Regulation of Oncogenic Targets by the Tumor-Suppressive miR-139 Duplex (miR-139-5p and miR-139-3p) in Renal Cell Carcinoma


Abstract
:1. Introduction
2. Materials and Methods
2.1. Human RCC Cell Lines
2.2. RNA Extraction and Quantitative Reverse Transcription-Polymerase Chain Reaction
2.3. Transfection of miRNAs, siRNAs, and Plasmid Vectors into RCC cells
2.4. Functional Assays (Cell Proliferation, Migration, and Invasion Assays) in RCC cells
2.5. Identification of miR-139-5p and miR-139-3p Gene Targets in RCC Cells
2.6. In silico Analysis of RCC Public Databases
2.7. Clinicopathological Analysis of RCC
2.8. Plasmid Construction and Dual-Luciferase Reporter Assays
2.9. Western Blotting
2.10. Statistical Analyses
3. Results
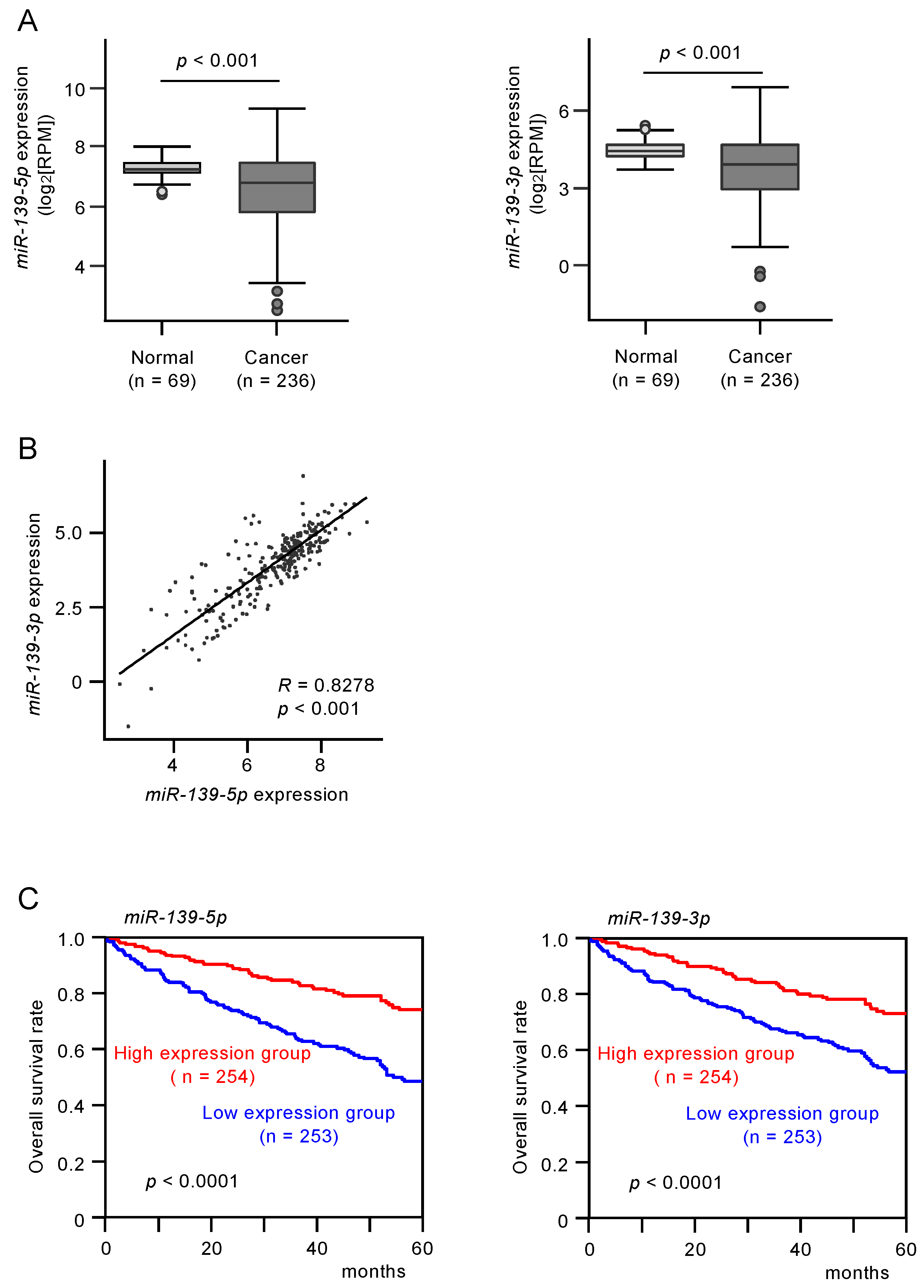
3.1. Analysis of miR-139-5p and miR-139-3p Expression Levels in Clinical RCC Tissues and Their Clinical Significance
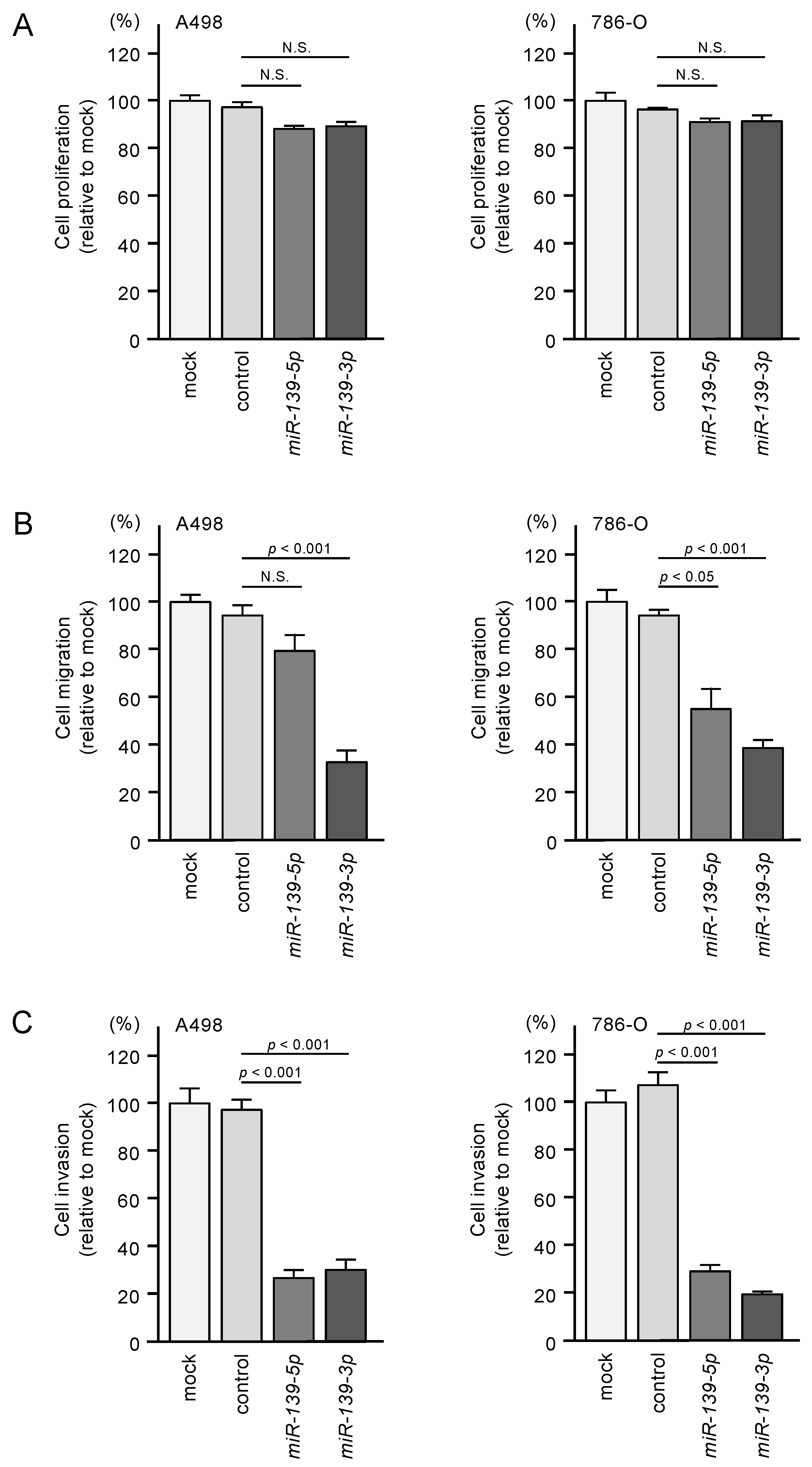
3.2. Tumor-Suppressive Functions of miR-139-5p and miR-139-3p in RCC Cells
3.3. Identification of Putative Oncogenic Targets Regulated by miR-139-3p and miR-139-3p in RCC Cells
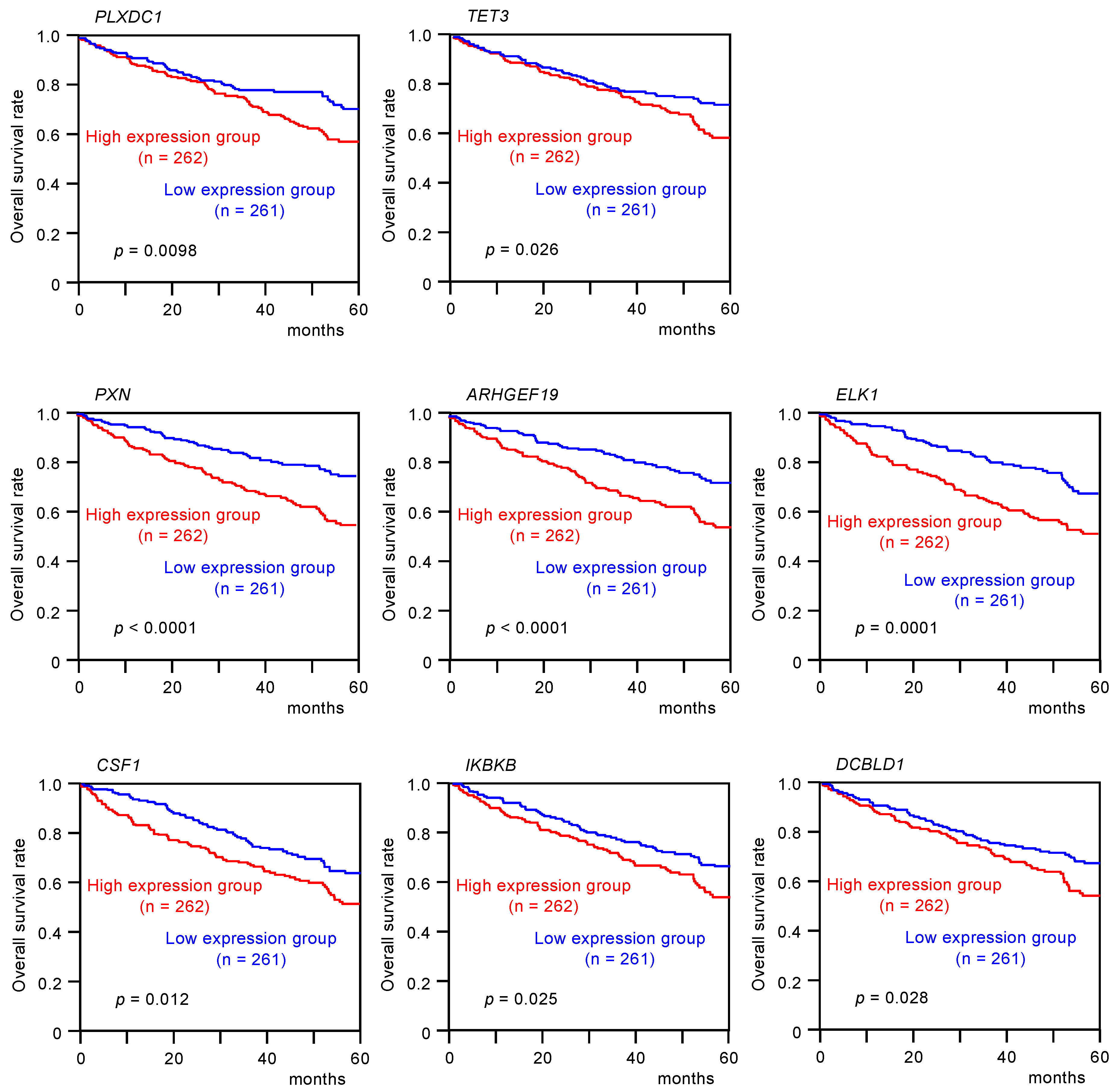
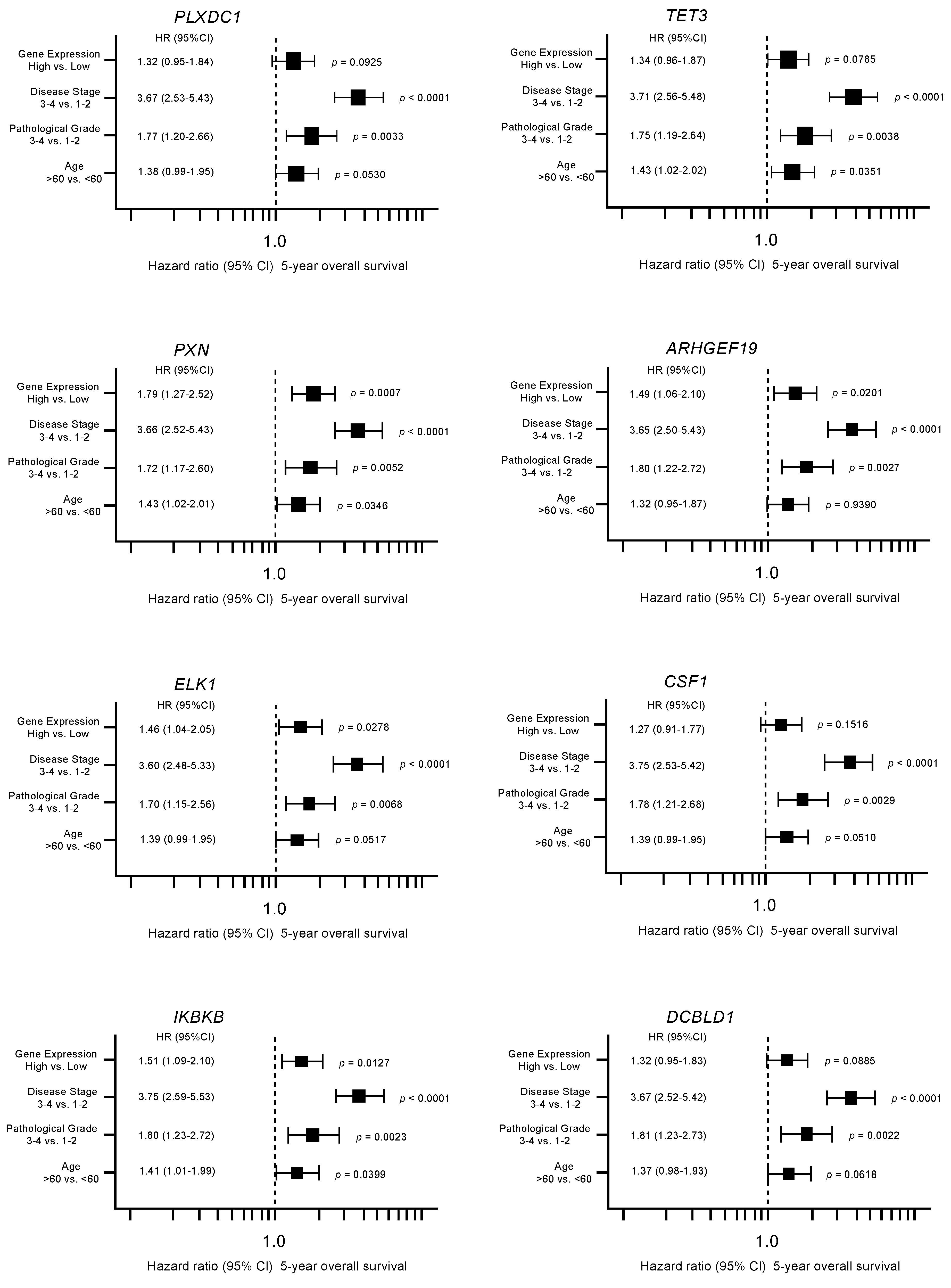
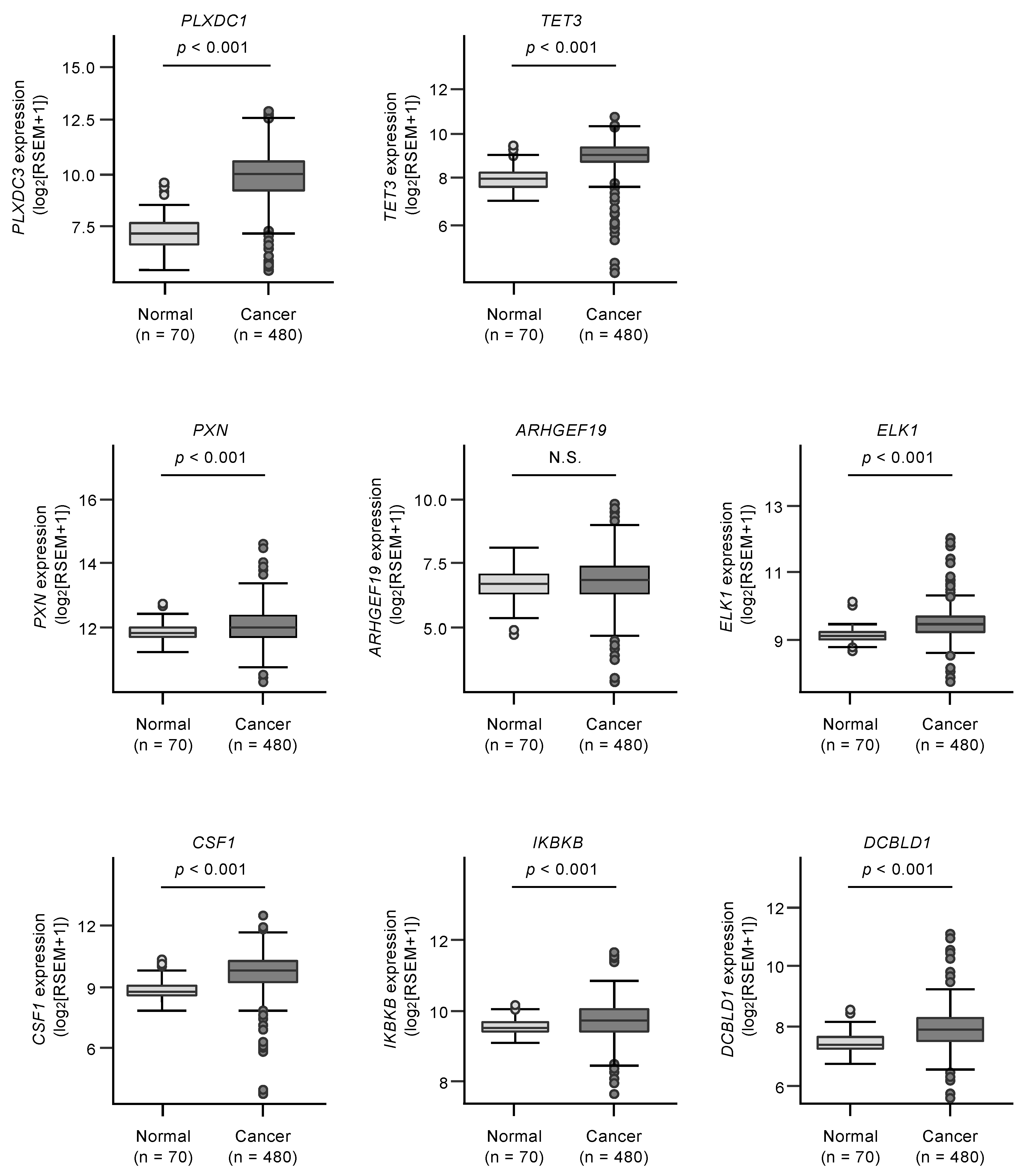
3.4. Clinical Significance of miR-139 Target Genes in RCC Pathogenesis
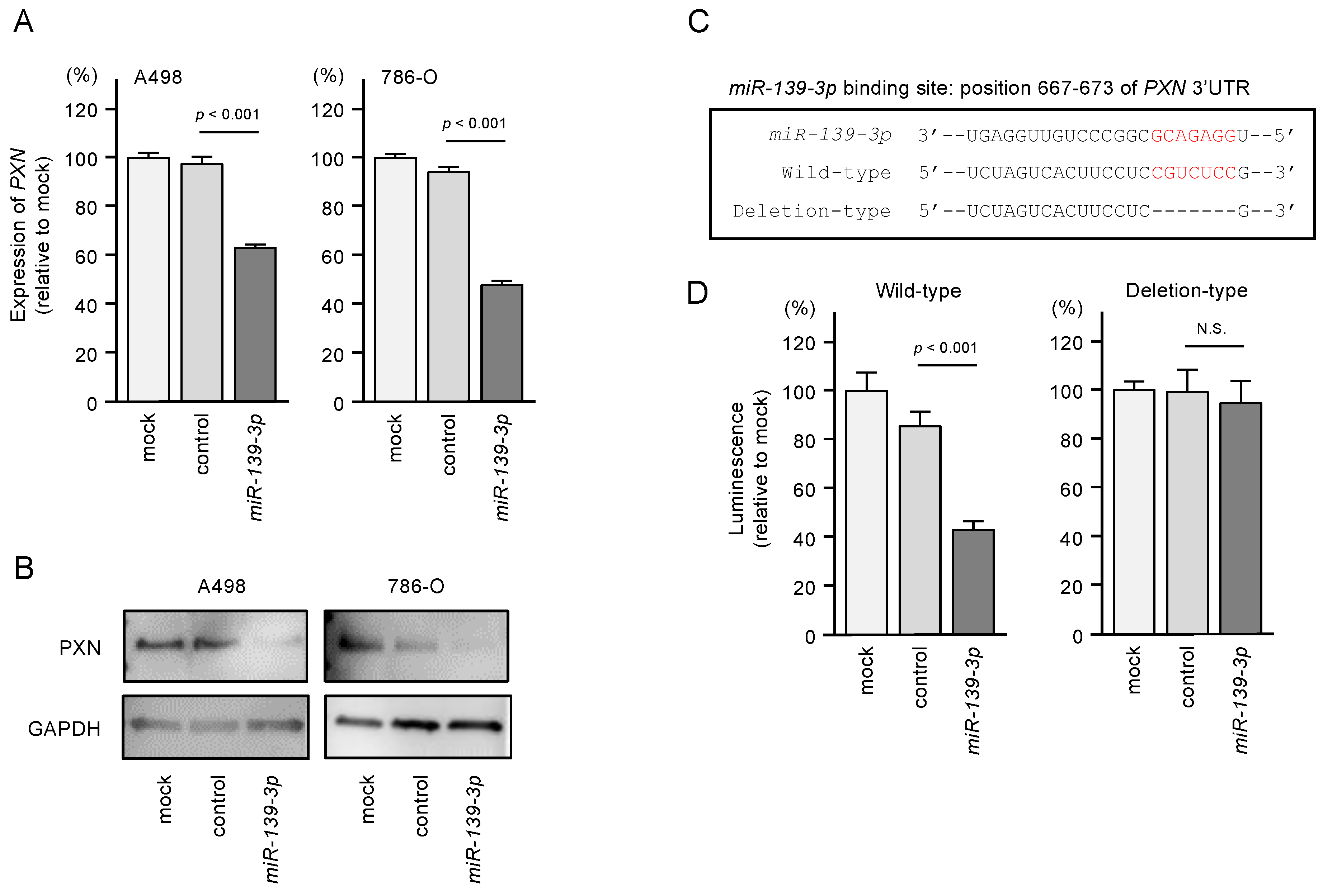
3.5. Direct Regulation of PXN by miR-139-3p in RCC Cells
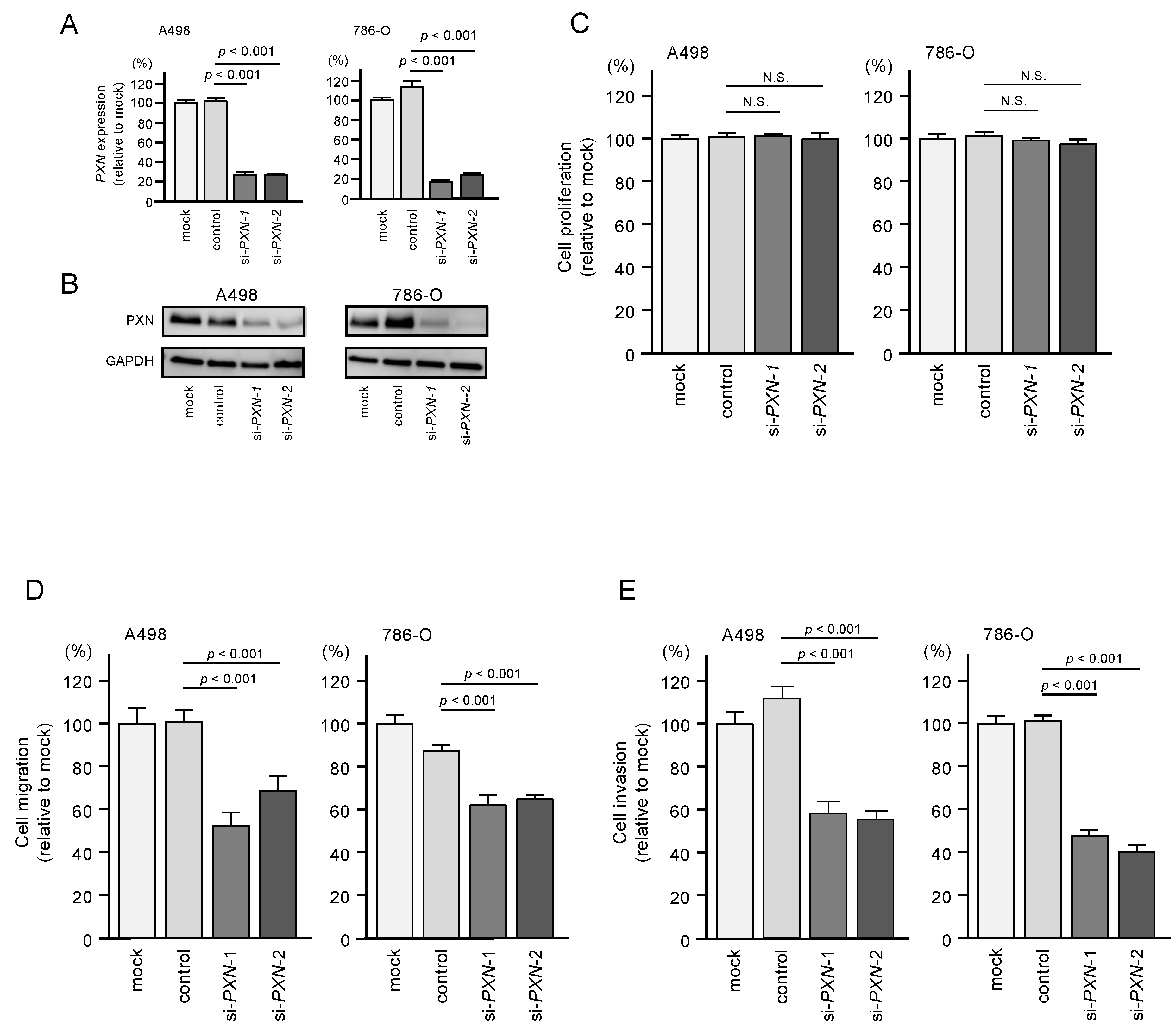
3.6. PXN Knockdown Assays in RCC Cells
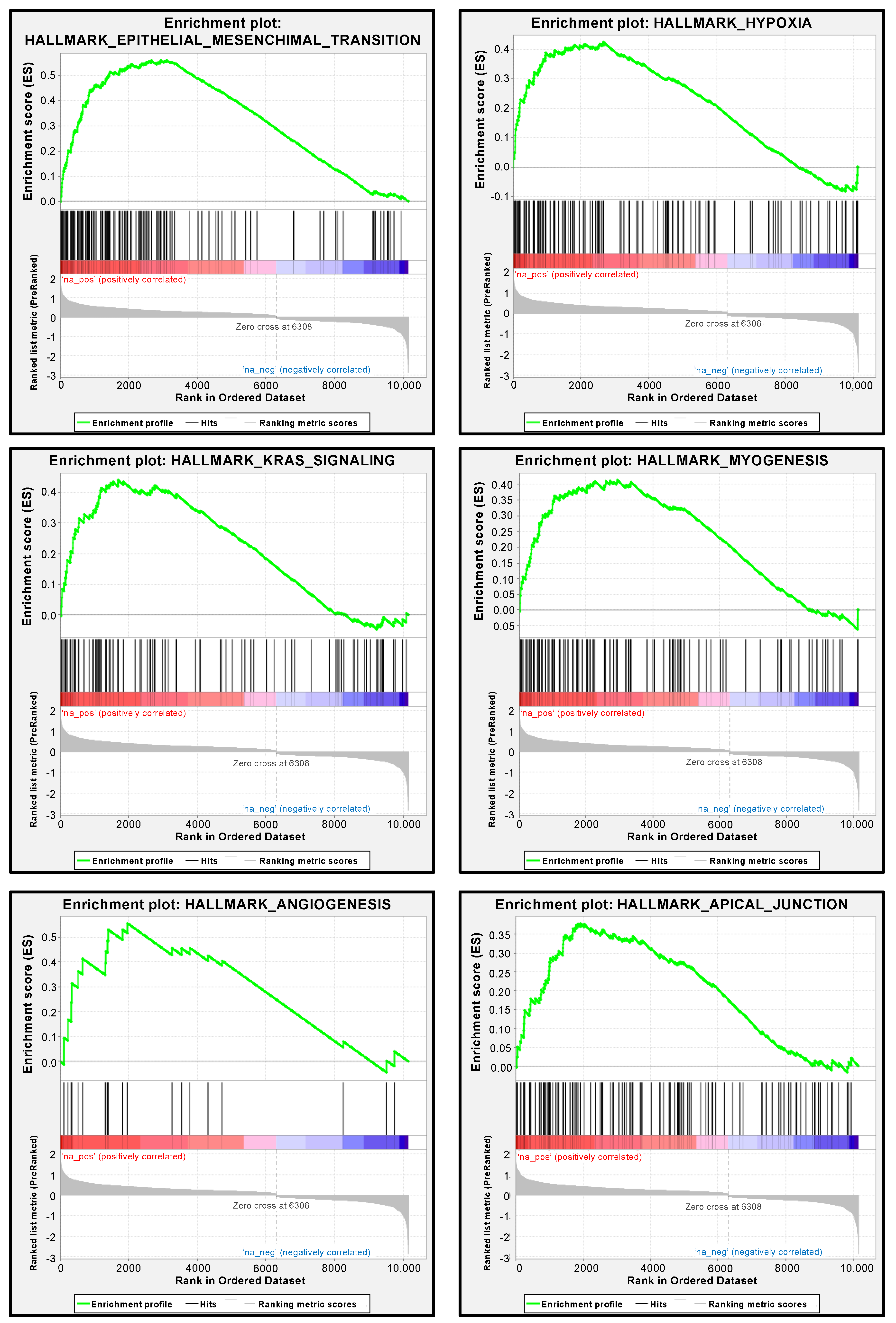
3.7. PXN-Mediated Pathways in RCC Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierorazio, P.M.; Johnson, M.H.; Patel, H.D.; Sozio, S.M.; Sharma, R.; Iyoha, E.; Bass, E.B.; Allaf, M.E. Management of renal masses and localized renal cancer: Systematic review and meta-analysis. J. Urol. 2016, 196, 989–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cairns, P. Renal cell carcinoma. Cancer Biomark 2010, 9, 461–473. [Google Scholar] [CrossRef]
- Cohen, H.T.; McGovern, F.J. Renal-cell carcinoma. N. Engl. J. Med. 2005, 353, 2477–2490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs-an update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Goto, Y.; Kurozumi, A.; Arai, T.; Nohata, N.; Kojima, S.; Okato, A.; Kato, M.; Yamazaki, K.; Ishida, Y.; Naya, Y.; et al. Impact of novel miR-145-3p regulatory networks on survival in patients with castration-resistant prostate cancer. Br. J. Cancer 2017, 117, 409–420. [Google Scholar] [CrossRef]
- Toda, H.; Seki, N.; Kurozumi, S.; Shinden, Y.; Yamada, Y.; Nohata, N.; Moriya, S.; Idichi, T.; Maemura, K.; Fujii, T.; et al. RNA-sequence-based microRNA expression signature in breast cancer: Tumor-suppressive miR-101-5p regulates molecular pathogenesis. Mol. Oncol. 2020, 14, 426–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, M.; Goto, Y.; Tanaka, T.; Okada, R.; Moriya, S.; Idichi, T.; Noda, M.; Sasaki, K.; Kita, Y.; Kurahara, H.; et al. RNA sequencing-based microRNA expression signature in esophageal squamous cell carcinoma: Oncogenic targets by antitumor miR-143-5p and miR-143-3p regulation. J. Hum. Genet. 2020, 65, 1019–1034. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Okada, R.; Hozaka, Y.; Wada, M.; Moriya, S.; Satake, S.; Idichi, T.; Kurahara, H.; Ohtsuka, T.; Seki, N. Molecular pathogenesis of pancreatic ductal adenocarcinoma: Impact of miR-30c-5p and miR-30c-2-3p regulation on oncogenic genes. Cancers (Basel) 2020, 12, 2731. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Tanigawa, K.; Nohata, N.; Misono, S.; Okada, R.; Asai, S.; Moriya, S.; Suetsugu, T.; Inoue, H.; Seki, N. FAM64A: A novel oncogenic target of lung adenocarcinoma regulated by both strands of miR-99a (miR-99a-5p and miR-99a-3p). Cells 2020, 9, 2083. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Nohata, N.; Uchida, A.; Kato, M.; Arai, T.; Moriya, S.; Mizuno, K.; Kojima, S.; Yamazaki, K.; Naya, Y.; et al. Replisome genes regulation by antitumor miR-101-5p in clear cell renal cell carcinoma. Cancer Sci. 2020, 111, 1392–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misono, S.; Seki, N.; Mizuno, K.; Yamada, Y.; Uchida, A.; Sanada, H.; Moriya, S.; Kikkawa, N.; Kumamoto, T.; Suetsugu, T.; et al. Molecular pathogenesis of gene regulation by the miR-150 duplex: miR-150-3p regulates TNS4 in lung adenocarcinoma. Cancers (Basel) 2019, 11, 601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, R.; Adams, C.M.; Jiang, W.; Greenawalt, E.; Eischen, C.M. Pan-cancer analysis reveals cooperativity of both strands of microRNA that regulate tumorigenesis and patient survival. Nat. Commun. 2020, 11, 968. [Google Scholar] [CrossRef] [Green Version]
- Alpha, K.M.; Xu, W.; Turner, C.E. Paxillin family of focal adhesion adaptor proteins and regulation of cancer cell invasion. Int. Rev. Cell Mol. Biol. 2020, 355, 1–52. [Google Scholar] [CrossRef]
- Noh, K.; Bach, D.H.; Choi, H.J.; Kim, M.S.; Wu, S.Y.; Pradeep, S.; Ivan, C.; Cho, M.S.; Bayraktar, E.; Rodriguez-Aguayo, C.; et al. The hidden role of paxillin: Localization to nucleus promotes tumor angiogenesis. Oncogene 2020. [Google Scholar] [CrossRef]
- Khalili, N.; Nouri-Vaskeh, M.; Hasanpour Segherlou, Z.; Baghbanzadeh, A.; Halimi, M.; Rezaee, H.; Baradaran, B. Diagnostic, prognostic, and therapeutic significance of miR-139-5p in cancers. Life Sci. 2020, 256, 117865. [Google Scholar] [CrossRef]
- Wu, X.; Weng, L.; Li, X.; Guo, C.; Pal, S.K.; Jin, J.M.; Li, Y.; Nelson, R.A.; Mu, B.; Onami, S.H.; et al. Identification of a 4-microRNA signature for clear cell renal cell carcinoma metastasis and prognosis. PLoS ONE 2012, 7, e35661. [Google Scholar] [CrossRef] [PubMed]
- Fridman, E.; Dotan, Z.; Barshack, I.; David, M.B.; Dov, A.; Tabak, S.; Zion, O.; Benjamin, S.; Benjamin, H.; Kuker, H.; et al. Accurate molecular classification of renal tumors using microRNA expression. J. Mol. Diagn 2010, 12, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Catanzaro, G.; Besharat, Z.M.; Miele, E.; Chiacchiarini, M.; Po, A.; Carai, A.; Marras, C.E.; Antonelli, M.; Badiali, M.; Raso, A.; et al. The miR-139-5p regulates proliferation of supratentorial paediatric low-grade gliomas by targeting the PI3K/AKT/mTORC1 signalling. Neuropathol. Appl. Neurobiol. 2018, 44, 687–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noorolyai, S.; Mokhtarzadeh, A.; Baghbani, E.; Asadi, M.; Baghbanzadeh Kojabad, A.; Mogaddam, M.M.; Baradaran, B. The role of microRNAs involved in PI3-kinase signaling pathway in colorectal cancer. J. Cell Physiol. 2019, 234, 5664–5673. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Guo, H.; Yin, S.; Du, H. miR-139-5p functions as a tumor suppressor in cervical cancer by targeting TCF4 and inhibiting Wnt/β-catenin signaling. Onco Targets Ther. 2019, 12, 7739–7748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirjang, S.; Mansoori, B.; Asghari, S.; Duijf, P.H.G.; Mohammadi, A.; Gjerstorff, M.; Baradaran, B. MicroRNAs in cancer cell death pathways: Apoptosis and necroptosis. Free Radic. Biol. Med. 2019, 139, 1–15. [Google Scholar] [CrossRef]
- Niveditha, D.; Jasoria, M.; Narayan, J.; Majumder, S.; Mukherjee, S.; Chowdhury, R.; Chowdhury, S. Common and unique microRNAs in multiple carcinomas regulate similar network of pathways to mediate cancer progression. Sci. Rep. 2020, 10, 2331. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Chen, Z.; Yu, G. microRNA-139-3p inhibits malignant behaviors of laryngeal cancer cells via the KDM5B/SOX2 axis and the Wnt/β-catenin pathway. Cancer Manag. Res. 2020, 12, 9197–9209. [Google Scholar] [CrossRef]
- Xu, Y.J.; Yu, H.; Liu, G.X. Hsa_circ_0031288/hsa-miR-139-3p/Bcl-6 regulatory feedback circuit influences the invasion and migration of cervical cancer HeLa cells. J. Cell Biochem. 2020, 121, 4251–4260. [Google Scholar] [CrossRef]
- Yonemori, M.; Seki, N.; Yoshino, H.; Matsushita, R.; Miyamoto, K.; Nakagawa, M.; Enokida, H. Dual tumor-suppressors miR-139-5p and miR-139-3p targeting matrix metalloprotease 11 in bladder cancer. Cancer Sci. 2016, 107, 1233–1242. [Google Scholar] [CrossRef] [Green Version]
- Hipskind, R.A.; Rao, V.N.; Mueller, C.G.; Reddy, E.S.; Nordheim, A. Ets-related protein Elk-1 is homologous to the c-fos regulatory factor p62TCF. Nature 1991, 354, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Gille, H.; Kortenjann, M.; Thomae, O.; Moomaw, C.; Slaughter, C.; Cobb, M.H.; Shaw, P.E. ERK phosphorylation potentiates Elk-1-mediated ternary complex formation and transactivation. EMBO J. 1995, 14, 951–962. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ye, Z.; Chen, S.; Pan, Z.; Zhou, Q.; Li, Y.Z.; Shuai, W.D.; Kuang, C.M.; Peng, Q.H.; Shi, W.; et al. ARHGEF19 interacts with BRAF to activate MAPK signaling during the tumorigenesis of non-small cell lung cancer. Int. J. Cancer 2018, 142, 1379–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiDonato, J.A.; Hayakawa, M.; Rothwarf, D.M.; Zandi, E.; Karin, M. A cytokine-responsive IkappaB kinase that activates the transcription factor NF-kappaB. Nature 1997, 388, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB system. Wiley. Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef] [Green Version]
- Oya, M.; Takayanagi, A.; Horiguchi, A.; Mizuno, R.; Ohtsubo, M.; Marumo, K.; Shimizu, N.; Murai, M. Increased nuclear factor-kappa B activation is related to the tumor development of renal cell carcinoma. Carcinogenesis 2003, 24, 377–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.J.; Zhong, J.; Yu, J.G.; Zhao, F.; Xiang, Y. The structure and functions of paxillin and its roles in neovascularization. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1768–1773. [Google Scholar]
- López-Colomé, A.M.; Lee-Rivera, I.; Benavides-Hidalgo, R.; López, E. Paxillin: A crossroad in pathological cell migration. J. Hematol. Oncol. 2017, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Deakin, N.O.; Pignatelli, J.; Turner, C.E. Diverse roles for the paxillin family of proteins in cancer. Genes Cancer 2012, 3, 362–370. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, A.; Nam, A.; Pozhitkov, A.; Yang, L.; Srivastava, S.; Nathan, A.; Wu, X.; Mambetsariev, I.; Nelson, M.; Subbalakshmi, A.R.; et al. A non-genetic mechanism involving the integrin β4/paxillin axis contributes to chemoresistance in lung cancer. iScience 2020, 23, 101496. [Google Scholar] [CrossRef]
- Wu, D.W.; Wu, T.C.; Wu, J.Y.; Cheng, Y.W.; Chen, Y.C.; Lee, M.C.; Chen, C.Y.; Lee, H. Phosphorylation of paxillin confers cisplatin resistance in non-small cell lung cancer via activating ERK-mediated Bcl-2 expression. Oncogene 2014, 33, 4385–4395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, L.; Zhang, X.; Zhang, L.; Chen, S.; Ma, Y.; Hu, J.; Yue, T.; Wang, J.; Zhu, J.; Wu, T.; et al. Paxillin knockdown suppresses metastasis and epithelial-mesenchymal transition in colorectal cancer via the ERK signalling pathway. Oncol. Rep. 2020, 44, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, Z.; Xiong, J.; Gong, B.; Zhang, G.; Cao, C.; Jie, Z.; Liu, Y.; Cao, Y.; Yan, Y.; et al. MicroRNA-212 functions as an epigenetic-silenced tumor suppressor involving in tumor metastasis and invasion of gastric cancer through down-regulating PXN expression. Am. J. Cancer Res. 2015, 5, 2980–2997. [Google Scholar] [PubMed]
- Qin, J.; Wang, F.; Jiang, H.; Xu, J.; Jiang, Y.; Wang, Z. MicroRNA-145 suppresses cell migration and invasion by targeting paxillin in human colorectal cancer cells. Int. J. Clin. Exp. Pathol. 2015, 8, 1328–1340. [Google Scholar]
- Qin, J.; Ke, J.; Xu, J.; Wang, F.; Zhou, Y.; Jiang, Y.; Wang, Z. Downregulation of microRNA-132 by DNA hypermethylation is associated with cell invasion in colorectal cancer. Onco. Targets Ther. 2015, 8, 3639–3648. [Google Scholar] [CrossRef] [Green Version]
- Bi, Y.; Han, Y.; Bi, H.; Gao, F.; Wang, X. miR-137 impairs the proliferative and migratory capacity of human non-small cell lung cancer cells by targeting paxillin. Hum. Cell 2014, 27, 95–102. [Google Scholar] [CrossRef]
- Wu, D.W.; Cheng, Y.W.; Wang, J.; Chen, C.Y.; Lee, H. Paxillin predicts survival and relapse in non-small cell lung cancer by microRNA-218 targeting. Cancer Res. 2010, 70, 10392–10401. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, H.; Enokida, H.; Itesako, T.; Kojima, S.; Kinoshita, T.; Tatarano, S.; Chiyomaru, T.; Nakagawa, M.; Seki, N. Tumor-suppressive microRNA-143/145 cluster targets hexokinase-2 in renal cell carcinoma. Cancer Sci. 2013, 104, 1567–1574. [Google Scholar] [CrossRef]
- Yamasaki, T.; Seki, N.; Yoshino, H.; Itesako, T.; Hidaka, H.; Yamada, Y.; Tatarano, S.; Yonezawa, T.; Kinoshita, T.; Nakagawa, M.; et al. MicroRNA-218 inhibits cell migration and invasion in renal cell carcinoma through targeting caveolin-2 involved in focal adhesion pathway. J. Urol. 2013, 190, 1059–1068. [Google Scholar] [CrossRef]
- Du, C.; Wang, Y.; Zhang, Y.; Zhang, J.; Zhang, L.; Li, J. LncRNA DLX6-AS1 contributes to epithelial-mesenchymal transition and cisplatin resistance in triple-negative breast cancer via modulating Mir-199b-5p/paxillin axis. Cell Transpl. 2020, 29. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. Candidate target genes of miR-139-5p | |||||||
| Gene Symbol | Gene Name | Entrez Gene ID | miR-139-5p- Transfected A498 Cells (log2 FC) | miR-139-5p- Transfected 786-O Cells (log2 FC) | GSE36895 (log2 FC) | Total Binding Sites | TCGA 5y OS p-Value |
| PLXDC1 | plexin domain containing 1 | 57125 | −0.615791 | −1.455703 | 1.6528159 | 1 | 0.0098 |
| TET3 | tet methylcytosine dioxygenase 3 | 200424 | −1.074818 | −1.152835 | 0.7350367 | 1 | 0.0261 |
| IRF4 | interferon regulatory factor 4 | 3662 | −2.491284 | −1.563216 | 0.72956836 | 1 | 0.0575 |
| RAB27B | RAB27B, member RAS oncogene family | 5874 | −0.620716 | −0.970745 | 0.7823266 | 1 | 0.1244 |
| FCHSD2 | FCH and double SH3 domains 2 | 9873 | −1.834753 | −1.364281 | 0.5673638 | 1 | 0.1565 |
| DMD | dystrophin | 1756 | −0.98734 | −1.12498 | 0.46094477 | 1 | 0.3891 |
| APOL6 | apolipoprotein L, 6 | 80830 | −1.363298 | −0.574163 | 0.43652818 | 2 | 0.4733 |
| AP1S2 | adaptor-related protein complex 1, sigma 2 subunit | 8905 | −0.571341 | −0.63469 | 0.57979524 | 1 | 0.5072 |
| PTPRU | protein tyrosine phosphatase, receptor type, U | 10076 | −1.430874 | −0.883479 | 0.8481535 | 1 | 0.6152 |
| TRAT1 | T cell receptor associated transmembrane adaptor 1 | 50852 | −1.192173 | −1.781637 | 1.9733018 | 2 | 0.7395 |
| SLC39A14 | solute carrier family 39 (zinc transporter), member 14 | 23516 | −0.536152 | −0.705082 | 1.0408258 | 1 | 0.8125 |
| OTUD4 | OTU deubiquitinase 4 | 54726 | −1.720465 | −1.59549 | 0.21931966 | 1 | 0.9338 |
| CDCA7L | cell division cycle associated 7-like | 55536 | −1.478013 | −0.564377 | 2.1733913 | 1 | 0.969 |
| ZNF678 | zinc finger protein 678 | 339500 | −1.585917 | −0.621832 | 0.25128952 | 2 | 0.0086 * |
| FGFBP2 | fibroblast growth factor binding protein 2 | 83888 | −0.830757 | −1.317887 | 1.5742466 | 1 | 0.0058 * |
| ATP2B2 | ATPase, Ca++ transporting, plasma membrane 2 | 491 | −1.109889 | −2.668194 | 1.3068246 | 3 | 0.0050 * |
| EML1 | echinoderm microtubule associated protein like 1 | 2009 | −0.56874 | −0.541978 | 0.38891175 | 2 | 0.0014 * |
| PCSK5 | proprotein convertase subtilisin/kexin type 5 | 5125 | −1.751102 | −0.577052 | 0.57546955 | 1 | 0.0006 * |
| FAM168A | family with sequence similarity 168, member A | 23201 | −1.390649 | −0.959992 | 0.25754136 | 1 | 0.0002 * |
| B. Candidate target genes of miR-139-3p | |||||||
| Gene Symbol | Gene Name | Entrez Gene ID | miR-139-3p- Transfected A498 Cells (log2 FC) | miR-139-3p- Transfected 786-O Cells (log2 FC) | GSE36895 (log2 FC) | Total Binding Sites | TCGA 5y OS p-Value |
| PXN | paxillin | 5829 | −1.164181 | −0.707167 | 0.481819 | 2 | <0.0001 |
| ARHGEF19 | Rho guanine nucleotide exchange factor (GEF) 19 | 128272 | −0.607603 | −1.826996 | 1.1049173 | 1 | <0.0001 |
| ELK1 | ELK1, member of ETS oncogene family | 2002 | −1.518329 | −0.839984 | 0.5987212 | 2 | 0.0001 |
| CSF1 | colony stimulating factor 1 (macrophage) | 1435 | −0.952795 | −0.537074 | 1.0153022 | 1 | 0.0124 |
| IKBKB | inhibitor of kappa light polypeptide gene enhancer in B-cells, kinase beta | 3551 | −0.511766 | −1.882033 | 0.23441868 | 1 | 0.0251 |
| DCBLD1 | discoidin, CUB and LCCL domain containing 1 | 285761 | −1.398862 | −0.668245 | 0.28628728 | 1 | 0.0285 |
| SYT11 | synaptotagmin XI | 23208 | −1.034462 | −0.578617 | 0.4411527 | 1 | 0.0556 |
| SERPINE1 | serpin peptidase inhibitor, clade E (nexin, plasminogen activator inhibitor type 1), member 1 | 5054 | −2.516404 | −0.663638 | 1.8049024 | 3 | 0.0731 |
| KDM6B | lysine (K)-specific demethylase 6B | 23135 | −0.62567 | −0.733171 | 0.42303625 | 1 | 0.1019 |
| RASSF5 | Ras association (RalGDS/AF-6) domain family member 5 | 83593 | −1.180982 | −4.605049 | 1.5059676 | 1 | 0.3643 |
| ACBD3 | acyl-CoA binding domain containing 3 | 64746 | −0.688812 | −0.542166 | 0.65537864 | 1 | 0.3841 |
| APOL6 | apolipoprotein L, 6 | 80830 | −1.350307 | −0.858142 | 0.43652818 | 1 | 0.4733 |
| EPN2 | epsin 2 | 22905 | −0.619177 | −0.520903 | 0.29098234 | 1 | 0.581 |
| GIT2 | G protein-coupled receptor kinase interacting ArfGAP 2 | 9815 | −0.64332 | −0.544951 | 1.4113243 | 1 | 0.7179 |
| KIF3C | kinesin family member 3C | 3797 | −0.694929 | −0.956568 | 0.29198763 | 1 | 0.9148 |
| ARAP2 | ArfGAP with RhoGAP domain, ankyrin repeat and PH domain 2 | 116984 | −1.161352 | −1.258001 | 0.3943753 | 1 | 0.0940 * |
| RFX2 | regulatory factor X, 2 (influences HLA class II expression) | 5990 | −0.937881 | −0.580092 | 1.4766915 | 1 | 0.0671 * |
| RNF125 | ring finger protein 125, E3 ubiquitin protein ligase | 54941 | −0.539067 | −1.811576 | 0.6103558 | 1 | 0.0394 * |
| ARSK | arylsulfatase family, member K | 153642 | −1.505581 | −0.887476 | 0.4327301 | 1 | 0.0219 * |
| STAG2 | stromal antigen 2 | 10735 | −0.527565 | −0.563985 | 0.38703138 | 1 | <0.0001 * |
| TNS1 | tensin 1 | 7145 | −0.764289 | −0.609893 | 0.28155625 | 1 | <0.0001 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okada, R.; Goto, Y.; Yamada, Y.; Kato, M.; Asai, S.; Moriya, S.; Ichikawa, T.; Seki, N. Regulation of Oncogenic Targets by the Tumor-Suppressive miR-139 Duplex (miR-139-5p and miR-139-3p) in Renal Cell Carcinoma. Biomedicines 2020, 8, 599. https://doi.org/10.3390/biomedicines8120599
Okada R, Goto Y, Yamada Y, Kato M, Asai S, Moriya S, Ichikawa T, Seki N. Regulation of Oncogenic Targets by the Tumor-Suppressive miR-139 Duplex (miR-139-5p and miR-139-3p) in Renal Cell Carcinoma. Biomedicines. 2020; 8(12):599. https://doi.org/10.3390/biomedicines8120599
Chicago/Turabian StyleOkada, Reona, Yusuke Goto, Yasutaka Yamada, Mayuko Kato, Shunichi Asai, Shogo Moriya, Tomohiko Ichikawa, and Naohiko Seki. 2020. "Regulation of Oncogenic Targets by the Tumor-Suppressive miR-139 Duplex (miR-139-5p and miR-139-3p) in Renal Cell Carcinoma" Biomedicines 8, no. 12: 599. https://doi.org/10.3390/biomedicines8120599