
Pitavastatin Ameliorates Lipopolysaccharide-Induced Blood-Brain Barrier Dysfunction


 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
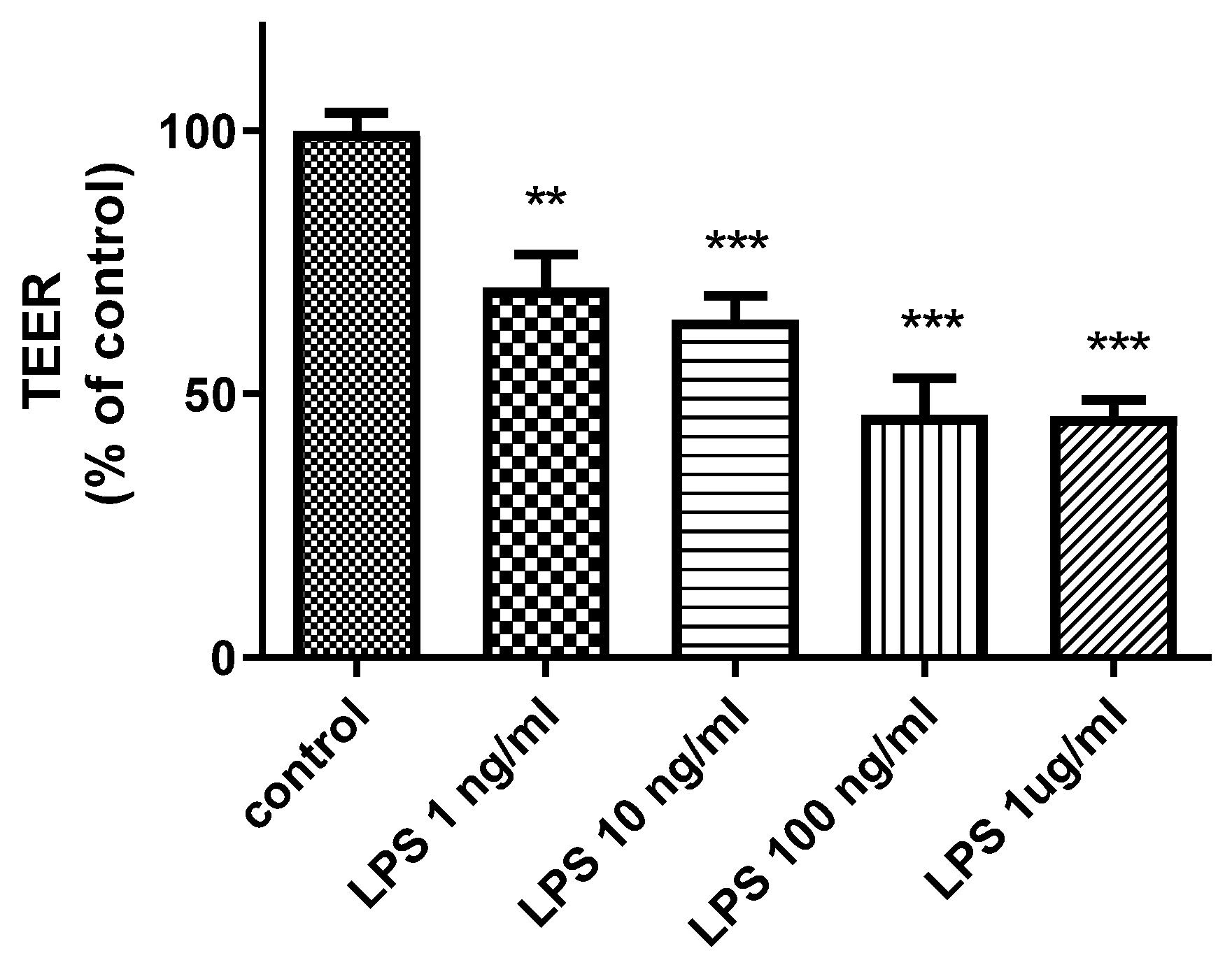
2.1. Effects of LPS on Cerebral Barrier Function
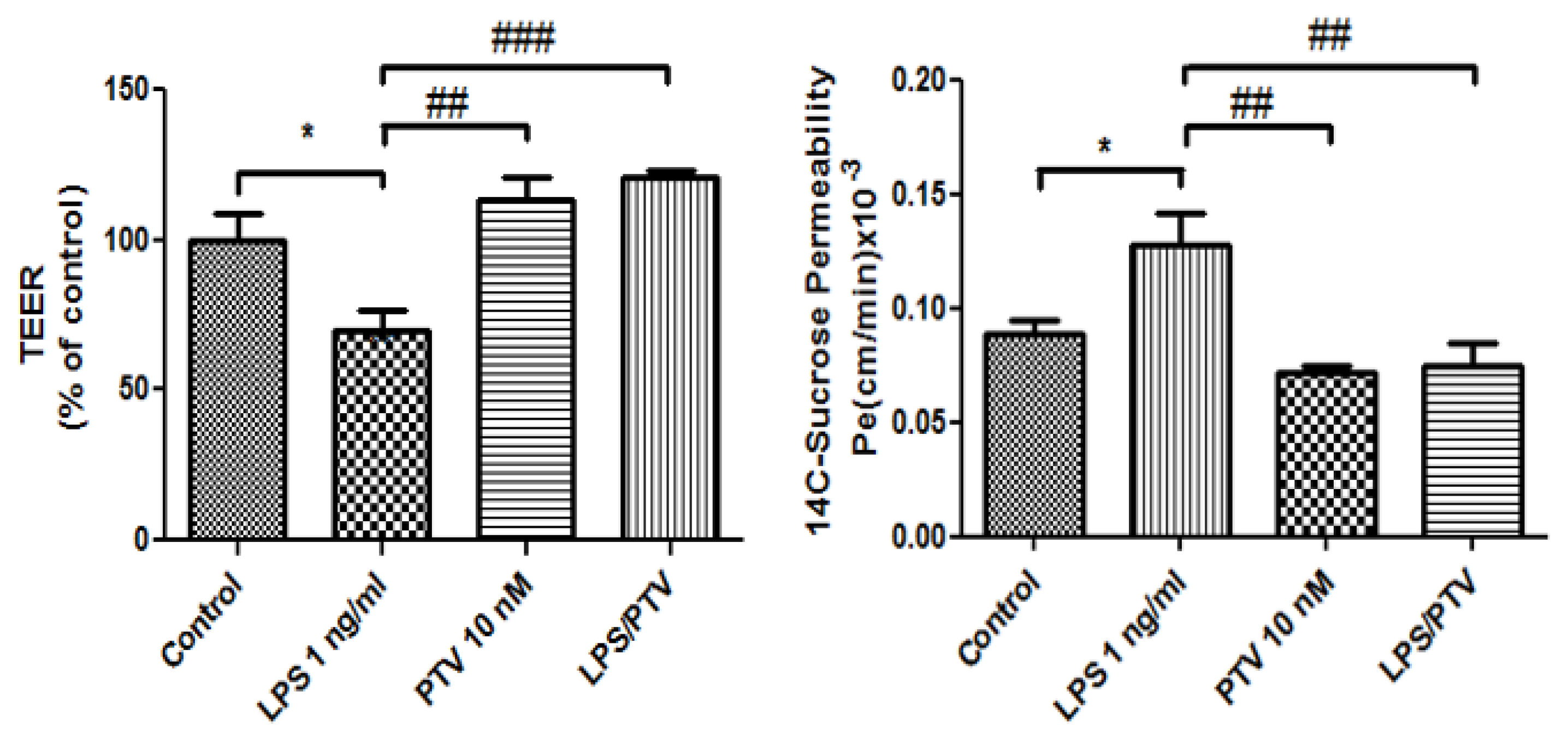
2.2. Effect of PTV on BBB Function
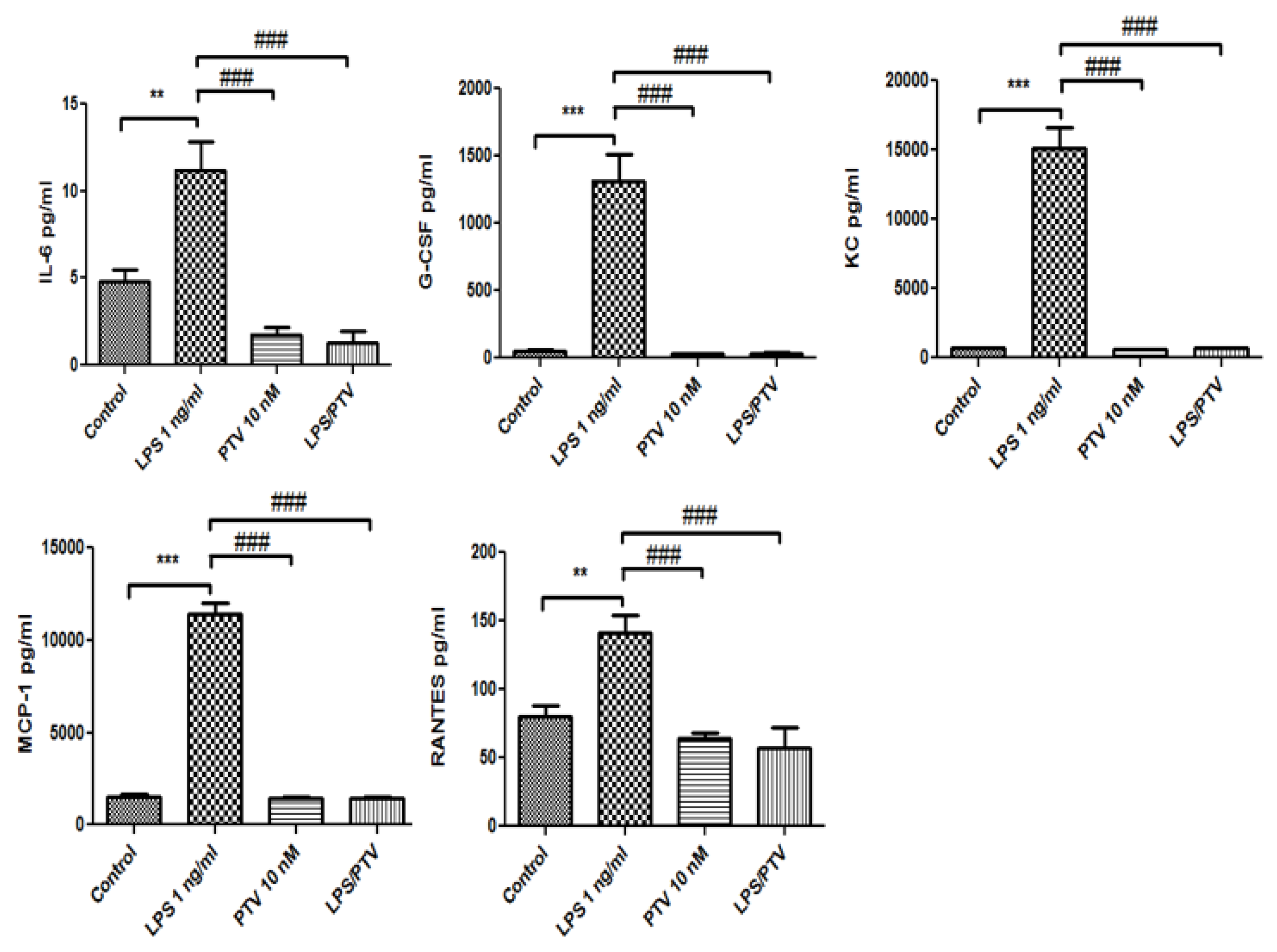
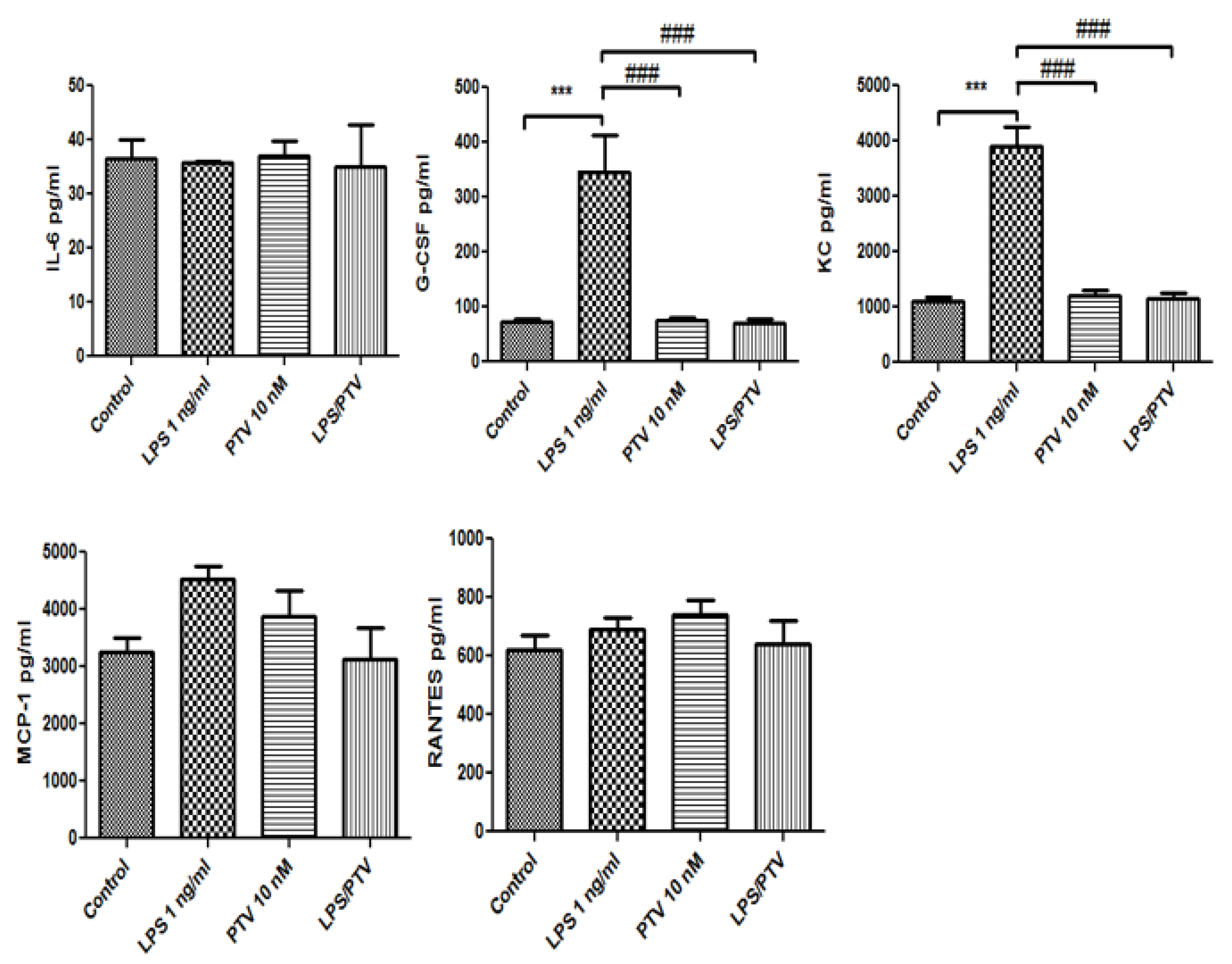
2.3. Secretion of Cytokines and Chemokines
3. Discussion
4. Materials and Methods
4.1. Mouse Brain Endothelial Cell Cultures
4.2. Mouse Brain Pericyte Culture
4.3. Mouse Glial Cell Cultures
4.4. In Vitro BBB Model Generation and Treatment
4.5. TEER and Sucrose Permeability
4.6. Enzyme-Linked Immunosorbent Assay (ELISA) of Cytokines
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flint, A.C.; Kamel, H.; Navi, B.B.; Rao, V.A.; Faigeles, B.S.; Conell, C.; Klingman, J.G.; Sidney, S.; Hills, N.K.; Sorel, M.; et al. Statin Use During Ischemic Stroke Hospitalization Is Strongly Associated With Improved Poststroke Survival. Stroke 2012, 43, 147–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, K.; Yoshimura, S.; Imamura, H.; Ohara, N.; Sakai, N.; Tanaka, K.; Yamagami, H.; Matsumoto, Y.; Takeuchi, M.; Morimoto, T.; et al. Effect of Statin Administration After Onset of Acute Ischemic Stroke With Large Vessel Occlusion: Insights From RESCUE-Japan Registry 2. J. Am. Heart Assoc. 2020, 9, e017472. [Google Scholar] [CrossRef]
- Torrandell-Haro, G.; Branigan, G.L.; Vitali, F.; Geifman, N.; Zissimopoulos, J.M.; Brinton, R.D. Statin therapy and risk of Alzheimer’s and age-related neurodegenerative diseases. Alzheimers Dement (N. Y.) 2020, 6, e12108. [Google Scholar] [CrossRef]
- McFarland, A.J.; Anoopkumar-Dukie, S.; Arora, D.S.; Grant, G.D.; McDermott, C.M.; Perkins, A.V.; Davey, A.K. Molecular mechanisms underlying the effects of statins in the central nervous system. Int. J. Mol. Sci. 2014, 15, 20607–20637. [Google Scholar] [CrossRef] [Green Version]
- Khokhar, B.; Simoni-Wastila, L.; Slejko, J.F.; Perfetto, E.; Zhan, M.; Smith, G.S. In-Hospital Mortality Following Traumatic Brain Injury Among Older Medicare Beneficiaries, Comparing Statin Users With Nonusers. J. Pharm. Technol. 2017, 33, 225–236. [Google Scholar] [CrossRef]
- Xu, X.; Gao, W.; Cheng, S.; Yin, D.; Li, F.; Wu, Y.; Sun, D.; Zhou, S.; Wang, D.; Zhang, Y.; et al. Anti-inflammatory and immunomodulatory mechanisms of atorvastatin in a murine model of traumatic brain injury. J. Neuroinflamm. 2017, 14, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, P.A.; Estato, V.; da Silva, T.I.; d’Avila, J.C.; Siqueira, L.D.; Assis, E.F.; Bozza, P.T.; Bozza, F.A.; Tibiriça, E.V.; Zimmerman, G.A.; et al. Statins decrease neuroinflammation and prevent cognitive impairment after cerebral malaria. PLoS Pathog. 2012, 8, e1003099. [Google Scholar] [CrossRef]
- Wani, M.A.; Mukherjee, S.; Mallick, S.; Akbar, I.; Basu, A. Atorvastatin ameliorates viral burden and neural stem/progenitor cell (NSPC) death in an experimental model of Japanese encephalitis. J. Biosci. 2020, 45, 1–17. [Google Scholar] [CrossRef]
- Tokgözoglu, L.; Zamorano, J.L. Current perspectives on the use of statins in the treatment of dyslipidaemic patients: Focus on pitavastatin. Drugs Context 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Chu, U.B.; Duellman, T.; Weaver, S.J.; Tao, Y.; Yang, J. Endothelial protective genes induced by statin are mimicked by ERK5 activation as triggered by a drug combination of FTI-277 and GGTI-298. Biochim. Biophys. Acta 2015, 1850, 1415–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayano, R.; Morofuji, Y.; Nakagawa, S.; Fukuda, S.; Watanabe, D.; Ozawa, H.; Niwa, M.; Matsuo, T. In vitro analysis of drugs that improve hyperglycemia-induced blood-brain barrier dysfunction. Biochem. Biophys. Res. Commun. 2018, 503, 1885–1890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morofuji, Y.; Nakagawa, S.; So, G.; Hiu, T.; Horai, S.; Hayashi, K.; Tanaka, K.; Suyama, K.; Deli, M.A.; Nagata, I.; et al. Pitavastatin strengthens the barrier integrity in primary cultures of rat brain endothelial cells. Cell Mol. Neurobiol. 2010, 30, 727–735. [Google Scholar] [CrossRef] [Green Version]
- van den Boogaard, M.; Ramakers, B.P.; van Alfen, N.; van der Werf, S.P.; Fick, W.F.; Hoedemaekers, C.W.; Verbeek, M.M.; Schoonhoven, L.; van der Hoeven, J.G.; Pickkers, P. Endotoxemia-induced inflammation and the effect on the human brain. Crit. Care 2010, 14, R81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banks, W.A.; Kovac, A.; Morofuji, Y. Neurovascular unit crosstalk: Pericytes and astrocytes modify cytokine secretion patterns of brain endothelial cells. J. Cereb. Blood Flow Metab. 2018, 38, 1104–1118. [Google Scholar] [CrossRef] [PubMed]
- Kovac, A.; Erickson, M.A.; Banks, W.A. Brain microvascular pericytes are immunoactive in culture: Cytokine, chemokine, nitric oxide, and LRP-1 expression in response to lipopolysaccharide. J. Neuroinflamm. 2011, 8, 139. [Google Scholar] [CrossRef] [Green Version]
- Jangula, A.; Murphy, E.J. Lipopolysaccharide-induced blood brain barrier permeability is enhanced by alpha-synuclein expression. Neurosci. Lett. 2013, 551, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Cerejeira, J.; Firmino, H.; Vaz-Serra, A.; Mukaetova-Ladinska, E.B. The neuroinflammatory hypothesis of delirium. Acta Neuropathol. 2010, 119, 737–754. [Google Scholar] [CrossRef] [Green Version]
- Koladiya, R.U.; Jaggi, A.S.; Singh, N.; Sharma, B.K. Ameliorative role of Atorvastatin and Pitavastatin in L-Methionine induced vascular dementia in rats. BMC Pharmacol. 2008, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, T.W.; Zhang, S.; Wang, Y.T. Excitotoxicity and stroke: Identifying novel targets for neuroprotection. Prog. Neurobiol. 2014, 115, 157–188. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, C.; Tercero, I.; Pineda, A.; Burgos, J.S. Simvastatin is the statin that most efficiently protects against kainate-induced excitotoxicity and memory impairment. J. Alzheimers Dis. 2011, 24, 161–174. [Google Scholar] [CrossRef]
- Malfitano, A.M.; Marasco, G.; Proto, M.C.; Laezza, C.; Gazzerro, P.; Bifulco, M. Statins in neurological disorders: An overview and update. Pharmacol. Res. 2014, 88, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.M.; Kho, D.; Graham, E.S.; Nicholson, L.F.; O’Carroll, S.J. Statins Inhibit Fibrillary β-Amyloid Induced Inflammation in a Model of the Human Blood Brain Barrier. PLoS ONE 2016, 11, e0157483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.J.; Shyu, K.G.; Wang, B.W.; Lai, L.P. Simvastatin inhibits C-reactive protein-induced pro-inflammatory changes in endothelial cells by decreasing mevalonate pathway products. Cardiology 2008, 110, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Dichtl, W.; Dulak, J.; Frick, M.; Alber, H.F.; Schwarzacher, S.P.; Ares, M.P.; Nilsson, J.; Pachinger, O.; Weidinger, F. HMG-CoA reductase inhibitors regulate inflammatory transcription factors in human endothelial and vascular smooth muscle cells. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 58–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banks, W.A.; Gray, A.M.; Erickson, M.A.; Salameh, T.S.; Damodarasamy, M.; Sheibani, N.; Meabon, J.S.; Wing, E.E.; Morofuji, Y.; Cook, D.G.; et al. Lipopolysaccharide-induced blood-brain barrier disruption: Roles of cyclooxygenase, oxidative stress, neuroinflammation, and elements of the neurovascular unit. J. Neuroinflamm. 2015, 12, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyth, L.C.D.; Rustenhoven, J.; Park, T.I.; Schweder, P.; Jansson, D.; Heppner, P.A.; O’Carroll, S.J.; Mee, E.W.; Faull, R.L.M.; Curtis, M.; et al. Unique and shared inflammatory profiles of human brain endothelia and pericytes. J. Neuroinflamm. 2018, 15, 138. [Google Scholar] [CrossRef] [PubMed]
- Coisne, C.; Dehouck, L.; Faveeuw, C.; Delplace, Y.; Miller, F.; Landry, C.; Morissette, C.; Fenart, L.; Cecchelli, R.; Tremblay, P.; et al. Mouse syngenic in vitro blood-brain barrier model: A new tool to examine inflammatory events in cerebral endothelium. Lab. Investig. 2005, 85, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, S.; Deli, M.A.; Kawaguchi, H.; Shimizudani, T.; Shimono, T.; Kittel, A.; Tanaka, K.; Niwa, M. A new blood-brain barrier model using primary rat brain endothelial cells, pericytes and astrocytes. Neurochem. Int. 2009, 54, 253–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, K.D.; de Vellis, J. Preparation of separate astroglial and oligodendroglial cell cultures from rat cerebral tissue. J. Cell Biol. 1980, 85, 890–902. [Google Scholar] [CrossRef] [Green Version]
- Dehouck, M.P.; Méresse, S.; Delorme, P.; Fruchart, J.C.; Cecchelli, R. An easier, reproducible, and mass-production method to study the blood-brain barrier in vitro. J. Neurochem. 1990, 54, 1798–1801. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujimoto, T.; Morofuji, Y.; Kovac, A.; Erickson, M.A.; Deli, M.A.; Niwa, M.; Banks, W.A. Pitavastatin Ameliorates Lipopolysaccharide-Induced Blood-Brain Barrier Dysfunction. Biomedicines 2021, 9, 837. https://doi.org/10.3390/biomedicines9070837
Fujimoto T, Morofuji Y, Kovac A, Erickson MA, Deli MA, Niwa M, Banks WA. Pitavastatin Ameliorates Lipopolysaccharide-Induced Blood-Brain Barrier Dysfunction. Biomedicines. 2021; 9(7):837. https://doi.org/10.3390/biomedicines9070837
Chicago/Turabian StyleFujimoto, Takashi, Yoichi Morofuji, Andrej Kovac, Michelle A. Erickson, Mária A. Deli, Masami Niwa, and William A. Banks. 2021. "Pitavastatin Ameliorates Lipopolysaccharide-Induced Blood-Brain Barrier Dysfunction" Biomedicines 9, no. 7: 837. https://doi.org/10.3390/biomedicines9070837