CD146/sCD146 in the Pathogenesis and Monitoring of Angiogenic and Inflammatory Diseases

 , , ,
, , ,
Abstract
:1. Introduction
2. Systemic Sclerosis
3. Diabetes Mellitus
4. Rheumatoid Arthritis
5. Inflammatory Bowel Disease
6. Multiple Sclerosis
7. Interest of sCD146 and CD146 in Autoimmune Diseases
8. Interest in CD146 and sCD146 as Molecular Targets
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kebir, A.; Harhouri, K.; Guillet, B.; Liu, J.W.; Foucault-Bertaud, A.; Lamy, E.; Kaspi, E.; Elganfoud, N.; Vely, F.; Sabatier, F.; et al. CD146 short isoform increases the proangiogenic potential of endothelial progenitor cells in vitro and in vivo. Circ. Res. 2010, 107, 66–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardin, N.; Anfosso, F.; Massé, J.M.; Cramer, E.; Sabatier, F.; Le Bivic, A.; Sampol, J.; Dignat-George, F. Identification of CD146 as a component of the endothelial junction involved in the control of cell-cell cohesion. Blood 2001, 98, 3677–3684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dagur, P.K.; Biancotto, A.; Wei, L.; Sen, H.N.; Yao, M.; Strober, W.; Nussenblatt, R.B.; McCoy, J.P. MCAM-expressing CD4(+) T cells in peripheral blood secrete IL-17A and are significantly elevated in inflammatory autoimmune diseases. J. Autoimmun. 2011, 37, 319–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Kim, K.-R.; McKeon, F.; Yang, A.; Boyd, T.K.; Crum, C.P.; Parast, M.M. A unifying concept of trophoblastic differentiation and malignancy defined by biomarker expression. Hum. Pathol. 2007, 38, 1003–1013. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, A.; Ferracin, M.; Castelli, G.; Biffoni, M.; Tomaselli, G.; Baiocchi, M.; Fatica, A.; Negrini, M.; Peschle, C.; Valtieri, M. Isolation and characterization of CD146+ multipotent mesenchymal stromal cells. Exp. Hematol. 2008, 36, 1035–1046. [Google Scholar] [CrossRef]
- Lehmann, J.M.; Riethmüller, G.; Johnson, J.P. MUC18, a marker of tumor progression in human melanoma, shows sequence similarity to the neural cell adhesion molecules of the immunoglobulin superfamily. Proc. Natl. Acad. Sci. USA 1989, 86, 9891–9895. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Ohuchida, K.; Chijiiwa, Y.; Zhao, M.; Mizuuchi, Y.; Cui, L.; Horioka, K.; Ohtsuka, T.; Mizumoto, K.; Oda, Y.; et al. CD146 attenuation in cancer-associated fibroblasts promotes pancreatic cancer progression. Mol. Carcinog. 2016, 55, 1560–1572. [Google Scholar] [CrossRef]
- Bardin, N.; Blot-Chabaud, M.; Despoix, N.; Kebir, A.; Harhouri, K.; Arsanto, J.-P.; Espinosa, L.; Perrin, P.; Robert, S.; Vely, F.; et al. CD146 and its soluble form regulate monocyte transendothelial migration. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 746–753. [Google Scholar] [CrossRef] [Green Version]
- Bardin, N.; Francès, V.; Combes, V.; Sampol, J.; Dignat-George, F. CD146: Biosynthesis and production of a soluble form in human cultured endothelial cells. FEBS Lett. 1998, 421, 12–14. [Google Scholar] [CrossRef] [Green Version]
- Daniel, L.; Bardin, N.; Moal, V.; Dignat-George, F.; Berland, Y.; Figarella-Branger, D. Tubular CD146 expression in nephropathies is related to chronic renal failure. Nephron Exp. Nephrol. 2005, 99, e105–e111. [Google Scholar] [CrossRef]
- Duan, H.; Luo, Y.; Hao, H.; Feng, L.; Zhang, Y.; Lu, D.; Xing, S.; Feng, J.; Yang, D.; Song, L.; et al. Soluble CD146 in cerebrospinal fluid of active multiple sclerosis. Neuroscience 2013, 235, 16–26. [Google Scholar] [CrossRef] [PubMed]
- So, J.-H.; Hong, S.-K.; Kim, H.-T.; Jung, S.-H.; Lee, M.-S.; Choi, J.-H.; Bae, Y.-K.; Kudoh, T.; Kim, J.-H.; Kim, C.-H. Gicerin/Cd146 is involved in zebrafish cardiovascular development and tumor angiogenesis. Genes Cells Devoted Mol. Cell. Mech. 2010, 15, 1099–1110. [Google Scholar] [CrossRef] [PubMed]
- Harhouri, K.; Kebir, A.; Guillet, B.; Foucault-Bertaud, A.; Voytenko, S.; Piercecchi-Marti, M.-D.; Berenguer, C.; Lamy, E.; Vely, F.; Pisano, P.; et al. Soluble CD146 displays angiogenic properties and promotes neovascularization in experimental hind-limb ischemia. Blood 2010, 115, 3843–3851. [Google Scholar] [CrossRef]
- Stalin, J.; Harhouri, K.; Hubert, L.; Subrini, C.; Lafitte, D.; Lissitzky, J.-C.; Elganfoud, N.; Robert, S.; Foucault-Bertaud, A.; Kaspi, E.; et al. Soluble melanoma cell adhesion molecule (sMCAM/sCD146) promotes angiogenic effects on endothelial progenitor cells through angiomotin. J. Biol. Chem. 2013, 288, 8991–9000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, T.; Wondimu, Z.; Oikawa, Y.; Gentilcore, G.; Kiessling, R.; Egyhazi Brage, S.; Hansson, J.; Patarroyo, M. Laminins 411 and 421 differentially promote tumor cell migration via α6β1 integrin and MCAM (CD146). Matrix Biol. J. Int. Soc. Matrix Biol. 2014, 38, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Guezguez, B.; Vigneron, P.; Lamerant, N.; Kieda, C.; Jaffredo, T.; Dunon, D. Dual role of melanoma cell adhesion molecule (MCAM)/CD146 in lymphocyte endothelium interaction: MCAM/CD146 promotes rolling via microvilli induction in lymphocyte and is an endothelial adhesion receptor. J. Immunol. 2007, 179, 6673–6685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Duan, H.; Qian, Y.; Feng, L.; Wu, Z.; Wang, F.; Feng, J.; Yang, D.; Qin, Z.; Yan, X. Macrophagic CD146 promotes foam cell formation and retention during atherosclerosis. Cell Res. 2017, 27, 352–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Xu, Q.; Zhang, N.; Du, X.; Xu, G.; Yan, X. CD146, from a melanoma cell adhesion molecule to a signaling receptor. Signal Transduct. Target. Ther. 2020, 5, 148. [Google Scholar] [CrossRef]
- Ye, Z.; Zhang, C.; Tu, T.; Sun, M.; Liu, D.; Lu, D.; Feng, J.; Yang, D.; Liu, F.; Yan, X. Wnt5a uses CD146 as a receptor to regulate cell motility and convergent extension. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Witze, E.S.; Litman, E.S.; Argast, G.M.; Moon, R.T.; Ahn, N.G. Wnt5a control of cell polarity and directional movement by polarized redistribution of adhesion receptors. Science 2008, 320, 365–369. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Zhuang, J.; Duan, H.; Luo, Y.; Zeng, Q.; Fan, K.; Yan, H.; Lu, D.; Ye, Z.; Hao, J.; et al. CD146 is a coreceptor for VEGFR-2 in tumor angiogenesis. Blood 2012, 120, 2330–2339. [Google Scholar] [CrossRef] [Green Version]
- Distler, O.; Del Rosso, A.; Giacomelli, R.; Cipriani, P.; Conforti, M.L.; Guiducci, S.; Gay, R.E.; Michel, B.A.; Brühlmann, P.; Müller-Ladner, U.; et al. Angiogenic and angiostatic factors in systemic sclerosis: Increased levels of vascular endothelial growth factor are a feature of the earliest disease stages and are associated with the absence of fingertip ulcers. Arthritis Res. 2002, 4, R11. [Google Scholar] [CrossRef] [Green Version]
- Abraham, D.; Dashwood, M. Endothelin—Role in vascular disease. Rheumatology 2008, 47, v23–v24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutolo, M.; Soldano, S.; Smith, V. Pathophysiology of systemic sclerosis: Current understanding and new insights. Expert Rev. Clin. Immunol. 2019, 15, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Derk, C.T.; Jimenez, S.A. Systemic sclerosis: Current views of its pathogenesis. Autoimmun. Rev. 2003, 2, 181–191. [Google Scholar] [CrossRef]
- Mouthon, L.; García De La Peña-Lefebvre, P.; Chanseaud, Y.; Tamby, M.C.; Boissier, M.-C.; Guillevin, L. Pathogenesis of systemic scleroderma: Immunological aspects. Ann. Med. Interne 2002, 153, 167–178. [Google Scholar]
- Lazzaroni, M.-G.; Cavazzana, I.; Colombo, E.; Dobrota, R.; Hernandez, J.; Hesselstrand, R.; Varju, C.; Nagy, G.; Smith, V.; Caramaschi, P.; et al. Malignancies in Patients with Anti-RNA Polymerase III Antibodies and Systemic Sclerosis: Analysis of the EULAR Scleroderma Trials and Research Cohort and Possible Recommendations for Screening. J. Rheumatol. 2017, 44, 639–647. [Google Scholar] [CrossRef] [Green Version]
- Meyer, O.; De Chaisemartin, L.; Nicaise-Roland, P.; Cabane, J.; Tubach, F.; Dieude, P.; Hayem, G.; Palazzo, E.; Chollet-Martin, S.; Kahan, A.; et al. Anti-RNA Polymerase III Antibody Prevalence and Associated Clinical Manifestations in a Large Series of French Patients with Systemic Sclerosis: A Cross-sectional Study. J. Rheumatol. 2010, 37, 125–130. [Google Scholar] [CrossRef]
- Kowal-Bielecka, O.; Fransen, J.; Avouac, J.; Becker, M.; Kulak, A.; Allanore, Y.; Distler, O.; Clements, P.; Cutolo, M.; Czirjak, L.; et al. Update of EULAR recommendations for the treatment of systemic sclerosis. Ann. Rheum. Dis. 2017, 76, 1327–1339. [Google Scholar] [CrossRef] [Green Version]
- Kaspi, E.; Heim, X.; Granel, B.; Guillet, B.; Stalin, J.; Nollet, M.; Bertaud-Foucault, A.; Robaglia-Schlupp, A.; Roll, P.; Cau, P.; et al. Identification of CD146 as a novel molecular actor involved in systemic sclerosis. J. Allergy Clin. Immunol. 2017, 140, 1448–1451.e6. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Tamura, N.; Okuda, S.; Tada, K.; Matsushita, M.; Yamaji, K.; Kato, K.; Takasaki, Y. Elevated serum levels of soluble CD146 in patients with systemic sclerosis. Clin. Rheumatol. 2017, 36, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Gabsi, A.; Heim, X.; Dlala, A.; Gati, A.; Sakhri, H.; Abidi, A.; Amri, S.; Neili, B.; Leroyer, A.S.; Bertaud, A.; et al. TH17 cells expressing CD146 are significantly increased in patients with Systemic sclerosis. Sci. Rep. 2019, 9, 17721. [Google Scholar] [CrossRef] [Green Version]
- Pearson, T.; Wattis, J.A.D.; King, J.R.; MacDonald, I.A.; Mazzatti, D.J. The Effects of Insulin Resistance on Individual Tissues: An Application of a Mathematical Model of Metabolism in Humans. Bull. Math. Biol. 2016, 78, 1189–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.D.; Harris-Hayes, M.; Schootman, M. Epidemiology of diabetes and diabetes-related complications. Phys. Ther. 2008, 88, 1254–1264. [Google Scholar] [CrossRef] [Green Version]
- Wada, J.; Makino, H. Inflammation and the pathogenesis of diabetic nephropathy. Clin. Sci. 2013, 124, 139–152. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Fei, Y.; Zheng, L.; Wang, J.; Xiao, W.; Wen, J.; Xu, Y.; Wang, Y.; He, L.; Guan, J.; et al. Expression of Endothelial Cell Injury Marker Cd146 Correlates with Disease Severity and Predicts the Renal Outcomes in Patients with Diabetic Nephropathy. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 48, 63–74. [Google Scholar] [CrossRef]
- Wang, N.; Fan, Y.; Ni, P.; Wang, F.; Gao, X.; Xue, Q.; Tang, L. High glucose effect on the role of CD146 in human proximal tubular epithelial cells in vitro. J. Nephrol. 2008, 21, 931–940. [Google Scholar]
- Saito, T.; Saito, O.; Kawano, T.; Tamemoto, H.; Kusano, E.; Kawakami, M.; Ishikawa, S.-E. Elevation of serum adiponectin and CD146 levels in diabetic nephropathy. Diabetes Res. Clin. Pract. 2007, 78, 85–92. [Google Scholar] [CrossRef]
- Hohenstein, B.; Hausknecht, B.; Boehmer, K.; Riess, R.; Brekken, R.A.; Hugo, C.P.M. Local VEGF activity but not VEGF expression is tightly regulated during diabetic nephropathy in man. Kidney Int. 2006, 69, 1654–1661. [Google Scholar] [CrossRef] [Green Version]
- Ciftci Dogansen, S.; Helvaci, A.; Adas, M.; Deniz Onal, S. The relationship between early atherosclerosis and endothelial dysfunction in type 1 diabetic patients as evidenced by measurement of carotid intima-media thickness and soluble CD146 levels: A cross sectional study. Cardiovasc. Diabetol. 2013, 12, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aletaha, D.; Funovits, J.; Smolen, J.S. Physical disability in rheumatoid arthritis is associated with cartilage damage rather than bone destruction. Ann. Rheum. Dis. 2011, 70, 733–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallach, D. The cybernetics of TNF: Old views and newer ones. Semin. Cell Dev. Biol. 2016, 50, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O.; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheum. 2010, 62, 2569–2581. [Google Scholar] [CrossRef] [PubMed]
- Moll, J.; Isailovic, N.; De Santis, M.; Selmi, C. Rheumatoid Factors in Hepatitis B and C Infections: Connecting Viruses, Autoimmunity, and Cancer. Isr. Med. Assoc. J. IMAJ 2019, 21, 480–486. [Google Scholar] [PubMed]
- Kroot, E.J.; de Jong, B.A.; van Leeuwen, M.A.; Swinkels, H.; van den Hoogen, F.H.; van’t Hof, M.; van de Putte, L.B.; van Rijswijk, M.H.; van Venrooij, W.J.; van Riel, P.L. The prognostic value of anti-cyclic citrullinated peptide antibody in patients with recent-onset rheumatoid arthritis. Arthritis Rheum. 2000, 43, 1831–1835. [Google Scholar] [CrossRef]
- Neidhart, M.; Wehrli, R.; Brühlmann, P.; Michel, B.A.; Gay, R.E.; Gay, S. Synovial fluid CD146 (MUC18), a marker for synovial membrane angiogenesis in rheumatoid arthritis. Arthritis Rheum. 1999, 42, 622–630. [Google Scholar] [CrossRef]
- Wu, C.; Goodall, J.C.; Busch, R.; Gaston, J.S.H. Relationship of CD146 expression to secretion of interleukin (IL)-17, IL-22 and interferon-γ by CD4(+) T cells in patients with inflammatory arthritis. Clin. Exp. Immunol. 2015, 179, 378–391. [Google Scholar] [CrossRef] [Green Version]
- Pickl, W.F.; Majdic, O.; Fischer, G.F.; Petzelbauer, P.; Faé, I.; Waclavicek, M.; Stöckl, J.; Scheinecker, C.; Vidicki, T.; Aschauer, H.; et al. MUC18/MCAM (CD146), an activation antigen of human T lymphocytes. J. Immunol. 1997, 158, 2107–2115. [Google Scholar]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Martinez, C.; Antolin, M.; Santos, J.; Torrejon, A.; Casellas, F.; Borruel, N.; Guarner, F.; Malagelada, J.-R. Unstable composition of the fecal microbiota in ulcerative colitis during clinical remission. Am. J. Gastroenterol. 2008, 103, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Mahida, Y.R. The key role of macrophages in the immunopathogenesis of inflammatory bowel disease. Inflamm. Bowel Dis. 2000, 6, 21–33. [Google Scholar] [CrossRef]
- Danese, S.; Sans, M.; de la Motte, C.; Graziani, C.; West, G.; Phillips, M.H.; Pola, R.; Rutella, S.; Willis, J.; Gasbarrini, A.; et al. Angiogenesis as a novel component of inflammatory bowel disease pathogenesis. Gastroenterology 2006, 130, 2060–2073. [Google Scholar] [CrossRef] [Green Version]
- Sands, B.E.; Sandborn, W.J.; Panaccione, R.; O’Brien, C.D.; Zhang, H.; Johanns, J.; Adedokun, O.J.; Li, K.; Peyrin-Biroulet, L.; Van Assche, G.; et al. Ustekinumab as Induction and Maintenance Therapy for Ulcerative Colitis. N. Engl. J. Med. 2019, 381, 1201–1214. [Google Scholar] [CrossRef] [PubMed]
- Danese, S.; Vuitton, L.; Peyrin-Biroulet, L. Biologic agents for IBD: Practical insights. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Bardin, N.; Reumaux, D.; Geboes, K.; Colombel, J.F.; Blot-Chabaud, M.; Sampol, J.; Duthilleul, P.; Dignat-George, F. Increased expression of CD146, a new marker of the endothelial junction in active inflammatory bowel disease. Inflamm. Bowel Dis. 2006, 12, 16–21. [Google Scholar] [CrossRef]
- Tsiolakidou, G.; Koutroubakis, I.E.; Tzardi, M.; Kouroumalis, E.A. Increased expression of VEGF and CD146 in patients with inflammatory bowel disease. Dig. Liver Dis. Off. J. Ital. Soc. Gastroenterol. Ital. Assoc. Study Liver 2008, 40, 673–679. [Google Scholar] [CrossRef]
- Reumaux, D.; Bardin, N.; Colombel, J.-F.; Dignat-George, F.; Duthilleul, P.; Vermeire, S. Restoration of soluble CD146 in patients with Crohn’s disease treated with the TNF-alpha antagonist infliximab. Inflamm. Bowel Dis. 2007, 13, 1315–1317. [Google Scholar] [CrossRef]
- Xing, S.; Luo, Y.; Liu, Z.; Bu, P.; Duan, H.; Liu, D.; Wang, P.; Yang, J.; Song, L.; Feng, J.; et al. Targeting endothelial CD146 attenuates colitis and prevents colitis-associated carcinogenesis. Am. J. Pathol. 2014, 184, 1604–1616. [Google Scholar] [CrossRef]
- Ghosh, S.; Panaccione, R. Anti-adhesion molecule therapy for inflammatory bowel disease. Ther. Adv. Gastroenterol. 2010, 3, 239–258. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.F.; Alvarez, E. The immunopathophysiology of multiple sclerosis. Neurol. Clin. 2011, 29, 257–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: 2010 Revisions to the McDonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Polman, C.H.; O’Connor, P.W.; Havrdova, E.; Hutchinson, M.; Kappos, L.; Miller, D.H.; Phillips, J.T.; Lublin, F.D.; Giovannoni, G.; Wajgt, A.; et al. A randomized, placebo-controlled trial of natalizumab for relapsing multiple sclerosis. N. Engl. J. Med. 2006, 354, 899–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, L.; Larochelle, C.; Prat, A. Update on treatments in multiple sclerosis. Presse Med. 2015, 44, e137–e151. [Google Scholar] [CrossRef] [PubMed]
- Brucklacher-Waldert, V.; Stuerner, K.; Kolster, M.; Wolthausen, J.; Tolosa, E. Phenotypical and functional characterization of T helper 17 cells in multiple sclerosis. Brain J. Neurol. 2009, 132, 3329–3341. [Google Scholar] [CrossRef] [PubMed]
- Larochelle, C.; Lécuyer, M.-A.; Alvarez, J.I.; Charabati, M.; Saint-Laurent, O.; Ghannam, S.; Kebir, H.; Flanagan, K.; Yednock, T.; Duquette, P.; et al. Melanoma cell adhesion molecule-positive CD8 T lymphocytes mediate central nervous system inflammation. Ann. Neurol. 2015, 78, 39–53. [Google Scholar] [CrossRef]
- Schneider-Hohendorf, T.; Rossaint, J.; Mohan, H.; Böning, D.; Breuer, J.; Kuhlmann, T.; Gross, C.C.; Flanagan, K.; Sorokin, L.; Vestweber, D.; et al. VLA-4 blockade promotes differential routes into human CNS involving PSGL-1 rolling of T cells and MCAM-adhesion of TH17 cells. J. Exp. Med. 2014, 211, 1833–1846. [Google Scholar] [CrossRef]
- Petersen, E.R.; Ammitzbøll, C.; Søndergaard, H.B.; Oturai, A.B.; Sørensen, P.S.; Nilsson, A.C.; Börnsen, L.; von Essen, M.; Sellebjerg, F. Expression of melanoma cell adhesion molecule-1 (MCAM-1) in natalizumab-treated multiple sclerosis. J. Neuroimmunol. 2019, 337, 577085. [Google Scholar] [CrossRef]
- Duan, H.; Xing, S.; Luo, Y.; Feng, L.; Gramaglia, I.; Zhang, Y.; Lu, D.; Zeng, Q.; Fan, K.; Feng, J.; et al. Targeting endothelial CD146 attenuates neuroinflammation by limiting lymphocyte extravasation to the CNS. Sci. Rep. 2013, 3, 1687. [Google Scholar] [CrossRef] [Green Version]
- Breuer, J.; Korpos, E.; Hannocks, M.-J.; Schneider-Hohendorf, T.; Song, J.; Zondler, L.; Herich, S.; Flanagan, K.; Korn, T.; Zarbock, A.; et al. Blockade of MCAM/CD146 impedes CNS infiltration of T cells over the choroid plexus. J. Neuroinflammation 2018, 15, 236. [Google Scholar] [CrossRef]
- Bardin, N.; Moal, V.; Anfosso, F.; Daniel, L.; Brunet, P.; Sampol, J.; Dignat George, F. Soluble CD146, a novel endothelial marker, is increased in physiopathological settings linked to endothelial junctional alteration. Thromb. Haemost. 2003, 90, 915–920. [Google Scholar] [CrossRef] [PubMed]
- Figarella-Branger, D.; Schleinitz, N.; Boutière-Albanèse, B.; Camoin, L.; Bardin, N.; Guis, S.; Pouget, J.; Cognet, C.; Pellissier, J.-F.; Dignat-George, F. Platelet-endothelial cell adhesion molecule-1 and CD146: Soluble levels and in situ expression of cellular adhesion molecules implicated in the cohesion of endothelial cells in idiopathic inflammatory myopathies. J. Rheumatol. 2006, 33, 1623–1630. [Google Scholar] [PubMed]
- Seftalioğlu, A.; Karakoç, L. Expression of CD146 adhesion molecules (MUC18 or MCAM) in the thymic microenvironment. Acta Histochem. 2000, 102, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Kamiyama, T.; Watanabe, H.; Iijima, M.; Miyazaki, A.; Iwamoto, S. Coexpression of CCR6 and CD146 (MCAM) is a marker of effector memory T-helper 17 cells. J. Dermatol. 2012, 39, 838–842. [Google Scholar] [CrossRef]
- Dagur, P.K.; McCoy, J.P. Endothelial-binding, proinflammatory T cells identified by MCAM (CD146) expression: Characterization and role in human autoimmune diseases. Autoimmun. Rev. 2015, 14, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Suryawanshi, A.; Manoharan, I.; Hong, Y.; Swafford, D.; Majumdar, T.; Taketo, M.M.; Manicassamy, B.; Koni, P.A.; Thangaraju, M.; Sun, Z.; et al. Canonical wnt signaling in dendritic cells regulates Th1/Th17 responses and suppresses autoimmune neuroinflammation. J. Immunol. 2015, 194, 3295–3304. [Google Scholar] [CrossRef] [Green Version]
- Flanagan, K.; Fitzgerald, K.; Baker, J.; Regnstrom, K.; Gardai, S.; Bard, F.; Mocci, S.; Seto, P.; You, M.; Larochelle, C.; et al. Laminin-411 is a vascular ligand for MCAM and facilitates TH17 cell entry into the CNS. PLoS ONE 2012, 7, e40443. [Google Scholar] [CrossRef]
- Garibaldi, S.; Barisione, C.; Ghigliotti, G.; Spallarossa, P.; Barsotti, A.; Fabbi, P.; Corsiglia, L.; Palmieri, D.; Palombo, D.; Brunelli, C. Soluble form of the endothelial adhesion molecule CD146 binds preferentially CD16+ monocytes. Mol. Biol. Rep. 2012, 39, 6745–6752. [Google Scholar] [CrossRef]
- Zhang, L.; Luo, Y.; Teng, X.; Wu, Z.; Li, M.; Xu, D.; Wang, Q.; Wang, F.; Feng, J.; Zeng, X.; et al. CD146: A potential therapeutic target for systemic sclerosis. Protein Cell 2018. [Google Scholar] [CrossRef] [Green Version]
- Stalin, J.; Traboulsi, W.; Vivancos-Stalin, L.; Nollet, M.; Joshkon, A.; Bachelier, R.; Guillet, B.; Lacroix, R.; Foucault-Bertaud, A.; Leroyer, A.S.; et al. Therapeutic targeting of soluble CD146/MCAM with the M2J-1 monoclonal antibody prevents metastasis development and procoagulant activity in CD146-positive invasive tumors. Int. J. Cancer 2020. [Google Scholar] [CrossRef]
- Larochelle, C.; Uphaus, T.; Broux, B.; Gowing, E.; Paterka, M.; Michel, L.; Dudvarski Stankovic, N.; Bicker, F.; Lemaître, F.; Prat, A.; et al. EGFL7 reduces CNS inflammation in mouse. Nat. Commun. 2018, 9, 819. [Google Scholar] [CrossRef] [PubMed] [Green Version]


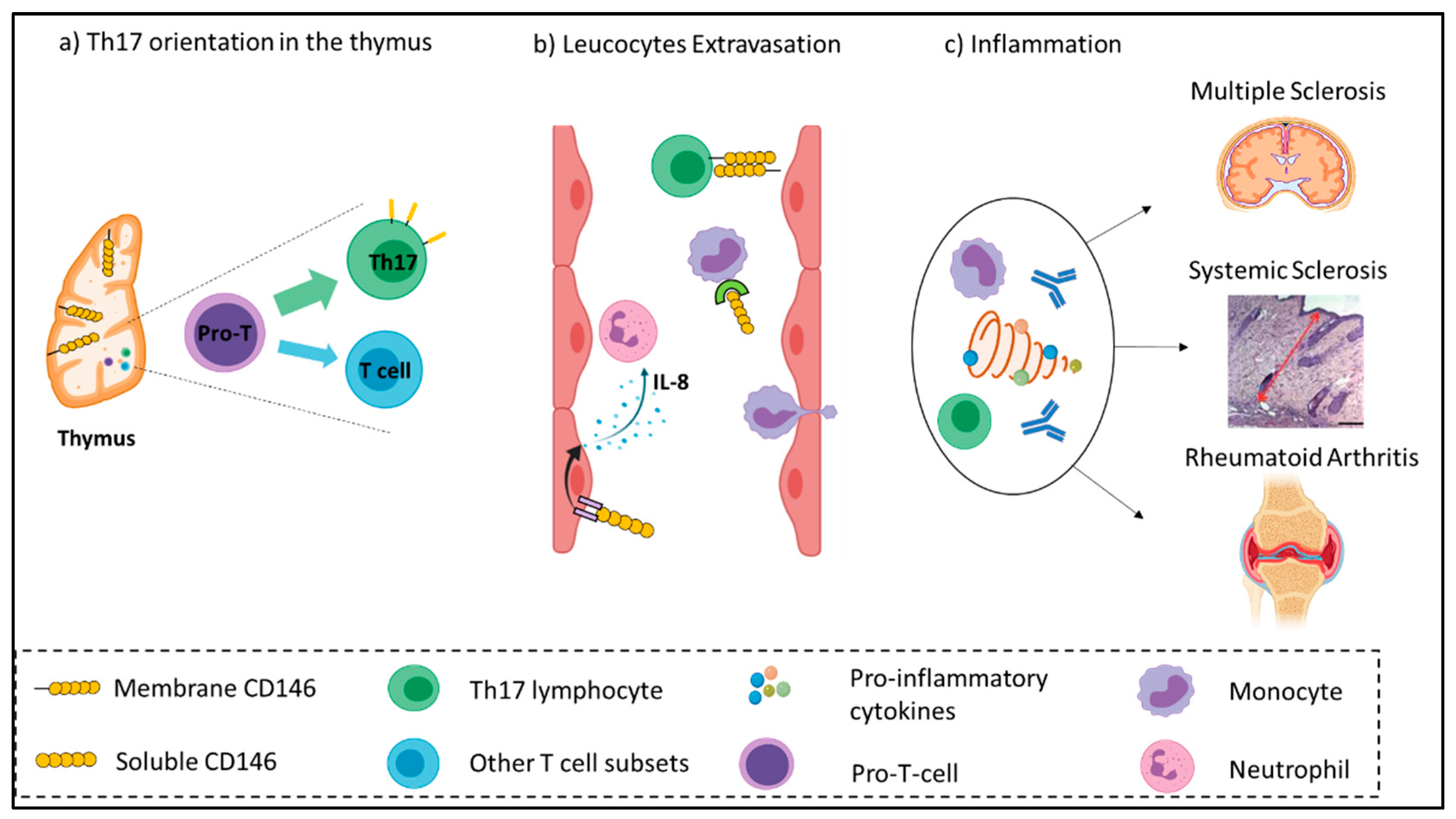{kind=link}
| Pathology | Membrane CD146 | Soluble CD146 | ||||
|---|---|---|---|---|---|---|
| Cell Population | Function | Ref | Fluid of Interest | Role | Ref | |
| Systemic sclerosis | Fibroblasts | Protects from fibrosis | Kaspi et al. [30] | Serum | Follow-up | Kaspi et al. [30] |
| Th17 cells | Migration | Gabsi et al. [32] | ||||
| Diabetes mellitus | Tubular epithelial cells | Not determined | Wang et al. [38] | Serum | Follow-up | Saito et al. [39] |
| Rheumatoid arthritis | Lymphocytes | Migration | Pickl et al. [49] | Synovial fluid | Prognosis | Neidhart et al. [47] |
| Inflammatory Bowel Disease | Endothelial cells | Not determined | Bardin et al. [56] | Serum | Follow-up | Bardin et al. [56] |
| Multiple sclerosis | Th17 cells | Migration | Breuer et al. [70] | CSF | Not determined | Duan et al. [11] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heim, X.; Joshkon, A.; Bermudez, J.; Bachelier, R.; Dubrou, C.; Boucraut, J.; Foucault-Bertaud, A.; Leroyer, A.S.; Dignat-George, F.; Blot-Chabaud, M.; et al. CD146/sCD146 in the Pathogenesis and Monitoring of Angiogenic and Inflammatory Diseases. Biomedicines 2020, 8, 592. https://doi.org/10.3390/biomedicines8120592
Heim X, Joshkon A, Bermudez J, Bachelier R, Dubrou C, Boucraut J, Foucault-Bertaud A, Leroyer AS, Dignat-George F, Blot-Chabaud M, et al. CD146/sCD146 in the Pathogenesis and Monitoring of Angiogenic and Inflammatory Diseases. Biomedicines. 2020; 8(12):592. https://doi.org/10.3390/biomedicines8120592
Chicago/Turabian StyleHeim, Xavier, Ahmad Joshkon, Julien Bermudez, Richard Bachelier, Cléa Dubrou, José Boucraut, Alexandrine Foucault-Bertaud, Aurélie S. Leroyer, Francoise Dignat-George, Marcel Blot-Chabaud, and et al. 2020. "CD146/sCD146 in the Pathogenesis and Monitoring of Angiogenic and Inflammatory Diseases" Biomedicines 8, no. 12: 592. https://doi.org/10.3390/biomedicines8120592