
Genomic and Personalized Medicine Approaches for Substance Use Disorders (SUDs) Looking at Genome-Wide Association Studies

 , and
, and
Abstract
:1. Introduction
2. The Genetics of Cannabis Use
3. The Genetics of Cocaine Use
4. The Genetics of Heroin and Opioids Use
5. The Genetics of the Treatment of Substance Use Disorders: A Pharmacogenetics Approach
6. Epigenetics and Addiction: An Overview
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Knopik, V.S.; Marceau, K.; Bidwell, L.C.; Rolan, E. Prenatal substance exposure and offspring development: Does DNA methyl-ation play a role? Neurotoxicol Teratol 2019, 71, 50–63. [Google Scholar] [CrossRef]
- Uffelmann, E.; Huang, Q.Q.; Munung, N.S.; de Vries, J.; Okada, Y.; Martin, A.R.; Martin, H.C.; Lappalainen, T.; Posthuma, D. Genome-wide association studies. Nat Rev Methods Primers 2021, 1, 59. [Google Scholar] [CrossRef]
- Manolio, T.A.; Collins, F.S.; Cox, N.J.; Goldstein, D.B.; Hindorff, L.A.; Hunter, D.J.; McCarthy, M.I.; Ramos, E.M.; Cardon, L.R.; Chakravarti, A.; et al. Visscher, Finding the missing heritability of complex diseases. Nature 2009, 461, 747–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bycroft, C.; Freeman, C.; Petkova, D.; Band, G.; Elliott, L.T.; Sharp, K.; Motyer, A.; Vukcevic, D.; Delaneau, O.; O’Connell, J.; et al. The UK Biobank resource with deep phenotyping and genomic data. Nat. Cell Biol. 2018, 562, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallares, L.F. Searching for solutions to the missing heritability problem. eLife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Burgess, D.J. The TOPMed genomic resource for human health. Nat. Rev. Genet. 2021, 22, 200. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, P.F. The Psychiatric GWAS Consortium: Big Science Comes to Psychiatry. Neuron 2010, 68, 182–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.J.; Wedow, R.; Okbay, A.; Kong, E.; Maghzian, O.; Zacher, M.; Nguyen-Viet, T.A.; Bowers, P.; Sidorenko, J.; Linnér, R.K.; et al. Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nat. Genet. 2018, 50, 1112–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardiñas, A.F.; GERAD1 Consortium; Holmans, P.; Pocklington, A.J.; Escott-Price, V.; Ripke, S.; Carrera, N.; Legge, S.E.; Bishop, S.; Cameron, D.; et al. Publisher Correction: Common schizophrenia alleles are enriched in mutation-intolerant genes and in regions under strong background selection. Nat. Genet. 2019, 51, 1193. [Google Scholar] [CrossRef]
- Welter, D.; MacArthur, J.; Morales, J.; Burdett, A.; Hall, P.; Junkins, H.; Klemm, A.; Flicek, P.; Manolio, T.; Hindorff, L.; et al. The NHGRI GWAS Catalog, a curated resource of SNP-trait associations. Nucleic Acids Res. 2014, 42, D1001–D1006. [Google Scholar] [CrossRef]
- Reich, T.; Edenberg, H.J.; Goate, A.; Williams, J.T.; Rice, J.P.; Van Eerdewegh, P.; Foroud, T.; Hesselbrock, V.; Schuckit, M.A.; Bucholz, K.; et al. Begleiter, Genome-wide search for genes affecting the risk for alcohol dependence. Am. J. Med. Genet 1998, 81, 207–215. [Google Scholar] [CrossRef]
- World Health Organization. World Drug Report 2021: Executive Summary Policy Impications; UNODC: Vienna, Austria, 2021. [Google Scholar]
- Verweij, K.J.; Zietsch, B.P.; Lynskey, M.T.; Medland, S.E.; Neale, M.C.; Martin, N.G.; Boomsma, D.I.; Vink, J.M. Genetic and environmental influences on cannabis use initiation and problematic use: A meta-analysis of twin studies. Addiction 2010, 105, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Kendler, K.S.; Ohlsson, H.; Maes, H.H.; Sundquist, K.; Lichtenstein, P.; Sundquist, J. A population-based Swedish Twin and Sibling Study of cannabis, stimulant and sedative abuse in men. Drug Alcohol Depend 2015, 149, 49–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasman, J.A.; Verweij, K.J.H.; Gerring, Z.; Stringer, S.; Sanchez-Roige, S.; Treur, J.L.; Abdellaoui, A.; Nivard, M.G.; Baselmans, B.M.L.; Ong, J.-S.; et al. GWAS of lifetime cannabis use reveals new risk loci, genetic overlap with psychiatric traits, and a causal effect of schizophrenia liability. Nat. Neurosci. 2018, 21, 1161–1170. [Google Scholar] [CrossRef]
- Brazel, D.M.; Jiang, Y.; Hughey, J.M.; Turcot, V.; Zhan, X.; Gong, J.; Batini, C.; Weissenkampen, J.D.; Liu, M.; Barnes, D.R.; et al. Behaviour, Exome Chip Meta-analysis Fine Maps Causal Variants and Elucidates the Genetic Architecture of Rare Coding Variants in Smoking and Alcohol Use. Biol. Psychiatry 2019, 85, 946–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Roige, S.; Fontanillas, P.; Elson, S.L.; Gray, J.C.; de Wit, H.; MacKillop, J.; Palmer, A.A. Genome-Wide Association Studies of Impulsive Personality Traits (BIS-11 and UPPS-P) and Drug Experimentation in up to 22,861 Adult Research Participants Identify Loci. J Neurosci. 2019, 39, 2562–2572. [Google Scholar]
- Liu, M.; Jiang, Y.; Wedow, R.; Li, Y.; Brazel, D.M.; Chen, F.; Datta, G.; Davila-Velderrain, J.; McGuire, D.; Tian, C.; et al. Psychiatry, Association studies of up to 1.2 million individuals yield new insights into the genetic etiology of tobacco and alcohol use. Nat. Genet. 2019, 51, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Nagel, M.; Jansen, P.R.; Stringer, S.; Watanabe, K.; de Leeuw, C.A.; Bryois, J.; Savage, J.E.; Hammerschlag, A.R.; Skene, N.G.; Muñoz-Manchado, A.B.; et al. Meta-analysis of genome-wide association studies for neuroticism in 449,484 individuals identifies novel genetic loci and pathways. Nat. Genet. 2018, 50, 920–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson Linnér, R.; Mallard, T.T.; Barr, P.B.; Sanchez-Roige, S.; Madole, J.W.; Driver, M.N.; Poore, H.E.; de Vlaming, R.; Grotzinger, A.D.; Tielbeek, J.J.; et al. Collaborators, Multivariate analysis of 1.5 million people identifies genetic associations with traits related to self-regulation and addiction. Nat. Neurosci. 2021, 24, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- Minică, C.C.; Verweij, K.J.H.; van der Most, P.J.; Mbarek, H.; Bernard, M.; van Eijk, K.R.; Lind, P.A.; Liu, M.Z.; Maciejewski, D.F.; Palviainen, T.; et al. Genome-wide association meta-analysis of age at first cannabis use. Addiction 2018, 113, 2073–2086. [Google Scholar] [CrossRef] [Green Version]
- Minică, C.C.; The Genome of the Netherlands Consortium; Dolan, C.V.; Hottenga, J.-J.; Pool, R.; Fedko, I.; Mbarek, H.; Huppertz, C.; Bartels, M.; Boomsma, D.I.; et al. Heritability, SNP- and Gene-Based Analyses of Cannabis Use Initiation and Age at Onset. Behav. Genet. 2015, 45, 503–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, A.; Chou, Y.-L.; Carey, C.E.; Baranger, D.A.A.; Zhang, B.; Sherva, R.; Wetherill, L.; Kapoor, M.; Wang, J.-C.; Bertelsen, S.; et al. Genome-wide association study identifies a novel locus for cannabis dependence. Mol. Psychiatry 2018, 23, 1293–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demontis, D.; Rajagopal, V.M.; Thorgeirsson, T.E.; Als, T.D.; Grove, J.; Leppälä, K.; Gudbjartsson, D.F.; Pallesen, J.; Hjorthøj, C.; Reginsson, G.W.; et al. Børglum, Genome-wide association study implicates CHRNA2 in cannabis use disorder. Nat. Neurosci. 2019, 22, 1066–1074. [Google Scholar] [CrossRef]
- Dudbridge, F.; Pashayan, N.; Yang, J. Predictive accuracy of combined genetic and environmental risk scores. Genet. Epidemiol. 2018, 42, 4–19. [Google Scholar] [CrossRef] [Green Version]
- Hodge, S.E.; Greenberg, D.A. How Can We Explain Very Low Odds Ratios in GWAS? I. Polygenic Models. Hum. Hered. 2016, 81, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Gizer, I.R.; Bizon, C.; Gilder, D.A.; Ehlers, C.L.; Wilhelmsen, K.C. Whole genome sequence study of cannabis dependence in two independent cohorts. Addict. Biol. 2017, 23, 461–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torsten, K.-A.U.; Denmark, T.-M.U. Betsy, UK, The SAGE Handbook of Drug & Alcohol Studies; SAGE Publications Ltd.: New York, NY, USA, 2016. [Google Scholar]
- Gelernter, J.; Yu, Y.; Weiss, R.; Brady, K.; Panhuysen, C.; Yang, B.Z.; Kranzler, H.R.; Farrer, L. Haplotype spanning TTC12 and ANKK1, flanked by the DRD2 and NCAM1 loci, is strongly associated to nicotine dependence in two distinct American populations. Hum. Mol. Genet. 2006, 15, 3498–3507. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.Z.; Han, S.; Kranzler, H.R.; Farrer, L.A.; Gelernter, J. A genomewide linkage scan of cocaine dependence and major de-pressive episode in two populations. Neuropsychopharmacology 2011, 36, 2422–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelernter, J.; Sherva, R.; Koesterer, R.; Almasy, L.; Zhao, H.; Kranzler, H.R.; Farrer, L. Genome-wide association study of cocaine dependence and related traits: FAM53B identified as a risk gene. Mol. Psychiatry 2013, 19, 717–723. [Google Scholar] [CrossRef] [Green Version]
- Kizil, C.; Küchler, B.; Yan, J.J.; Özhan, G.; Moro, E.; Argenton, F.; Brand, M.; Weidinger, G.; Antos, C.L. Simplet/Fam53b is required for Wnt signal transduction by regulating β-catenin nuclear localization. Development 2014, 141, 3529–3539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernàndez-Castillo, N.; Cabana-Domínguez, J.; Soriano, J.; Sànchez-Mora, C.; Roncero, C.; Grau-López, L.; Ros-Cucurull, E.; Daigre, C.; Van Donkelaar, M.M.J.; Franke, B.; et al. Transcriptomic and genetic studies identify NFAT5 as a candidate gene for cocaine dependence. Transl. Psychiatry 2015, 5, e667. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.; Kim, D.; Kim, W.-U. Role of NFAT5 in the Immune System and Pathogenesis of Autoimmune Diseases. Front. Immunol. 2019, 10, 270. [Google Scholar] [CrossRef] [PubMed]
- Huggett, S.B.; Stallings, M.C. Cocaine’omics: Genome-wide and transcriptome-wide analyses provide biological insight into cocaine use and dependence. Addict. Biol. 2020, 25, e12719. [Google Scholar] [CrossRef]
- Sun, J.; Kranzler, H.R.; Gelernter, J.; Bi, J. A genome-wide association study of cocaine use disorder accounting for phenotypic heterogeneity and gene–environment interaction. J. Psychiatry Neurosci. 2020, 45, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Goldman, D.; Oroszi, G.; Ducci, F. The genetics of addictions: Uncovering the genes. Nat. Rev. Genet. 2005, 6, 521–532. [Google Scholar] [CrossRef]
- Jones, J.D.; Luba, R.R.; Vogelman, J.L.; Comer, S.D. Searching for evidence of genetic mediation of opioid withdrawal by opioid receptor gene polymorphisms. Am. J. Addict. 2016, 25, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendler, K.S.; Prescott, C.A.; Myers, J.; Neale, M.C. The structure of genetic and environmental risk factors for common psychiatric and substance use disorders in men and women. Arch. Gen. Psychiatry 2003, 60, 929–937. [Google Scholar] [CrossRef] [Green Version]
- Tsuang, M.T.; Lyons, M.J.; Meyer, J.M.; Doyle, T.; Eisen, S.A.; Goldberg, J.; True, W.; Lin, N.; Toomey, R.; Eaves, L. Co-occurrence of abuse of different drugs in men: The role of drug-specific and shared vulnerabilities. Arch. Gen. Psychiatry 1998, 55, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Kreek, M.J.; Nielsen, D.; Butelman, E.R.; LaForge, K.S. Genetic influences on impulsivity, risk taking, stress responsivity and vulnerability to drug abuse and addiction. Nat. Neurosci. 2005, 8, 1450–1457. [Google Scholar] [CrossRef]
- Merikangas, K.R.; Dierker, L.C.; Szatmari, P. Psychopathology among offspring of parents with substance abuse and/or anxiety disorders: A high-risk study. J. Child Psychol. Psychiatry 1998, 39, 711–720. [Google Scholar]
- Merikangas, K.R.; Mehta, R.L.; Molnar, B.E.; Walters, E.E.; Swendsen, J.D.; Aguilar-Gaziola, S.; Bijl, R.; Borges, G.; Caraveo-Anduaga, J.J.; Dewit, D.J.; et al. Comorbidity of substance use disorders with mood and anxiety disorders: Results of the international consortium in psychiatric epidemiology. Addict. Behav. 1998, 23, 893–907. [Google Scholar] [CrossRef] [Green Version]
- Charbogne, P.; Kieffer, B.L.; Befort, K. 15 years of genetic approaches in vivo for addiction research: Opioid receptor and peptide gene knockout in mouse models of drug abuse. Neuropharmacology 2014, 76, 204–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prom-Wormley, E.C.; Ebejer, J.; Dick, D.M.; Bowers, M.S. The genetic epidemiology of substance use disorder: A review. Drug Alcohol Depend. 2017, 180, 241–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachman, H.M.; Fann, C.S.; Bartzis, M.; Evgrafov, O.V.; Rosenthal, R.N.; Nunes, E.V.; Miner, C.; Santana, M.; Gaffney, J.; Riddick, A.; et al. Genomewide suggestive linkage of opioid dependence to chromosome 14q. Hum. Mol. Genet. 2007, 16, 1327–1334. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, D.A.; Ji, F.; Yuferov, V.; Ho, A.; Chen, A.; Levran, O.; Ott, J.; Kreek, M.J. Genotype patterns that contribute to increased risk for or protection from developing heroin addiction. Mol. Psychiatry 2008, 13, 417–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz-Gordillo, P.; Fedrigo, O.; Wray, G.A.; Babbitt, C.C. Extensive changes in the expression of the opioid genes between humans and chimpanzees. Brain Behav. Evol. 2010, 76, 154–162. [Google Scholar] [CrossRef]
- Nielsen, D.; Ji, F.; Yuferov, V.; Ho, A.; He, C.; Ott, J.; Kreek, M.J. Genome-wide association study identifies genes that may contribute to risk for developing heroin addiction. Psychiatr. Genet. 2010, 20, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Zhao, H.; Kranzler, H.R.; Li, M.D.; Jensen, K.P.; Zayats, T.; Farrer, L.A.; Gelernter, J. Genome-Wide Association Study of Copy Number Variations (CNVs) with Opioid Dependence. Neuropsychopharmacology 2014, 40, 1016–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brick, L.A.; Micalizzi, L.; Knopik, V.S.; Palmer, R.H.C. Characterization of DSM-IV Opioid Dependence among Individuals of European Ancestry. J. Stud. Alcohol Drugs 2019, 80, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Zhou, H.; Sherva, R.; Farrer, L.A.; Kranzler, H.R.; Gelernter, J. Genome-wide Association Study Identifies a Regulatory Variant of RGMA Associated with Opioid Dependence in European Americans. Biol. Psychiatry 2018, 84, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Polimanti, R.; Walters, R.K.; Johnson, E.C.; McClintick, J.N.; Adkins, A.E.; Adkins, D.E.; Bacanu, S.A.; Bierut, L.J.; Bigdeli, T.B.; Brown, S.; et al. Workgroup, Leveraging genome-wide data to investigate differences between opioid use vs. opioid dependence in 41,176 individuals from the Psychiatric Genomics Consortium. Mol. Psychiatry 2020, 25, 1673–1687. [Google Scholar] [CrossRef]
- Zhou, H.; Rentsch, C.T.; Cheng, Z.; Kember, R.L.; Nunez, Y.Z.; Sherva, R.M.; Tate, J.P.; Dao, C.; Xu, K.; Polimanti, R.; et al. Program, Association of OPRM1 Functional Coding Variant with Opioid Use Disorder: A Genome-Wide Association Study. JAMA Psychiatry 2020, 77, 1072–1080. [Google Scholar] [CrossRef]
- Anton, R.F.; Oroszi, G.; O’Malley, S.; Couper, D.; Swift, R.; Pettinati, H.; Goldman, D. An evaluation of mu-opioid receptor (OPRM1) as a predictor of naltrexone response in the treatment of alcohol dependence: Results from the Combined Pharma-cotherapies and Behavioral Interventions for Alcohol Dependence (COMBINE) study. Arch. Gen. Psychiatry 2008, 65, 135–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eap, C.B.; Buclin, T.; Baumann, P. Interindividual variability of the clinical pharmacokinetics of methadone: Implications for the treatment of opioid dependence. Clin. Pharm. 2002, 41, 1153–1193. [Google Scholar] [CrossRef] [PubMed]
- Silverman, D.A.; Nettleton, R.T.; Spencer, K.B.; Wallisch, M.; Olsen, G.D. S-Methadone augments R-methadone induced respiratory depression in the neonatal guinea pig. Respir. Physiol. Neurobiol. 2009, 169, 252–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eap, C.B.; Finkbeiner, T.; Gastpar, M.; Scherbaum, N.; Powell, K.; Baumann, P. Replacement of (R)-methadone by a double dose of (R,S)-methadone in addicts: Interindividual variability of the (R)/(S) ratios and evidence of adaptive changes in methadone pharmacokinetics. Eur. J. Clin. Pharmacol. 1996, 50, 385–389. [Google Scholar] [CrossRef]
- Dennis, B.B.; Bawor, M.; Thabane, L.; Sohani, Z.; Samaan, Z. Impact of ABCB1 and CYP2B6 Genetic Polymorphisms on Methadone Metabolism, Dose and Treatment Response in Patients with Opioid Addiction: A Systematic Review and Meta-Analysis. PLoS ONE 2014, 9, e86114. [Google Scholar] [CrossRef]
- Smith, A.H.; Jensen, K.P.; Li, J.; Nunez, Y.; Farrer, L.A.; Hakonarson, H.; Cook-Sather, S.D.; Kranzler, H.R.; Gelernter, J. Genome-wide association study of therapeutic opioid dosing identifies a novel locus upstream of OPRM1. Mol. Psychiatry 2017, 22, 346–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.-C.; Chu, S.-K.; Huang, C.-L.; Kuo, H.-W.; Wang, S.-C.; Liu, S.-W.; Ho, I.-K.; Liu, Y.-L. Genome-Wide Pharmacogenomic Study on Methadone Maintenance Treatment Identifies SNP rs17180299 and Multiple Haplotypes on CYP2B6, SPON1, and GSG1L Associated with Plasma Concentrations of Methadone R- and S-enantiomers in Heroin-Dependent Patients. PLoS Genet. 2016, 12, e1005910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, F.; Torrens, M. Pharmacogenetics of Methadone Response. Mol. Diagn. Ther. 2018, 22, 57–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Kantelip, J.P.; van Schieveen, P.G.; Davani, S. Interindividual variability of methadone response: Impact of genetic polymorphism. Mol. Diagn Ther. 2008, 12, 109–124. [Google Scholar] [CrossRef]
- Orman, J.S.; Keating, G.M. Buprenorphine/naloxone: A review of its use in the treatment of opioid dependence. Drugs 2009, 69, 577–607. [Google Scholar] [CrossRef]
- Orman, J.S.; Keating, G.M. Spotlight on Buprenorphine/Naloxone in the Treatment of Opioid Dependence. CNS Drugs 2009, 23, 899–902. [Google Scholar] [CrossRef]
- Peterson, R.E.; Kuchenbaecker, K.; Walters, R.K.; Chen, C.-Y.; Popejoy, A.B.; Periyasamy, S.; Lam, M.; Iyegbe, C.; Strawbridge, R.J.; Brick, L.; et al. Genome-wide Association Studies in Ancestrally Diverse Populations: Opportunities, Methods, Pitfalls, and Recommendations. Cell 2019, 179, 589–603. [Google Scholar] [CrossRef]
- Hyman, S.E.; Malenka, R.C.; Nestler, E.J. NEURAL MECHANISMS OF ADDICTION: The Role of Reward-Related Learning and Memory. Annu. Rev. Neurosci. 2006, 29, 565–598. [Google Scholar] [CrossRef] [Green Version]
- Jaenisch, R.; Bird, A. Epigenetic regulation of gene expression: How the genome integrates intrinsic and environmental signals. Nat. Genet. 2003, 33, 245–254. [Google Scholar] [CrossRef]
- Egervari, G.; Landry, J.; Callens, J.; Fullard, J.F.; Roussos, P.; Keller, E.; Hurd, Y.L. Striatal H3K27 Acetylation Linked to Glu-tamatergic Gene Dysregulation in Human Heroin Abusers Holds Promise as Therapeutic Target. Biol. Psychiatry 2017, 81, 585–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, P.; Nestler, E.J. Epigenetics and addiction. Curr. Opin. Neurobiol. 2019, 59, 128–136. [Google Scholar] [CrossRef]
- Browne, C.J.; Godino, A.; Salery, M.; Nestler, E.J. Epigenetic Mechanisms of Opioid Addiction. Biol. Psychiatry 2020, 87, 22–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, J.; Lv, Z.G.; Wang, L.; Zhou, Y.; Hui, B. Histone H3 phosphoacetylation is critical for heroin-induced place preference. NeuroReport 2011, 22, 575–580. [Google Scholar] [CrossRef]
- Wang, Z.; Yan, P.; Hui, T.; Zhang, J. Epigenetic upregulation of PSD-95 contributes to the rewarding behavior by morphine conditioning. Eur. J. Pharmacol. 2014, 732, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Zhao, M.; Li, M.; Song, T.; Zhang, M.; Quan, L.; Li, S.; Sun, Z. Reversal of Cocaine-Conditioned Place Preference through Methyl Supplementation in Mice: Altering Global DNA Methylation in the Prefrontal Cortex. PLoS ONE 2012, 7, e33435. [Google Scholar] [CrossRef]
- Wright, K.N.; Hollis, F.; Duclot, F.; Dossat, A.M.; Strong, C.E.; Francis, T.; Mercer, R.; Feng, J.; Dietz, D.M.; Lobo, M.K.; et al. Methyl Supplementation Attenuates Cocaine-Seeking Behaviors and Cocaine-Induced c-Fos Activation in a DNA Methylation-Dependent Manner. J. Neurosci. 2015, 35, 8948–8958. [Google Scholar] [CrossRef]
- Massart, R.; Barnea, R.; Dikshtein, Y.; Suderman, M.; Meir, O.; Hallett, M.; Kennedy, P.J.; Nestler, E.J.; Szyf, M.; Yadid, G. Role of DNA Methylation in the Nucleus Accumbens in Incubation of Cocaine Craving. J. Neurosci. 2015, 35, 8042–8058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, D.M.; Nestler, E.J. Neuroepigenetics and addiction. Cerebellum: Embryol. Diagn. Investig. 2018, 148, 747–765. [Google Scholar] [CrossRef]
- Tapocik, J.D.; Luu, T.V.; Mayo, C.L.; Wang, B.-D.; Doyle, E.; Lee, A.D.; Lee, N.H.; Elmer, G.I. Neuroplasticity, axonal guidance and micro-RNA genes are associated with morphine self-administration behavior. Addict. Biol. 2013, 18, 480–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Hwang, C.K.; Zheng, H.; Wagley, Y.; Lin, H.Y.; Kim, D.K.; Law, P.Y.; Loh, H.H.; Wei, L.N. MicroRNA 339 down-regulates μ-opioid receptor at the post-transcriptional level in response to opioid treatment. FASEB J. 2013, 27, 522–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, E.M.; Larson, E.; Guzman, D.; Wissman, A.M.; Neve, R.L.; Nestler, E.J.; Self, D.W. Overexpression of the Histone Dimethyltransferase G9a in Nucleus Accumbens Shell Increases Cocaine Self-Administration, Stress-Induced Reinstatement, and Anxiety. J. Neurosci. 2018, 38, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, M.; Carreira, M.B.; Cooper, Y.A.; Bobadilla, A.-C.; Heinsbroek, J.A.; Koike, N.; Larson, E.; Balmuth, E.A.; Hughes, B.W.; Penrod, R.D.; et al. HDAC5 and Its Target Gene, Npas4, Function in the Nucleus Accumbens to Regulate Cocaine-Conditioned Behaviors. Neuron 2017, 96, 130–144.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberti, A.; Valdes, A.F.; Torrecillas, R.; Fraga, M.F.; Fernandez, A.F. Epigenetics in cancer therapy and nanomedicine. Clin. Epigenetics 2019, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]


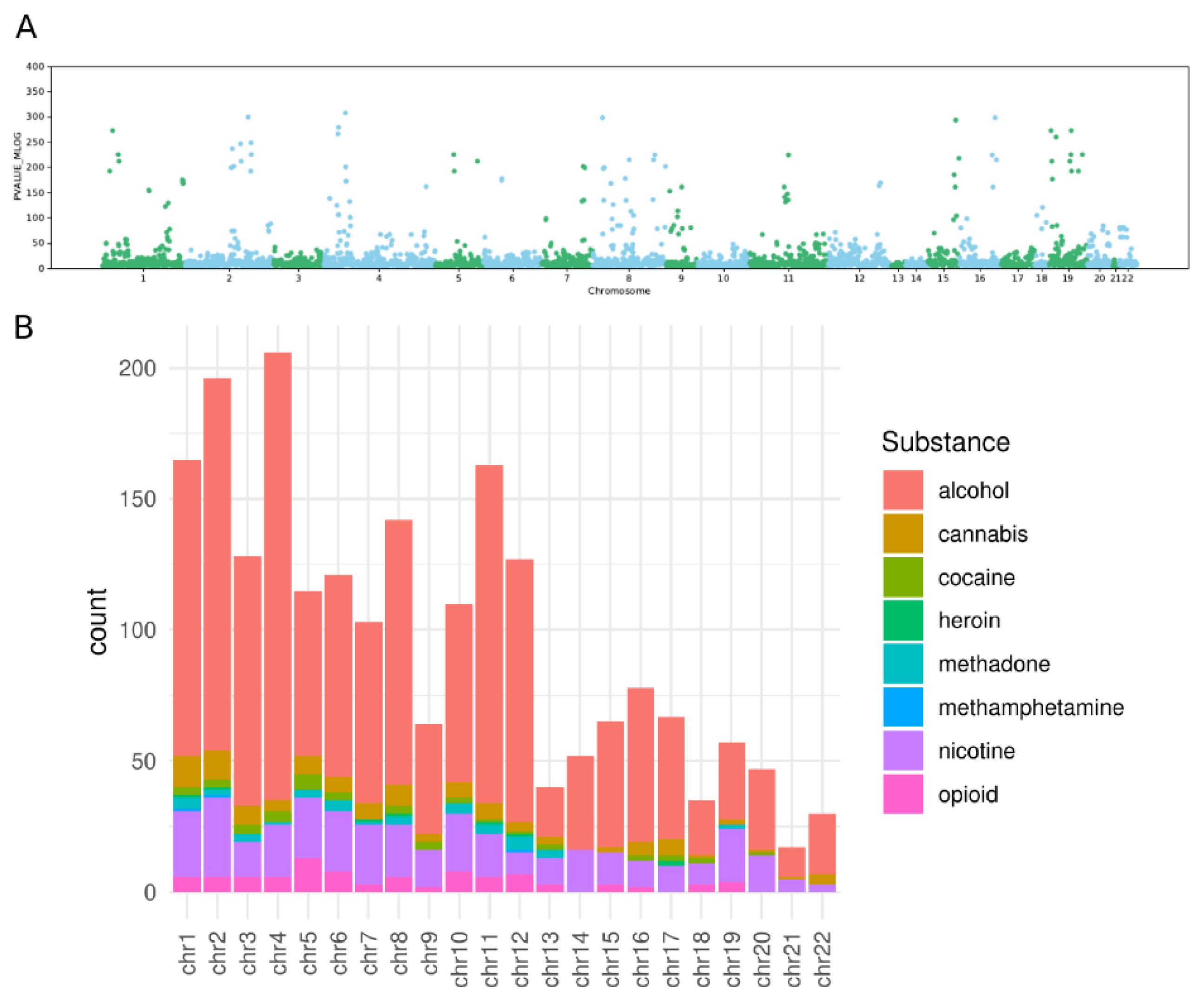{kind=link}
| Trait | Gene | Extended Gene Name | Biological Role in Drug Addiction | Sample size (# Individuals) | References |
|---|---|---|---|---|---|
| Cannabis use | CADM2 | Cell adhesion molecule 2 | Unknown | 184,765 Europeans | Pasman, J.A., et al., 2018 |
| SDK1 | Sidekick cell adhesion molecule 1 | Unknown | |||
| ZNF704 | Zinc finger protein 704 | Unknown | |||
| NCAM1 | Neural cell adhesion molecule 1 | Unknown | |||
| NCAM1 | Neural cell adhesion molecule 1 | Unknown | 32,330 Europeans | Stringer, S., et al., 2016 | |
| RABEP2 | RAB GTPase binding effector protein 2 | Unknown | 184,765 Europeans | Pasman, J.A., et al., 2018 | |
| ATP2A1 | ATPase sarcoplasmic/endoplasmic reticulum Ca2+ transporting 1 | Unknown | |||
| SMG6 | SMG6 nonsense mediated mRNA decay factor | Unknown | |||
| Cannabis use (age at onset) | ATP2C2 | ATPase secretory pathway Ca2+ transporting 2 | Possible involvement of calcium signalling mechanism | 24,953 individuals | Minică, C.C., et al., 2013 |
| Cannabis use disorder | CHRNA2 | Cholinergic receptor nicotinic alpha 2 subunit | Possible direct and indirect interaction of cannabis with the alpha 2 subunit Possible biological link between CHRNA2 and cannabioid receptor genes | 2387 cases 48,985 controls | Demontis, D., et al., 2019 |
| Cocaine dependence | FAM53B | Family with sequence similarity 53 member B | Unknown | 4498 Europeans 2114 African Americans | Gelernter, J., et al., 2014 |
| NFAT5 | Nuclear factor of activated T cells 5 | Unknown | 806 cocaine-dependents 817 controls | Fernàndez-Castillo, N., et al., 2015 | |
| C1QL2 | Complement C1q Like 2 | Unknown | 4498 Europeans 2114 African Americans | Gelernter, J., et al., 2014 | |
| KCTD20 | Potassium channel tetramerization domain containing 20 | Possiblly involved in the distruption of hippocampal gene networks | |||
| STK38 | Serine/threonine kinase 38 | Unknown | |||
| Cocaine use disorder | LINC01411 | Long intergenic non-protein coding RNA 1411 | Unknown | 2070 African Americans 1570 European Americans | Sun, J., et al., 2019 |
| TMEM51 | Transmembrane protein 51 | Unknown | |||
| TRAK2 | Trafficking kinesin protein 2 | Possible interaction with GABA-A receptors when increasing the inhibition of dopamine neuron in response to cocaine | |||
| Opioid sensitivity | KCNG2 | Potassium voltage-gated channel modifier subfamily G member 2 | Possible involvement of calcium and potassium transport and signalling mechanism | 1383 European cases 996 European controls 683 African American cases 2635 African American controls | Gelernter, J., et al., 2013 |
| KCNC1 | Potassium voltage-gated channel subfamily C member 1 | ||||
| APBB2 | Amyloid beta precursor protein binding family B member 2 | ||||
| Opioid use disorder | OPRM1 | Opioid receptor mu 1 | Unknown | 10,544 European cases 72,163 Europeans controls 32,088 African Americans | Zhou, H., et al., 2020 |
| Drug use measurement | PARVA | Parvin alpha | Possible involvement of calcium and potassium transport and signalling mechanism | 175 Netherlands cases 6268 Netherlands controls | Noordam, R., et al., 2015 |
| Opioid dependence | CNIH3 | Cornichon family AMPA receptor auxiliary protein 3 | Unknown | 1167 Europeans cases 161 European controls | Nelson, E.C., et al., 2015 |
| RGMA | Repulsive guidance molecule BMP co-receptor A | Unknown | 3058 opioid-exposed European Americans | Cheng, Z., et al., 2018 | |
| Opioid exposure | BEND4 | BEN domain containing 4 | Unknown | 1297 African cases 2876 European cases 7063 African controls 25,437 European controls | Polimanti, R., et al., 2020 |
| SLC30A9 | Solute carrier family 30 member 9 | Unknown | |||
| MMT | CYP3A4 | Cytochrome P450 family 3 subfamily A member 4 | Unknown | 366 Han Chinese in MMT | Chen, C.H., et al., 2011 |
| Methadone Metabolism, Dose and Treatment Response | ABCB1 | ATP binding cassette subfamily B member 1 | Unknown | 1052 opioid dependents | Dennis, B.B., et al., 2014 |
| CYP2B6 | Cytochrome P450 family 2 subfamily B member 6 | Unknown | |||
| Therapeutic methadone dose | OPRM1 | Opioid receptor mu 1 | Unknown | 383 African-American Opioid Dependents 1027 European-Americans Opioid Dependent 241 opioid-naive African American children | Smith, A.H., et al., 2017 |
| MMT | GSG1L | GSG1 like | Unknown | 360 MMT patients | Yang, H.C., et al., 2016 |
| CYP450 | CYP450 genes | Unknown | |||
| OPRD1 | Opioid receptor delta 1 | Unknown | 70 African Americans Cocaine and opioid codependent | Thomas, P.S., et al., 2021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cozzoli, D.; Daponte, A.; De Fazio, S.; Ariano, V.; Quaranta, M.R.; Leone, V.; Ostuni, A.; Casanova, M.; Catacchio, C.R.; Ventura, M.; et al. Genomic and Personalized Medicine Approaches for Substance Use Disorders (SUDs) Looking at Genome-Wide Association Studies. Biomedicines 2021, 9, 1799. https://doi.org/10.3390/biomedicines9121799
Cozzoli D, Daponte A, De Fazio S, Ariano V, Quaranta MR, Leone V, Ostuni A, Casanova M, Catacchio CR, Ventura M, et al. Genomic and Personalized Medicine Approaches for Substance Use Disorders (SUDs) Looking at Genome-Wide Association Studies. Biomedicines. 2021; 9(12):1799. https://doi.org/10.3390/biomedicines9121799
Chicago/Turabian StyleCozzoli, Danilo, Alessia Daponte, Salvatore De Fazio, Vincenza Ariano, Maria Rita Quaranta, Vincenzo Leone, Angelo Ostuni, Margherita Casanova, Claudia Rita Catacchio, Mario Ventura, and et al. 2021. "Genomic and Personalized Medicine Approaches for Substance Use Disorders (SUDs) Looking at Genome-Wide Association Studies" Biomedicines 9, no. 12: 1799. https://doi.org/10.3390/biomedicines9121799