1. Introduction
Telomeres are specialised nucleoprotein complexes at the chromosomal ends containing repeated nucleotide sequences ((TTAGGG)n) and shelterin proteins [
1]. They protect the genome from nucleolytic degradation, undesired recombination, repair, and end-to-end chromosome fusion [
2] while they are maintained by a specialised enzyme, telomerase. The human telomerase enzyme is composed of 3 core subunits: (1) The telomerase RNA component (hTERC), (2) the catalytic subunit telomerase reverse transcriptase (hTERT) and (3) the dyskerin protein [
3].
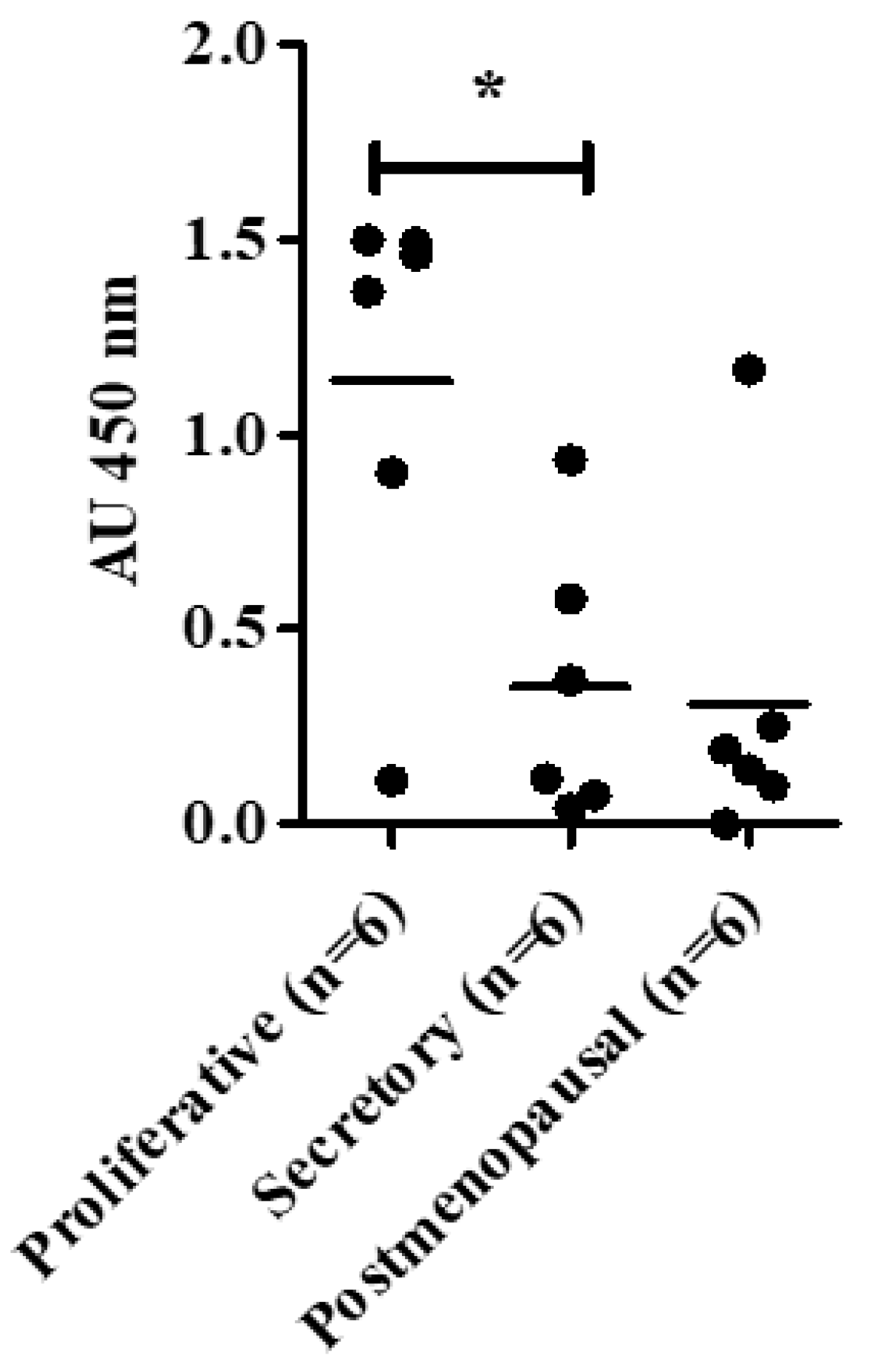
Telomerase activity (TA) shows dynamic changes in human endometrium correlating with the ovarian cycle and with glandular proliferation [
4]. Within the endometrium, stromal cells, regardless of the cycle phase, possess very low levels or absent TA and hTERC in comparison with epithelial cells [
4,
5,
6,
7]. Proliferating endometrial epithelial cells have the highest amount of TA [
4] and it was suggested that the elevated TA in epithelial cells could preferentially cap and maintain their short telomeres in order to protect endometrial epithelial cell telomeres from shortening to a critical length [
4]. Direct in vitro inhibition of TA using the TERC inhibitor “imetelstat” resulted in inhibition of cell proliferation and prevention of gland formation in healthy human endometrial epithelial cells [
4]. The relatively quiescent postmenopausal endometrium shows low levels of TA [
8].
Endometriosis is a common gynaecological disease defined as the presence of endometrial glands and stroma-like lesions outside the uterus and it is an oestrogen-dependent disease, which can cause significant morbidity in a large number of women of reproductive age [
9,
10]. Unfortunately, there are no curative treatments and none of the available treatments is effective in managing the symptoms associated with this disease [
11]. The pathogenesis of endometriosis is not yet fully elucidated in order to develop new treatments for endometriosis-associated pain and infertility. Therefore, women with endometriosis continue to suffer and they have a decreased quality of life [
11].
The best-accepted theory of the pathogenesis of endometriosis is retrograde menstruation, by seeding of eutopic endometrium in the pelvis that initiates endometriotic deposits [
12]. However, this phenomenon of retrograde menstruation occurs in almost all women [
13] while the prevalence of endometriosis in the general population is only 10–15% of women. Thus, this theory had subsequently been modified to propose that eutopic endometrium shed by women with endometriosis to have particular aberrations, enhancing its ectopic growth [
14,
15]. Supporting this theory, there is a plethora of evidence confirming that the endometrium of women with endometriosis differs from that of healthy fertile women [
16,
17]. High TA levels, higher
hTERT gene expression and hTERT protein levels associated with longer mean endometrial telomere length (TL) have all been reported to be characteristics of such abnormalities specific to eutopic secretory endometrium of women with endometriosis when compared with the endometrium of healthy women [
7,
14,
18,
19,
20,
21].
During retrograde menstruation, elevated TA in late-secretory endometrium of women with endometriosis [
19] may cause the sloughed cells to survive in the peritoneal cavity and establish ectopic lesions [
4,
14]. While TA, hTERT, and TL have been studied in the context of endometriosis; there have not yet been any reports published on the levels of the remaining core components, hTERC and dyskerin, in the endometrium of women with endometriosis to date.
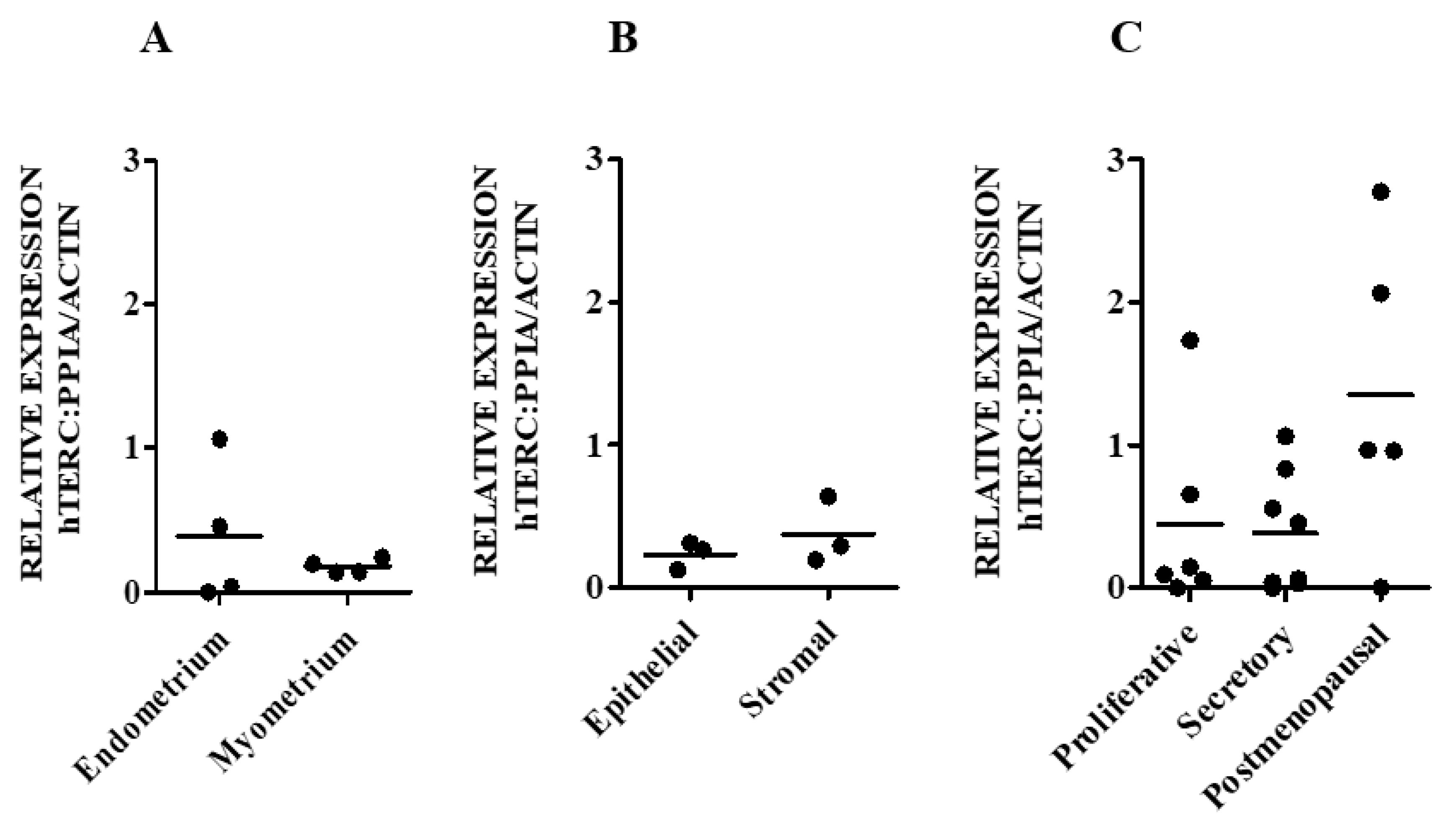
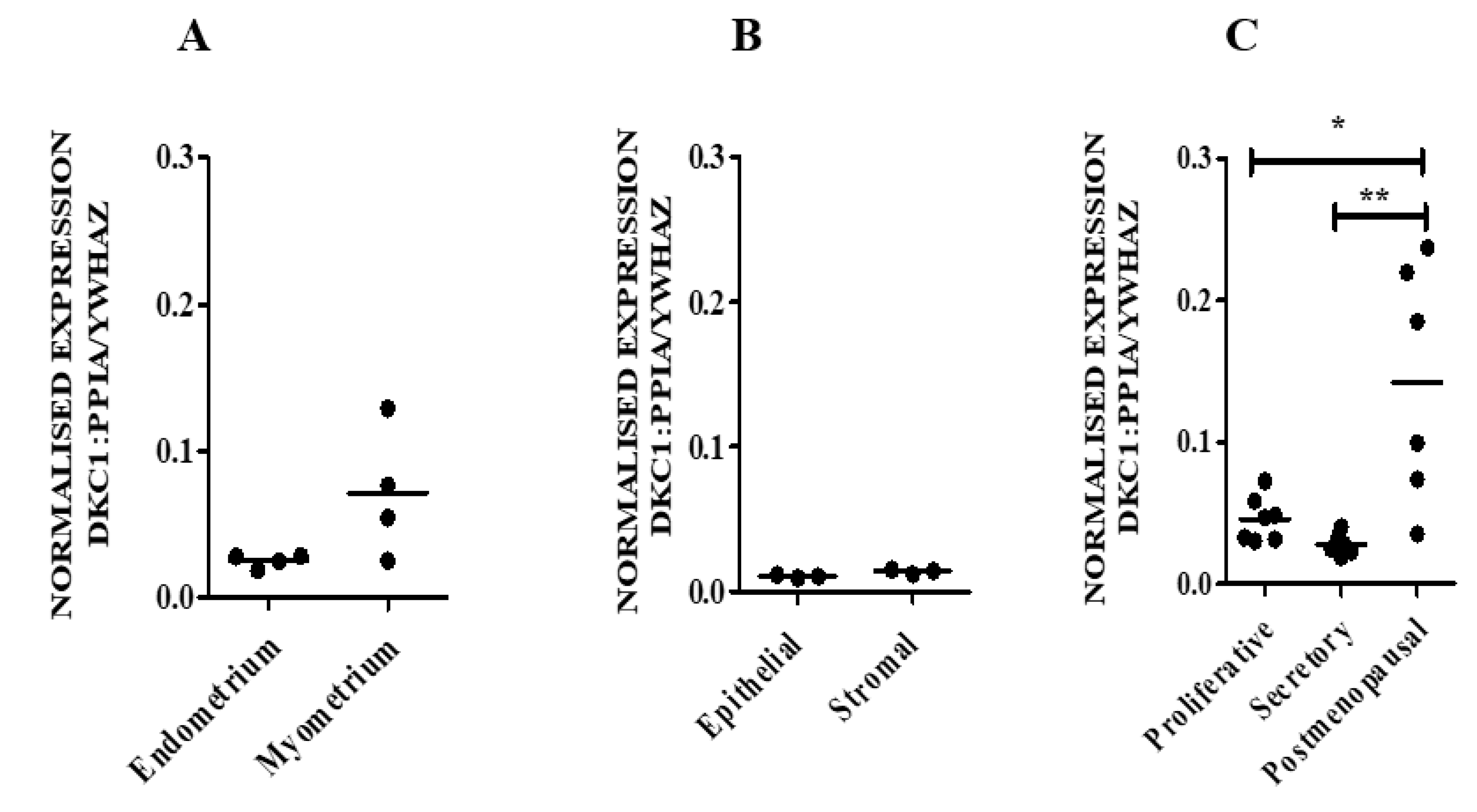
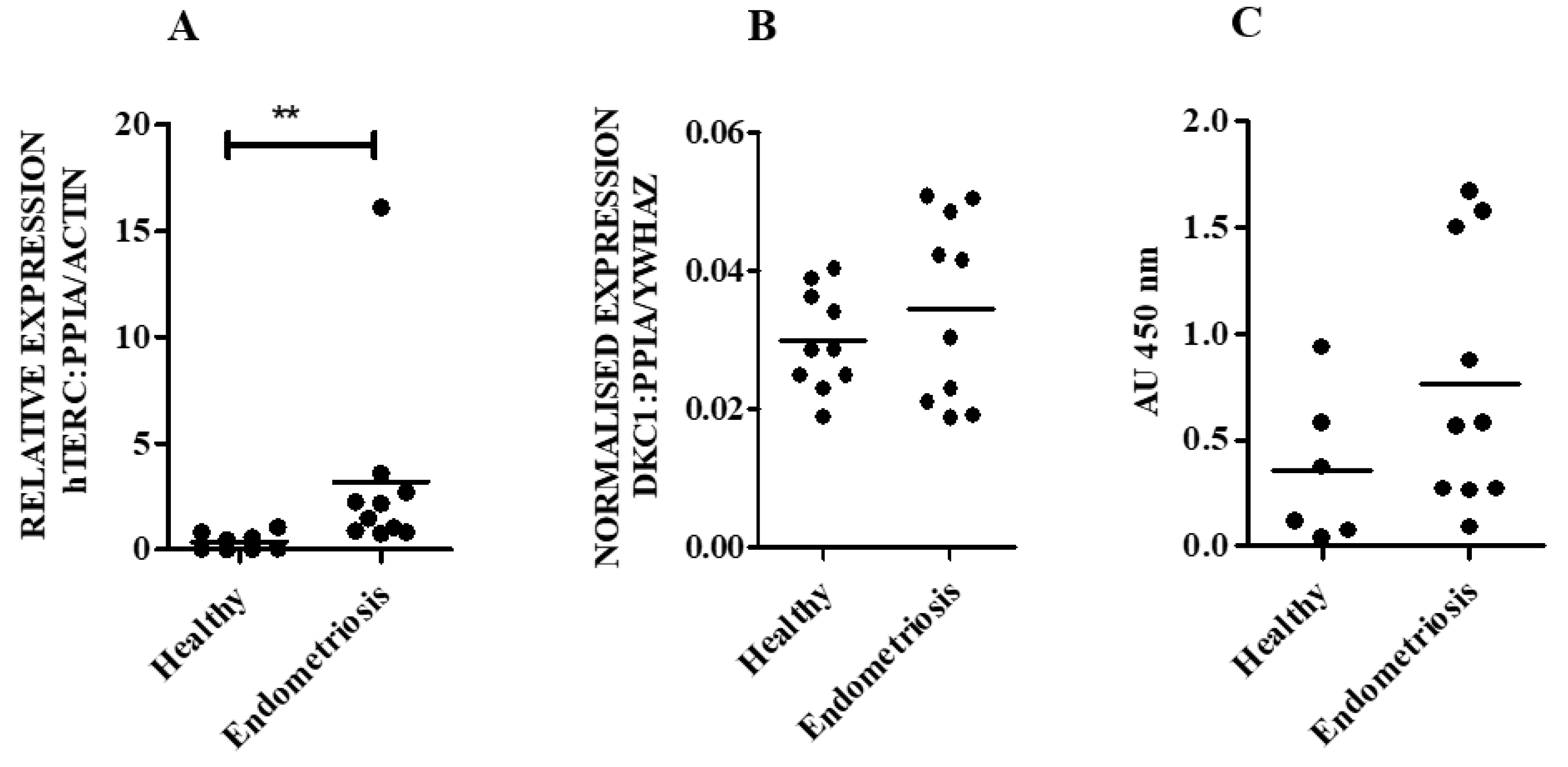
Therefore, our aim was initially to examine the levels of hTERC and DKC1 RNA as well as dyskerin protein levels in normal endometrial tissue obtained from healthy women and in endometria collected from those with a surgical diagnosis of benign proliferative endometrial disease, endometriosis. We then progressed to an in silico study, where endometriosis-specific telomere- and telomerase-associated transcriptional aberrations of secretory phase eutopic endometrium were explored. Using a comprehensive telomere- and telomerase-associated genes (TTAGs) list that was compiled in our previous work [
22], differentially regulated TTAGs were examined in published human endometrial microarray datasets from women with and without endometriosis, which were selected according to a strict predetermined criteria.
4. Discussion
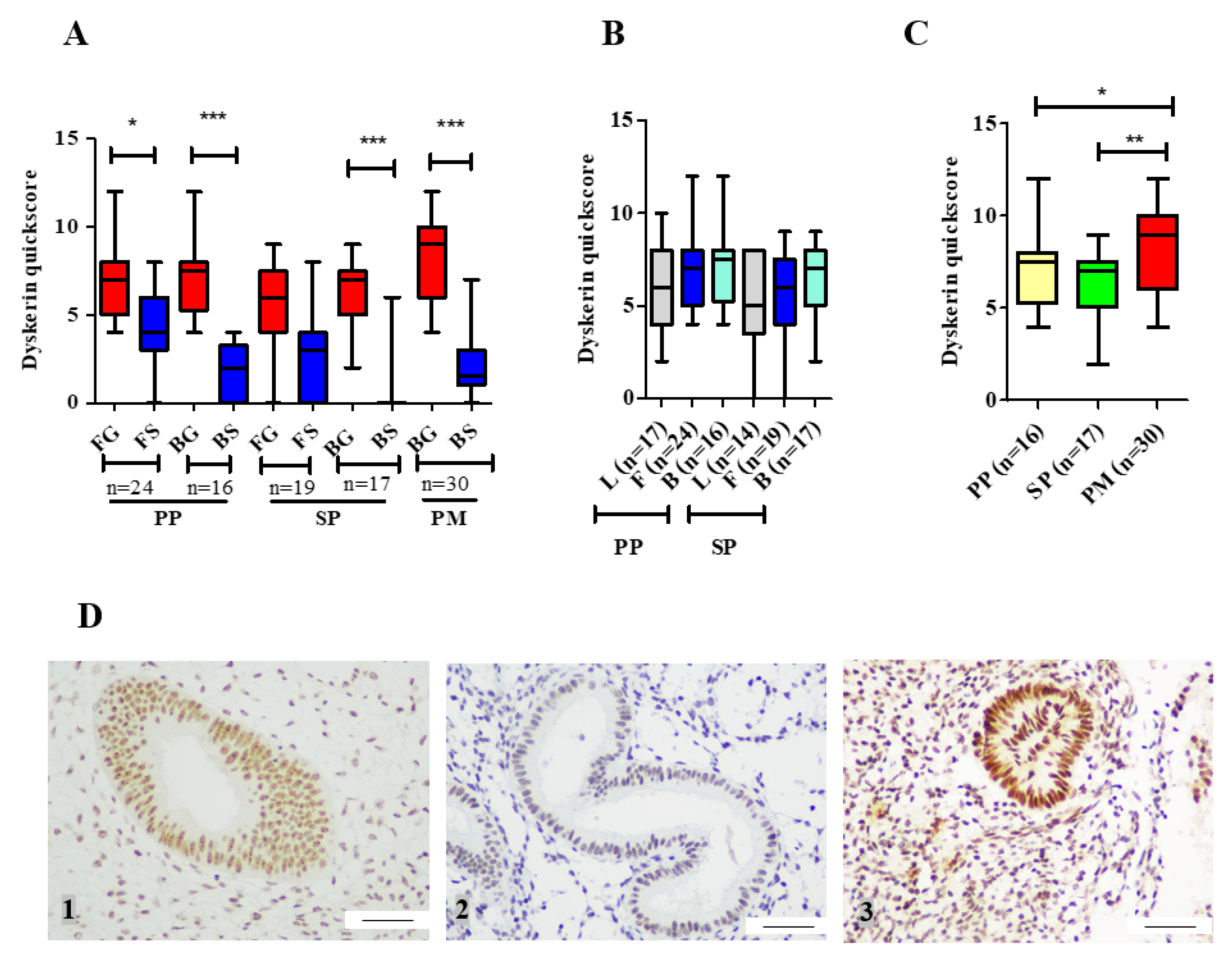
Involvement of telomerase in endometrial biology is well established. Previous reports have demonstrated human endometrial epithelial proliferation to be associated with high TA and increased hTERT expression [
4,
8]. Consequently, increased TA and
hTERT levels were observed in the endometrium of women with the proliferative endometrial disease, endometriosis [
18,
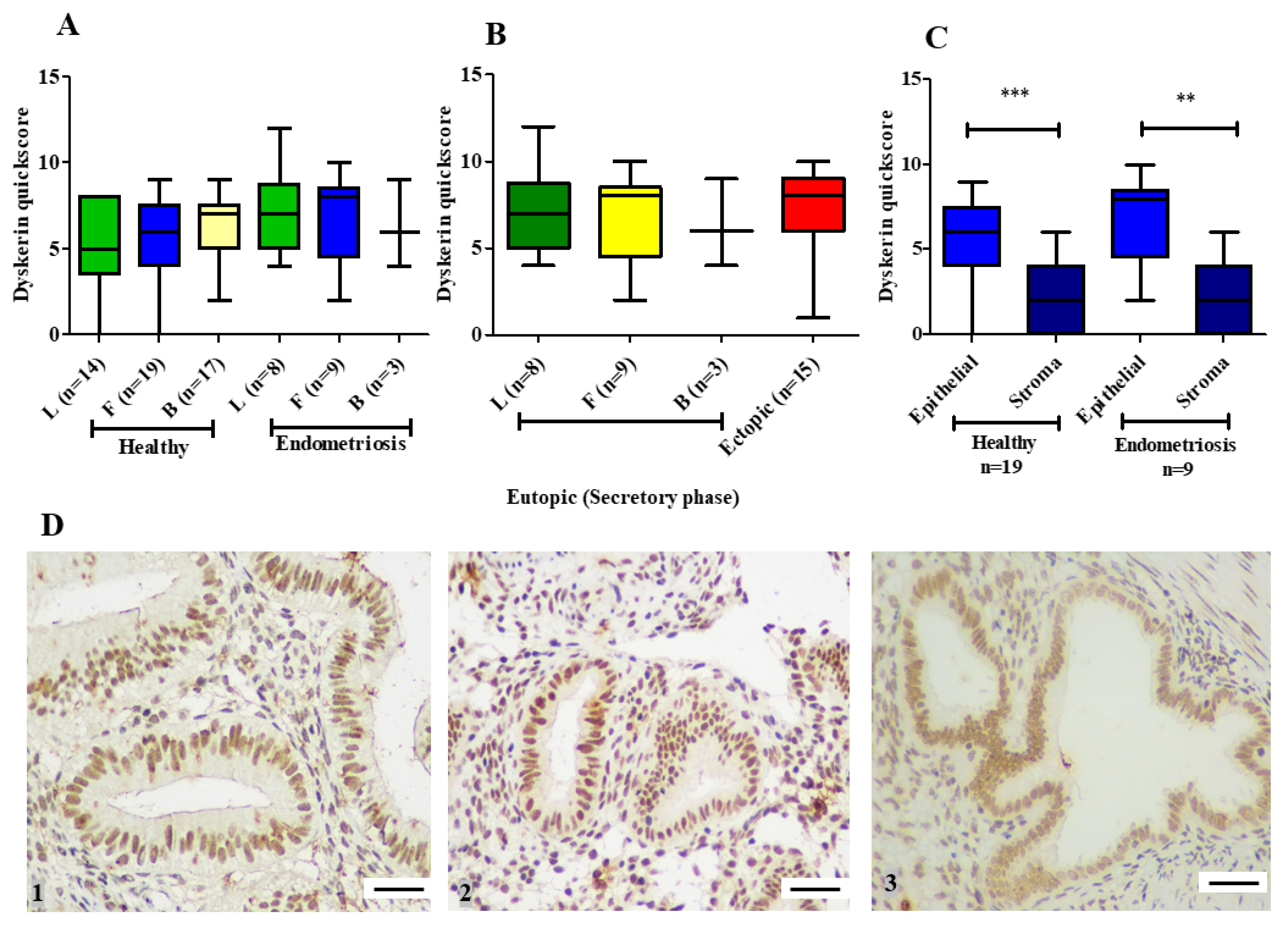
19]. Our ex-vivo study with patient-derived endometrial samples initially examines the involvement of the remaining two main core components of telomerase holoenzyme (hTERC and dyskerin) in healthy human endometrium across all stages of the premenopausal menstrual cycle, in postmenopausal endometrium as well as the involvement of these two components in eutopic endometrium of women with endometriosis. The dyskerin immunostaining of ectopic endometriotic lesions was also examined. Our data demonstrate dynamic changes in these components in pre/postmenopausal samples and reveals that eutopic secretory endometrial hTERC RNA levels are significantly increased in women with endometriosis. Contrastingly, dyskerin (DKC1) mRNA and protein levels were unperturbed in the secretory phase eutopic endometrium of women with endometriosis. Subsequently, we expanded this work with an in silico study, analysing available microarray datasets to determine the differentially regulated TTAGs in the mid-secretory endometrium of women with endometriosis to determine the involvement of these in known aberrations of telomerase biology and cellular function in the pathogenesis of endometriosis. Interestingly,
hTERT was not upregulated in either of the datasets; this is contrary to prior findings that
hTERT expression is higher in the eutopic secretory endometrium of women with endometriosis when compared with the endometrium of healthy women [
7,
14,
18,
19,
20,
21]. This is consistent with previous observations in endometrial cancer, a condition associated with high TA Even the large TCGA dataset did not show an increased expression of
hTERT in women with endometrial cancer [
22]. Consistent with experimental data, mid-secretory expression of
DKC1 in mid-secretory endometrium from women with endometriosis was not significantly different when compared with the control healthy samples according to published datasets. Although no expression data were available for
hTERC, 10 additional TTAGs were found to be altered in mid-secretory endometrium of diseased patients (
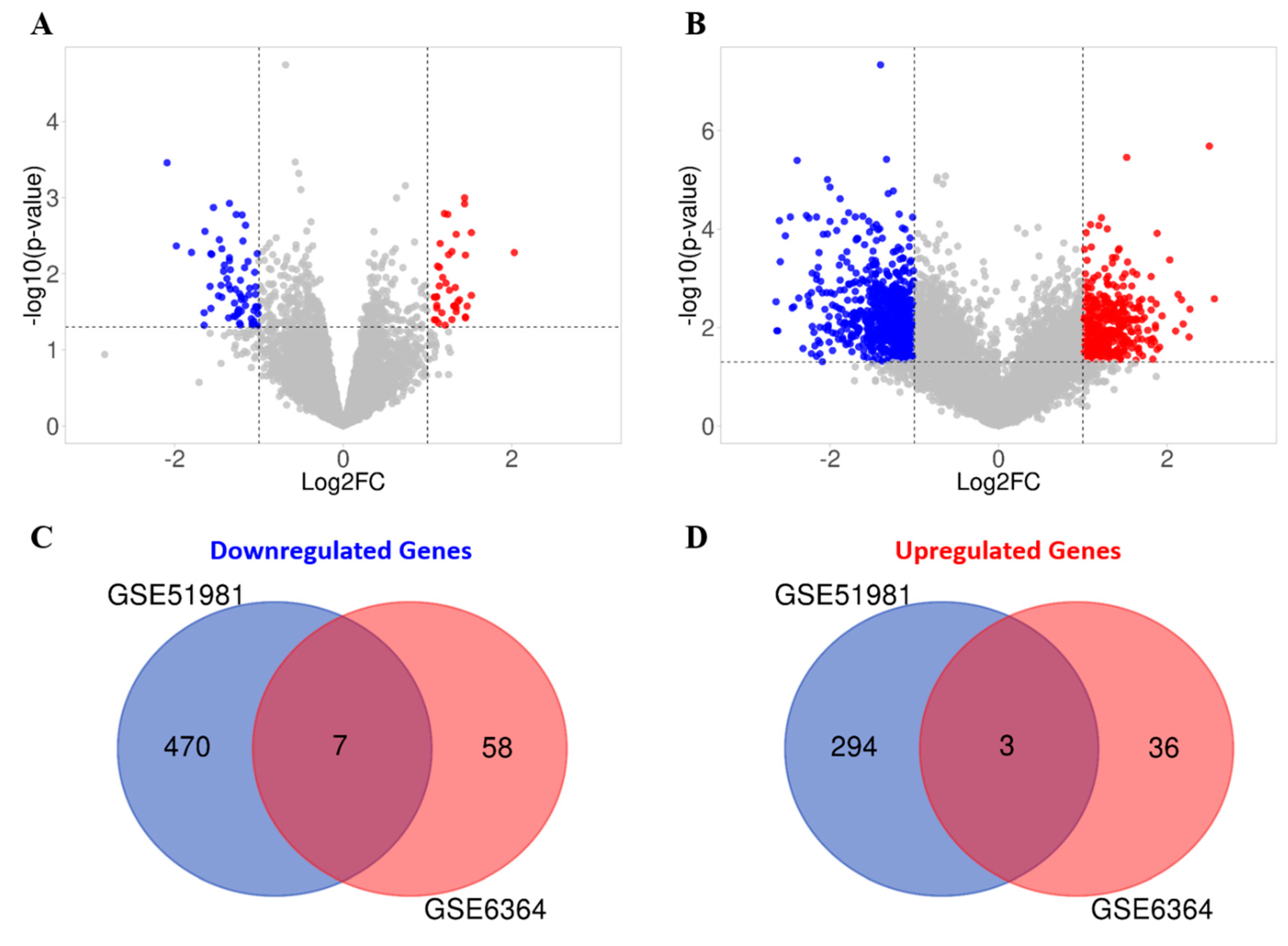
Figure 8).
The current consensus is that all core components of the telomerase enzyme will contribute to TA [
3]. The endometrial TA and hTERC RNA levels did not correlate in our study, and previous authors have reported that the enzymatic level of TA measured by TRAP assay, does not necessarily correlate with
hTERC expression [
39,
40]. hTERC is ubiquitously expressed in all tissues, thus some have suggested hTERC levels to be not directly relevant to TA [
39], therefore, supporting our observation in the endometrium. In vitro studies have suggested that hTERT needs hTERC as a minimal requisite for TA but that the main catalytic subunit hTERT might be the rate limiting feature relevant to TA [
41]. In the human endometrium, epithelial TA and hTERT RNA levels have been relevant to cell proliferation and are regulated by ovarian steroid hormones, progesterone and estradiol [
4,
7]. Eutopic endometrium of women with endometriosis appears to be different to that of women without endometriosis and the aberration in the endometrial functionalis layer in these women have been proposed to play a role in the pathogenesis of endometriosis [
7,
9,
14,
15,
42,
43]. Previous studies have also demonstrated that TA, hTERT protein and mRNA levels were increased in the secretory phase endometrium of women with endometriosis [
18,
19]. The high TA accompanying the pro-proliferative, anti-apoptotic, and senescence-evading phenotype found in the secretory phase endometrium of women with endometriosis, has been proposed to contribute to endometriotic lesion formation due to deposition of sloughed endometrial fragments after retrograde menstruation [
14,
20]. Our study examined hTERC and
DKC1 expression levels in eutopic endometrium of women with endometriosis demonstrating significantly higher hTERC RNA in the secretory phase compared with healthy secretory endometrium. RNA interference of hTERC reduced cancer cell proliferation rate quickly, without altering TA [
44]. We therefore hypothesise that the high hTERC RNA levels found in the eutopic endometrium of women with the benign proliferative disease, endometriosis, may facilitate the postulated increased proliferative potential of these cells, which may also be a reflection of increased TA, but dyskerin may not play a major role in the pathology of endometriosis.
Dyskerin is the main component of the dyskerin core complex (dyskerin-NHP2-NOP2) that binds and stabilises hTERC, and thus enhances TA. The expression levels of both genes are expected to follow a similar pattern to endometrial
hTERT and TA. However, the regulation of the steady-state levels of hTERC is far more complex [
45]. hTERC transcripts undergo a crucial multistep process of maturation, which collectively controls the levels of hTERC and dysregulation of these steps in various pathologies may influence TA. Functional TA in vivo, however, requires more than just hTERT and hTERC, but dyskerin and other associated components of the larger telomerase holoenzyme as well as some proteins that are only transiently associated with the holoenzyme [
45].
There is previous data however, supporting an intricate relationship between hTERC and dyskerin, where a reduction in DKC1 mRNA or dyskerin protein was shown to be associated with a concomitant reduction in hTERC levels. For example, the A353V mutation in the
DKC1 gene, which is the most common mutation in X linked dyskeratosis congenita (X-DC) patients, causes a reduction in the levels of hTERC RNA [
46], while another report revealed that loss of dyskerin caused degradation of hTERC in vivo [
47]. In breast cancer tissue, when DKC1 mRNA levels were very low, TA was significantly decreased, independently of the level of
hTERT expression [
48]. Additionally, in vitro
DKC1 gene knock-down using siRNA in MCF-7 breast cancer cells, reduced TA via a reduction in hTERC levels [
48]. Taken together, this evidence suggests the simultaneous reduction of DKC1 and hTERC was associated with a concomitant reduction in TA. In contrast, in our study we only found changes in hTERC RNA levels but not for dyskerin expression and protein levels. Unlike premenopausal endometrial TA levels, hTERC and DKC1 RNA levels do not demonstrate a significant change across the menstrual cycle. Furthermore, in complete contrast to TA, healthy premenopausal endometrial hTERC and DKC1 RNA levels were low in premenopausal endometrium while significantly increased levels were observed in proliferatively quiescent postmenopausal samples. When previous evidence is considered in this context, although knocking down
DKC1 influenced TA via hTERC in MCF-7 cells as mentioned above, an excessive reduction of hTERC levels was required to decrease TA [
48]. Thus, we propose that the reduced levels seen in hTERC in premenopausal proliferative endometrium in the context of high TA levels, may still not have reached the threshold level to affect endometrial TA. In contrast, in the postmenopausal endometrium, where TA was low, DKC1 RNA level was high in comparison with premenopausal samples; therefore, we hypothesise that the high DKC1 levels could possibly have a direct consequence on endometrial TA. Since dyskerin plays an important role in stabilising hTERC, high levels of dyskerin protein may be important in resisting carcinogenesis in the postmenopausal endometrium via the non-telomerase related pathway. Dyskerin acts also as pseudouridine synthase which catalyses the isomerisation of uridine to pseudouridine in rRNA.
DKC1 mutations cause ribosomal dysfunction and result in a reduction of tumour suppressive proteins. For example, in a X-DC mouse model, defective translation of specific mRNAs harbouring internal ribosomal entry site (IRES) elements occurred due to decrease in dyskerin activity, and also reduced the tumour suppressor, p27, levels and caused a marked increase in spontaneous pituitary tumorigenesis in p27 heterozygous mice [
49]. Further studies are therefore required to examine the functional consequence of over-expression of dyskerin, such as cellular proliferation.
Our novel finding that hTERC RNA levels are significantly higher in eutopic endometrial samples in the secretory phase of women with endometriosis, without altered levels of DKC1 mRNA, seems interesting. The high TA [
19], hTERT [
18,
19], and hTERC (present study) levels may promote endometrial epithelial proliferation in women with endometriosis independent of dyskerin.
It is an established fact that not all excised endometriotic ectopic lesions contain endometrial epithelial and stromal-like cells are found during histological scrutiny. Techniques such as immunohistochemistry directly visualise the specimen to ascertain the cellular content, but other techniques such as PCR do not allow such assessment. Considering this discrepancy, we did not assess the RNA levels of hTERC or DKC1 in ectopic lesions. This inherent difficulty in ascertaining whether the excised presumed endometriotic lesions actually contained endometrium-like cells can be overcome in the future with employing specialist techniques such as in situ hybridisation.
By interrogating the published endometrial microarray datasets, we have identified 3 TTAGs that are specifically upregulated in the mid-secretory endometrium.
CDKN2A being the most significantly upregulated (
Table 4), is a known tumour suppressor gene, known to promote cell senescence [
50]. The gene is known to decrease during cell proliferation therefore, our findings of CDKN2A upregulation corresponds to the expected halt in endometrial proliferation during the secretory phase [
51]. Regardless, the loss of p16 has been described in many cancer entities [
51] while in contrast, its high expression has been linked to poor prognosis of endometrial cancer, where telomerase and telomere activity are known to play an important role [
52]. Moreover, there is a significant amount of evidence on altered p16 levels in eutopic endometrium in women with endometriosis [
53]. Promyelocytic leukaemia (PML) is a critical component to the alternative telomere lengthening (ATL) pathway; its effects are achieved through ALT-associated PML bodies (APBs) [
54]. APBs have recently been shown to drive BLM–TOP3A–RMI (BTR) accumulation which is known to promote break-induced replication-mediated telomere elongation [
55]. Upregulation of PML may therefore suggest increased telomere-lengthening in endometriosis-positive mid-secretory endometrium. ZNHIT2 is a member of the zinc finger HIT domain-containing protein family and has not been well-characterised [
56]. A recent study has demonstrated its role in regulating spliceosome activity [
56]; spliceosome is a complex small nuclear RNA-protein machine responsible for removing introns from pre-mRNA; hence, known to regulate alternative splicing of a gene [
57]. Therefore, altered ZNHIT2 levels seen in endometriosis may dysregulate alternative splicing of hTERT which is often found in cancers [
58]. A total of 7 TTAGs were found to be commonly downregulated in both GEO datasets, suggesting that they may contribute to endometriosis pathogenesis. FGFR2 and PIK3C2A have both previously been proposed to contribute to endometriosis-related carcinogenesis [
59,
60], but their role in benign endometriosis has not been elucidated. PIK3C2A is associated with telomerase as a result of its yeast homologue, VPS34, being linked with telomere biology [
61,
62,
63]. However, FGFR2 was thought to be associated with telomerase due to its downregulation in telomerase deficient cells in a previous study [
63]. This is in contrast with our finding that FGFR2 was downregulated in the mid-secretory phase eutopic endometrium of women with endometriosis—a telomerase abundant tissue. FGFR2, which encodes fibroblast growth factor receptor 2, is involved in a wide array of cellular processes, such as cellular proliferation, differentiation, wound repair and regulation of tissue fibrosis [
64]; its downregulation in endometriosis may be related to an alternative cellular process, irrespective of telomerase regulation. Both HNRNPA2B1 and HSPC159 have previously been linked with epithelial–mesenchymal transition (EMT) in cancer [
65,
66]; EMT is thought to drive malignant invasion and metastasis and has also been proposed to contribute, in part, to the detachment and dissemination of endometrial cells in endometriosis [
67]. HNRNPA2B1 is linked with telomere biology as it has been shown to be upregulated upon telomere shortening in vitro [
68] HSPC159 has been found in a fluorescence localisation screen to be located in close proximity to TINF2 [
69], a protein from the shelterin protein complex that is bound to the telomere-binding proteins TRF1 and TRF2 [
70]. The remaining identified genes,
UBE3A,
MCCC2, and
RALGAPA1, have not previously been linked with endometrial disease, and very little is known about the actions of
MCCC2 and
RALGAPA1.
UBE3A is linked with telomere function as it has previously been shown to induce
hTERT transcription and telomerase activity [
71,
72], whereas MCCC2 and RALGAPA1 have previously been found in a protein network surrounding TRF1, TRF2, and POT1 [
73], which are all components of the shelterin complex [
70]. A limitation to these in silico findings is that some of these TTAGs are only weakly associated with telomere function; thus, their dysregulation in endometriosis may be representative of other cellular functions. Our findings are also limited by the small sample size of the datasets. Nonetheless, we have identified multiple novel genes that potentially contribute to endometriosis and may warrant further study. Although the sample size is a major limitation of our ex-vivo study, we have selected a well characterised population of women at a specific time point in the cycle to reduce variability and identified a statistically significant difference in hTERC levels between women with and without endometriosis, which, in the context of prior data on endometrial telomerase and progesterone resistance, is biologically plausible. Future studies thus are warranted to further confirm this data by appropriately increasing the sample size.

 and
and 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}