
A Cell Membrane-Level Approach to Cicatricial Alopecia Management: Is Caveolin-1 a Viable Therapeutic Target in Frontal Fibrosing Alopecia?

 , ,
, , {kind=link}
{kind=link}
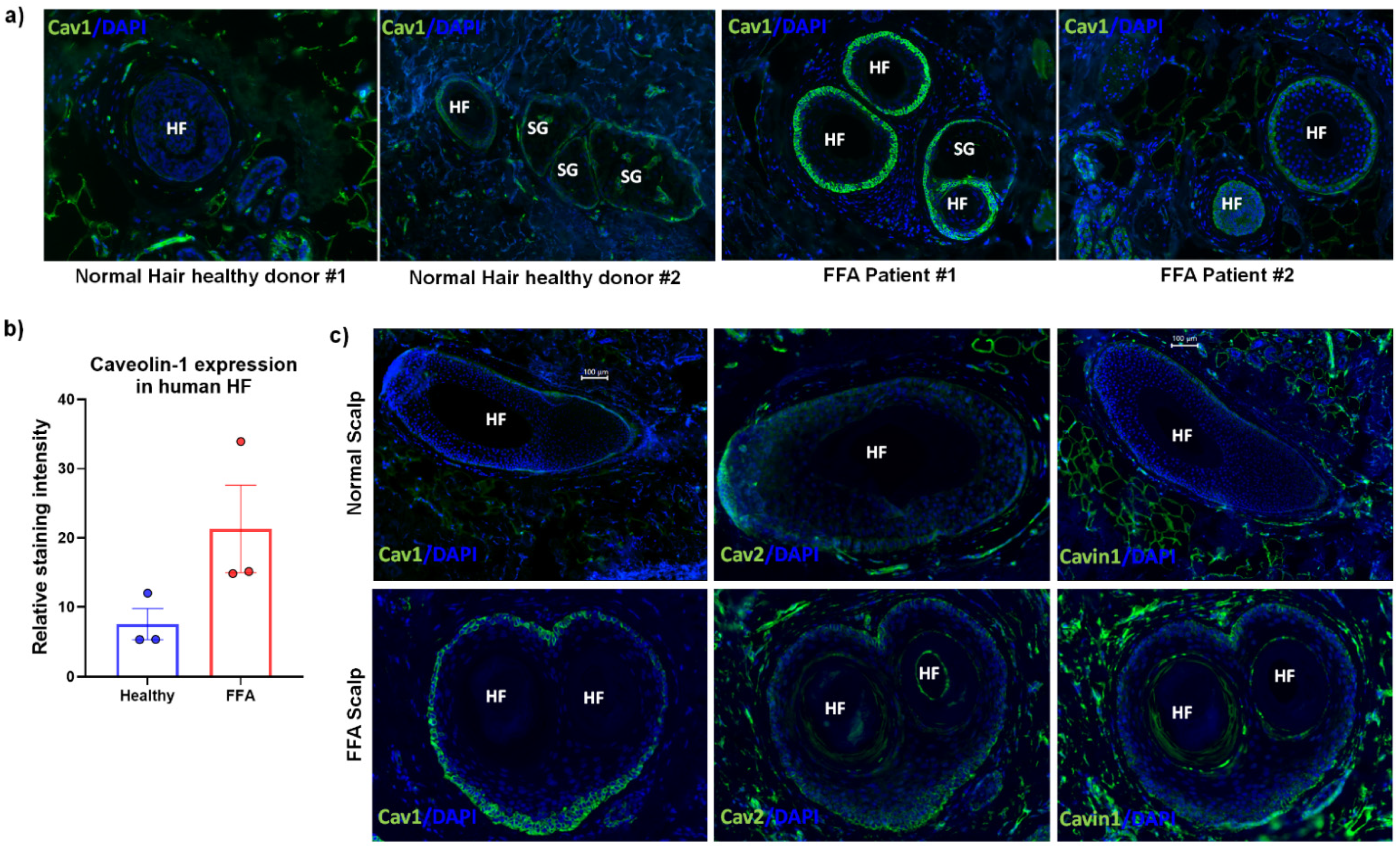{kind=link}
{kind=link}
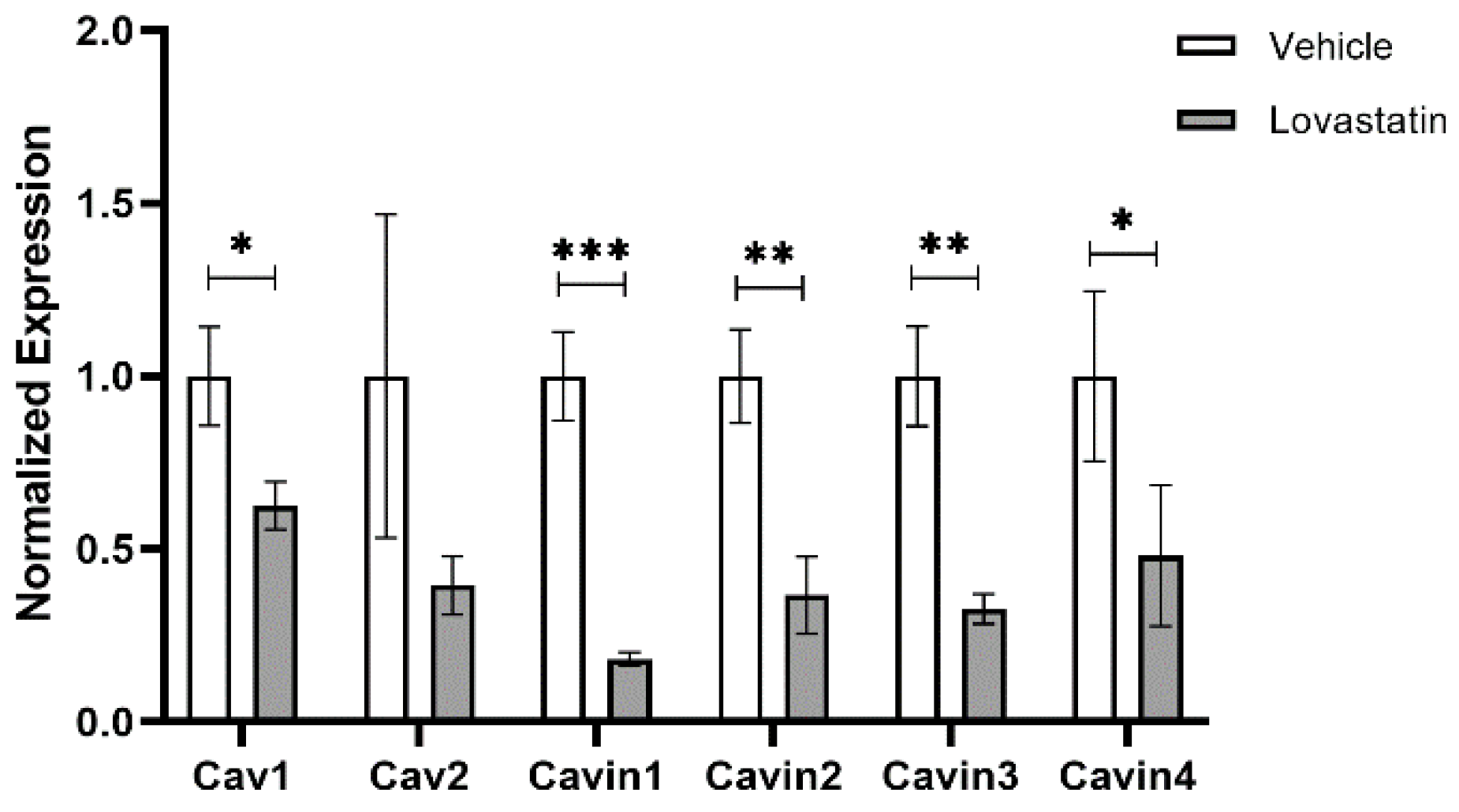{kind=link}
Abstract
:1. Background
2. Premises
3. Hypothesis
4. How to Probe the Hypothesis?
5. Translational Significance and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Harries, M.; Trüeb, R.; Tosti, A.; Messenger, A.; Chaudhry, I.; Whiting, D.; Sinclair, R.; Griffiths, C.; Paus, R. How not to get scar(r)ed: Pointers to the correct diagnosis in patients with suspected primary cicatricial alopecia. Br. J. Dermatol. 2009, 160, 482–501. [Google Scholar] [CrossRef]
- Doche, I.; Wilcox, G.L.; Ericson, M.; Valente, N.S.; Romiti, R.; McAdams, B.D. Hordinsky MK. Evidence for neurogenic inflammation in lichen planopilaris and frontal fibrosing alopecia patho-genic mechanism. Exp. Dermatol. 2020, 29, 282–285. [Google Scholar] [CrossRef]
- Egger, A.; Stojadinovic, O.; Miteva, M. Folliculitis Decalvans and Lichen Planopilaris Phenotypic Spectrum—A Series of 7 New Cases with Focus on Histopathology. Am. J. Dermatopathol. 2020, 42, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Miteva, M.; Torres, F.; Tosti, A. The ‘eyes’ or ‘goggles’ as a clue to the histopathological diagnosis of primary lymphocytic cicatricial alopecia. Br. J. Dermatol. 2011, 166, 454–455. [Google Scholar] [CrossRef]
- Miteva, M.; Tosti, A. The follicular triad: A pathological clue to the diagnosis of early frontal fibrosing alopecia. Br. J. Dermatol. 2012, 166, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Harries, M.J.; Jimenez, F.; Izeta, A.; Hardman, J.; Panicker, S.P.; Poblet, E.; Paus, R. Lichen Planopilaris and Frontal Fibrosing Alopecia as Model Epithelial Stem Cell Diseases. Trends Mol. Med. 2018, 24, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Harries, M.J.; Meyer, K.; Chaudhry, I.; Kloepper, J.; Poblet, E.; Griffiths, C.E.; Paus, R. Lichen planopilaris is characterized by immune privilege collapse of the hair follicle’s epithelial stem cell niche. J. Pathol. 2013, 231, 236–247. [Google Scholar] [CrossRef]
- Imanishi, H.; Ansell, D.M.; Chéret, J.; Harries, M.; Bertolini, M.; Sepp, N.; Bíró, T.; Poblet, E.; Jimenez, F.; Hardman, J.; et al. Epithelial-to-Mesenchymal Stem Cell Transition in a Human Organ: Lessons from Lichen Planopilaris. J. Investig. Dermatol. 2018, 138, 511–519. [Google Scholar] [CrossRef] [Green Version]
- Chiang, Y.; Bundy, C.; Griffiths, C.; Paus, R.; Harries, M. The role of beliefs: Lessons from a pilot study on illness perception, psychological distress and quality of life in patients with primary cicatricial alopecia. Br. J. Dermatol. 2015, 172, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Bolduc, C.; Sperling, L.C.; Shapiro, J. Primary cicatricial alopecia: Lymphocytic primary cicatricial alopecias, including chronic cutaneous lupus erythematosus, lichen planopilaris, frontal fibrosing alopecia, and Graham-Little syndrome. J. Am. Acad. Dermatol. 2016, 75, 1081–1099. [Google Scholar] [CrossRef]
- Olsen, E.A.; Bergfeld, W.F.; Cotsarelis, G.; Price, V.H.; Shapiro, J.; Sinclair, R.; Solomon, A.; Sperling, L.; Stenn, K.; Whiting, D.A. Summary of North American Hair Research Society (NAHRS)-sponsored Workshop on Cicatri-cial Alopecia, Duke University Medical Center, February 10 and 11, 2001. J. Am. Acad. Dermatol. 2003, 48, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Harries, M.J.; Sinclair, R.D.; Macdonald-Hull, S.; Whiting, D.A.; Griffiths, C.E.M.; Paus, R. Management of primary cicatricial alopecias: Options for treatment. Br. J. Dermatol. 2008, 159, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Lucky, A.W.; Piacquadio, D.J.; Ditre, C.M.; Dunlap, F.; Kantor, I.; Pandya, A.G.; Savin, R.C.; Tharp, M.D. A randomized, placebo-controlled trial of 5% and 2% topical minoxidil solutions in the treat-ment of female pattern hair loss. J. Am. Acad. Dermatol. 2004, 50, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.; Martinka, M.; Ball, N.; Shapiro, J. Primary cicatricial alopecias: Clinicopathology of 112 cases. J. Am. Acad. Dermatol. 2004, 50, 25–32. [Google Scholar] [CrossRef]
- Vañó-Galván, S.; Molina-Ruiz, A.M.; Serrano-Falcón, C.; Arias-Santiago, S.; Rodrigues-Barata, A.R.; Garnacho-Saucedo, G.; Martorell-Calatayud, A.; Fernández-Crehuet, P.; Grimalt, R.; Aranegui, B.; et al. Frontal fibrosing alopecia: A multicenter review of 355 patients. J. Am. Acad. Dermatol. 2014, 70, 670–678. [Google Scholar] [CrossRef]
- Jiménez, F.; Poblet, E. Is Hair Transplantation Indicated in Frontal Fibrosing Alopecia? The Results of Test Grafting in Three Patients. Dermatol. Surg. 2013, 39, 1115–1118. [Google Scholar] [CrossRef]
- Ho, A.; Shapiro, J. Medical therapy for frontal fibrosing alopecia: A review and clinical approach. J. Am. Acad. Dermatol. 2019, 81, 568–580. [Google Scholar] [CrossRef]
- Cheret, J.; Piccini, I.; Strutz-Seebohm, N.; Röpke, A.; Liashkovich, I.; Gosain, H.; Rieger, B.; Klingel, K.; Eggers, B.; Marcus, K.; et al. Preclinical evidence that the PPARgamma modulator, N-Acetyl-GED-0507-34-Levo, may protect human hair follicle epithelial stem cells against lichen planopilaris-associated damage. J. Eur. Acad. Dermatol. Venereol. 2020, 34, e195–e197. [Google Scholar] [CrossRef] [Green Version]
- Vañó-Galván, S.; Saceda-Corralo, D.; Blume-Peytavi, U.; Cucchía, J.; Dlova, N.C.; Dias, M.F.R.G.; Grimalt, R.; Guzmán-Sánchez, D.; Harries, M.; Ho, A.; et al. Frequency of the Types of Alopecia at Twenty-Two Specialist Hair Clinics: A Multicenter Study. Ski. Appendage Disord. 2019, 5, 309–315. [Google Scholar] [CrossRef]
- Griffin, L.; Michaelides, C.; Griffiths, C.; Paus, R.; Harries, M. Primary cicatricial alopecias: A U.K. survey. Br. J. Dermatol. 2012, 167, 694–697. [Google Scholar] [CrossRef]
- Holmes, S.M. Frontal Fibrosing Alopecia. J. Pigment. Disord. 2017, 4, 5–7. [Google Scholar] [CrossRef] [Green Version]
- Kossard, S. Postmenopausal frontal fibrosing alopecia. Scarring alopecia in a pattern distribution. Arch. Dermatol. 1994, 130, 770–774. [Google Scholar] [CrossRef]
- Kossard, S.; Lee, M.-S.; Wilkinson, B. Postmenopausal frontal fibrosing alopecia: A frontal variant of lichen planopilaris. J. Am. Acad. Dermatol. 1997, 36, 59–66. [Google Scholar] [CrossRef]
- Desai, K.; Almeida, B.; Miteva, M. Understanding Hormonal Therapies: Overview for the Dermatologist Focused on Hair. Dermatology 2021, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.M.; Randolph, M.; Rajabi-Estarabadi, A.; Keri, J.; Tosti, A. Hormonal Contraceptives and Dermatology. Am. J. Clin. Dermatol. 2021, 22, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Aldoori, N.; Dobson, K.; Holden, C.R.; McDonagh, A.J.; Harries, M.; Messenger, A.G. Frontal fibrosing alopecia: Possible association with leave-on facial skin care products and sun-screens; a questionnaire study. Br. J. Dermatol. 2016, 175, 762–767. [Google Scholar] [CrossRef]
- Moreno-Arrones, O.M.; Saceda-Corralo, D.; Rodrigues-Barata, A.R.; Castellanos-González, M.; Fernández-Pugnaire, M.A.; Grimalt, R.; Hermosa-Gelbard, A.; Bernárdez, C.; Molina-Ruiz, A.M.; Ormaechea-Pérez, N.; et al. Risk factors associated with frontal fibrosing alopecia: A multicentre case–control study. Clin. Exp. Dermatol. 2019, 44, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Debroy Kidambi, A.; Dobson, K.; Holmes, S.; Carauna, D.; Del Marmol, V.; Vujovic, A.; Kaur, M.R.; Takwale, A.; Farrant, P.; Champagne, C. Frontal fibrosing alopecia in men: An association with facial moisturizers and sun-screens. Br. J. Dermatol. 2017, 177, 260–261. [Google Scholar] [CrossRef] [Green Version]
- Prasad, S.; Marks, D.H.; Burns, L.J.; De Souza, B.; Flynn, E.A.; Scheinman, P.; Silvestri, D.; Yu, J.; LoSicco, K.; Senna, M.M. Patch testing and contact allergen avoidance in patients with lichen planopilaris and/or frontal fibrosing alopecia: A cohort study. J. Am. Acad. Dermatol. 2020, 83, 659–661. [Google Scholar] [CrossRef]
- Tziotzios, C.; Petridis, C.; Dand, N.; Ainali, C.; Saklatvala, J.R.; Pullabhatla, V.; Onoufriadis, A.; Pramanik, R.; Baudry, D.; Lee, S.H.; et al. Genome-wide association study in frontal fibrosing alopecia identifies four susceptibility loci including HLA-B*07: 02. Nat. Commun. 2019, 10, 1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McSweeney, S.M.; Christou, E.A.A.; Dand, N.; Boalch, A.; Holmes, S.; Harries, M.; Palamaras, I.; Cunningham, F.; Parkins, G.; Kaur, M. Frontal fibrosing alopecia: A descriptive cross-sectional study of 711 cases in female pa-tients from the UK. Br. J. Dermatol. 2020, 183, 1136–1138. [Google Scholar] [CrossRef]
- Chéret, J.; Bertolini, M.; Ponce, L.; Lehmann, J.; Tsai, T.; Alam, M.; Hatt, H.; Paus, R. Olfactory receptor OR2AT4 regulates human hair growth. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, F.; López, E.; Bertolini, M.; Alam, M.; Chéret, J.; Westgate, G.; Rinaldi, F.; Marzani, B.; Paus, R. Topical odorant application of the specific olfactory receptor OR2AT4 agonist, Sandalore®, improves telogen effluvium-associated parameters. J. Cosmet. Dermatol. 2021, 20, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Fessa, C.; Lim, P.; Kossard, S.; Richards, S.; Peñas, P.F. Lichen planus-like drug eruptions due to beta-blockers: A case report and literature review. Am. J. Clin. Dermatol. 2012, 13, 417–421. [Google Scholar] [CrossRef]
- Clayton, R.; Chaudhry, S.; Ali, I.; Cooper, S.; Hodgson, T.; Wojnarowska, F. Mucosal (oral and vulval) lichen planus in women: Are angiotensin-converting enzyme inhibitors protective, and beta-blockers and non-steroidal anti-inflammatory drugs associated with the condition? Clin. Exp. Dermatol. 2010, 35, 384–387. [Google Scholar] [CrossRef] [PubMed]
- Botchkarev, V.A.; Peters, E.M.; Botchkareva, N.V.; Maurer, M.; Paus, R. Hair Cycle-Dependent Changes in Adrenergic Skin Innervation, and Hair Growth Modulation by Adrenergic Drugs. J. Investig. Dermatol. 1999, 113, 878–887. [Google Scholar] [CrossRef] [Green Version]
- Gilhar, A.; Etzioni, A.; Paus, R. Alopecia areata. N. Engl. J. Med. 2012, 366, 1515–1525. [Google Scholar] [CrossRef] [Green Version]
- Pozdnyakova, O.; Mahalingam, M. Involvement of the bulge region in primary scarring alopecia. J. Cutan. Pathol. 2008, 35, 922–925. [Google Scholar] [CrossRef] [PubMed]
- Hoang, M.; Keady, M.; Mahalingam, M. Stem cell markers (cytokeratin 15, CD34 and nestin) in primary scarring and nonscarring alopecia. Br. J. Dermatol. 2009, 160, 609–615. [Google Scholar] [CrossRef]
- Harries, M.J.; Paus, R. The Pathogenesis of Primary Cicatricial Alopecias. Am. J. Pathol. 2010, 177, 2152–2162. [Google Scholar] [CrossRef] [PubMed]
- Purba, T.S.; Haslam, I.S.; Poblet, E.; Jiménez, F.; Gandarillas, A.; Izeta, A.; Paus, R. Human epithelial hair follicle stem cells and their progeny: Current state of knowledge, the widening gap in translational research and future challenges. BioEssays 2014, 36, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.C.; Klatte, J.E.; Dinh, H.V.; Harries, M.J.; Reithmayer, K.; Meyer, W.; Sinclair, R.; Paus, R. Evidence that the bulge region is a site of relative immune privilege in human hair follicles. Br. J. Dermatol. 2008, 159, 1077–1085. [Google Scholar] [CrossRef]
- Gonzales, K.A.U.; Fuchs, E. Skin and Its Regenerative Powers: An Alliance between Stem Cells and Their Niche. Dev. Cell 2017, 43, 387–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, H.; Mohri, Y.; Binh, N.T.; Morinaga, H.; Fukuda, M.; Ito, M.; Kurata, S.; Hoeijmakers, J.; Nishimura, E.K. Hair follicle aging is driven by transepidermal elimination of stem cells via COL17A1 pro-teolysis. Science 2016, 351, aad4395. [Google Scholar] [CrossRef]
- Zhang, B.; Ma, S.; Rachmin, I.; He, M.; Baral, P.; Choi, S.; Gonçalves, W.A.; Shwartz, Y.; Fast, E.M.; Su, Y.; et al. Hyperactivation of sympathetic nerves drives depletion of melanocyte stem cells. Nat. Cell Biol. 2020, 577, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Tokura, Y. Expression of Snail1 in the fibrotic dermis of postmenopausal frontal fibrosing al-opecia: Possible involvement of an epithelial-mesenchymal transition and a review of the Japanese patients. Br. J. Dermatol. 2010, 162, 1152–1154. [Google Scholar] [CrossRef]
- Gheldof, A.; Berx, G. Cadherins and Epithelial-to-Mesenchymal Transition. Prog. Mol. Biol. Transl. Sci. 2013, 116, 317–336. [Google Scholar] [CrossRef] [PubMed]
- Wheelock, M.J.; Shintani, Y.; Maeda, M.; Fukumoto, Y.; Johnson, K.R. Cadherin switching. J. Cell Sci. 2008, 121, 727–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egger, A.N.; Rajabi-Estarabadi, A.; Williams, N.M.; Resnik, S.R.; Fox, J.D.; Wong, L.L.; Jozic, I. The importance of caveolins and caveolae to dermatology: Lessons from the caves and beyond. Exp. Dermatol. 2020, 29, 136–148. [Google Scholar] [CrossRef]
- Kruglikov, I.L.; Scherer, P.E. Caveolin as a Universal Target in Dermatology. Int. J. Mol. Sci. 2019, 21, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codrici, E.; Albulescu, L.; Popescu, I.D.; Mihai, S.; Enciu, A.-M.; Albulescu, R.; Tanase, C.; Hinescu, M.E. Caveolin-1-Knockout Mouse as a Model of Inflammatory Diseases. J. Immunol. Res. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gai, X.; Lu, Z.; Tu, K.; Liang, Z.; Zheng, X. Caveolin-1 Is Up-Regulated by GLI1 and Contributes to GLI1-Driven EMT in Hepatocellular Carcinoma. PLoS ONE 2014, 9, e84551. [Google Scholar] [CrossRef]
- Han, F.; Zhang, L.; Zhou, Y.; Yi, X. Caveolin-1 regulates cell apoptosis and invasion ability in paclitaxel-induced multidrug-resistant A549 lung cancer cells. Int. J. Clin. Exp. Pathol. 2015, 8, 8937–8947. [Google Scholar] [PubMed]
- Tang, W.; Feng, X.; Zhang, S.; Ren, Z.; Liu, Y.; Yang, B.; Lv, B.; Cai, Y.; Xia, J.; Ge, N. Caveolin-1 Confers Resistance of Hepatoma Cells to Anoikis by Activating IGF-1 Pathway. Cell. Physiol. Biochem. 2015, 36, 1223–1236. [Google Scholar] [CrossRef] [PubMed]
- Jozic, I.; Sawaya, A.P.; Pastar, I.; Head, C.R.; Wong, L.L.; Glinos, G.D.; Wikramanayake, T.C.; Brem, H.; Kirsner, R.S.; Tomic-Canic, M. Pharmacological and Genetic Inhibition of Caveolin-1 Promotes Epithelialization and Wound Closure. Mol. Ther. 2019, 27, 1992–2004. [Google Scholar] [CrossRef]
- Sawaya, A.P.; Jozic, I.; Stone, R.C.; Pastar, I.; Egger, A.N.; Stojadinovic, O.; Glinos, G.D.; Kirsner, R.S.; Tomic-Canic, M. Mevastatin promotes healing by targeting caveolin-1 to restore EGFR signaling. JCI Insight 2019, 4, e129320. [Google Scholar] [CrossRef] [Green Version]
- Kruglikov, I.L.; Scherer, P.E. Caveolin-1 as a target in prevention and treatment of hypertrophic scarring. NPJ Regen. Med. 2019, 4, 1–7. [Google Scholar] [CrossRef]
- Kruglikov, I.L.; Scherer, P.E. Caveolin-1 as a pathophysiological factor and target in psoriasis. NPJ Aging Mech. Dis. 2019, 5, 1–7. [Google Scholar] [CrossRef]
- Kruglikov, I.L.; Zhang, Z.; Scherer, P.E. Caveolin-1 in skin aging—From innocent bystander to major contributor. Ageing Res. Rev. 2019, 55, 100959. [Google Scholar] [CrossRef]
- Rhim, J.H.; Kim, J.H.; Yeo, E.-J.; Park, S.C. Caveolin-1 as a Novel Indicator of Wound-Healing Capacity in Aged Human Corneal Epithelium. Mol. Med. 2010, 16, 527–534. [Google Scholar] [CrossRef]
- Abbasi, S.; Biernaskie, J. Injury modifies the fate of hair follicle dermal stem cell progeny in a hair cycle-dependent manner. Exp. Dermatol. 2019, 28, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Garcin, C.L.; Ansell, D.M.; Headon, D.J.; Paus, R.; Hardman, M.J. Hair Follicle Bulge Stem Cells Appear Dispensable for the Acute Phase of Wound Re-epithelialization. Stem Cells 2016, 34, 1377–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez, F.; Poblet, E.; Izeta, A. Reflections on how wound healing-promoting effects of the hair follicle can be translated into clinical practice. Exp. Dermatol. 2015, 24, 91–94. [Google Scholar] [CrossRef]
- Ohyama, M. Hair follicle bulge: A fascinating reservoir of epithelial stem cells. J. Dermatol. Sci. 2007, 46, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Purba, T.S.; Brunken, L.; Hawkshaw, N.J.; Peake, M.; Hardman, J.A.; Paus, R. A primer for studying cell cycle dynamics of the human hair follicle. Exp. Dermatol. 2016, 25, 663–668. [Google Scholar] [CrossRef] [Green Version]
- Purba, T.S.; Peake, M.; Farjo, B.; Farjo, N.; Bhogal, R.K.; Jenkins, G.; Paus, R. Divergent proliferation patterns of distinct human hair follicle epithelial progenitor niches in situ and their differential responsiveness to prostaglandin D2. Sci. Rep. 2017, 7, 15197. [Google Scholar] [CrossRef] [Green Version]
- Paus, R.; van der Veen, C.; Eichmüller, S.; Kopp, T.; Hagen, E.; Müller-Röver, S.; Hofmann, U. Generation and Cyclic Remodeling of the Hair Follicle Immune System in Mice. J. Investig. Dermatol. 1998, 111, 7–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paus, R.; Nickoloff, B.J.; Ito, T. A ’hairy’ privilege. Trends Immunol. 2005, 26, 32–40. [Google Scholar] [CrossRef]
- Christoph, T.; Müller-Röver, S.; Audring, H.; Tobin, D.J.; Hermes, B.; Cotsarelis, G.; Rückert, R.; Paus, R. The human hair follicle immune system: Cellular composition and immune privilege. Br. J. Dermatol. 2000, 142, 862–873. [Google Scholar] [CrossRef]
- He, K.; Yan, X.; Li, N.; Dang, S.; Xu, L.; Zhao, B.; Li, Z.; Lv, Z.; Fang, X.; Zhang, Y.; et al. Internalization of the TGF-beta type I receptor into caveolin-1 and EEA1 double-positive early endo-somes. Cell Res. 2015, 25, 738–752. [Google Scholar] [CrossRef] [Green Version]
- Razani, B.; Zhang, X.L.; Bitzer, M.; von Gersdorff, G.; Böttinger, E.P.; Lisanti, M.P. Caveolin-1 regulates transforming growth factor (TGF)-beta/SMAD signaling through an inter-action with the TGF-beta type I receptor. J. Biol. Chem. 2001, 276, 6727–6738. [Google Scholar] [CrossRef] [Green Version]
- Rückert, R.; Hofmann, U.; van der Veen, C.; Paus, R.; Bulfone-Paus, S. MHC Class I Expression in Murine Skin: Developmentally Controlled and Strikingly Restricted Intraepithelial Expression During Hair Follicle Morphogenesis and Cycling, and Response to Cytokine Treatment In Vivo. J. Investig. Dermatol. 1998, 111, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Ito, N.; Bettermann, A.; Tokura, Y.; Takigawa, M.; Paus, R. Collapse and restoration of MHC class-I-dependent immune privilege: Exploiting the human hair fol-licle as a model. Am. J. Pathol. 2004, 164, 623–634. [Google Scholar] [CrossRef] [Green Version]
- Peters, E.M.; Liotiri, S.; Bodó, E.; Hagen, E.; Bíró, T.; Arck, P.C.; Paus, R. Probing the effects of stress mediators on the human hair follicle: Substance P holds central po-sition. Am. J. Pathol. 2007, 171, 1872–1886. [Google Scholar] [CrossRef] [Green Version]
- Bertolini, M.; McElwee, K.; Gilhar, A.; Bulfone-Paus, S.; Paus, R. Hair follicle immune privilege and its collapse in alopecia areata. Exp. Dermatol. 2020, 29, 703–725. [Google Scholar] [CrossRef] [PubMed]
- Stang, E.; Kartenbeck, J.; Parton, R.G. Major histocompatibility complex class I molecules mediate association of SV40 with caveolae. Mol. Biol. Cell 1997, 8, 47–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oakley, F.D.; Smith, R.L.; Engelhardt, J.F. Lipid rafts and caveolin-1 coordinate interleukin-1beta (IL-1beta)-dependent activation of NFkappaB by controlling endocytosis of Nox2 and IL-1beta receptor 1 from the plasma membrane. J. Biol. Chem. 2009, 284, 33255–33264. [Google Scholar] [CrossRef] [Green Version]
- Tomassian, T.; Humphries, L.A.; Liu, S.D.; Silva, O.; Brooks, D.G.; Miceli, M.C. Caveolin-1 Orchestrates TCR Synaptic Polarity, Signal Specificity, and Function in CD8 T Cells. J. Immunol. 2011, 187, 2993–3002. [Google Scholar] [CrossRef] [Green Version]
- Monastyrskaya, K.; Hostettler, A.; Buergi, S.; Draeger, A. The NK1 Receptor Localizes to the Plasma Membrane Microdomains, and Its Activation Is Dependent on Lipid Raft Integrity. J. Biol. Chem. 2005, 280, 7135–7146. [Google Scholar] [CrossRef] [Green Version]
- Bae, G.D.; Park, E.-Y.; Kim, K.; Jang, S.-E.; Jun, H.-S.; Oh, Y.S. Upregulation of caveolin-1 and its colocalization with cytokine receptors contributes to beta cell apoptosis. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Ortega, S.; Varela-Guruceaga, M.; Algarabel, M.; Milagro, F.I.; Martínez, J.A.; De Miguel, C. Effect of TNF-Alpha on Caveolin-1 Expression and Insulin Signaling During Adipocyte Differentiation and in Mature Adipocytes. Cell. Physiol. Biochem. 2015, 36, 1499–1516. [Google Scholar] [CrossRef]
- Lotz, M.; Vaughan, J.H.; Carson, D. Effect of neuropeptides on production of inflammatory cytokines by human monocytes. Science 1988, 241, 1218–1221. [Google Scholar] [CrossRef] [PubMed]
- Ansel, J.C.; Brown, J.R.; Payan, D.G.; Brown, M. Substance P selectively activates TNF-alpha gene expression in murine mast cells. J. Immunol. 1993, 150, 4478–4485. [Google Scholar] [PubMed]
- Cipriani, G.; Serboiu, C.S.; Gherghiceanu, M.; Faussone-Pellegrini, M.S.; Vannucchi, M.G. NK receptors, Substance P, Ano1 expression and ultrastructural features of the muscle coat in Cav-1(-/-) mouse ileum. J. Cell Mol. Med. 2011, 15, 2411–2420. [Google Scholar] [CrossRef] [Green Version]
- Arck, P.C.; Handjiski, B.; Kuhlmei, A.; Peters, E.M.J.; Knackstedt, M.; Peter, A.; Hunt, S.P.; Klapp, B.F.; Paus, R. Mast cell deficient and neurokinin-1 receptor knockout mice are protected from stress-induced hair growth inhibition. J. Mol. Med. 2005, 83, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Bailey, K.M.; Liu, J. Caveolin-1 Up-regulation during Epithelial to Mesenchymal Transition Is Mediated by Focal Adhesion Kinase. J. Biol. Chem. 2008, 283, 13714–13724. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.; Hao, Z.; Han, J.L.; Zhu, D.J.; Jin, Z.F.; Xie, W.L. CAV-1 contributes to bladder cancer progression by inducing epithelial-to-mesenchymal transi-tion. Urol. Oncol. 2014, 32, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.-N.; Chen, Z.-X.; Liu, X.-C.; Liu, H.-Y.; Guan, G.-J.; Liu, G. Curcumin ameliorates epithelial-to-mesenchymal transition of podocytes in vivo and in vitro via regulating caveolin-1. Biomed. Pharmacother. 2014, 68, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Cokakli, M.; Erdal, E.; Nart, D.; Yilmaz, F.; Sagol, O.; Kilic, M.; Karademir, S.; Atabey, N. Differential expression of Caveolin-1 in hepatocellular carcinoma: Correlation with differentiation state, motility and invasion. BMC Cancer 2009, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Kaur, P.; Li, A. Adhesive Properties of Human Basal Epidermal Cells: An Analysis of Keratinocyte Stem Cells, Transit Amplifying Cells, and Postmitotic Differentiating Cells. J. Investig. Dermatol. 2000, 114, 413–420. [Google Scholar] [CrossRef] [Green Version]
- Samuelov, L.; Sprecher, E.; Tsuruta, D.; Bíró, T.; Kloepper, J.E.; Paus, R. P-cadherin regulates human hair growth and cycling via canonical Wnt signaling and trans-forming growth factor-beta2. J. Investig. Dermatol. 2012, 132, 2332–2341. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, K.; Bíró, T.; Tsuruta, D.; Tóth, B.I.; Kromminga, A.; Zákány, N.; Zimmer, A.; Funk, W.; Gibbs, B.F.; Zimmer, A.; et al. Endocannabinoids limit excessive mast cell maturation and activation in human skin. J. Allergy Clin. Immunol. 2012, 129, 726–738.e8. [Google Scholar] [CrossRef] [Green Version]
- Bertolini, M.; Pretzlaff, M.; Sulk, M.; Bähr, M.; Gherardini, J.; Uchida, Y.; Reibelt, M.; Kinori, M.; Rossi, A.; Bíró, T.; et al. Vasoactive intestinal peptide, whose receptor-mediated signalling may be defective in alopecia areata, provides protection from hair follicle immune privilege collapse. Br. J. Dermatol. 2016, 175, 531–541. [Google Scholar] [CrossRef]
- Subash, J.; Alexander, T.; Beamer, V.; McMichael, A. A proposed mechanism for central centrifugal cicatricial alopecia. Exp. Dermatol. 2020, 29, 190–195. [Google Scholar] [CrossRef] [Green Version]
- Lenart-Migdalska, A.; Drabik, L.; Kaźnica-Wiatr, M.; Tomkiewicz-Pająk, L.; Podolec, P.; Olszowska, M. Flow Cytometric Assessment of Endothelial and Platelet Microparticles in Patients with Atrial Fibrillation Treated with Dabigatran. Clin. Appl. Thromb. 2020, 26, 1076029620972467. [Google Scholar] [CrossRef] [PubMed]
- Gery, I.; Caspi, R.R. Tolerance Induction in Relation to the Eye. Front. Immunol. 2018, 9, 2304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keino, H.; Horie, S.; Sugita, S. Immune Privilege and Eye-Derived T-Regulatory Cells. J. Immunol. Res. 2018, 2018, 1–12. [Google Scholar] [CrossRef]
- Nikitorowicz-Buniak, J.; Denton, C.P.; Abraham, D.; Stratton, R. Partially Evoked Epithelial-Mesenchymal Transition (EMT) Is Associated with Increased TGFbeta Signaling within Lesional Scleroderma Skin. PLoS ONE 2015, 10, e0134092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzzi, F.; Zanchetta, D.; Cassoni, P.; Guzzi, V.; Francolini, M.; Parenti, M.; Chini, B. Localization of the human oxytocin receptor in caveolin-1 enriched domains turns the recep-tor-mediated inhibition of cell growth into a proliferative response. Oncogene 2002, 21, 1658–1667. [Google Scholar] [CrossRef] [Green Version]
- Burgueño, J.; Canela, E.I.; Mallol, J.; Lluís, C.; Franco, R.; Ciruela, F. Mutual regulation between metabotropic glutamate type 1α receptor and caveolin proteins: From traffick to constitutive activity. Exp. Cell Res. 2004, 300, 23–34. [Google Scholar] [CrossRef]
- Mahavadi, S.; Bhattacharya, S.; Kim, J.; Fayed, S.; Al-Shboul, O.; Grider, J.R.; Murthy, K.S. Caveolae-dependent internalization and homologous desensitization of VIP/PACAP receptor, VPAC2, in gastrointestinal smooth muscle. Peptides 2013, 43, 137–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, G.; Borroto-Escuela, D.O.; Fuxe, K.; Franco, R. Potential of caveolae in the therapy of cardiovascular and neurological diseases. Front. Physiol. 2014, 5, 370. [Google Scholar] [CrossRef] [Green Version]
- Moo, E.V.; van Senten, J.R.; Bräuner-Osborne, H.; Møller, T.C. Arrestin-Dependent and -Independent Internalization of G Protein-Coupled Receptors: Methods, Mechanisms, and Implications on Cell Signaling. Mol. Pharmacol. 2021, 99, 242–255. [Google Scholar] [CrossRef]
- Domingues, L.; Hurbain, I.; Gilles-Marsens, F.; Sirés-Campos, J.; André, N.; Dewulf, M.; Romao, M.; De Lesegno, C.V.; Macé, A.-S.; Blouin, C.; et al. Coupling of melanocyte signaling and mechanics by caveolae is required for human skin pigmentation. Nat. Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Busse, D.; Kudella, P.; Grüning, N.-M.; Gisselmann, G.; Ständer, S.; Luger, T.; Jacobsen, F.; Steinsträßer, L.; Paus, R.; Gkogkolou, P.; et al. A Synthetic Sandalwood Odorant Induces Wound-Healing Processes in Human Keratinocytes via the Olfactory Receptor OR2AT4. J. Investig. Dermatol. 2014, 134, 2823–2832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Tomisato, W.; Su, L.; Sun, L.; Choi, J.H.; Zhang, Z.; Wang, K.-W.; Zhan, X.; Choi, M.; Li, X.; et al. Skin-specific regulation of SREBP processing and lipid biosynthesis by glycerol kinase 5. Proc. Natl. Acad. Sci. USA 2017, 114, E5197–E5206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cakmak, S.; Gul, U.; Gönül, M.; Demiriz, M.; Cakmak, A. Statin Therapy and Diabetic Skin. Adv. Ther. 2008, 25, 17–22. [Google Scholar] [CrossRef]
- Feingold, K.R.; Man, M.Q.; Proksch, E.; Menon, G.K.; Brown, B.E.; Elias, P.M.; Mao-Qiang, M. The Lovastatin-Treated Rodent: A New Model of Barrier Disruption and Epidermal Hyperplasia. J. Investig. Dermatol. 1991, 96, 201–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jozic, I.; Chéret, J.; Abujamra, B.A.; Miteva, M.; Gherardini, J.; Paus, R. A Cell Membrane-Level Approach to Cicatricial Alopecia Management: Is Caveolin-1 a Viable Therapeutic Target in Frontal Fibrosing Alopecia? Biomedicines 2021, 9, 572. https://doi.org/10.3390/biomedicines9050572
Jozic I, Chéret J, Abujamra BA, Miteva M, Gherardini J, Paus R. A Cell Membrane-Level Approach to Cicatricial Alopecia Management: Is Caveolin-1 a Viable Therapeutic Target in Frontal Fibrosing Alopecia? Biomedicines. 2021; 9(5):572. https://doi.org/10.3390/biomedicines9050572
Chicago/Turabian StyleJozic, Ivan, Jérémy Chéret, Beatriz Abdo Abujamra, Mariya Miteva, Jennifer Gherardini, and Ralf Paus. 2021. "A Cell Membrane-Level Approach to Cicatricial Alopecia Management: Is Caveolin-1 a Viable Therapeutic Target in Frontal Fibrosing Alopecia?" Biomedicines 9, no. 5: 572. https://doi.org/10.3390/biomedicines9050572