
Expression of miRNAs Targeting ATP Binding Cassette Transporter 1 (ABCA1) among Patients with Significant Carotid Artery Stenosis

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
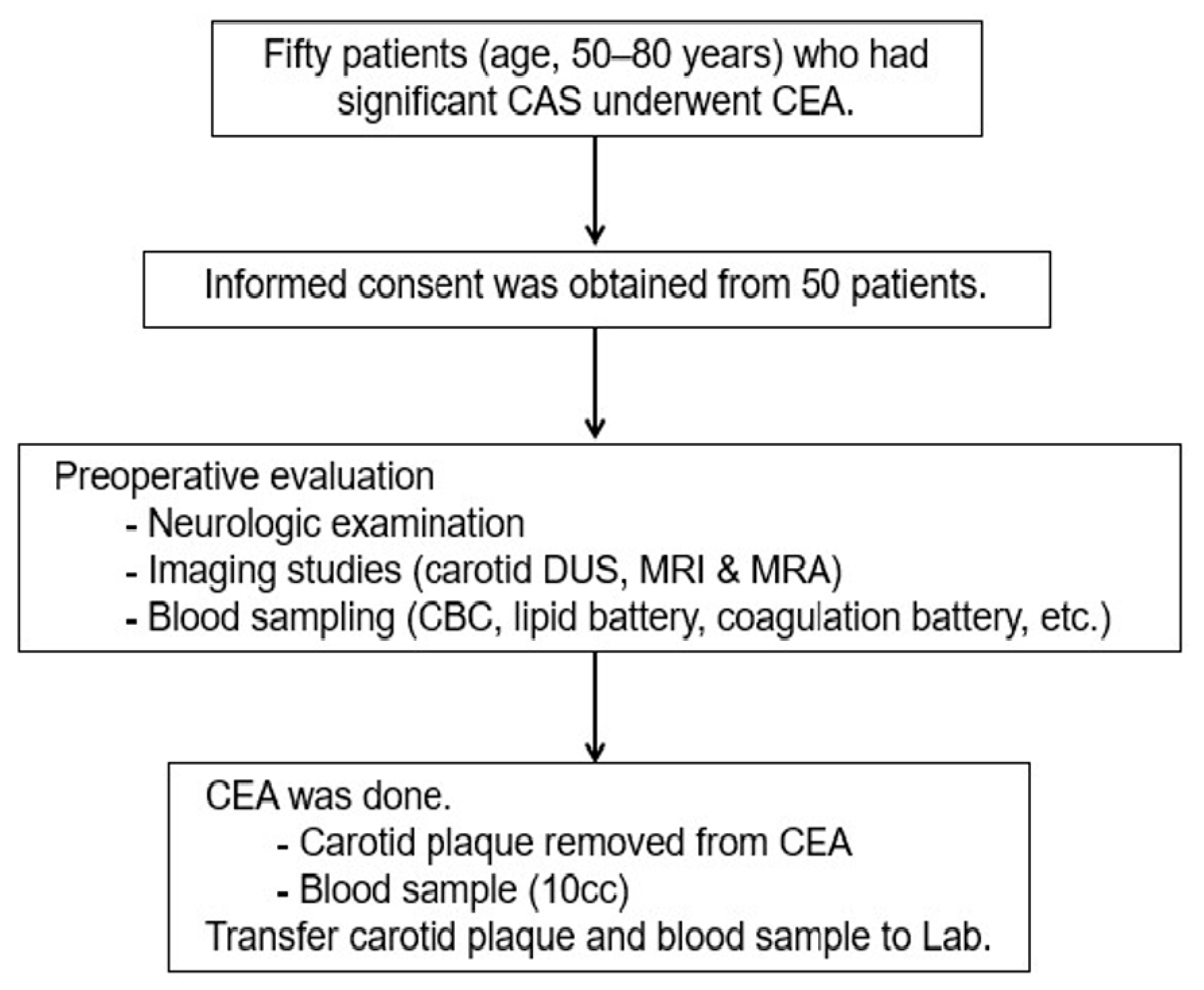
2.1. Study Design and Population
2.2. Index Procedures and Tissue Sampling
2.3. Clinical Outcomes
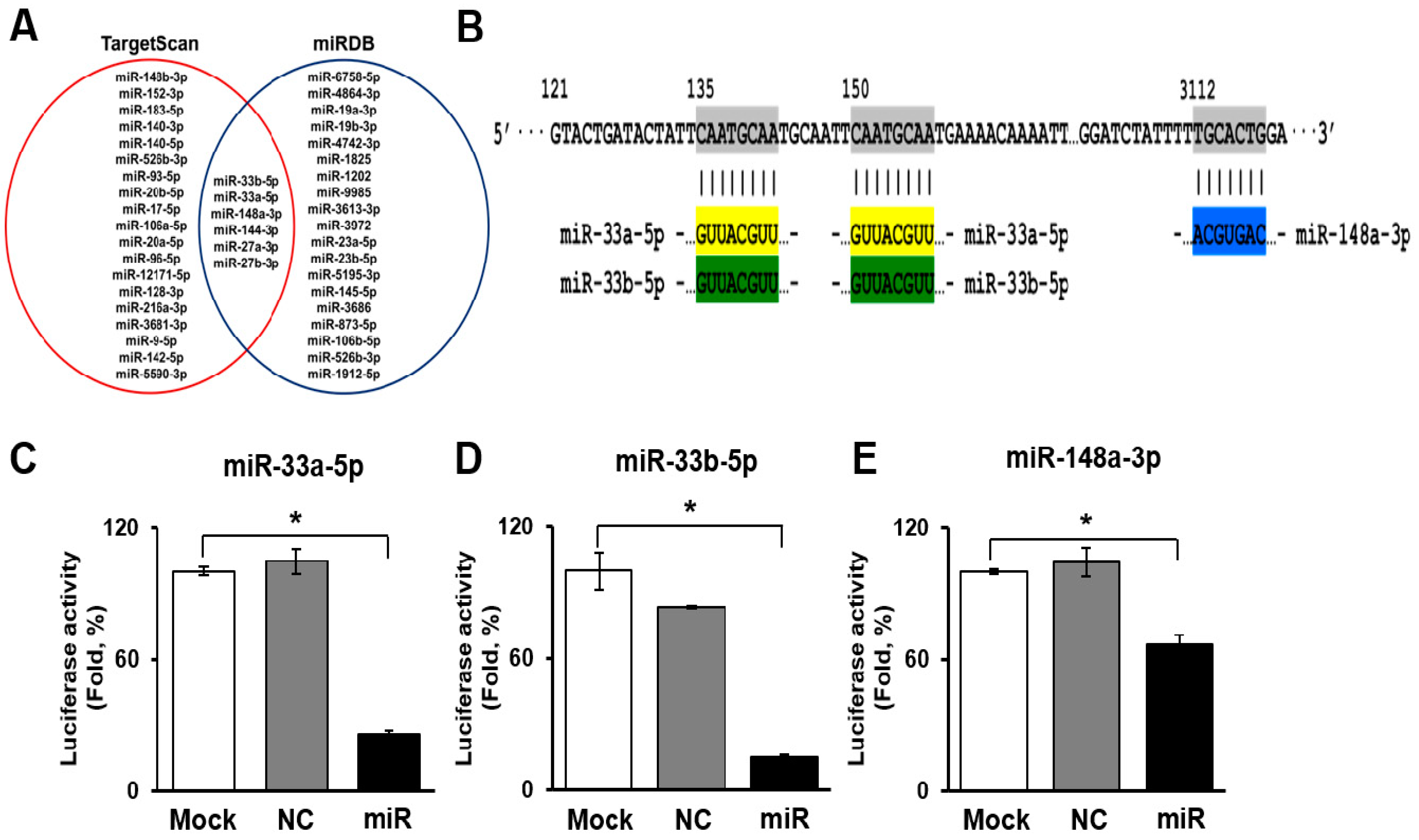
2.4. Prediction of miRNAs Targeting ATP Binding Cassette Transporter 1 (ABCA1)
2.5. ABCA1 ELISA Analysis
2.6. Quantitative Real-Time Chain Reaction (qRT-PCR)
2.7. Immunohistochemistry
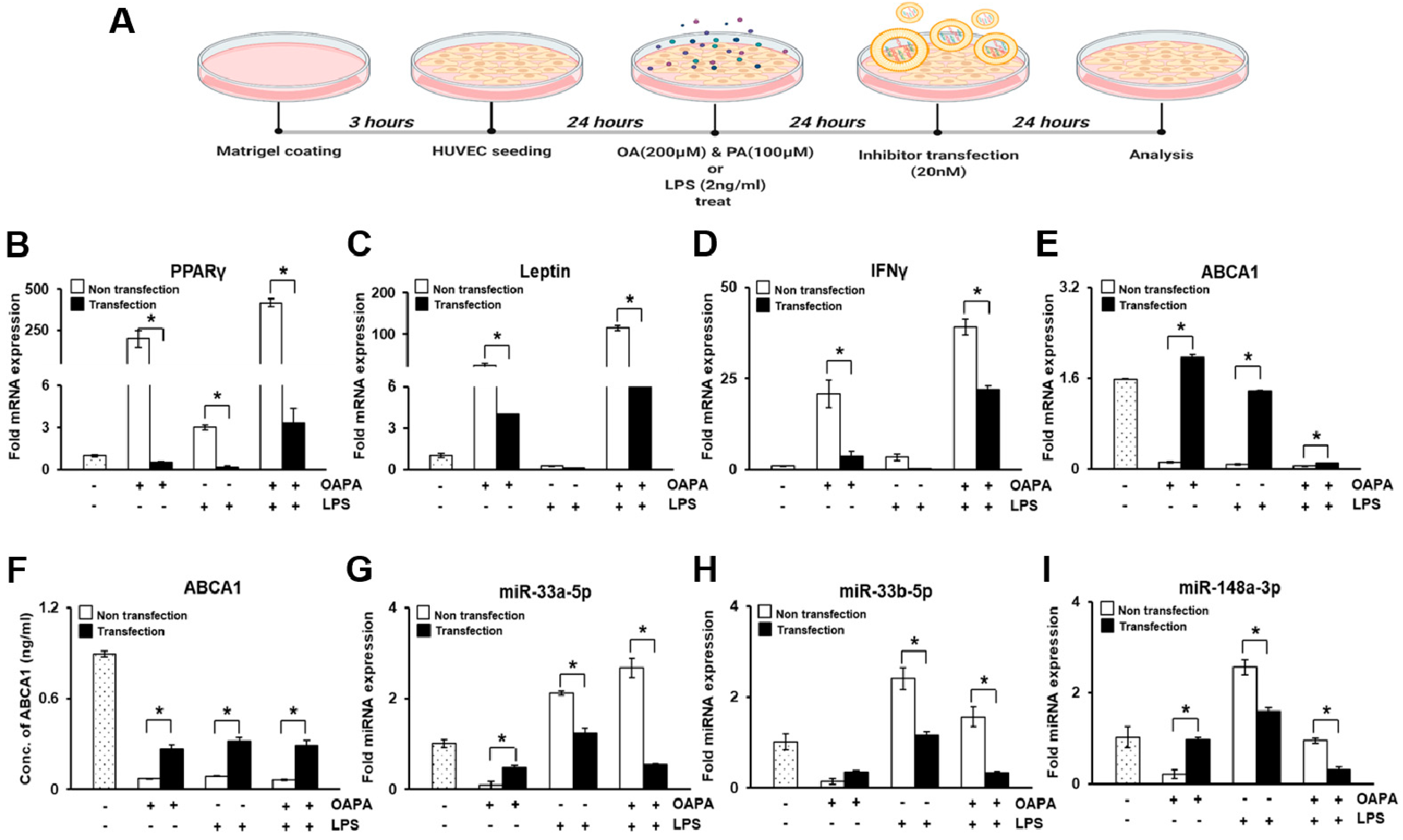
2.8. In Vitro Study with HUVECs
2.9. Cloning
2.10. Dual Luciferase Assay
2.11. Statistical Analysis
3. Results
3.1. Characteristics of Carotid Artery Stenosis
3.2. Selection and Validation of miRNAs Targeting ATP Binding Cassette Transporter 1 (ABCA1)
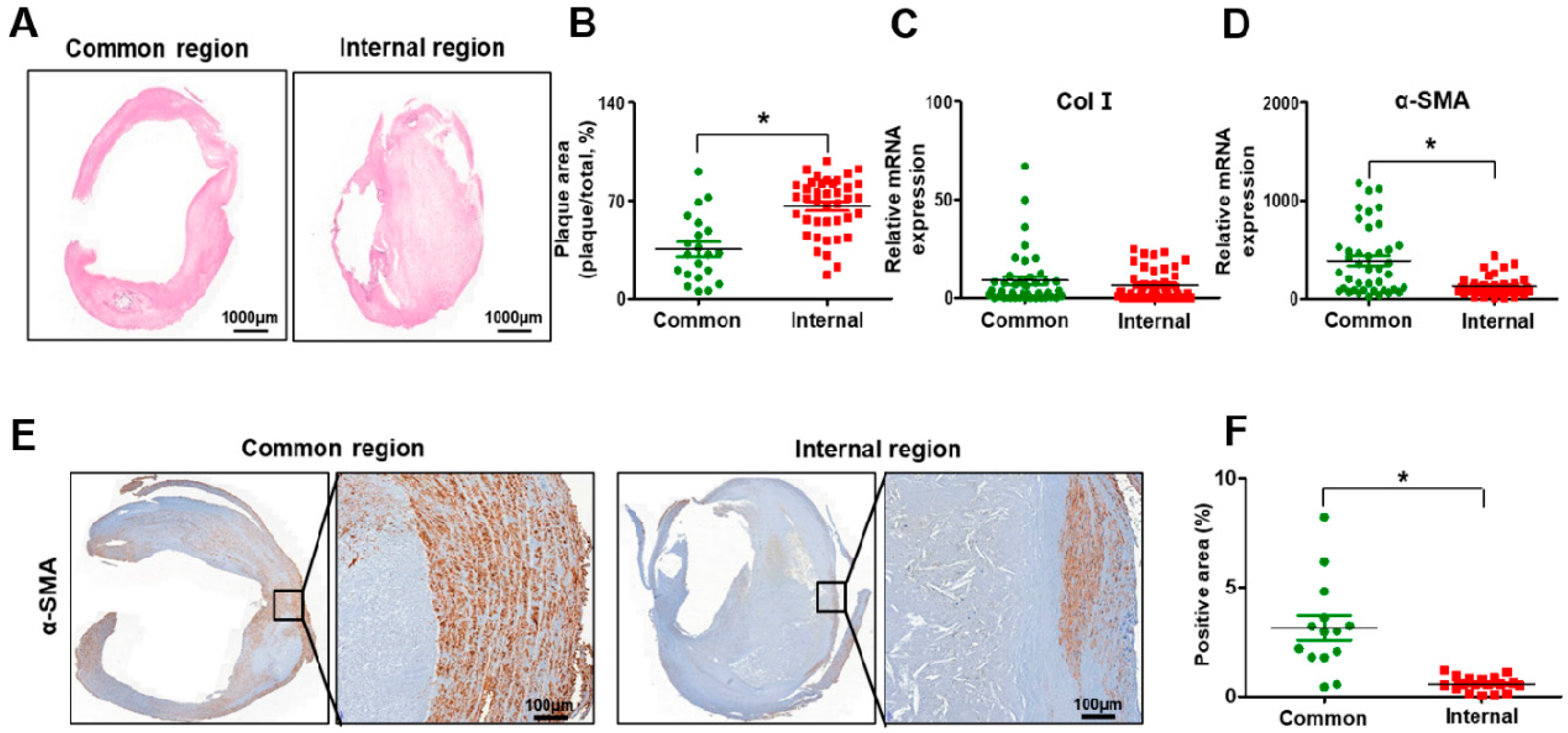
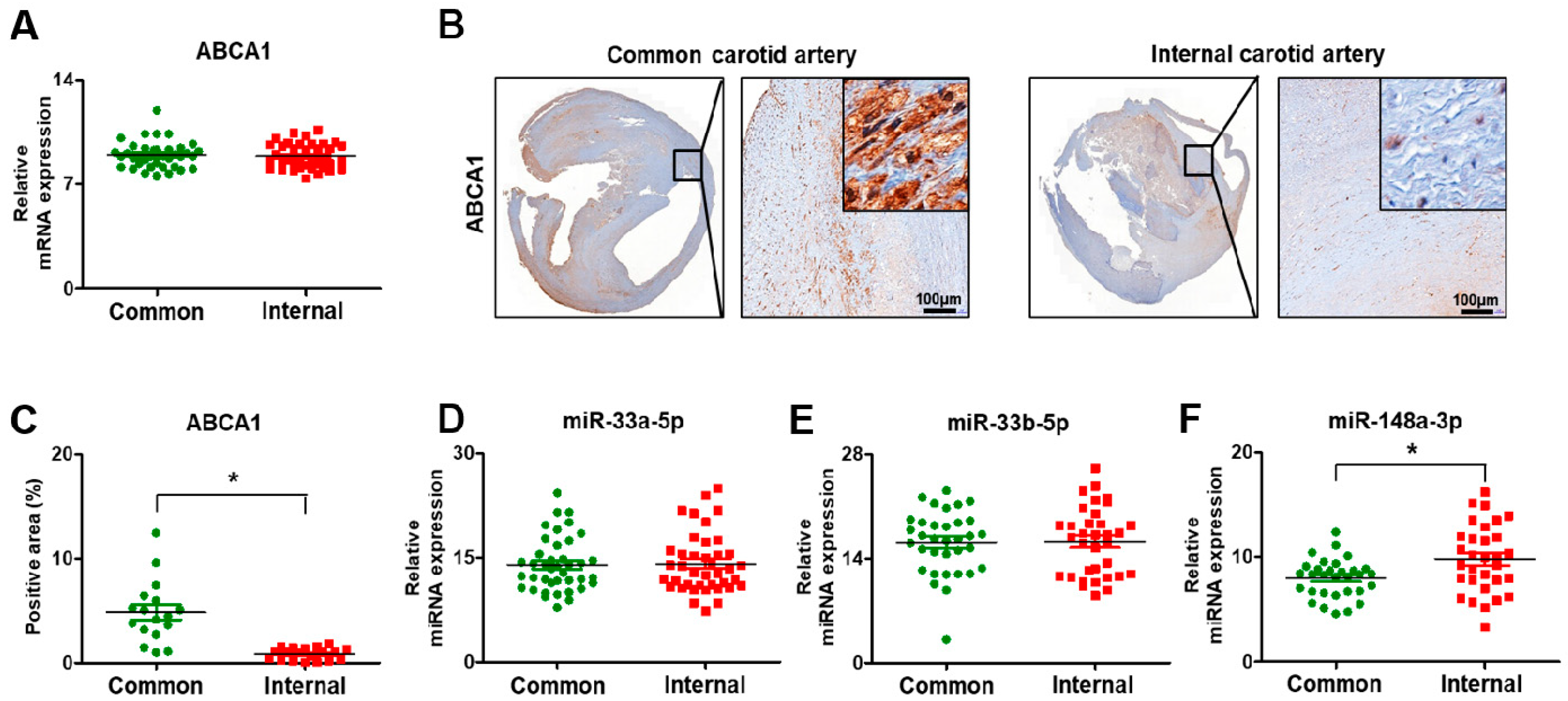
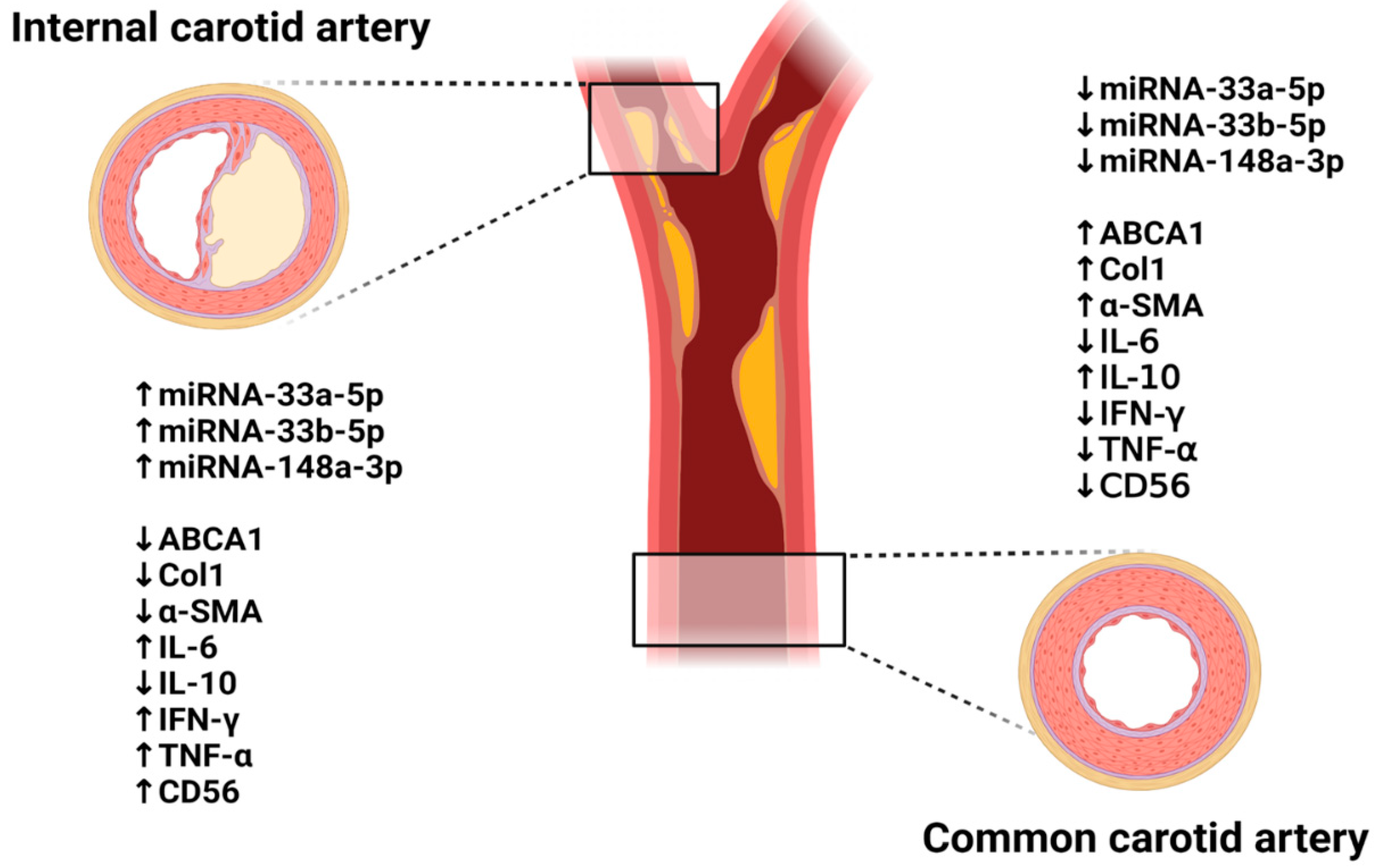
3.3. Histopathological Transitions in Carotid Stenosis Arteries
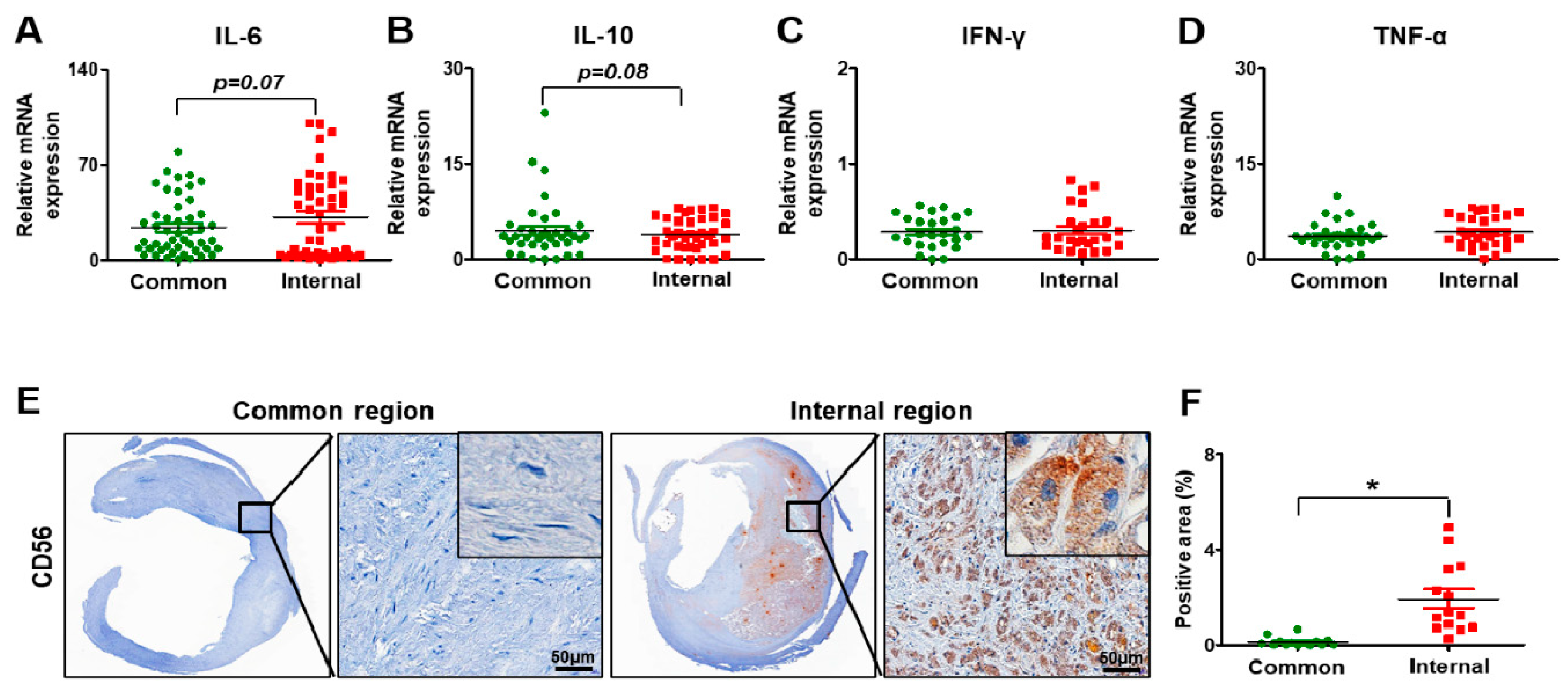
3.4. Inflammation in Carotid Artery Stenosis
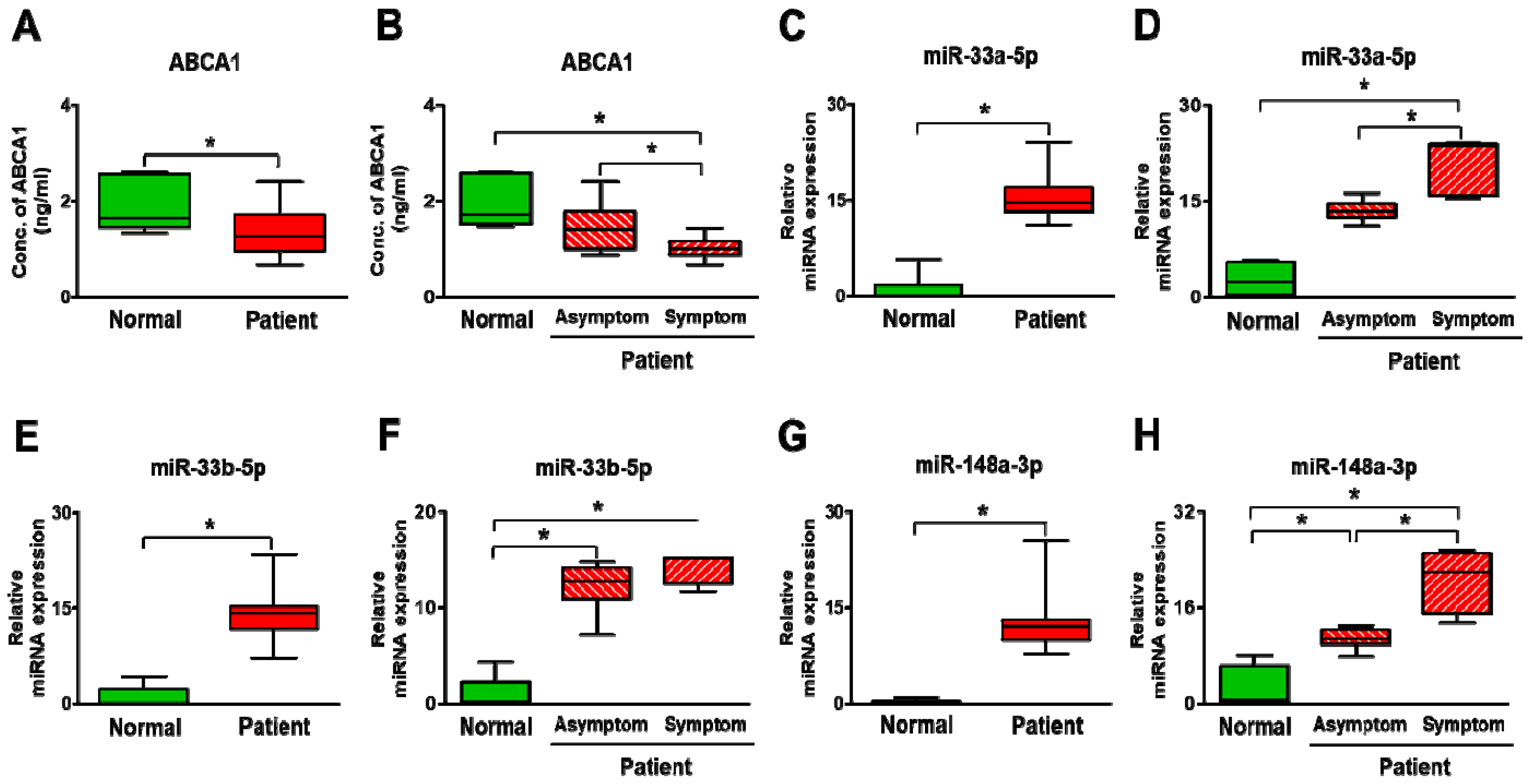
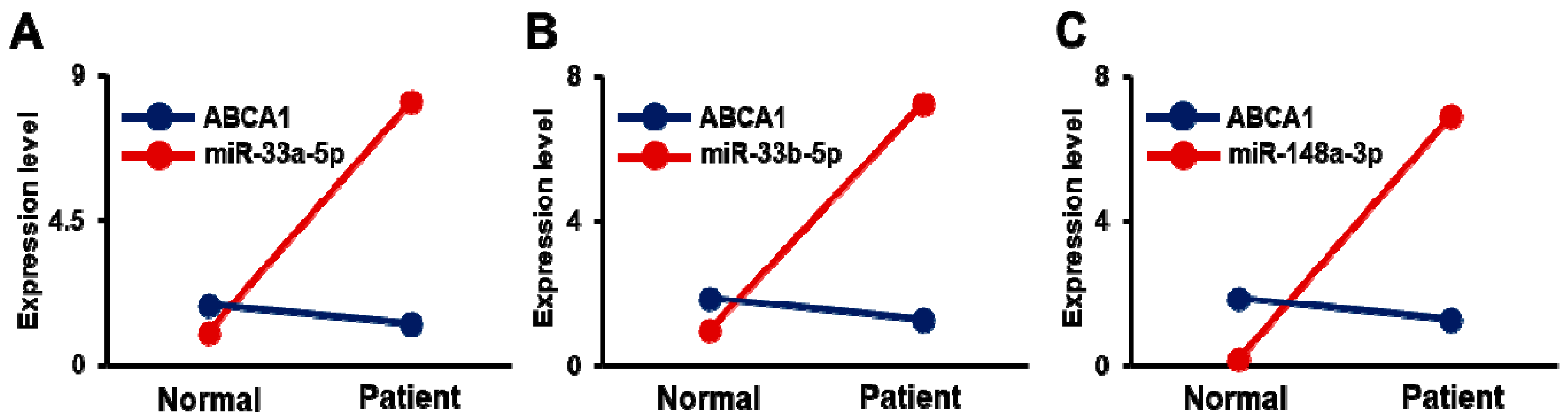
3.5. Expression of ABCA1 and Its Targeting miRNAs in Carotid Artery Stenosis
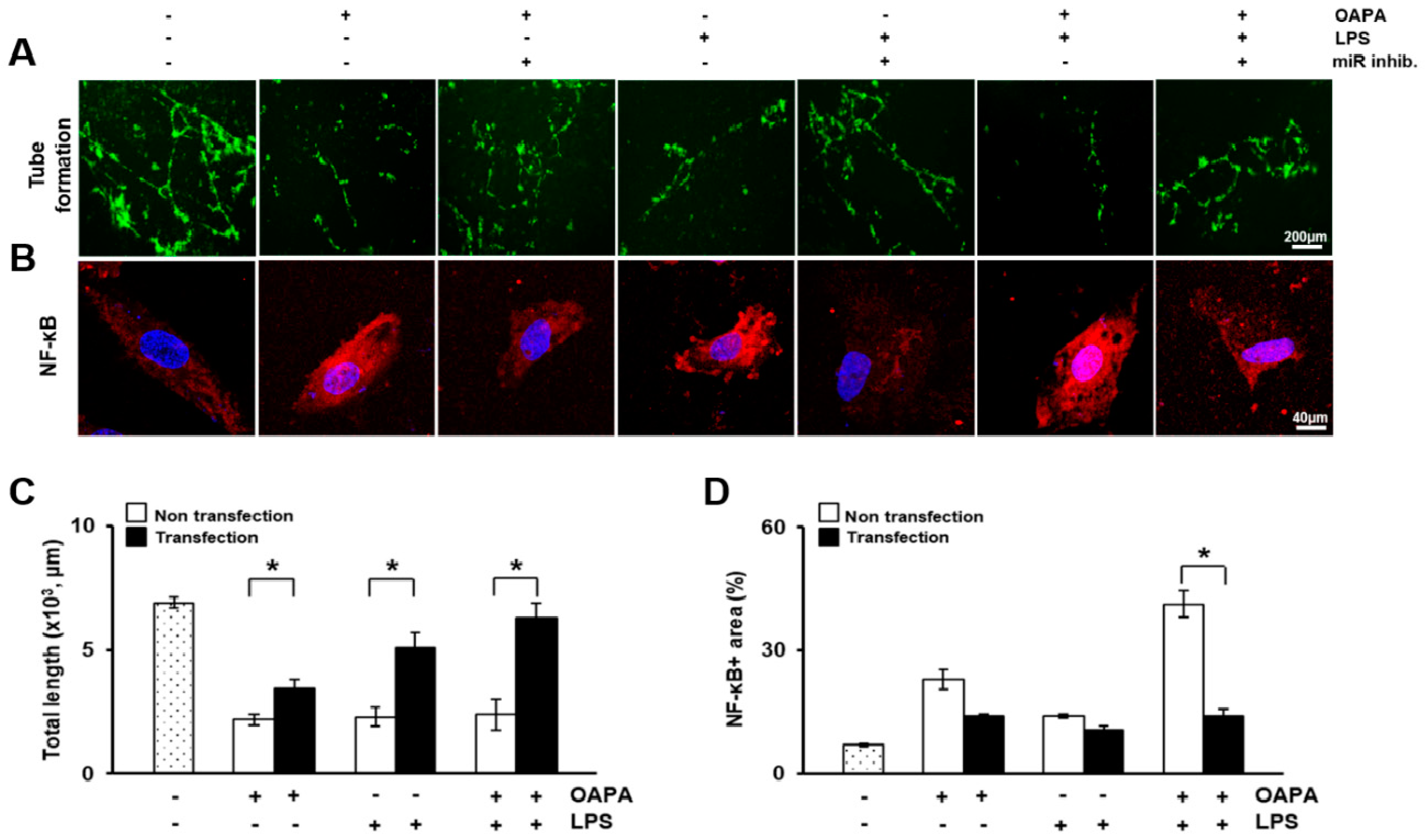
3.6. In Vitro Study in HUVECs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rundek, T.; Arif, H.; Boden-Albala, B.; Elkind, M.S.; Paik, M.C.; Sacco, R.L. Carotid plaque, a subclinical precursor of vascular events: The Northern Manhattan Study. Neurology 2008, 70, 1200–1207. [Google Scholar] [CrossRef] [Green Version]
- Spence, J.D.; Eliasziw, M.; DiCicco, M.; Hackam, D.G.; Galil, R.; Lohmann, T. Carotid plaque area: A tool for targeting and evaluating vascular preventive therapy. Stroke 2002, 33, 2916–2922. [Google Scholar] [CrossRef]
- den Hartog, A.G.; Achterberg, S.; Moll, F.L.; Kappelle, L.J.; Visseren, F.L.; van der Graaf, Y.; Algra, A.; de Borst, G.J.; Group, S.S. Asymptomatic carotid artery stenosis and the risk of ischemic stroke according to subtype in patients with clinical manifest arterial disease. Stroke 2013, 44, 1002–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noh, M.; Kwon, H.; Jung, C.H.; Kwon, S.U.; Kim, M.S.; Lee, W.J.; Park, J.Y.; Han, Y.; Kim, H.; Kwon, T.W.; et al. Impact of diabetes duration and degree of carotid artery stenosis on major adverse cardiovascular events: A single-center, retrospective, observational cohort study. Cardiovasc. Diabetol. 2017, 16, 74. [Google Scholar] [CrossRef] [PubMed]
- Saba, L.; Sanfilippo, R.; Sannia, S.; Anzidei, M.; Montisci, R.; Mallarini, G.; Suri, J.S. Association between carotid artery plaque volume, composition, and ulceration: A retrospective assessment with MDCT. AJR Am. J. Roentgenol. 2012, 199, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Hirt, L.S. Progression rate and ipsilateral neurological events in asymptomatic carotid stenosis. Stroke 2014, 45, 702–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwee, R.M.; van Oostenbrugge, R.J.; Hofstra, L.; Teule, G.J.; van Engelshoven, J.M.; Mess, W.H.; Kooi, M.E. Identifying vulnerable carotid plaques by noninvasive imaging. Neurology 2008, 70, 2401–2409. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Kweon, M.; Kim, J.Y.; Jun, J.H.; Kim, G.J. Research Trends in the Efficacy of Stem Cell Therapy for Hepatic Diseases Based on MicroRNA Profiling. Int. J. Mol. Sci. 2020, 22, 239. [Google Scholar] [CrossRef]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [Green Version]
- Cannell, I.G.; Kong, Y.W.; Bushell, M. How do microRNAs regulate gene expression? Biochem. Soc. Trans. 2008, 36, 1224–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The microRNA spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, J.; Sen, S. MicroRNA as Biomarkers and Diagnostics. J. Cell Physiol. 2016, 231, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.K.; Robertson, A.K.; Soderberg-Naucler, C. Inflammation and atherosclerosis. Annu. Rev. Pathol. 2006, 1, 297–329. [Google Scholar] [CrossRef]
- Badimon, L.; Vilahur, G. LDL-cholesterol versus HDL-cholesterol in the atherosclerotic plaque: Inflammatory resolution versus thrombotic chaos. Ann. N. Y. Acad. Sci. 2012, 1254, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Brunham, L.R.; Kruit, J.K.; Pape, T.D.; Timmins, J.M.; Reuwer, A.Q.; Vasanji, Z.; Marsh, B.J.; Rodrigues, B.; Johnson, J.D.; Parks, J.S.; et al. Beta-cell ABCA1 influences insulin secretion, glucose homeostasis and response to thiazolidinedione treatment. Nat. Med. 2007, 13, 340–347. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, C. Regulation of ABCA1 functions by signaling pathways. Biochim. Biophys. Acta 2012, 1821, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Singaraja, R.R.; Fievet, C.; Castro, G.; James, E.R.; Hennuyer, N.; Clee, S.M.; Bissada, N.; Choy, J.C.; Fruchart, J.C.; McManus, B.M.; et al. Increased ABCA1 activity protects against atherosclerosis. J. Clin. Investig. 2002, 110, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.; Moon, D.H.; Han, Y.; Lee, J.Y.; Kwon, S.U.; Kang, D.W.; Choo, S.J.; Kwon, T.W.; Kim, M.J.; Cho, Y.P. Impact of subclinical coronary artery disease on the clinical outcomes of carotid endarterectomy. J. Neurosurg. 2017, 126, 1560–1565. [Google Scholar] [CrossRef] [Green Version]
- Barkat, M.; Hajibandeh, S.; Hajibandeh, S.; Torella, F.; Antoniou, G.A. Systematic Review and Meta-analysis of Dual Versus Single Antiplatelet Therapy in Carotid Interventions. Eur. J. Vasc. Endovasc. Surg. 2017, 53, 53–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, H.; Kim, H.K.; Kwon, S.U.; Lee, S.W.; Kim, M.J.; Park, J.W.; Noh, M.; Han, Y.; Kwon, T.W.; Cho, Y.P. Risk of major adverse cardiovascular events in subjects with asymptomatic mild carotid artery stenosis. Sci. Rep. 2018, 8, 4700. [Google Scholar] [CrossRef] [Green Version]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.S.; Sander, C. The microRNA.org resource: Targets and expression. Nucleic Acids Res. 2008, 36, D149–D153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goedeke, L.; Fernandez-Hernando, C. Regulation of cholesterol homeostasis. Cell. Mol. Life Sci. 2012, 69, 915–930. [Google Scholar] [CrossRef]
- Oram, J.F. Molecular basis of cholesterol homeostasis: Lessons from Tangier disease and ABCA1. Trends Mol. Med. 2002, 8, 168–173. [Google Scholar] [CrossRef]
- Jacobo-Albavera, L.; Dominguez-Perez, M.; Medina-Leyte, D.J.; Gonzalez-Garrido, A.; Villarreal-Molina, T. The Role of the ATP-Binding Cassette A1 (ABCA1) in Human Disease. Int. J. Mol. Sci. 2021, 22, 1593. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, J.T.; Ning, H.; Stone, N.J.; Criqui, M.H.; Zhao, L.; Greenland, P.; Lloyd-Jones, D.M. Coronary heart disease risks associated with high levels of HDL cholesterol. J. Am. Heart Assoc. 2014, 3, e000519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumitrescu, L.; Carty, C.L.; Taylor, K.; Schumacher, F.R.; Hindorff, L.A.; Ambite, J.L.; Anderson, G.; Best, L.G.; Brown-Gentry, K.; Buzkova, P.; et al. Genetic determinants of lipid traits in diverse populations from the population architecture using genomics and epidemiology (PAGE) study. PLoS Genet. 2011, 7, e1002138. [Google Scholar] [CrossRef] [PubMed]
- Voight, B.F.; Peloso, G.M.; Orho-Melander, M.; Frikke-Schmidt, R.; Barbalic, M.; Jensen, M.K.; Hindy, G.; Holm, H.; Ding, E.L.; Johnson, T.; et al. Plasma HDL cholesterol and risk of myocardial infarction: A mendelian randomisation study. Lancet 2012, 380, 572–580. [Google Scholar] [CrossRef] [Green Version]
- van Rooij, E.; Olson, E.N. MicroRNAs: Powerful new regulators of heart disease and provocative therapeutic targets. J. Clin. Investig. 2007, 117, 2369–2376. [Google Scholar] [CrossRef] [Green Version]
- Fichtlscherer, S.; De Rosa, S.; Fox, H.; Schwietz, T.; Fischer, A.; Liebetrau, C.; Weber, M.; Hamm, C.W.; Roxe, T.; Muller-Ardogan, M.; et al. Circulating microRNAs in patients with coronary artery disease. Circ. Res. 2010, 107, 677–684. [Google Scholar] [CrossRef] [Green Version]
- Maitrias, P.; Metzinger-Le Meuth, V.; Massy, Z.A.; M’Baya-Moutoula, E.; Reix, T.; Caus, T.; Metzinger, L. MicroRNA deregulation in symptomatic carotid plaque. J. Vasc. Surg. 2015, 62, 1245–1250.e1241. [Google Scholar] [CrossRef] [Green Version]
- Canfran-Duque, A.; Ramirez, C.M.; Goedeke, L.; Lin, C.S.; Fernandez-Hernando, C. microRNAs and HDL life cycle. Cardiovasc. Res. 2014, 103, 414–422. [Google Scholar] [CrossRef] [Green Version]
- Rayner, K.J.; Suarez, Y.; Davalos, A.; Parathath, S.; Fitzgerald, M.L.; Tamehiro, N.; Fisher, E.A.; Moore, K.J.; Fernandez-Hernando, C. MiR-33 contributes to the regulation of cholesterol homeostasis. Science 2010, 328, 1570–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Porto, F.; Cifani, N.; Proietta, M.; Dezi, T.; Tritapepe, L.; Raffa, S.; Micaloni, A.; Taurino, M. Lag3(+) regulatory T lymphocytes in critical carotid artery stenosis. Clin. Exp. Med. 2019, 19, 463–468. [Google Scholar] [CrossRef]
- Parahuleva, M.S.; Lipps, C.; Parviz, B.; Holschermann, H.; Schieffer, B.; Schulz, R.; Euler, G. MicroRNA expression profile of human advanced coronary atherosclerotic plaques. Sci. Rep. 2018, 8, 7823. [Google Scholar] [CrossRef]
- Ouimet, M.; Ediriweera, H.N.; Gundra, U.M.; Sheedy, F.J.; Ramkhelawon, B.; Hutchison, S.B.; Rinehold, K.; van Solingen, C.; Fullerton, M.D.; Cecchini, K.; et al. MicroRNA-33-dependent regulation of macrophage metabolism directs immune cell polarization in atherosclerosis. J. Clin. Investig. 2015, 125, 4334–4348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, S.; Horie, T.; Nishino, T.; Baba, O.; Sowa, N.; Miyasaka, Y.; Kuwabara, Y.; Nakao, T.; Nishiga, M.; Nishi, H.; et al. Identification of Differential Roles of MicroRNA-33a and -33b During Atherosclerosis Progression With Genetically Modified Mice. J. Am. Heart Assoc. 2019, 8, e012609. [Google Scholar] [CrossRef] [PubMed]
- Ono, K. Functions of microRNA-33a/b and microRNA therapeutics. J. Cardiol. 2016, 67, 28–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayner, K.J.; Moore, K.J. The plaque “micro” environment: microRNAs control the risk and the development of atherosclerosis. Curr. Atheroscler. Rep. 2012, 14, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Numata, T.; Takahashi, K.; Inoue, R. “TRP inflammation” relationship in cardiovascular system. Semin. Immunopathol. 2016, 38, 339–356. [Google Scholar] [CrossRef] [Green Version]
- Lamers, D.; Schlich, R.; Greulich, S.; Sasson, S.; Sell, H.; Eckel, J. Oleic acid and adipokines synergize in inducing proliferation and inflammatory signalling in human vascular smooth muscle cells. J. Cell. Mol. Med. 2011, 15, 1177–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayden, M.S.; Ghosh, S. NF-kappaB in immunobiology. Cell Res. 2011, 21, 223–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, N.; Xu, S.; Xu, Y.; Little, P.J.; Jin, Z.G. Targeting Mechanosensitive Transcription Factors in Atherosclerosis. Trends Pharmacol. Sci. 2019, 40, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Ouimet, M.; Ediriweera, H.; Afonso, M.S.; Ramkhelawon, B.; Singaravelu, R.; Liao, X.; Bandler, R.C.; Rahman, K.; Fisher, E.A.; Rayner, K.J.; et al. microRNA-33 Regulates Macrophage Autophagy in Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1058–1067. [Google Scholar] [CrossRef] [Green Version]
- Rotllan, N.; Price, N.; Pati, P.; Goedeke, L.; Fernandez-Hernando, C. microRNAs in lipoprotein metabolism and cardiometabolic disorders. Atherosclerosis 2016, 246, 352–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total (N = 50) | |
|---|---|
| Mean age, years | 68.2 ± 6.3 |
| Male | 44 (88) |
| BMI, kg/m2 | 24.0 ± 3.2 |
| Risk factors | |
| Hypertension | 32 (64) |
| Diabetes mellitus | 25 (50) |
| CVD | 20 (40) |
| Heart failure | 10 (20) |
| CKD | 5 (10) |
| Smoking | 32 (64) |
| BMI (kg/m2) | 24.2 ± 3.2 |
| Lipid battery * | |
| LDL (mg/dL) | 76.2 ± 25.6 |
| HDL (mg/dL) | 43.7 ± 10.6 |
| Triglyceride (mg/dL) | 126.54 ± 55.7 |
| Total cholesterol (mg/dL) | 129.9 ± 28.0 |
| Use of statin † | 49 (98) |
| Carotid stenosis | |
| Degree of stenosis, % | |
| ICA | 75.3 ± 7.9 |
| ECA | 27.9 ± 12.5 |
| Symptomatic | 13 (26) |
| Stroke | 6 (12) |
| TIA | 7 (14) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, S.; Jun, J.H.; Kim, J.Y.; Park, H.J.; Cho, Y.-P.; Kim, G.J. Expression of miRNAs Targeting ATP Binding Cassette Transporter 1 (ABCA1) among Patients with Significant Carotid Artery Stenosis. Biomedicines 2021, 9, 920. https://doi.org/10.3390/biomedicines9080920
Jeong S, Jun JH, Kim JY, Park HJ, Cho Y-P, Kim GJ. Expression of miRNAs Targeting ATP Binding Cassette Transporter 1 (ABCA1) among Patients with Significant Carotid Artery Stenosis. Biomedicines. 2021; 9(8):920. https://doi.org/10.3390/biomedicines9080920
Chicago/Turabian StyleJeong, Seonjeong, Ji Hye Jun, Jae Yeon Kim, Hee Jung Park, Yong-Pil Cho, and Gi Jin Kim. 2021. "Expression of miRNAs Targeting ATP Binding Cassette Transporter 1 (ABCA1) among Patients with Significant Carotid Artery Stenosis" Biomedicines 9, no. 8: 920. https://doi.org/10.3390/biomedicines9080920