M2 Monocyte Polarization in Dialyzed Patients Is Associated with Increased Levels of M-CSF and Myeloperoxidase-Associated Oxidative Stress: Preliminary Results

 , , ,
, , ,  and
and 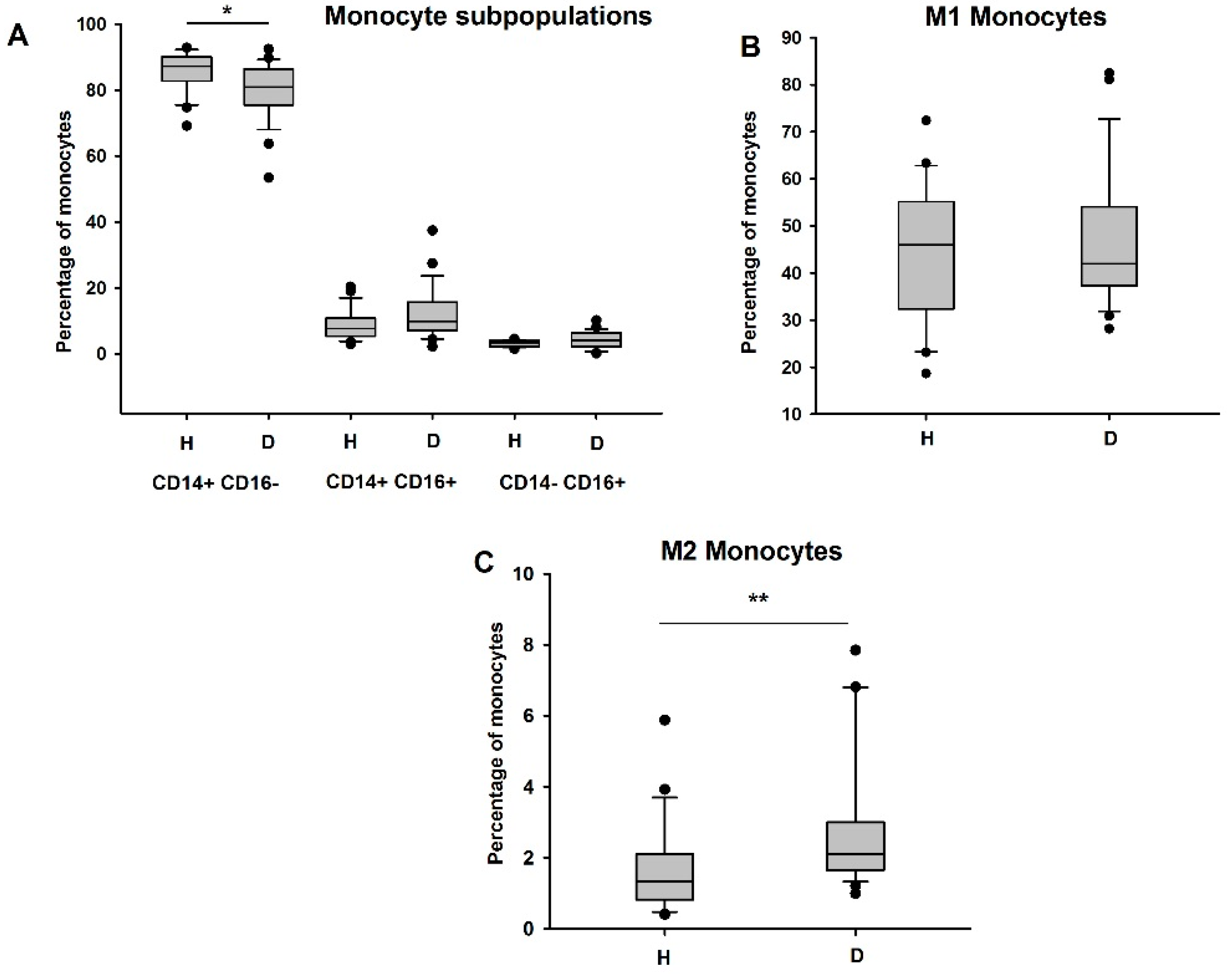{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Study Participants
2.2. Flow Cytometry
2.3. Measurements of Myeloperoxidase-Modified LDLs, M-CSF and IL-8 (ELISA Assays)
2.4. Quantitative Analysis of CRP, Apo-AI and Apo-B
2.5. Homocitrulline, Lysine (Lys), Chloro-Tyrosine (Cl-Tyr), and Tyrosine (Tyr) Quantification by LC-MS/MS
2.6. Statistical Analysis
3. Results
3.1. Volunteers and Patients
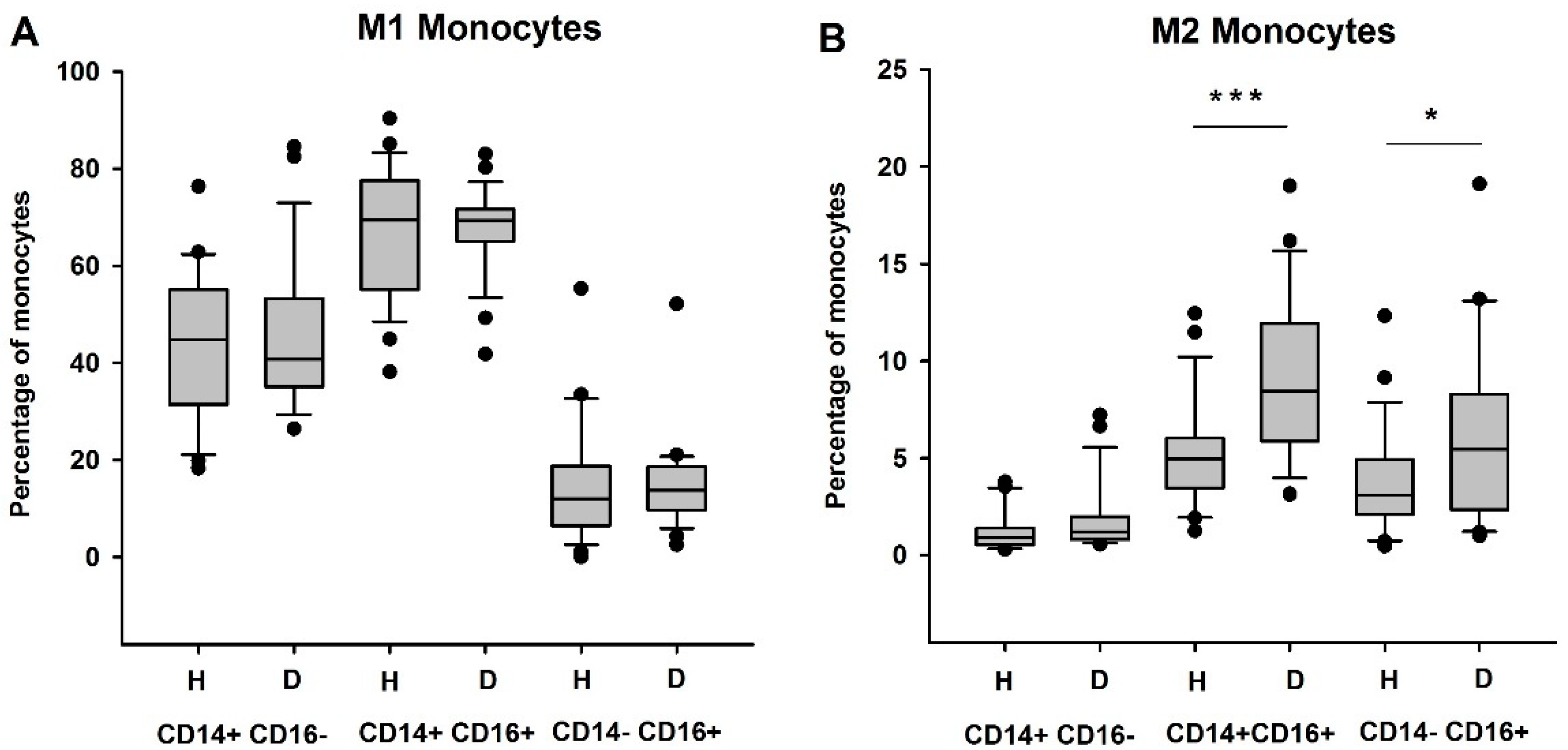
3.2. Circulating Anti-Inflammatory and Immunosuppressive Monocytes Are Increased in HD Patients
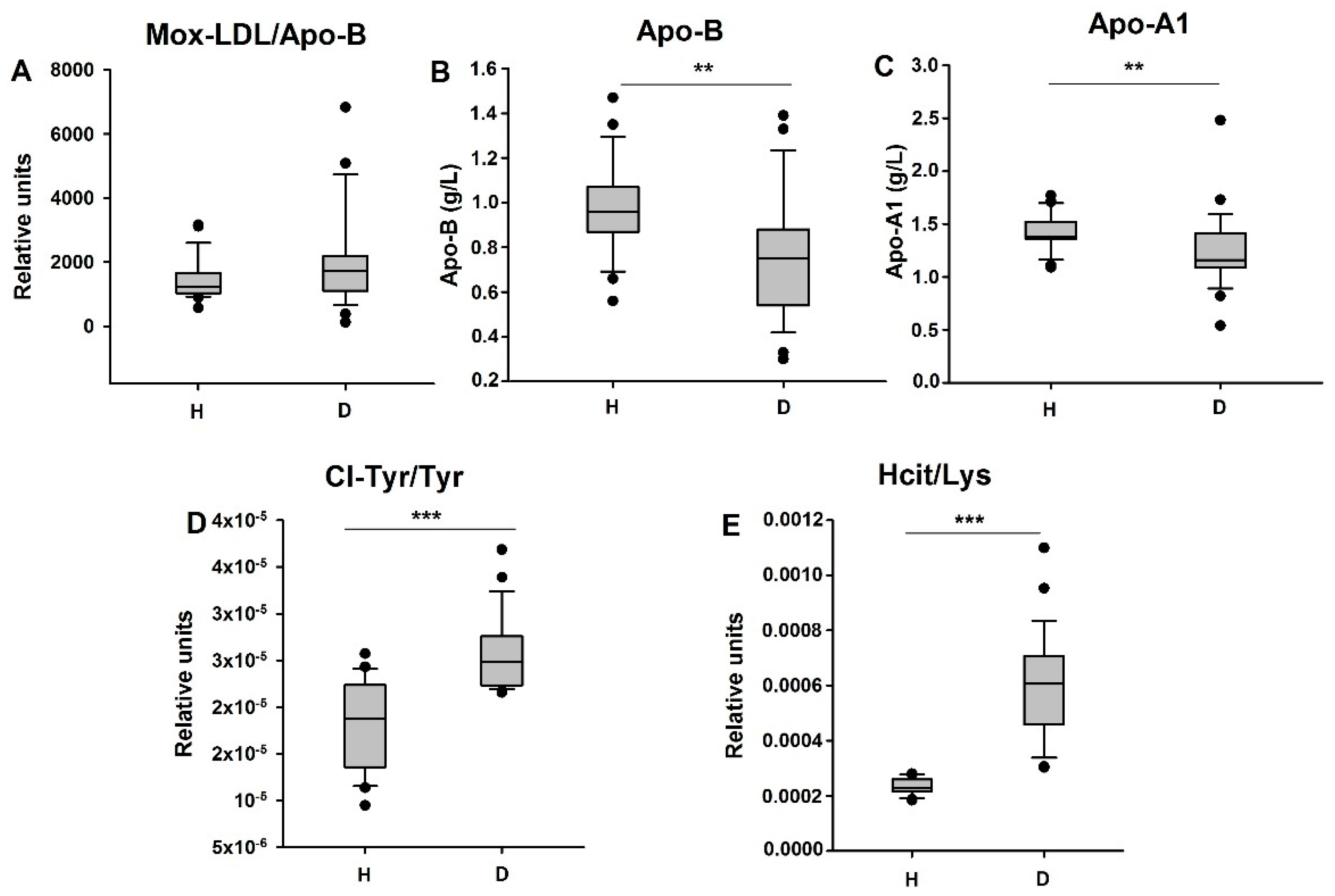
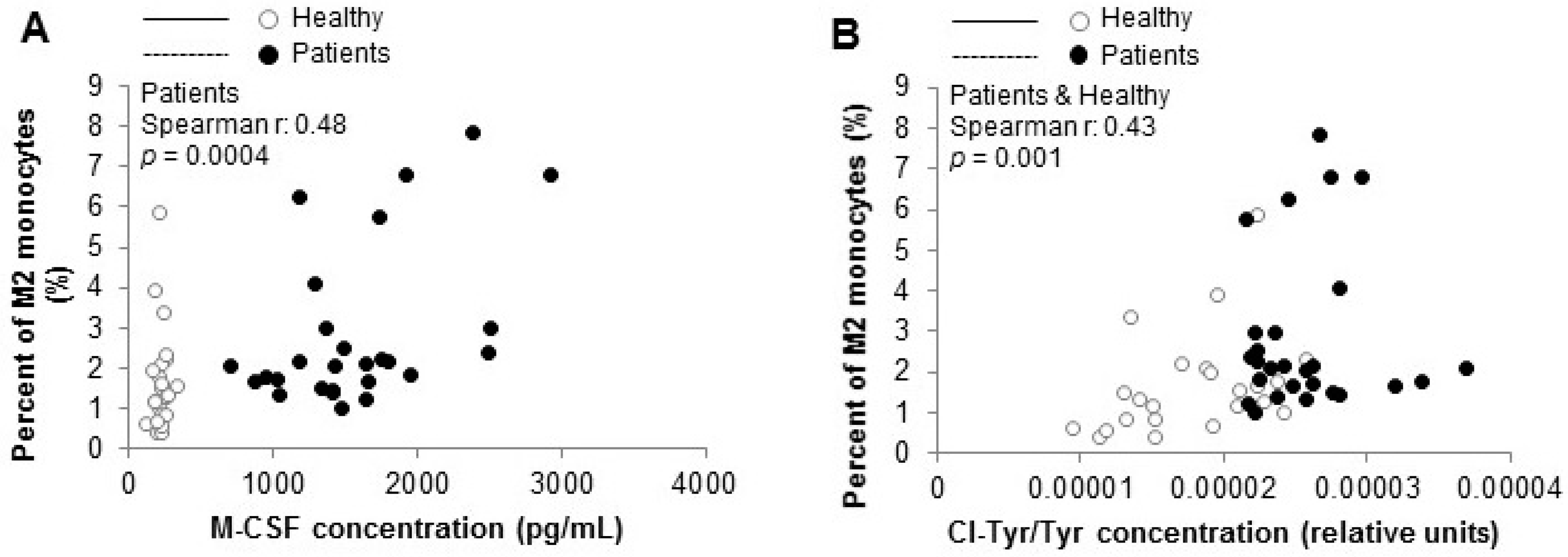
3.3. Biomarkers of Inflammation, Including M-CSF, Are Increased in HD Patients
3.4. HD Patients Undergo Myeloperoxidase-Dependent Oxidative Stress
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- USRDS. 2015 United States Renal Data System Annual Data Report. Available online: https://www.usrds.org/2015/view/Default.aspx (accessed on 19 September 2016).
- Walter, R.; Mischak, H.; Haller, H. Haemodialysis, atherosclerosis and inflammation-identifying molecular mechanisms of chronic vascular disease in ESDR patients. Nephrol. Dial. Transpl. 2002, 17 (Suppl. 3), 24–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawagishi, T.; Nishizawa, Y.; Konishi, T.; Kawasaki, K.; Emoto, M.; Shoji, T.; Tabata, T.; Inoue, T.; Morii, H. High-resolution B-mode ultrasonography in evaluation of atherosclerosis in uremia. Kidney Int. 1995, 48, 820–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, W.G.; Goldin, J.; Kuizon, B.D.; Yoon, C.; Gales, B.; Sider, D.; Wang, Y.; Chung, J.; Emerick, A.; Greaser, L.; et al. Coronary-artery calcification in young adults with end-stage renal disease who are undergoing dialysis. N. Engl. J. Med. 2000, 342, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Savage, T.; Clarke, A.L.; Giles, M.; Tomson, C.R.; Raine, A.E. Calcified plaque is common in the carotid and femoral arteries of dialysis patients without clinical vascular disease. Nephrol. Dial. Transpl. 1998, 13, 2004–2012. [Google Scholar] [CrossRef] [Green Version]
- Stenvinkel, P.; Heimburger, O.; Paultre, F.; Diczfalusy, U.; Wang, T.; Berglund, L.; Jogestrand, T. Strong association between malnutrition, inflammation, and atherosclerosis in chronic renal failure. Kidney Int. 1999, 55, 1899–1911. [Google Scholar] [CrossRef] [Green Version]
- Lusis, A.J. Atherosclerosis. Nature 2000, 407, 233–241. [Google Scholar] [CrossRef]
- Woollard, K.J.; Geissmann, F. Monocytes in atherosclerosis: Subsets and functions. Nat. Rev. Cardiol. 2010, 7, 77–86. [Google Scholar] [CrossRef]
- Tabas, I.; Garcia-Cardena, G.; Owens, G.K. Recent insights into the cellular biology of atherosclerosis. J. Cell Biol. 2015, 209, 13–22. [Google Scholar] [CrossRef]
- Fleetwood, A.J.; Dinh, H.; Cook, A.D.; Hertzog, P.J.; Hamilton, J.A. GM-CSF- and M-CSF-dependent macrophage phenotypes display differential dependence on type I interferon signaling. J. Leukoc. Biol. 2009, 86, 411–421. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Labonte, A.C.; Tosello-Trampont, A.C.; Hahn, Y.S. The role of macrophage polarization in infectious and inflammatory diseases. Mol. Cells 2014, 37, 275–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberale, L.; Dallegri, F.; Montecucco, F.; Carbone, F. Pathophysiological relevance of macrophage subsets in atherogenesis. Thromb. Haemost. 2017, 117, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Wolfs, I.M.; Donners, M.M.; de Winther, M.P. Differentiation factors and cytokines in the atherosclerotic plaque micro-environment as a trigger for macrophage polarisation. Thromb. Haemost. 2011, 106, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.C.; Zou, X.B.; Chai, Y.F.; Yao, Y.M. Macrophage polarization in inflammatory diseases. Int. J. Biol. Sci. 2014, 10, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K.; Chittezhath, M.; Shalova, I.N.; Lim, J.Y. Macrophage polarization and plasticity in health and disease. Immunol. Res. 2012, 53, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Martinez, F.O. Alternative activation of macrophages: Mechanism and functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, S.; Taylor, P.R. Monocyte and macrophage heterogeneity. Nat. Rev. Immunol. 2005, 5, 953–964. [Google Scholar] [CrossRef]
- Harmankaya, O.; Akalin, N.; Akay, H.; Okuturlar, Y.; Erturk, K.; Kaptanogullari, H.; Kocoglu, H. Comparison of risk factors for cardiovascular disease in hemodialysis and peritoneal dialysis patients. Clinics 2015, 70, 601–605. [Google Scholar] [CrossRef]
- Garcia-Lopez, E.; Carrero, J.J.; Suliman, M.E.; Lindholm, B.; Stenvinkel, P. Risk factors for cardiovascular disease in patients undergoing peritoneal dialysis. Perit. Dial. Int. 2007, 27 (Suppl. 2), S205–S209. [Google Scholar] [CrossRef]
- Turunen, S.; Koivula, M.-K.; Nicholas, A.; Risteli, L.; Risteli, J. Homocitrulline: An analog and confounder related to citrulline. In Protein Deimination in Human Health and Disease; Nicholas, A., Bhattacharya, S., Eds.; Springer Science Business Media LLC: New York, NY, USA, 2014; pp. 367–376. [Google Scholar]
- Moguilevsky, N.; Zouaoui Boudjeltia, K.; Babar, S.; Delree, P.; Legssyer, I.; Carpentier, Y.; Vanhaeverbeek, M.; Ducobu, J. Monoclonal antibodies against LDL progressively oxidized by myeloperoxidase react with ApoB-100 protein moiety and human atherosclerotic lesions. Biochem. Biophys. Res. Commun. 2004, 323, 1223–1228. [Google Scholar] [CrossRef] [PubMed]
- Delporte, C.; Franck, T.; Noyon, C.; Dufour, D.; Rousseau, A.; Madhoun, P.; Desmet, J.M.; Serteyn, D.; Raes, M.; Nortier, J.; et al. Simultaneous measurement of protein-bound 3-chlorotyrosine and homocitrulline by LC-MS/MS after hydrolysis assisted by microwave: Application to the study of myeloperoxidase activity during hemodialysis. Talanta 2012, 99, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Nishida, M.; Ando, M.; Iwamoto, Y.; Tsuchiya, K.; Nitta, K. New Insight into Atherosclerosis in Hemodialysis Patients: Overexpression of Scavenger Receptor and Macrophage Colony-Stimulating Factor Genes. Nephron Extra 2016, 6, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Nitta, K.; Akiba, T.; Kawashima, A.; Kimata, N.; Miwa, N.; Uchida, K.; Honda, K.; Takei, T.; Otsubo, S.; Yumura, W.; et al. Serum levels of macrophage colony-stimulating factor and aortic calcification in hemodialysis patients. Am. J. Nephrol. 2001, 21, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Mia, S.; Warnecke, A.; Zhang, X.M.; Malmstrom, V.; Harris, R.A. An optimized protocol for human M2 macrophages using M-CSF and IL-4/IL-10/TGF-beta yields a dominant immunosuppressive phenotype. Scand. J. Immunol. 2014, 79, 305–314. [Google Scholar] [CrossRef] [Green Version]
- Ito, A.; Shimokawa, H.; Meno, H.; Inou, T. Possible involvement of macrophage-colony stimulating factor in the pathogenesis of cardiac dysfunction in hemodialysis patients. Jpn. Heart J. 2004, 45, 497–503. [Google Scholar] [CrossRef] [Green Version]
- Chitu, V.; Stanley, E.R. Colony-stimulating factor-1 in immunity and inflammation. Curr. Opin. Immunol. 2006, 18, 39–48. [Google Scholar] [CrossRef]
- Hamilton, J.A. Colony-stimulating factors in inflammation and autoimmunity. Nat. Rev. Immunol. 2008, 8, 533–544. [Google Scholar] [CrossRef]
- Hume, D.A.; MacDonald, K.P. Therapeutic applications of macrophage colony-stimulating factor-1 (CSF-1) and antagonists of CSF-1 receptor (CSF-1R) signaling. Blood 2012, 119, 1810–1820. [Google Scholar] [CrossRef]
- Kihara, T.; Miyata, Y.; Furukawa, M.; Noguchi, M.; Nishikido, M.; Koga, S.; Kanetake, H. Predictive value of serum macrophage colony-stimulating factor for development of aortic calcification in haemodialysis patients: A 6 year longitudinal study. Nephrol. Dial. Transpl. 2005, 20, 1647–1652. [Google Scholar] [CrossRef] [Green Version]
- Clinton, S.K.; Underwood, R.; Hayes, L.; Sherman, M.L.; Kufe, D.W.; Libby, P. Macrophage colony-stimulating factor gene expression in vascular cells and in experimental and human atherosclerosis. Am. J. Pathol. 1992, 140, 301–316. [Google Scholar] [PubMed]
- Shyy, Y.J.; Wickham, L.L.; Hagan, J.P.; Hsieh, H.J.; Hu, Y.L.; Telian, S.H.; Valente, A.J.; Sung, K.L.; Chien, S. Human monocyte colony-stimulating factor stimulates the gene expression of monocyte chemotactic protein-1 and increases the adhesion of monocytes to endothelial monolayers. J. Clin. Investig. 1993, 92, 1745–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irvine, K.M.; Andrews, M.R.; Fernandez-Rojo, M.A.; Schroder, K.; Burns, C.J.; Su, S.; Wilks, A.F.; Parton, R.G.; Hume, D.A.; Sweet, M.J. Colony-stimulating factor-1 (CSF-1) delivers a proatherogenic signal to human macrophages. J. Leukoc. Biol. 2009, 85, 278–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, B.; Li, Y.; Buono, C.; Waldo, S.W.; Jones, N.L.; Mori, M.; Kruth, H.S. Constitutive receptor-independent low density lipoprotein uptake and cholesterol accumulation by macrophages differentiated from human monocytes with macrophage-colony-stimulating factor (M-CSF). J. Biol. Chem. 2006, 281, 15757–15762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.D.; Trogan, E.; Ginsberg, M.; Grigaux, C.; Tian, J.; Miyata, M. Decreased atherosclerosis in mice deficient in both macrophage colony-stimulating factor (op) and apolipoprotein E. Proc. Natl. Acad. Sci. USA 1995, 92, 8264–8268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devaraj, S.; Yun, J.M.; Duncan-Staley, C.; Jialal, I. C-reactive protein induces M-CSF release and macrophage proliferation. J. Leukoc. Biol. 2009, 85, 262–267. [Google Scholar] [CrossRef] [PubMed]
- N’Guessan, P.D.; Riediger, F.; Vardarova, K.; Scharf, S.; Eitel, J.; Opitz, B.; Slevogt, H.; Weichert, W.; Hocke, A.C.; Schmeck, B.; et al. Statins control oxidized LDL-mediated histone modifications and gene expression in cultured human endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 380–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryoo, S.W.; Kim, D.U.; Won, M.; Chung, K.S.; Jang, Y.J.; Oh, G.T.; Park, S.K.; Maeng, P.J.; Yoo, H.S.; Hoe, K.L. Native LDL induces interleukin-8 expression via H2O2, p38 Kinase, and activator protein-1 in human aortic smooth muscle cells. Cardiovasc. Res. 2004, 62, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.H.; Chang, C.L.; Yu, Y.S.; Lin, T.Y.; Chong, C.P.; Lin, Y.S.; Su, M.Y.; Yang, J.Y.; Shu, T.Y.; Lu, X.; et al. Chemical analysis of C-reactive protein synthesized by human aortic endothelial cells under oxidative stress. Anal. Chem. 2012, 84, 9646–9654. [Google Scholar] [CrossRef]
- Obradovic, M.M.; Trpkovic, A.; Bajic, V.; Soskic, S.; Jovanovic, A.; Stanimirovic, J.; Panic, M.; Isenovic, E.R. Interrelatedness between C-reactive protein and oxidized low-density lipoprotein. Clin. Chem. Lab. Med. 2015, 53, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Boudjeltia, K.Z.; Legssyer, I.; Van Antwerpen, P.; Kisoka, R.L.; Babar, S.; Moguilevsky, N.; Delree, P.; Ducobu, J.; Remacle, C.; Vanhaeverbeek, M.; et al. Triggering of inflammatory response by myeloperoxidase-oxidized LDL. Biochem. Cell Biol. 2006, 84, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Hazen, S.L.; Heinecke, J.W. 3-Chlorotyrosine, a specific marker of myeloperoxidase-catalyzed oxidation, is markedly elevated in low density lipoprotein isolated from human atherosclerotic intima. J. Clin. Investig. 1997, 99, 2075–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apostolov, E.O.; Shah, S.V.; Ok, E.; Basnakian, A.G. Carbamylated low-density lipoprotein induces monocyte adhesion to endothelial cells through intercellular adhesion molecule-1 and vascular cell adhesion molecule-1. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 826–832. [Google Scholar] [CrossRef] [Green Version]
- Holzer, M.; Gauster, M.; Pfeifer, T.; Wadsack, C.; Fauler, G.; Stiegler, P.; Koefeler, H.; Beubler, E.; Schuligoi, R.; Heinemann, A.; et al. Protein carbamylation renders high-density lipoprotein dysfunctional. Antioxid. Redox Signal. 2011, 14, 2337–2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desmons, A.; Jaisson, S.; Pietrement, C.; Rieu, P.; Wynckel, A.; Gillery, P. Homocitrulline: A new marker for differentiating acute from chronic renal failure. Clin. Chem. Lab. Med. 2016, 54, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Pireaux, V.; Sauvage, A.; Bihin, B.; Van Steenbrugge, M.; Rousseau, A.; Van Antwerpen, P.; Zouaoui Boudjeltia, K.; Raes, M. Myeloperoxidase-oxidized LDLs enhance an antiinflammatory M2 and antioxidant phenotype in murine macrophages. Mediat. Inflamm. 2016, 2016, 20. [Google Scholar] [CrossRef]
- Olmes, G.; Buttner-Herold, M.; Ferrazzi, F.; Distel, L.; Amann, K.; Daniel, C. CD163+ M2c-like macrophages predominate in renal biopsies from patients with lupus nephritis. Arthritis Res. Ther. 2016, 18, 90. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Ma, F.Y.; Tesch, G.H.; Manthey, C.L.; Nikolic-Paterson, D.J. Role of macrophages in the fibrotic phase of rat crescentic glomerulonephritis. Am. J. Physiol. Ren. Physiol. 2013, 304, F1043–F1053. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Cao, Q.; Zheng, D.; Sun, Y.; Wang, C.; Yu, X.; Wang, Y.; Lee, V.W.; Zheng, G.; Tan, T.K.; et al. Discrete functions of M2a and M2c macrophage subsets determine their relative efficacy in treating chronic kidney disease. Kidney Int. 2013, 84, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Khallou-Laschet, J.; Varthaman, A.; Fornasa, G.; Compain, C.; Gaston, A.T.; Clement, M.; Dussiot, M.; Levillain, O.; Graff-Dubois, S.; Nicoletti, A.; et al. Macrophage plasticity in experimental atherosclerosis. PLoS ONE 2010, 5, e8852. [Google Scholar] [CrossRef] [Green Version]
- Feig, J.E.; Parathath, S.; Rong, J.X.; Mick, S.L.; Vengrenyuk, Y.; Grauer, L.; Young, S.G.; Fisher, E.A. Reversal of hyperlipidemia with a genetic switch favorably affects the content and inflammatory state of macrophages in atherosclerotic plaques. Circulation 2011, 123, 989–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoger, J.L.; Gijbels, M.J.; van der Velden, S.; Manca, M.; van der Loos, C.M.; Biessen, E.A.; Daemen, M.J.; Lutgens, E.; de Winther, M.P. Distribution of macrophage polarization markers in human atherosclerosis. Atherosclerosis 2012, 225, 461–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pello, O.M.; Silvestre, C.; De Pizzol, M.; Andres, V. A glimpse on the phenomenon of macrophage polarization during atherosclerosis. Immunobiology 2011, 216, 1172–1176. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Lu, Y.; Zhou, G.; Liao, X.; Kapil, P.; Anand, P.; Mahabeleshwar, G.H.; Stamler, J.S.; Jain, M.K. Myeloid Kruppel-like factor 4 deficiency augments atherogenesis in ApoE-/- mice--brief report. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2836–2838. [Google Scholar] [CrossRef] [Green Version]
- Costantini, A.; Viola, N.; Beretta, A.; Galeazzi, R.; Matacchione, G.; Sabbatinelli, J.; Storci, G.; De Matteis, S.; Butini, L.; Rita Rippo, M.; et al. Age-related M1/M2 phenotype changes in circulating monocytes from healthy/unhealthy individuals. Aging 2018, 10, 1268–1280. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Alabdullah, M.; Grossmann, J.; Spieler, F.; Abdosh, R.; Lutz, V.; Kamies, K.; Knöpp, K.; Rieckmann, M.; Koch, S.; et al. The differential statin effect on cytokine production of monocytes or macrophages is mediated by differential genarylgenarylation-dependent Rac1 activation. Cell Death Dis. 2019, 10, 880. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pireaux, V.; Delporte, C.; Rousseau, A.; Desmet, J.-M.; Van Antwerpen, P.; Raes, M.; Zouaoui Boudjeltia, K. M2 Monocyte Polarization in Dialyzed Patients Is Associated with Increased Levels of M-CSF and Myeloperoxidase-Associated Oxidative Stress: Preliminary Results. Biomedicines 2021, 9, 84. https://doi.org/10.3390/biomedicines9010084
Pireaux V, Delporte C, Rousseau A, Desmet J-M, Van Antwerpen P, Raes M, Zouaoui Boudjeltia K. M2 Monocyte Polarization in Dialyzed Patients Is Associated with Increased Levels of M-CSF and Myeloperoxidase-Associated Oxidative Stress: Preliminary Results. Biomedicines. 2021; 9(1):84. https://doi.org/10.3390/biomedicines9010084
Chicago/Turabian StylePireaux, Valérie, Cédric Delporte, Alexandre Rousseau, Jean-Marc Desmet, Pierre Van Antwerpen, Martine Raes, and Karim Zouaoui Boudjeltia. 2021. "M2 Monocyte Polarization in Dialyzed Patients Is Associated with Increased Levels of M-CSF and Myeloperoxidase-Associated Oxidative Stress: Preliminary Results" Biomedicines 9, no. 1: 84. https://doi.org/10.3390/biomedicines9010084