in patients diagnosed with diffuse large B cell lymphoma. Non-Coding RNA Res. 2020, 5, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Karstensen, K.T.; Schein, A.; Petri, A.; Bøgsted, M.; Dybkær, K.; Uchida, S.; Kauppinen, S. Long Non-Coding RNAs in Diffuse Large B-Cell Lymphoma. Non-Coding RNA 2021, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Mourtada-Maarabouni, M.; Williams, G.T. Role of GAS5 Noncoding RNA in Mediating the Effects of Rapamycin and Its Analogues on Mantle Cell Lymphoma Cells. Clin. Lymphoma Myeloma Leuk. 2014, 14, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Gong, M.; Li, Z. Knockdown of lncRNA MCM3AP-AS1 Attenuates Chemoresistance of Burkitt Lymphoma to Doxorubicin Treatment via Targeting the miR-15a/EIF4E Axis. Cancer Manag. Res. 2020, 12, 5845–5855. [Google Scholar] [CrossRef] [PubMed]
- Study Biomarkers in Samples From Patients With B-Cell Acute Lymphoblastic Leukemia. Available online: https://ClinicalTrials.gov/show/NCT01505699 (accessed on 3 June 2021).
- Studying Biomarkers in Cell Samples From Patients With Acute Myeloid Leukemia. Available online: https://ClinicalTrials.gov/show/NCT01057199 (accessed on 3 June 2021).
- Circulating microRNAs as Disease Markers in Pediatric Cancers. Available online: https://ClinicalTrials.gov/show/NCT01541800 (accessed on 3 June 2021).
- Identification of Hematological Malignancies and Therapy Predication Using microRNAs as a Diagnostic Tool. Available online: https://ClinicalTrials.gov/show/NCT02791217 (accessed on 3 June 2021).
- Mechanisms of Action of Photo(Chemo)Therapy in Skin Diseases. Available online: https://ClinicalTrials.gov/show/NCT03340155 (accessed on 3 June 2021).
- Treatment Resistance Related With Gene Expression Profile of Diffuse Large B-cell Lymphoma. Available online: https://ClinicalTrials.gov/show/NCT01606605 (accessed on 3 June 2021).
- Qian, Y.; Shi, L.; Luo, Z. Long Non-coding RNAs in Cancer: Implications for Diagnosis, Prognosis, and Therapy. Front. Med. 2020, 7, 612393. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov. 2021. Available online: https://clinicaltrials.gov/ct2/about-site/new (accessed on 3 June 2021).
- Pal, R.P.; Maitra, N.U.; Mellon, J.K.; Khan, M.A. Defining prostate cancer risk before prostate biopsy. Urol. Oncol. 2013, 31, 1408–1418. [Google Scholar] [CrossRef]
- Immunophenotyping and Xist Gene in AML. Available online: https://ClinicalTrials.gov/show/NCT04288739 (accessed on 3 June 2021).




{kind=link}
{kind=link}
{kind=link}
| NHL | miRNA | Expression | Biomarker Utility | Source Material | Refs. |
|---|---|---|---|---|---|
| DLBCL | miR-155 | Upregulated | Diagnostic | Serum | [110,111] |
| Subclassification | Serum | [112] | |||
| Prognostic of OS, PFS and RFS | Plasma, serum | [113,114] | |||
| miR-210 | Upregulated | Diagnostic | Serum | [110,115] | |
| let-7b/c | Upregulated | Diagnostic | Serum | [116] | |
| miR-15a | Upregulated | Diagnostic | Serum | [111,115,116] | |
| miR-16-1 | Upregulated | Diagnostic | Serum | [111] | |
| miR-18a | Upregulated | Diagnostic | Serum | [116] | |
| miR-20a/b | Upregulated | Prognostic of OS | Serum | [117] | |
| miR-21 | Upregulated | Diagnostic | Serum | [110,115] | |
| Subclassification | Serum | [118] | |||
| Monitoring | Plasma | [119] | |||
| Prognostic of OS, PFS and RFS | Serum | [110,114,118,120] | |||
| miR-22 | Upregulated | Prognostic of PFS | Serum | [121] | |
| miR-24 | Upregulated | Diagnostic | Serum | [116] | |
| miR-28 | Downregulated | Prognostic of OS, PFS and RFS | Serum | [114] | |
| miR-29c | Upregulated | Diagnostic | Serum | [111] | |
| miR-33a | Downregulated | Prognostic of RFS | Serum | [122] | |
| miR-34 | Downregulated | Diagnostic | Serum | [111] | |
| miR-92a | Downregulated | Diagnostic | Plasma | [123] | |
| Monitoring | |||||
| miR-93 | Upregulated | Prognostic of OS | Serum | [117] | |
| miR-106a/b | Upregulated | Prognostic of OS | Serum | [117] | |
| miR-125b | Upregulation | Prognostic of OS | Serum | [124] | |
| miR-130a | Upregulated | Monitoring | Serum | [124] | |
| miR-130b | upregulation | Prognostic of OS, PFS and RFS | Serum | [114] | |
| miR-181-5p | Downregulated | Subclassification | Serum | [112] | |
| miR-199-5p | Upregulated | Prognostic of OS | Plasma | [113] | |
| miR-224 | Upregulated | Prognostic of RFS | Serum | [122] | |
| miR-323b | Downregulated | Diagnostic | Serum | [125] | |
| miR-326 | Upregulated | Diagnostic | Serum | [126] | |
| miR-375 | Downregulated | Diagnostic | Serum | [126] | |
| miR-431 | Downregulated | Diagnostic | Serum | [125] | |
| miR-455-3p | downregulated | Prognostic of RFS | Serum | [122] | |
| miR-494 | upregulated | Monitoring | Plasma | [119] | |
| miR-520d-3p | Upregulated | Prognostic of RFS | Serum | [122] | |
| miR-1236 | Upregulated | Prognostic of RFS | Serum | [122] | |
| CLL | miR-34a | Upregulated | Diagnostic | Serum | [127] |
| miR-31-5p | Upregulated | Diagnostic | Serum | [127] | |
| miR-150-5p | Upregulated | Diagnostic | Serum | [127] | |
| miR-155-5p | Upregulated | Diagnostic | Serum | [127] | |
| miR-15a-3p | Upregulated | Diagnostic | Serum | [127] | |
| miR-29a-3p | Upregulated | Diagnostic | Serum | [127] |
| NHL | LncRNA | Expression | Biomarker Utility | Source Material | Refs. |
|---|---|---|---|---|---|
| DLBCL | PEG10 | Upregulated | Diagnostic | Tissue Cell lines | [128] |
| Prognostic of OS | |||||
| LUNAR1 | Upregulated | Diagnostic | Tissue Cell lines | [86] | |
| Prognostic of OS and PFS | |||||
| FIRRE | Upregulated | Diagnostic | Tissue Cell lines | [77] | |
| Prognostic of OS | |||||
| HULC | Upregulated | Diagnostic | Tissue Cell lines | [76] | |
| Prognostic of OS and PFS | |||||
| LINC01857 | Upregulated | Diagnostic | Tissue Cell lines | [94] | |
| OR3A4 | Upregulated | Diagnostic | Tissue Cell lines | [129] | |
| Prognostic of OS | |||||
| ENST00000424690 | Upregulated | Diagnostic | Tissue Cell lines | [130] | |
| ENST00000425358 | |||||
| NR_026892 | |||||
| ENST00000464929 | Downregulated | ||||
| ENST00000475089 | |||||
| SubSigLnc-17 | - | Diagnostic | Tissue Cell lines | [131] | |
| Subclassification | |||||
| Prognostic of OS and PFS | |||||
| NONHSAG026900 | Upregulated | Diagnostic | Tissue Cell lines | [132] | |
| Prognostic of OS and PFS | |||||
| NEAT1_1 | Upregulated | Diagnostic | Tissue Cell lines | [133] | |
| Prognostic of OS | |||||
| GAS5 | Upregulated | Diagnostic | Tissue Cell lines | [134] | |
| MIR17HG | Upregulated | Diagnostic | Tissue Cell lines | ||
| HULC | Upregulation | Diagnostic | Tissue Cell lines | ||
| PCA3 | Upregulated | Diagnostic | Tissue Cell lines | ||
| PANDA | Downregulation | Diagnostic | Plasma Tissue | [75] | |
| Prognostic of OS and RFS | |||||
| TUG1 | Upregulated | Diagnostic | Plasma | [75] | |
| HOTAIR | Upregulated | Diagnostic | Plasma Tissue | [80,135] | |
| Predictive of Treatment response | |||||
| Prognostic of OS | |||||
| XIST | Upregulated | Diagnostic | Plasma | [135] | |
| GAS5 | Downregulated | Diagnostic | Plasma | ||
| Predictive of Treatment response | |||||
| 6-lncRNA signature | - | Prognostic of OS | Tissue | [136] | |
| FL | RP11-625 L16.3 | Upregulated | Diagnostic | Tissue | [137] |
| RP4-694A7.2 | Upregulated | Diagnostic and subclassification | Tissue | [138] | |
| MCL | LINK-A | Upregulated | Diagnostic | Plasma | [139] |
| GATA6-AS | Downregulated | Diagnostic | Plasma | ||
| MALAT1 | Upregulated | Prognostic of OS and DFS | Tissue Cell lines | [83] | |
| FOXP4-AS1 | Upregulated | Prognostic of OS and DFS | Plasma | [93] | |
| MORT | Downregulated | Diagnostic | Plasma | [140] | |
| CLL | lincRNA-p21 | Downregulated | Diagnostic | Plasma | [141] |
| MM | TUG1 | Upregulated | Diagnostic | Plasma | |
| MALAT1 | Downregulated | Diagnostic | Plasma | ||
| HOTAIR | Diagnostic | Plasma | |||
| GAS5 | Diagnostic | Plasma |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, M.; Marques, H.; Teixeira, A.L.; Medeiros, R. Competitive Endogenous RNA Network Involving miRNA and lncRNA in Non-Hodgkin Lymphoma: Current Advances and Clinical Perspectives. Biomedicines 2021, 9, 1934. https://doi.org/10.3390/biomedicines9121934
Fernandes M, Marques H, Teixeira AL, Medeiros R. Competitive Endogenous RNA Network Involving miRNA and lncRNA in Non-Hodgkin Lymphoma: Current Advances and Clinical Perspectives. Biomedicines. 2021; 9(12):1934. https://doi.org/10.3390/biomedicines9121934
Chicago/Turabian StyleFernandes, Mara, Herlander Marques, Ana Luísa Teixeira, and Rui Medeiros. 2021. "Competitive Endogenous RNA Network Involving miRNA and lncRNA in Non-Hodgkin Lymphoma: Current Advances and Clinical Perspectives" Biomedicines 9, no. 12: 1934. https://doi.org/10.3390/biomedicines9121934