
MicroRNAs as Molecular Biomarkers for the Characterization of Basal-like Breast Tumor Subtype

Discipline of Surgery, Lambe Institute for Translational Research, H91 TK33 Galway, Ireland
*
Authors to whom correspondence should be addressed.
Biomedicines 2023, 11(11), 3007; https://doi.org/10.3390/biomedicines11113007
Submission received: 11 October 2023
/
Revised: 3 November 2023
/
Accepted: 7 November 2023
/
Published: 9 November 2023
(This article belongs to the Special Issue Molecular Research of Triple-Negative Breast Cancer)
Abstract
:Breast cancer is a heterogeneous disease highlighted by the presence of multiple tumor variants and the basal-like breast cancer (BLBC) is considered to be the most aggressive variant with limited therapeutics and a poor prognosis. Though the absence of detectable protein and hormonal receptors as biomarkers hinders early detection, the integration of genomic and transcriptomic profiling led to the identification of additional variants in BLBC. The high-throughput analysis of tissue-specific micro-ribonucleic acids (microRNAs/miRNAs) that are deemed to have a significant role in the development of breast cancer also displayed distinct expression profiles in each subtype of breast cancer and thus emerged to be a robust approach for the precise characterization of the BLBC subtypes. The classification schematic of breast cancer is still a fluid entity that continues to evolve alongside technological advancement, and the transcriptomic profiling of tissue-specific microRNAs is projected to aid in the substratification and diagnosis of the BLBC tumor subtype. In this review, we summarize the current knowledge on breast tumor classification, aim to collect comprehensive evidence based on the microRNA expression profiles, and explore their potential as prospective biomarkers of BLBC.
1. Introduction
Breast cancer is the most commonly diagnosed malignancy in women globally with over 2 million new cases diagnosed in 2020 [1]. The American Cancer Society projected 300,590 new cases of breast cancer estimated to account for 43,700 deaths in the United States alone in 2023 [2]. Breast cancer is a highly heterogeneous disease and can be further categorized based on various parameters including and not limited to location of origin, degree of cellular pleomorphism, molecular composition, etc. [3,4]. The classification of breast tumors based on the expression of certain hormone receptors, analyzed via immunohistochemistry (IHC) is a conventionally recognized method [5]. The hormone receptors used are the following: estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2) [5,6]. Based on the expression of the aforementioned receptors, four subtypes of breast cancer are widely recognized and are as follows: luminal A, luminal B, HER2+, and triple-negative breast cancer (TNBC) [7]. The triple-negative breast tumor subtype lack the expression of ER, PR, or HER2 receptors and account for 10–15% of total breast cancer cases [8]. TNBC records a high recurrence rate with an inferior prognosis relative to non-TNBC [9]. The prognosis and treatment regimen for breast cancer can differ significantly from patient to patient owing to the substantial heterogeneity between primary malignant tumors [10].
The term “triple-negative breast cancer” is often used synonymously with “basal-like breast cancer”, and although the two possess a degree of resemblance to one another, they are not entirely interchangeable terms [11]. The exact definition of basal-like breast cancer (BLBC) is a topic of debate, but the title was initially designated to tumors characterized by an expression of markers for basal epithelial cells including cytokeratins (CK) 5/6 and CK17 with low or negative ER expression [12]. TNBCs and BLBCs are both associated with higher grade disease, larger size tumors, and aggressive histopathologic characteristics [13]. Individuals with BLBCs have decreased overall survival, with a decreased disease-free period alongside a statistically significant increased risk of recurrence post-chemotherapy relative to other subtypes [14]. Considering the aggressive nature and disease process of BLBCs, there still exists a gap in the therapeutic options for the pathology and, as of yet, chemotherapy remains the mainstay [15]. Therefore, early disease detection and the development of targeted treatment are imperative to reducing death rates and improving BLBC patient survival outcomes and prognosis [16].
Researchers have worked to segregate breast cancers in many ways, including the IHC-mediated proteomic approach of classifying tumors based on hormone receptor expression as previously mentioned [7]. The genome-based partition of breast cancer is another potential approach, and a well-accepted genomic design is the 50-gene microarray panel PAM50 (prediction analysis of microarray 50) [17]. A study by Krug et al. highlights a proteogenomic technique that may be used to identify clinical and molecular features of specific tumors for the development of tumor-specific treatment targets [18]. Transcriptomic characterization of malignant breast tumors based on microRNAs is a novel approach currently under ongoing investigation [19,20,21,22].
MicroRNAs are established post-transcriptional modulators of gene and consequentially protein expression that play a pivotal role in the maintenance of bodily homeostasis and the evolution of various pathological processes [23,24]. MicroRNAs have the ability to travel extracellularly in blood and plasma, prompting them to be the ideal subject for the development of a non-invasive molecular-based system for precise identification and classification [22,25]. This review aims to explore the use of a miRNA-based model to diagnose, subclassify, predict treatment response, evaluate the prognosis, and formulate potential targeted therapeutics for the BLBC subtype.
2. Evolution of the BLBC Classification System
From the initial four subtype classification system based on IHC data, breast cancer subtype classification has continued to develop throughout the years. Various other panels have been suggested to specifically identify BLBCs, one of which was introduced by Nielsen et al. that claims an IHC-based panel of ER, human epidermal growth factor receptor 1 (HER1), HER2, and cytokeratin (CK) 5/6 can distinguish BLBC tumors with high specificity [26]. Cheang et al. introduced a similar expanded immunopanel that consisted of ER, PR, HER2, EGFR, and CK5/6 in order to implement a precise definition of BLBC [27]. Livasy et al. found that BLBC tumors most coherently exhibited an immunophenotype of negative ER and HER2 expression in addition to CK5/6, CK8/18, epidermal growth factor receptor (EGFR), and vimentin positivity [28]. In 2007, Herschkowitz et al. discovered a subtype of breast cancer and referred to this as “claudin-low” tumors that are characterized by low expression of claudins 3, 4, 7, occludin, and E-cadherin [29]. Prat et al. then characterized the “claudin-low” subtype phenotypically and molecularly and found it to be ER negative [ER−], PR negative [PR−], and HER2 negative [HER2−], but showed an inconsistent expression of basal cytokeratins [30]. Claudin-low tumors were found to display phenotypic and molecular characteristics shared between the basal-like and luminal subtypes, henceforth bringing upon the emergence of another intrinsic subtype of breast cancer [30].
Parker JS introduced a 50-gene centroid-based breast cancer subtype identification model to compare the reproducibility of subtype assignment through the use of a prediction analysis of microarray (PAM) algorithm termed PAM50 [17]. They used Spearman’s rank correlation to identify the nearest simple centroid, which was then assigned as one of the following molecular intrinsic subtypes: luminal A, luminal B, HER2-enriched, basal-like, and normal-like [17,31]. The PAM50 gene microarray panel has been shown to subclassify breast cancer and the risk of relapse (ROR) with a high degree of reproducibility and validity, making it a commonly implemented system [17,32].
The Cancer Genome Atlas (TCGA) project incorporated molecular data of 825 breast cancer patient samples across five technological platforms and rendered four distinct heterogeneous breast cancer subtypes: luminal A, luminal B, HER2-enriched, and basal-like [33]. A novel breast cancer subtype classification system based on messenger RNA (mRNA) signatures incorporated with the PAM50 50-gene signatures identified seven breast cancer subtypes with clearly defined groups of gene expression patterns [34]. The distinct taxonomic groups proposed are as follows: basal/HER2, basal/myoepithelial, myoepithelial/luminal/HER2, myoepithelial/luminal, luminal, basal/luminal, and basal/luminal/HER2 [34]. A phenogenomic analysis of 483 breast cancer tumors by Ali et al. incorporated single-cell phenotypical analysis alongside genomic profiling and rendered their own distinct subgroups, which is a testament to the ongoing progression of breast tumor subclassification [35]. New microRNA-based panels may yield a more efficient, uniform, and clinically pertinent BLBC signature for the identification and prognostic indication and are in continued development [22,36]. To that point, Søkilde R et al. conducted transcriptomic analysis and sequencing of miRNAs on 186 PAM50 classified breast cancer samples from the Sweden Cancerome Analysis Network–Breast (SCAN-B) trials and categorized breast cancer based on the miRNA expression profiles [19,37]. The work conducted by Søkilde R et al. has paved the way for future validation studies and the progression of miRNA-based breast cancer research (Figure 1).
3. Significance of MicroRNAs in Breast Tumor Pathogenesis
3.1. Biogenesis of MicroRNAs
MicroRNAs (miRNAs) are small non-coding RNA molecules, typically 21–24 nucleotides in length, that are endogenously expressed and play a crucial role in post-transcriptional gene expression regulation, leading to variations in protein expression levels [38,39]. In the human genome, genes encoding miRNAs are predominantly located within intronic sequences, with a minority originating from exons or as intergenic sequences, each possessing its own promoter [40]. MicroRNAs undergo a well-planned series of events throughout their formation [41].
The transcription of miRNA genes yields primary miRNAs (pri-miRNAs) that undergo a two-step cleavage process, ultimately forming mature miRNA [41]. The process begins with RNA Polymerase II (RNA Pol II) transcription of the miRNA genes, which is followed by nuclear processing made possible by the Drosha ribonuclease III (DROSHA)-DiGeorge syndrome critical region 8 (DGCR8) complex [42]. The protein DGCR8, which binds to double-stranded RNA, aids in identifying the stem-loop structure present in pri-miRNAs [42]. Pre-miRNAs are shorter hairpin-shaped RNA molecules that are produced as a result of the ribonuclease DROSHA cleaving the pri-miRNA at a particular point inside the stem loop [43]. Exportin-5 (XPO5) then transports these pre-miRNAs from the nucleus to the cytoplasm [44]. These mature miRNAs integrate into the RNA-induced silencing complex (RISC), a key effector complex [45]. MiRNAs exhibit versatile modes of action, including binding to the 3′ untranslated region (UTR) of target mRNAs to repress translation and induce mRNA destabilization [46,47]. Additionally, they can bind to the 5′ UTR, resulting in gene expression silencing, or directly interact with promoter regions of protein-coding genes, eliciting transcription [40,48]. Remarkably, a single miRNA has the potential to target hundreds of distinct mRNAs, rendering the overall impact of miRNAs on gene regulation highly intricate [49].
3.2. The Dual Regulatory Roles of MicroRNAs in Cancer
MiRNAs play pivotal roles in various biological processes, including nervous system regulation, cell differentiation, development, viral infection, angiogenesis, cancer, gastrointestinal diseases, and diabetes [50]. Eugene Makeyev and Tom Maniatis enrich our understanding of miRNA versatility and also offer a compelling explanation for the surprisingly minimal disparities in protein-encoding genes across organisms of varying morphological and behavioral complexities, emphasizing the pivotal role miRNAs play in shaping intricate gene regulatory networks during cellular differentiation [51]. The evolution of cancer within an individual is the product of the malignant cells and the extrinsic factors interacting with and potentially dictating the behavior of said cells. The hallmarks of cancer (tumor-promoting inflammation, inducing or accessing vasculature, genome instability and mutation, resisting cell death, etc.) may all be possible targets of miRNAs [52]. MicroRNA dysregulation can result from multiple mechanisms, including gene locus expansion inducing gene overexpression, mutations leading to loss of function or deregulation, epigenetic modifications influencing gene activation or suppression, and trans-acting elements on gene expression control [53].
Studies have found that miRNAs function by altering the translation/transcription of oncogenes and/or tumor suppressor genes (TSGs) [54]. Oncogenic miRNAs (oncomiRs) tend to be overexpressed in cancer with the inverse relationship being true for tumor-suppressive miRNAs [49]. Disruption of miRNA regulation may wield significant influence over cancer progression, and an example of such is the oncomiR miR-21 which has been shown to drive tumor invasion, metastasis, and create epithelial mesenchymal transition (EMT) [55]. MiRNAs may also operate to antagonize or downregulate oncogenes (anti-oncomiR) and TSGs, further highlighting their role as modulators of carcinogenesis, cell apoptosis, invasive potential, and treatment resistance or efficacy [56]. The miRNA expression profile of specific tumors is the subject of ongoing investigation as a means of potential treatment avenue, early detection, or sub-classification.
The C19 miRNA cluster (C19MC) is the most tightly co-expressed miRNA set among all miRNAs that are expressed and a high expression of C19MC miRNAs also marked the basal-like TNBC subtype [57]. Similarly, the miR-17-92 cluster exhibits oncogenic behavior and has been observed to show substantial overexpression in human lung cancer and also contributes to the oncogenesis of acute myeloid leukemia with mixed-lineage leukemia (MLL) rearrangement [58]. The elevated amplification of the miR-23a locus, in conjunction with other miRNAs, has been found to stimulate gastric cancer tumor growth while simultaneously diminishing the expression of the metallothionein 2A (MT2A) gene, a known tumor progression suppressor [59,60]. In primary breast cancer cells, forced expression of miR-191 and miR-425 was shown to suppress the expression of dicer 1, ribonuclease III (DICER1) post-transcriptionally, consequently leading to the advancement of tumor progression and metastasis [61]. Repression of miR-191/425 was seen to also repress the carcinogenic activity of primary breast cancer cells, implicating miR-191/425 as oncomiRs [61]. MiRNAs such as miR-520h have been found to exhibit a dualistic role in cancer, alternating between tumor-suppressing and tumor-promoting functions [62]. In contrast, miR-15a and miR-16-1 consistently act as potent tumor suppressors, particularly in chronic lymphocytic leukemia, where they target the anti-apoptotic B-cell lymphoma 2 (BCL-2) mRNA [63]. Alternatively, miR-29 serves as a tumor suppressor in cholangiocarcinoma by targeting myeloid leukemia 1 (MCL-1) and in acute myeloid leukemia by orchestrating DNA methyltransferases, thereby restoring the expression of tumor suppressor genes through DNA hypomethylation [64].
Despite their ability to affect a wide range of target genes, individual miRNAs frequently exert their physiological effects by subtly altering the concentrations of crucial cellular proteins that serve as essential elements within complex cellular signaling pathways [65]. In hormone receptor positive breast cancer, the concentration of circulating miR-195 and let-7a was significantly elevated and was observed to discern breast cancer patients with a specificity of 100% for both and a sensitivity of 85.5% for miR-195 and 77.6% for let-7a [66]. In TNBC, hsa-miR-29b-3p has been found to elicit oncogenic activity through modulation of tumor necrosis factor receptor associated factor 3 (TRAF3), while miR-29a was found to downregulate the TSG suppressor of variegation 4–20 homolog 2 (SUV420H2) [47,67,68]. In summary, miRNAs operate to inflect the physiological, pathophysiological, and pathological processes within the body and are subject to continued research for the identification of their exact functional and pathological mechanism of action [69] (Table 1).
4. MicroRNAs in Basal-like Breast Cancer
4.1. MicroRNAs in the Diagnosis and Substratification of BLBC/TNBC Subtype
Due to the aggressive nature of basal-like breast cancers (BLBCs) and triple-negative breast cancers (TNBCs), early detection and diagnosis are imperative [83]. A screening assay or miRNA signature panel that differentiates BLBCs or TNBCs from the other subtypes or normal tissue may allow for quicker diagnosis and earlier intervention [56]. An example of such is the seven-miRNA panel developed by Kahraman et al. that was able to distinguish healthy women from patients with basal-like TNBC with a sensitivity and specificity of 84% and 74%, respectively [84]. Illustrating the miRNA profile of BLBCs/TNBCs can pave the way to understanding its meticulous disease process and lead to future studies validating the distinct role of miRNAs on tumorigenesis and tumor suppression [85,86].
One avenue to address this is through the analysis of miRNAs over- and under-expressed in TNBC and one such study was conducted by Hu J et al. [87]. The expression of seven miRNAs (hsa-miR-93, hsa-miR-25, hsa-miR-328, hsa-miR-16-2, hsa-miR-BART6-3p, hsa-miR-106b, and hsa-miR-7) were found to be upregulated by more than two-fold, while the expression of ten miRNAs (hsa-miR-145, hsa-miR-205, hsa-miR-27a, hsa-miR-195, hsvl-miR-H8, hsa-miR-376c, hsa-Let-7a, hsa-miR-585, hsa-miR-144, and hsa-miR-371-5p) were found to be downregulated [87]. A similar study by Radojicic J et al., using quantitative polymerase chain reaction (qPCR), observed significant overexpression of miR-21, miR-210, and miR-221 with reciprocal under expression of miR-10b, miR-122a, miR-145, and miR-205 in an analysis of 49 primary TNBC samples [88]. Ouyang M et al. analyzed the expression of 1513 miRNAs in TNBCs and found 41 miRNAs to be significantly dysregulated, 18 upregulated, and 23 downregulated, relative to normal adjacent tissue samples [89]. Du Y et al. performed a similar investigation of miRNAs differentially expressed in TNBC relative to normal, and found miR-105-5p, miR-210-3p, and miR-767-5p to be specifically upregulated with reciprocal under expression of miR-5683 [90]. Søkilde R et al.’s miRNA sequencing analysis of PAM50-classified BLBC samples found over one thousand miRNAs differentially over- and under-expressed in basal-like samples [19]. Similar studies on miRNAs up- and downregulated in BLBC/TNBC are featured in Table 2.
4.2. MicroRNAs Identified as OncomiRs or Tumor Suppressors in BLBC/TNBC
MiRNA expression signatures have been implicated as a means for the development of a potentially effective screening assay for the detection of breast cancer and its subtypes via a liquid blood/plasma-based biopsy sample [96]. In order for such a panel to be constructed, the identification of miRNAs that play a role in BLBC/TNBC and their specific functions and targets is imperative. Overexpression of miR-20a-5p was found to downregulate the expression of Bim and p21, in turn promoting metastasis and invasiveness of TNBC tumor cells in vitro, implicating miR-20a-5p as an oncomiR for TNBC [97]. In a miRnome analysis of miR-135b expression, it was found to be exclusively related to TNBC with basal-like phenotype and identified to possess carcinogenic activity [98]. Further target mapping of miR-135b implied its mechanism of tumorigenesis was through modulation of the wingless-related integration site (WNT), transforming growth factor-beta (TGFβ), and erythroblastic leukemia viral oncogene homologue (ERBB) pathways, all of which have been associated with breast cancer oncogenesis [98,99,100,101]. Hu et al. found miR-93 to be overexpressed in TNBCs relative to non-TNBCs and found MCF-7 cells transfected with miR-93 plasmid (pS-miR-93) displayed increased proliferation and invasion relative to the control [87]. The Warburg effect, a surging hallmark of cancer and a phenomenon in which malignant cells undergo increased aerobic glycolysis was found to be heavily correlated to miR-210-3p, miR-105-5p, and miR-767-5p in TNBC [90].
The stem-cell factors sex-determining region Y-related high-mobility group box 9 (SOX9) and aldehyde dehydrogenase 1 (ALDH1), the most activated factors in ductal carcinoma in situ (DCIS) stem-like cells, were identified to be objective targets for miR-140 [102]. The expression levels of miR-140 were seen to be downregulated in cancer stem-like cells relative to normal, and the reinstatement of miR-140 expression was shown to reduce in vivo tumor progression and exhibit tumor suppressive function in basal-like DCIS [102]. Telomeric repeat-binding factor 2 (TRF2), a known cancer advancement contributor, regulates telomere maintenance and has been identified to be downregulated by miR-182-3p in TNBC [103,104]. Impaired TNBC tumor expansion was seen in models treated with lipid nanoparticles containing miR-182-3p within [104]. Further miRNAs identified as oncomiRs and tumor suppressors in BLBC and/or TNBC summarized with their validated target and regulated expression can be found in Table 3.
4.3. Role of MicroRNAs as Prognostic Indicators in BLBC/TNBC
Prognostic indicators may be used to estimate the probability of crucial parameters such as overall survival (OS), recurrence-free survival (RFS), response to specific treatment regimens, and metastasis [123]. MiRNAs are a prime candidate for future enrichment and validation studies into their potential prognostic implications as their function is quite nuanced and not entirely defined [85,124]. Previous literature exploring the prognostic connotation of miRNA expression signatures has already shown great promise, and some such studies are showcased in Table 4.
5. MicroRNAs in Modulation of Drug Resistance in Breast Cancer
MiRNAs also play a pivotal role in modulating drug resistance in cancer cells and an example of such is miR-519c which was identified as a key factor in enhancing drug sensitivity in colon cancer cells through its regulation of ATP-binding cassette super-family G member 2 (ABCG2) [129]. Notably, miR-21 inhibition proves efficacious in reducing topotecan resistance in breast cancer [130]. The functional dynamics of miR-451 in doxorubicin-resistant breast cancer cells are nuanced, as it can either enhance sensitivity to anticancer drugs or provide protection [131]. Doxorubicin-resistant breast cancer tumors display suppressed or decreased expression of miR-451 and miR-200c and the transfection of miR-451 and miR-200c to MCF-7/doxorubicin-resistant cells elicits a heightened sensitivity to doxorubicin in said cells [131,132,133]. The heightened sensitivity underscores the critical role of correcting miRNA expression anomalies in the development of effective therapeutic approaches against drug-resistant cancer cells [131,132,133].
Beyond doxorubicin, other chemotherapeutics have also harnessed the potential of miRNAs to combat breast cancer [134]. Another such example is that of miR-328 demonstrating the capacity to enhance cellular sensitivity to mitoxantrone by downregulating the expression of ABCG2 proteins, resulting in an increased susceptibility of malignant tissue to mitoxantrone [134,135]. Furthermore, miR-132-3p enhances the susceptibility of breast cancer cells to etoposide-induced apoptosis, thereby increasing their responsiveness to the cytotoxic effects of etoposide [134,136]. Hu et al. highlighted the crucial role of miRNA-452 in influencing the responsiveness of breast cancer cells to docetaxel (DOC) where elevated miRNA-452 levels reduced sensitivity to DOC treatment, and conversely decreased miRNA-452 levels enhanced drug sensitivity [134,137]. Additionally, an analysis of miRNA and mRNA expression patterns identified a group of miRNAs, including let-7i, miR-346, miR-638, miR-181a, miR-191, miR-199b, miR-204, miR-211, miR-212, miR-216, miR-328, miR-373, miR-424, miR-768-3p, and miR-221/222, whose reduced levels significantly contributed to the development of antiestrogen resistance in breast cancer, presenting potential avenues for improved therapeutic strategies aimed at addressing this clinical challenge [134,138]. Identifying the role of miRNAs in determining iatrogenic complications of chemotherapy has significant clinical implications for patient satisfaction and prognosis [139,140]. Decreased concentration of circulating miR-195 and miR-21 was found to be statistically significant in forecasting the development of neutropenia and mucositis, respectively, in patients with primary breast cancer receiving neoadjuvant chemotherapy [140]. In the same study, elevated concentrations of miR-10b were found to correlate well to the development of anemia while raised circulating miR-145 expression accurately identified patients encountering nausea and vomiting [140].
6. MicroRNAs as Potential Treatment Options for Breast Cancer
Novel targeted treatment has already shown promising results and is well highlighted in a phase 3 trial of adjuvant pembrolizumab, an antiprogrammed death 1 checkpoint inhibitor, immunotherapy alongside neoadjuvant chemotherapy which showed increased rates of pathological complete response (pCR) and survival in early TNBCs [141]. Through functional mapping of miRNAs, therapeutics targeting miRNAs can be developed and instilled to improve the prognosis of BLBC and TNBC [142]. miR-10b exhibits therapeutic potential by targeting the homeobox D10 (HOXD10) gene, utilizing an Antagomir delivery system via the pcDNA5-CMV-d2eGFP vector [143]. MiR-19a-3p holds promise through its modulation of the fos-related antigen 1 (FRA-1) proto-oncogene, employing nanoparticles as the delivery platform within the category of miRNA mimics [144]. Furthermore, miRNAs such as miR-27a and miR-451 have demonstrated therapeutic relevance by targeting the multidrug resistance protein 1 (MDR1)/P-glycoprotein gene, facilitated through lipid-based delivery systems, categorized as miRNA mimics/antagomirs [145]. MiR-34a, on the other hand, exerts its influence by targeting multiple genes, including E2F transcription factor 3 (E2F3), cluster of differentiation 44 (CD44), and silent mating-type information regulation 2 homolog 1 (SIRT1), by employing the T-VISA system (plasmid) within the miRNA mimics category [146]. MiR-145 showcases its therapeutic potential through the regulation of various genes, including fascin-1 (FSCN1), C-MYC, small mothers against decapentaplegic homolog 2/3 (SMAD2/3), insulin-like growth factor 1 receptor (IGF-1R), and tumor protein 53 (TP53), utilizing adenoviral and lentiviral vectors as the transfection systems in the miRNA mimics category [147,148]. MiR-326 takes aim at the multidrug resistance protein 1 (MRP-1)/ATP-binding cassette subfamily member 1 (ABCC1) gene, employing the pGL2-control vector for transfection [149]. Additionally, miR-298 and miR-1253 demonstrate their therapeutic relevance by targeting the MDR1/P-glycoprotein (P-gp) gene, making use of the lipofectamine vector in the miRNA mimics category [150]. Even though it is a promising field for the ongoing battle against cancer, it is not without its limitations which include challenges with reproducibility, delivery of therapeutics, knowledge gaps, and cost efficiency [151,152,153,154].
7. Limitations and Challenges of MicroRNA-Based Research
The implication of miRNAs in cancer is a relatively recent concept, evidenced by the fact that the first study showing dysregulation of miRNAs in cancer was published as recently as 2002, which suggested the role miR-15 and miR-16 in chronic lymphocytic leukemia [155]. Validation studies have shown inconsistent results, which may be a consequence of variation in methodology and lack of a standardized approach [151,153]. Disparities in the miRNA quantification method used such as hybridization, amplification, and sequencing can directly contribute to sub-satisfactory reproducibility [153]. Discrepancy of sample extraction and preparation, miRNA retrieval, miRNA quantitative and qualitative protocols, and normalization of post-analytical data all may give rise to conflicting findings [151]. Clinical implementation of miRNA-based medications faces significant obstacles including identifying viable mediums for administration, impaired transfection, adverse side effects, and bioavailability/biodistribution, as well as navigating patient-specific discrepancies in metabolism [155]. These challenges are exemplified due to the apparent inadequate understanding of the complete functionality for any given miRNA, and the in vivo antagonization or introduction of one or more miRNAs possesses the capacity to generate considerable drug-associated toxicity [154,155]. This is well evidenced from previous clinical trials involving miRNA interference drugs; an example of such is the miR-34 mimic, MRX34, which showed substantial efficacy against primary liver cancer, melanoma, renal cell carcinoma, and non-small cell lung cancer but induced five significant immune-related adverse events leading to trial termination [154]. Cost optimization without compromising integrity and quality also poses as a hinderance to miRNA-based research [154]. Investigations to surmount the above-mentioned impediments, to facilitate the clinical translation of miRNAs, are ongoing and promising [156].
8. Conclusions
Breast cancer is an ever-prevalent pathology that persistently claims lives by the thousands yearly. Researchers have investigated the complex, heterogenous disease process of breast cancer to taxonomically segregate tumors based on their proteomic, genomic, phenotypic, transcriptomic, and epigenetic profile. Of all the resultant variants, BLBC and TNBC carry an unfortunate prognosis and the lack of targeted therapy continues to pose a challenge for management. Early diagnosis is imperative to reducing the disease burden of BLBC and the intricate dual functionality of miRNAs makes them a prime candidate for the production of an early detection model. Circulating miRNA expression signatures have been implicated as a means for the development of a potentially effective screening assay to detect BLBC via a liquid blood/plasma-based biopsy sample. Individualized and personalized targeted therapy for BLBC may considerably reduce the associated morbidity and mortality, and miRNAs stand as a potential means to achieve such. Drugs capable of effectively and safely silencing oncomiRs and/or potentiating tumor suppressive miRNAs associated with BLBC can substantially alter the present BLBC therapeutic landscape. The impact of miRNAs on BLBC tumorigenesis, tumor suppression, malignant transformation, and drug modulation, as highlighted in this review, is still subject to continued ongoing investigation. In conclusion, BLBC is an aggressive and complicated condition with no targeted therapy currently available and carries a significant burden to patients, but through previous and ongoing miRNA-based research, there is still hope.
Author Contributions
M.T. (writing—original draft, conceptualization, data curation, validation, visualization), V.R. (writing—review and editing, conceptualization, data curation, validation, supervision) and M.J.K. (supervision, visualization, grant acquisition, writing—review and editing). All authors have read and agreed to the published version of the manuscript.
Funding
The research in M.J. Kerin’s laboratory at the University of Galway, Ireland, is supported by the National Breast Cancer Research Institute [NBCRI] and Science Foundation Ireland [SFI] Strategic Partnership Programme, under Grant number [18/SPP/3522]. M.T. received a HRB Summer student scholarship 2023 (Grant Reference SS-2023-093). V.R. received postdoctoral fellowship support from the National Breast Cancer Research Institute [NBCRI] and Precision Oncology Ireland under Grant number [18/SPP/3522].
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef]
- Testa, U.; Castelli, G.; Pelosi, E. Breast Cancer: A Molecularly Heterogenous Disease Needing Subtype-Specific Treatments. Med. Sci. 2020, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Kong, D.; Liu, J.; Zhan, L.; Luo, L.; Zheng, W.; Zheng, Q.; Chen, C.; Sun, S. Breast cancer heterogeneity and its implication in personalized precision therapy. Exp. Hematol. Oncol. 2023, 12, 3. [Google Scholar] [CrossRef]
- Bouchal, P.; Schubert, O.T.; Faktor, J.; Čápková, L.; Imrichová, H.; Zoufalová, K.; Páralová, V.; Hrstka, R.; Liu, Y.; Ebhardt, H.A.; et al. Breast Cancer Classification Based on Proteotypes Obtained by SWATH Mass Spectrometry. Cell Rep. 2019, 28, 832–843.e7. [Google Scholar] [CrossRef] [PubMed]
- Walter, V.; Fischer, C.; Deutsch, T.M.; Ersing, C.; Nees, J.; Schütz, F.; Fremd, C.; Grischke, E.; Sinn, H.; Brucker, S.Y.; et al. Estrogen, Progesterone, and Human Epidermal Growth Factor Receptor 2 Discordance between Primary and Metastatic Breast Cancer. Breast Cancer Res. Treat. 2020, 183, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Orrantia-Borunda, E.; Anchondo-Nuñez, P.; Acuña-Aguilar, L.E.; Gómez-Valles, F.O.; Ramírez-Valdespino, C.A. Subtypes of Breast Cancer; Exon Publications: Brisbane, Australia, 2022; pp. 31–42. [Google Scholar] [CrossRef]
- Zagami, P.; Carey, L.A. Triple Negative Breast Cancer: Pitfalls and Progress. NPJ Breast Cancer 2022, 8, 95. [Google Scholar] [CrossRef]
- Dass, S.A.; Tan, K.L.; Rajan, R.S.; Mokhtar, N.; Adzmi, E.R.M.; Rahman, W.F.W.A.; Din, T.A.D.A.-A.T.; Balakrishnan, V. Triple Negative Breast Cancer: A Review of Present and Future Diagnostic Modalities. Medicina 2021, 57, 62. [Google Scholar] [CrossRef]
- Fumagalli, C.; Barberis, M. Breast Cancer Heterogeneity. Diagnostics 2021, 11, 1555. [Google Scholar] [CrossRef]
- Zadeh, A.H.; Alsabi, Q.; Ramírez-Vick, J.E.; Nosoudi, N. Characterizing Basal-like Triple Negative Breast Cancer Using Gene Expression Analysis: A Data Mining Approach. Expert Syst. Appl. 2020, 148, 113253. [Google Scholar] [CrossRef]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; Van De Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular Portraits of Human Breast Tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Alluri, P.; Newman, L.A. Basal-Like and Triple-Negative Breast Cancers. Surg. Oncol. Clin. N. Am. 2014, 23, 567–577. [Google Scholar] [CrossRef]
- Banerjee, S.; Reis-Filho, J.S.; Ashley, S.; Steele, D.; Ashworth, A.; Lakhani, S.R.; Smith, I. Basal-like Breast Carcinomas: Clinical Outcome and Response to Chemotherapy. J. Clin. Pathol. 2006, 59, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.R.; Hwang, C.Y.; Lee, J.; Cho, K.-H. Network Analysis Identifies Regulators of Basal-Like Breast Cancer Reprogramming and Endocrine Therapy Vulnerability. Cancer Res. 2022, 82, 320–333. [Google Scholar] [CrossRef] [PubMed]
- Wang, L. Early Diagnosis of Breast Cancer. Sensors 2017, 17, 1572. [Google Scholar] [CrossRef]
- Parker, J.S.; Mullins, M.E.; Cheang, M.C.U.; Leung, S.; David, V.; Vickery, T.L.; Davies, S.R.; Fauron, C.; He, X.; Hu, Z.; et al. Supervised Risk Predictor of Breast Cancer Based on Intrinsic Subtypes. J. Clin. Oncol. 2009, 27, 1160–1167. [Google Scholar] [CrossRef]
- Krug, K.; Jaehnig, E.J.; Satpathy, S.; Blumenberg, L.M.; Karpova, A.; Anurag, M.; Miles, G.; Mertins, P.; Geffen, Y.; Tang, L.; et al. Proteogenomic Landscape of Breast Cancer Tumorigenesis and Targeted Therapy. Cell 2020, 183, 1436–1456.e31. [Google Scholar] [CrossRef]
- Søkilde, R.; Persson, H.; Ehinger, A.; Pirona, A.C.; Fernö, M.; Hegardt, C.; Larsson, C.; Loman, N.; Malmberg, M.; Rydén, L.; et al. Refinement of Breast Cancer Molecular Classification by MiRNA Expression Profiles. BMC Genom. 2019, 20, 503. [Google Scholar] [CrossRef] [PubMed]
- Mkabaah, L.B.; Davey, M.G.; Lennon, J.C.; Bouz, G.; Miller, N.; Kerin, M.J. Assessing the Role of MicroRNAs in Predicting Breast Cancer Recurrence—A Systematic Review. Int. J. Mol. Sci. 2023, 24, 7115. [Google Scholar] [CrossRef]
- Syed, R.A.; Davey, M.G.; Richard, V.; Miller, N.; Kerin, M.J. Biological Implications of MicroRNAs as Regulators and Biomarkers of Therapeutic Toxicities in Breast Cancer. Int. J. Mol. Sci. 2023, 24, 12694. [Google Scholar] [CrossRef]
- Richard, V.; Davey, M.G.; Annuk, H.; Miller, N.; Dwyer, R.M.; Lowery, A.; Kerin, M.J. MicroRNAs in Molecular Classification and Pathogenesis of Breast Tumors. Cancers 2021, 13, 5332. [Google Scholar] [CrossRef] [PubMed]
- Annese, T.; Tamma, R.; De Giorgis, M.; Ribatti, D. MicroRNAs Biogenesis, Functions and Role in Tumor Angiogenesis. Front. Oncol. 2020, 10, 581007. [Google Scholar] [CrossRef]
- Bhat, S.A.; Majid, S.; Hassan, T. MicroRNAs and Its Emerging Role as Breast Cancer Diagnostic Marker- A Review. Adv. Biomark. Sci. Technol. 2019, 1, 1–8. [Google Scholar] [CrossRef]
- Hamam, R.; Hamam, D.; Alsaleh, K.; Kassem, M.; Zaher, W.; Alfayez, M.; Aldahmash, A.; Alajez, N.M. Circulating MicroRNAs in Breast Cancer: Novel Diagnostic and Prognostic Biomarkers. Cell Death Dis. 2017, 8, e3045. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, T.O.; Hsu, F.D.; Jensen, K.C.; Cheang, M.C.U.; Karaca, G.; Hu, Z.; Hernandez-Boussard, T.; Livasy, C.; Cowan, D.; Dressler, L.G.; et al. Immunohistochemical and Clinical Characterization of the Basal-Like Subtype of Invasive Breast Carcinoma. Clin. Cancer Res. 2004, 10, 5367–5374. [Google Scholar] [CrossRef] [PubMed]
- Cheang, M.C.U.; David, V.; Bajdik, C.; Leung, S.; McKinney, S.; Chia, S.; Perou, C.M.; Nielsen, T.O. Basal-Like Breast Cancer Defined by Five Biomarkers Has Superior Prognostic Value than Triple-Negative Phenotype. Clin. Cancer Res. 2008, 14, 1368–1376. [Google Scholar] [CrossRef] [PubMed]
- Livasy, C.; Karaca, G.; Nanda, R.; Tretiakova, M.; Olopade, O.I.; Moore, D.T.; Perou, C.M. Phenotypic Evaluation of the Basal-like Subtype of Invasive Breast Carcinoma. Mod. Pathol. 2006, 19, 264–271. [Google Scholar] [CrossRef]
- Herschkowitz, J.I.; Simin, K.; Weigman, V.; Mikaelian, I.; Usary, J.; Hu, Z.; Rasmussen, K.; Jones, L.P.; Assefnia, S.; Chandrasekharan, S.; et al. Identification of Conserved Gene Expression Features between Murine Mammary Carcinoma Models and Human Breast Tumors. Genome Biol. 2007, 8, R76. [Google Scholar] [CrossRef]
- Prat, A.; Parker, J.S.; Karginova, O.; Fan, C.; Livasy, C.; Herschkowitz, J.I.; He, X.; Perou, C.M. Phenotypic and Molecular Characterization of the Claudin-Low Intrinsic Subtype of Breast Cancer. Breast Cancer Res. 2010, 12, R68. [Google Scholar] [CrossRef]
- Kensler, K.H.; Sankar, V.; Wang, J.; Zhang, X.; Rubadue, C.A.; Baker, G.M.; Parker, J.S.; Hoadley, K.A.; Stancu, A.L.; Pyle, M.E.; et al. PAM50 Molecular Intrinsic Subtypes in the Nurses’ Health Study Cohorts. Cancer Epidemiol. Biomark. Prev. 2019, 28, 798–806. [Google Scholar] [CrossRef]
- Hurson, A.N.; Hamilton, A.M.; Olsson, L.T.; Kirk, E.L.; Sherman, M.E.; Calhoun, B.C.; Geradts, J.; Troester, M.A. Reproducibility and Intratumoral Heterogeneity of the PAM50 Breast Cancer Assay. Breast Cancer Res. Treat. 2023, 199, 147–154. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Network: Koboldt, D.C.; Fulton, R.S.; McLellan, M.D.; Schmidt, H.; Kalicki-Veizer, J.; McMichael, J.F.; Fulton, L.L.; Dooling, D.J.; Li, D.; Mardis, E.R.; et al. Comprehensive Molecular Portraits of Human Breast Tumours. Nature 2012, 490, 61–70. [Google Scholar] [CrossRef]
- Mathews, J.; Nadeem, S.; Levine, A.J.; Pouryahya, M.; Deasy, J.O.; Tannenbaum, A. Robust and Interpretable PAM50 Reclassification Exhibits Survival Advantage for Myoepithelial and Immune Phenotypes. NPJ Breast Cancer 2019, 5, 30. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.R.; Jackson, H.W.; Zanotelli, V.R.T.; Danenberg, E.; Fischer, J.; Bardwell, H.; Provenzano, E.; Rueda, O.M.; Chin, S.-F.; Aparicio, S.; et al. Imaging Mass Cytometry and Multiplatform Genomics Define the Phenogenomic Landscape of Breast Cancer. Nat. Cancer 2020, 1, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Zou, R.; Loke, S.Y.; Tan, V.K.; Quek, S.T.; Jagmohan, P.; Tang, Y.C.; Madhukumar, P.; Tan, B.K.; Yong, W.S.; Sim, Y.; et al. Development of a MicroRNA Panel for Classification of Abnormal Mammograms for Breast Cancer. Cancers 2021, 13, 2130. [Google Scholar] [CrossRef]
- Saal, L.H.; Vallon-Christersson, J.; Häkkinen, J.; Hegardt, C.; Grabau, D.; Winter, C.; Brueffer, C.; Tang, M.-H.E.; Reuterswärd, C.; Schulz, R.; et al. The Sweden Cancerome Analysis Network—Breast (SCAN-B) Initiative: A Large-Scale Multicenter Infrastructure towards Implementation of Breast Cancer Genomic Analyses in the Clinical Routine. Genome Med. 2015, 7, 20. [Google Scholar] [CrossRef]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef]
- De Rie, D.; Abugessaisa, I.; Alam, T.; Arner, P.; Arner, P.; Ashoor, H.; Åström, G.; Babina, M.; Bertin, N.; Burroughs, A.M.; et al. An Integrated Expression Atlas of MiRNAs and Their Promoters in Human and Mouse. Nat. Biotechnol. 2017, 35, 872–878. [Google Scholar] [CrossRef]
- O’Brien, J.A.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Lou, W.; Liu, J.; Gao, Y.; Zhong, G.; Chen, D.; Shen, J.; Bao, C.; Xu, L.; Pan, J.; Cheng, J.; et al. MicroRNAs in Cancer Metastasis and Angiogenesis. Oncotarget 2017, 8, 115787–115802. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Lee, Y.; Yeom, K.-H.; Kim, Y.; Jin, H.; Kim, V.N. The Drosha-DGCR8 Complex in Primary MicroRNA Processing. Genes Dev. 2004, 18, 3016–3027. [Google Scholar] [CrossRef]
- Gregory, R.I.; Shiekhattar, R. MicroRNA Biogenesis and Cancer. Cancer Res. 2005, 65, 3509–3512. [Google Scholar] [CrossRef] [PubMed]
- Yi, R.; Qin, Y.; Macara, I.G.; Cullen, B.R. Exportin-5 Mediates the Nuclear Export of Pre-MicroRNAs and Short Hairpin RNAs. Genes Dev. 2003, 17, 3011–3016. [Google Scholar] [CrossRef]
- MacFarlane, L.A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef]
- Huntzinger, E.; Izaurralde, E. Gene Silencing by MicroRNAs: Contributions of Translational Repression and MRNA Decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Richard, V.; Nair, M.G.; Jaikumar, V.S.; Jones, S.E.; Prabhu, J.S.; Kerin, M.J. Cell State Transitions and Phenotypic Heterogeneity in Luminal Breast Cancer Implicating MicroRNAs as Potential Regulators. Int. J. Mol. Sci. 2023, 24, 3497. [Google Scholar] [CrossRef]
- Dharap, A.; Pokrzywa, C.; Murali, S.; Pandi, G.; Vemuganti, R. MicroRNA MIR-324-3P Induces Promoter-Mediated Expression of RELA Gene. PLoS ONE 2013, 8, e79467. [Google Scholar] [CrossRef] [PubMed]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OnCOMIR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef]
- Huang, Y.; Shen, X.J.; Zou, Q.; Wang, S.P.; Tang, S.M.; Zhang, G.Z. Biological Functions of MicroRNAs: A Review. J. Physiol. Biochem. 2010, 67, 129–139. [Google Scholar] [CrossRef]
- Makeyev, E.V.; Maniatis, T. Multilevel Regulation of Gene Expression by MicroRNAs. Science 2008, 319, 1789–1790. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Aziz, A.A.; Rajasegaran, Y.; Zi, K.K.; Rosli, A.A.; Yik, M.Y.; Yusoff, N.M.; Heidenreich, O.; Moses, E.J. Elucidating MIRNA Function in Cancer Biology via the Molecular Genetics’ Toolbox. Biomedicines 2022, 10, 915. [Google Scholar] [CrossRef]
- Otmani, K.; Lewalle, P. Tumor Suppressor MIRNA in Cancer Cells and the Tumor Microenvironment: Mechanism of Deregulation and Clinical Implications. Front. Oncol. 2021, 11, 708765. [Google Scholar] [CrossRef] [PubMed]
- Debnath, T.; Nath, N.C.D.; Kim, M.J.; Lee, K. Role of Phytochemicals in the Modulation of MiRNA Expression in Cancer. Food Funct. 2017, 8, 3432–3442. [Google Scholar] [CrossRef]
- Angius, A.; Cossu-Rocca, P.; Arru, C.; Muroni, M.R.; Rallo, V.; Carru, C.; Uva, P.; Pira, G.; Orrù, S.; De Miglio, M.R. Modulatory Role of MicroRNAs in Triple Negative Breast Cancer with Basal-Like Phenotype. Cancers 2020, 12, 3298. [Google Scholar] [CrossRef] [PubMed]
- Jinesh, G.G.; Flores, E.R.; Brohl, A.S. Chromosome 19 MiRNA Cluster and CEBPB Expression Specifically Mark and Potentially Drive Triple Negative Breast Cancers. PLoS ONE 2018, 13, e0206008. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Li, Z.; Chen, P.; He, C.; Cao, D.; Elkahloun, A.G.; Lü, J.; Pelloso, L.A.F.; Wunderlich, M.; Huang, H.; et al. Aberrant Overexpression and Function of the MiR-17-92 Cluster in MLL-Rearranged Acute Leukemia. Proc. Natl. Acad. Sci. USA 2010, 107, 3710–3715. [Google Scholar] [CrossRef]
- An, J.; Pan, Y.; Yan, Z.; Li, W.; Cui, J.; Yuan, J.; Tian, L.; Xing, R.; Lü, Y. MiR-23a in Amplified 19p13.13 Loci Targets Metallothionein 2A and Promotes Growth in Gastric Cancer Cells. J. Cell. Biochem. 2013, 114, 2160–2169. [Google Scholar] [CrossRef]
- Shimizu, M.; Koma, Y.; Sakamoto, H.; Tsukamoto, S.; Kitamura, Y.; Urakami, S.; Tanigawa, K.; Kodama, T.; Higashino, N.; Nishio, M.; et al. Metallothionein 2A Expression in Cancer-Associated Fibroblasts and Cancer Cells Promotes Esophageal Squamous Cell Carcinoma Progression. Cancers 2021, 13, 4552. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, M.; Chong, Q.-Y.; Zhang, W.; Qian, P.; Zhang, P.; Qian, W.; Zhang, M.; Lobie, P.E.; Zhu, T. Amplification of Hsa-MiR-191/425 Locus Promotes Breast Cancer Proliferation and Metastasis by Targeting DICER1. Carcinogenesis 2018, 39, 1506–1516. [Google Scholar] [CrossRef]
- Ting, S.; Chen, P.S.; Johansson, G.; Chu, C.-Y.; Wang, M.Y.; Jeng, Y.; Yu, S.-L.; Chen, J.; Chang, K.; Jee, S.; et al. MicroRNA-519C Suppresses Hypoxia-Inducible Factor-1A Expression and Tumor Angiogenesis. Cancer Res. 2010, 70, 2675–2685. [Google Scholar] [CrossRef]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M.; et al. MiR-15 and MiR-16 Induce Apoptosis by Targeting BCL2. Proc. Natl. Acad. Sci. USA 2005, 102, 13944–13949. [Google Scholar] [CrossRef]
- Garzon, R.; Liu, S.; Fabbri, M.; Liu, Z.; Heaphy, C.E.A.; Callegari, E.; Schwind, S.; Pang, J.; Yu, J.; Muthusamy, N.; et al. MicroRNA-29b Induces Global DNA Hypomethylation and Tumor Suppressor Gene Reexpression in Acute Myeloid Leukemia by Targeting Directly DNMT3A and 3B and Indirectly DNMT1. Blood 2009, 113, 6411–6418. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Rajewsky, K. MicroRNA Control in the Immune System: Basic Principles. Cell 2009, 136, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Heneghan, H.; Miller, N.; Lowery, A.; Sweeney, K.; Newell, J.; Kerin, M.J. Circulating MicroRNAs as Novel Minimally Invasive Biomarkers for Breast Cancer. Ann. Surg. 2010, 251, 499–505. [Google Scholar] [CrossRef]
- Bao, Z.; Shetti, D.; Fan, C.; Wei, K. MiR-29b-3p Promotes Progression of MDA-MB-231 Triple-Negative Breast Cancer Cells through Downregulating TRAF3. Biol. Res. 2019, 52, 38. [Google Scholar] [CrossRef]
- Wu, Y.; Shi, W.; Tang, T.; Wang, Y.; Yin, X.; Chen, Y.; Zhang, Y.; Xing, Y.; Shen, Y.; Xia, T.; et al. MiR-29a Contributes to Breast Cancer Cells Epithelial–Mesenchymal Transition, Migration, and Invasion via down-Regulating Histone H4K20 Trimethylation through Directly Targeting SUV420H2. Cell Death Dis. 2019, 10, 176. [Google Scholar] [CrossRef]
- Bhaskaran, M.; Mohan, M. MicroRNAs: History, Biogenesis, and Their Evolving Role in Animal Development and Disease. Vet. Pathol. 2013, 51, 759–774. [Google Scholar] [CrossRef]
- Ma, L.; Teruya-Feldstein, J.; Weinberg, R.A. Tumour Invasion and Metastasis Initiated by MicroRNA-10b in Breast Cancer. Nature 2007, 449, 682–688. [Google Scholar] [CrossRef]
- Weidle, U.H.; Dickopf, S.; Hintermair, C.; Kollmorgen, G.; Birzele, F.; Brinkmann, U. The Role of Micro RNAs in Breast Cancer Metastasis: Preclinical Validation and Potential Therapeutic Targets. Cancer Genom. Proteom. 2018, 15, 17–39. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, X.; Huang, Z.; Wang, J.; Zhu, W.; Shu, Y.; Liu, P. Prognostic Value of MIR-21 in Various Cancers: An Updating Meta-Analysis. PLoS ONE 2014, 9, e102413. [Google Scholar] [CrossRef]
- Shah, N.R.; Chen, H. MicroRNAs in Pathogenesis of Breast Cancer: Implications in Diagnosis and Treatment. World J. Clin. Oncol. 2014, 5, 48. [Google Scholar] [CrossRef]
- Sharifi, M.; Moridnia, A. Apoptosis-Inducing and Antiproliferative Effect by Inhibition of MiR-182-5p through the Regulation of CASP9 Expression in Human Breast Cancer. Cancer Gene Ther. 2017, 24, 75–82. [Google Scholar] [CrossRef]
- Zehentmayr, F.; Hauser-Kronberger, C.; Zellinger, B.; Hlubek, F.; Schuster, C.; Bodenhofer, U.; Fastner, G.; Deutschmann, H.; Steininger, P.; Reitsamer, R.; et al. Hsa-MiR-375 Is a Predictor of Local Control in Early Stage Breast Cancer. Clin. Epigenetics 2016, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Raval, A.; Joshi, J.; Shah, F.D. Significance of MetastamiR-10b in Breast Cancer Therapeutics. J. Egypt. Natl. Cancer Inst. 2022, 34, 19. [Google Scholar] [CrossRef]
- Feliciano, A.; Castellví, J.; Castro, A.A.; Leal, J.A.; Romagosa, C.; Hernández-Losa, J.; Peg, V.; Fabra, À.; Vidal, F.; Kondoh, H.; et al. MIR-125B Acts as a Tumor Suppressor in Breast Tumorigenesis via Its Novel Direct Targets ENPEP, CK2-A, CCNJ, and MEGF9. PLoS ONE 2013, 8, e76247. [Google Scholar] [CrossRef]
- Xu, W.-X.; Zhen, L.; Deng, F.; Wang, D.-D.; Li, X.-W.; Tian, T.; Zhang, J.; Tang, J.-H. MiR-145: A Potential Biomarker of Cancer Migration and Invasion. Am. J. Transl. Res. 2019, 11, 6739. [Google Scholar]
- Kuang, W.; Deng, Q.; Deng, C.; Ws, L.; Yg, Z.; Shu, S.; Zhou, M. MiRNA Regulates OCT4 Expression in Breast Cancer Cells. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1351–1357. [Google Scholar] [CrossRef] [PubMed]
- Klicka, K.; Grzywa, T.M.; Mielniczuk, A.; Klinke, A.; Włodarski, P. The Role of MiR-200 Family in the Regulation of Hallmarks of Cancer. Front. Oncol. 2022, 12, 965231. [Google Scholar] [CrossRef] [PubMed]
- Fontana, A.; Barbano, R.; Dama, E.; Pasculli, B.; Rendina, M.; Morritti, M.; Melocchi, V.; Castelvetere, M.; Valori, V.M.; Ravaioli, S.; et al. Combined Analysis of MiR-200 Family and Its Significance for Breast Cancer. Sci. Rep. 2021, 11, 2980. [Google Scholar] [CrossRef]
- Plantamura, I.; Cataldo, A.; Cosentino, G.; Iorio, M.V. MIR-205 in Breast Cancer: State of the Art. Int. J. Mol. Sci. 2020, 22, 27. [Google Scholar] [CrossRef] [PubMed]
- Afifi, N.; Barrero, C. Understanding Breast Cancer Aggressiveness and Its Implications in Diagnosis and Treatment. J. Clin. Med. 2023, 12, 1375. [Google Scholar] [CrossRef] [PubMed]
- Kahraman, M.; Röske, A.; Laufer, T.; Fehlmann, T.; Backes, C.; Kern, F.; Kohlhaas, J.; Schrörs, H.; Saiz, A.; Zabler, C.; et al. MicroRNA in Diagnosis and Therapy Monitoring of Early-Stage Triple-Negative Breast Cancer. Sci. Rep. 2018, 8, 11584. [Google Scholar] [CrossRef]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.E.; Creţoiu, D.; Suciu, N.; Crețoiu, S.M.; Voinea, S. MIRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef]
- Loh, H.-Y.; Norman, B.P.; Lai, K.-S.; Rahman, N.M.A.N.A.; Alitheen, N.B.M.; Osman, M. The Regulatory Role of MicroRNAs in Breast Cancer. Int. J. Mol. Sci. 2019, 20, 4940. [Google Scholar] [CrossRef]
- Hu, J.; Xu, J.; Wu, Y.; Chen, Q.; Zheng, W.; Lu, X.; Zhou, C.H.; Jiao, D. Identification of MicroRNA-93 as a Functional Dysregulated MiRNA in Triple-Negative Breast Cancer. Tumor Biol. 2014, 36, 251–258. [Google Scholar] [CrossRef]
- Radojicic, J.; Zaravinos, A.; Vrekoussis, T.; Kafousi, M.; Spandidos, D.A.; Stathopoulos, E.N. MicroRNA Expression Analysis in Triple-Negative (ER, PR and Her2/Neu) Breast Cancer. Cell Cycle 2011, 10, 507–517. [Google Scholar] [CrossRef]
- Ouyang, M.; Li, Y.; Ye, S.; Ma, J.; Lu, L.; Lv, W.; Chang, G.; Li, X.; Li, Q.; Wang, S.; et al. MicroRNA Profiling Implies New Markers of Chemoresistance of Triple-Negative Breast Cancer. PLoS ONE 2014, 9, e96228. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Wei, N.; Ma, R.; Jiang, S.; Song, D.E. A MiR-210-3p Regulon That Controls the Warburg Effect by Modulating HIF-1α and P53 Activity in Triple-Negative Breast Cancer. Cell Death Dis. 2020, 11, 731. [Google Scholar] [CrossRef]
- Shaaban, N.Z.; Ibrahim, N.K.; Saada, H.N.; El-Rashidy, F.H.; Shaaban, H.M.; Elbakary, N.M.; Kodous, A.S. The Implication of MicroRNAs as Non-Invasive Biomarkers in 179 Egyptian Breast Cancer Female Patients. Oncol. Res. 2022, 30, 269–276. [Google Scholar] [CrossRef]
- Turashvili, G.; Lightbody, E.D.; Tyryshkin, K.; Sengupta, S.; Elliott, B.E.; Madarnas, Y.; Ghaffari, A.; Day, A.G.; Nicol, C.J.B. Novel Prognostic and Predictive MicroRNA Targets for Triple-negative Breast Cancer. FASEB J. 2018, 32, 5937–5954. [Google Scholar] [CrossRef]
- M’hamed, I.F.; Privat, M.; Trimèche, M.; Penault-Llorca, F.; Bignon, Y.; Kénani, A. MiR-10b, MiR-26a, MiR-146a And MiR-153 Expression in Triple Negative Vs Non Triple Negative Breast Cancer: Potential Biomarkers. Pathol. Oncol. Res. 2017, 23, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Aydoğdu, E.; Katchy, A.; Tsouko, E.; Lin, C.-Y.; Haldosén, L.-A.; Helguero, L.A.; Williams, C. MicroRNA-Regulated Gene Networks during Mammary Cell Differentiation Are Associated with Breast Cancer. Carcinogenesis 2012, 33, 1502–1511. [Google Scholar] [CrossRef]
- Lowery, A.; Miller, N.; Devaney, A.; McNeill, R.E.; Davoren, P.A.; Lemetre, C.; Beneš, V.; Schmidt, S.; Blake, J.; Ball, G.; et al. MicroRNA Signatures Predict Oestrogen Receptor, Progesterone Receptor and HER2/Neureceptor Status in Breast Cancer. Breast Cancer Res. 2009, 11, R27. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Papukashvili, D.; Wang, Z.; Liu, Y.; Chen, X.; Lǐ, J.; Li, Z.; Hu, L.; Li, Z.; Rcheulishvili, N.; et al. Potential Utility of MiRNAs for Liquid Biopsy in Breast Cancer. Front. Oncol. 2022, 12, 940314. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Han, G.; Liu, Y.; Jiang, H.; He, Q. MiRNA-20a-5p Promotes the Growth of Triple-Negative Breast Cancer Cells through Targeting RUNX3. Biomed. Pharmacother. 2018, 103, 1482–1489. [Google Scholar] [CrossRef] [PubMed]
- Uva, P.; Cossu-Rocca, P.; Loi, F.; Pira, G.; Murgia, L.; Orrù, S.; Floris, M.; Muroni, M.R.; Sanges, F.; Carru, C.; et al. MiRNA-135b Contributes to Triple Negative Breast Cancer Molecular Heterogeneity: Different Expression Profile in Basal-like Versus Non-Basal-like Phenotypes. Int. J. Med. Sci. 2018, 15, 536–548. [Google Scholar] [CrossRef]
- Jamdade, V.S.; Sethi, N.; Mundhe, N.; Kumar, P.; Lahkar, M.; Sinha, N. Therapeutic Targets of Triple-Negative Breast Cancer: A Review. Br. J. Pharmacol. 2015, 172, 4228–4237. [Google Scholar] [CrossRef]
- Ding, M.; Su, K.; Cui, G.; Yang, W.; Chen, L.; Yang, M.; Liu, Y.; Dai, D. Association between Transforming Growth Factor-Β1 Expression and the Clinical Features of Triple Negative Breast Cancer. Oncol. Lett. 2016, 11, 4040–4044. [Google Scholar] [CrossRef]
- Rangel, M.C.; Bertolette, D.C.; Castro, N.P.; Klauzińska, M.; Cuttitta, F.; Salomon, D. Developmental Signaling Pathways Regulating Mammary Stem Cells and Contributing to the Etiology of Triple-Negative Breast Cancer. Breast Cancer Res. Treat. 2016, 156, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yao, Y.; Eades, G.; Liu, Z.; Zhang, Y.; Zhou, Q. Downregulation of MiR-140 Promotes Cancer Stem Cell Formation in Basal-like Early Stage Breast Cancer. Oncogene 2013, 33, 2589–2600. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, O.H.; Xin, H.; Chen, L.; Qin, J.; Chae, H.K.; Lin, S.Y.; Safari, A.; Liu, D.; Songyang, Z. TRF2 Functions as a Protein Hub and Regulates Telomere Maintenance by Recognizing Specific Peptide Motifs. Nat. Struct. Mol. Biol. 2009, 16, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Dinami, R.; Pompili, L.; Petti, E.; Porru, M.; D’Angelo, C.; Di Vito, S.; Rizzo, A.; Campani, V.; De Rosa, G.; Bruna, A.; et al. MiR -182-3p Targets TRF2 and Impairs Tumor Growth of Triple-negative Breast Cancer. EMBO Mol. Med. 2022, 15, e16033. [Google Scholar] [CrossRef]
- Rezaei, F.; Forouzesh, F.; Abbasvandi, F. Up-Regulation of Mir-105-5p Promotes The Development of Breast Cancer By Targeting AKT1 / GRB2 Genes In Patients With Breast Cancer. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Hua, K.; Jin, J.; Zhao, J.; Song, J.; Song, H.; Li, D.; Maskey, N.; Zhao, B.; Wu, C.-C.; Xu, H.; et al. MiR-135b, Upregulated in Breast Cancer, Promotes Cell Growth and Disrupts the Cell Cycle by Regulating LATS2. Int. J. Oncol. 2016, 48, 1997–2006. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Ouyang, H.; He, D.; Yu, C.; Tang, G. MicroRNA-Based Potential Diagnostic, Prognostic and Therapeutic Applications in Triple-Negative Breast Cancer. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2800–2809. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Bahroudi, Z.; Shoorei, H.; Abak, A.; Ahin, M.; Taheri, M. MicroRNA-140: A MiRNA with Diverse Roles in Human Diseases. Biomed. Pharmacother. 2021, 135, 111256. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, B.; Wang, Y.; Chen, G.; Lian, Q.; Wang, H. MiR-140-3p Inhibits Breast Cancer Proliferation and Migration by Directly Regulating the Expression of Tripartite Motif 28. Oncol. Lett. 2019, 17, 3835–3841. [Google Scholar] [CrossRef]
- Do, Y.; Cho, J.G.; Park, J.Y.; Oh, S.; Park, D.; Yoo, K.H.; Lee, M.S.; Kwon, B.S.; Kim, J.; Yang, Y. MIR-146A Regulates Migration and Invasion by Targeting NRP2 in Circulating-Tumor Cell Mimicking Suspension Cells. Genes 2020, 12, 45. [Google Scholar] [CrossRef]
- Si, C.; Ye, Q.; Yao, Y. Effect of MiR-146a-5p on Proliferation and Metastasis of Triple-Negative Breast Cancer via Regulation of SOX5. Exp. Ther. Med. 2018, 15, 4515–4521. [Google Scholar] [CrossRef]
- Chiang, C.-C.; Hou, M.; Hung, W. Up-Regulation of MiR-182 by β-Catenin in Breast Cancer Increases Tumorigenicity and Invasiveness by Targeting the Matrix Metalloproteinase Inhibitor RECK. Biochim. Biophys. Acta (BBA)—Gen. General. Subj. 2013, 1830, 3067–3076. [Google Scholar] [CrossRef] [PubMed]
- Sameti, P.; Tohidast, M.; Amini, M.; Mahdavi, S.Z.B.; Najafi, S.; Mokhtarzadeh, A. The Emerging Role of MicroRNA-182 in Tumorigenesis; a Promising Therapeutic Target. Cancer Cell Int. 2023, 23, 134. [Google Scholar] [CrossRef]
- Humphries, B.A.; Wang, Z.; Oom, A.L.; Fisher, T.; Tan, D.; Cui, Y.; Jiang, Y.; Yang, C. MicroRNA-200b Targets Protein Kinase Cα and Suppresses Triple-Negative Breast Cancer Metastasis. Carcinogenesis 2014, 35, 2254–2263. [Google Scholar] [CrossRef]
- Pasculli, B.; Barbano, R.; Rendina, M.; Fontana, A.; Copetti, M.; Mazza, T.; Valori, V.M.; Morritti, M.; Maiello, E.; Graziano, P.; et al. Hsa-MiR-210-3p Expression in Breast Cancer and Its Putative Association with Worse Outcome in Patients Treated with Docetaxel. Sci. Rep. 2019, 9, 14913. [Google Scholar] [CrossRef]
- Kim, J.; Jung, E.J.; Kim, J.; Son, Y.; Lee, H.S.; Kwag, S.-J.; Park, J.-H.; Jeong, C.Y.; Kim, H.G.; Park, T.; et al. MiR-221 and MiR-222 Regulate Cell Cycle Progression and Affect Chemosensitivity in Breast Cancer by Targeting ANXA3. Exp. Ther. Med. 2023, 25, 127. [Google Scholar] [CrossRef]
- Li, B.; Lü, Y.; Yu, L.; Han, X.; Wang, H.; Murata, J.; Shen, J.; Wang, B.; Tang, J.; Li, C.; et al. MiR-221/222 Promote Cancer Stem-like Cell Properties and Tumor Growth of Breast Cancer via Targeting PTEN and Sustained Akt/NF-ΚB/COX-2 Activation. Chem.-Biol. Interact. 2017, 277, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Stinson, S.; Lackner, M.R.; Adai, A.; Yu, N.; Kim, H.; O’Brien, C.; Spoerke, J.M.; Jhunjhunwala, S.; Boyd, Z.; Januario, T.; et al. MiR-221/222 Targeting of Trichorhinophalangeal 1 (TRPS1) Promotes Epithelial-to-Mesenchymal Transition in Breast Cancer. Sci. Signal. 2011, 4, pt5. [Google Scholar] [CrossRef]
- Dinami, R.; Buemi, V.; Sestito, R.; Zappone, A.; Ciani, Y.; Mano, M.; Petti, E.; Sacconi, A.; Blandino, G.; Giacca, M.; et al. Epigenetic Silencing of MiR-296 and MiR-512 Ensures HTERT Dependent Apoptosis Protection and Telomere Maintenance in Basal-Type Breast Cancer Cells. Oncotarget 2017, 8, 95674–95691. [Google Scholar] [CrossRef]
- Crippa, E.; Lusa, L.; De Cecco, L.; Marchesi, E.; Calin, G.A.; Radice, P.; Manoukian, S.; Peissel, B.; Daidone, M.G.; Gariboldi, M.; et al. MiR-342 Regulates BRCA1 Expression through Modulation of ID4 in Breast Cancer. PLoS ONE 2014, 9, e87039. [Google Scholar] [CrossRef]
- Wang, F.; Wang, X.; Li, J.; Lv, P.; Han, M.; Li, L.; Chen, Z.; Li, D.; Wang, N.; Gu, Y. CircNOL10 Suppresses Breast Cancer Progression by Sponging MiR-767-5p to Regulate SOCS2/JAK/STAT Signaling. J. Biomed. Sci. 2021, 28, 4. [Google Scholar] [CrossRef]
- Hu, J.; Huang, H.; Xi, Z.; Ma, S.; Ming, J.; Dong, F.; Guo, H.; Zhang, H.; Zhao, E.; Yao, G.; et al. LncRNA SEMA3B-AS1 Inhibits Breast Cancer Progression by Targeting MiR-3940/KLLN Axis. Cell Death Dis. 2022, 13, 800. [Google Scholar] [CrossRef] [PubMed]
- Santana, T.A.B.D.S.; De Oliveira Passamai, L.; De Miranda, F.S.; Borin, T.F.; Borges, G.F.; Luiz, W.B.; Campos, L.C.G. The Role of MIRNAs in the Prognosis of Triple-Negative Breast Cancer: A Systematic Review and Meta-Analysis. Diagnostics 2022, 13, 127. [Google Scholar] [CrossRef]
- Improta-Caria, A.C.; Nonaka, C.K.V.; Cavalcante, B.R.R.; De Sousa, R.A.L.; Júnior, R.A.B.; De Freitas Souza, B.S. Modulation of MicroRNAs as a Potential Molecular Mechanism Involved in the Beneficial Actions of Physical Exercise in Alzheimer Disease. Int. J. Mol. Sci. 2020, 21, 4977. [Google Scholar] [CrossRef]
- Gasparini, P.; Cascione, L.; Fassan, M.; Lovat, F.; Güler, G.; Balcı, S.; Irkkan, Ç.; Morrison, C.; Croce, C.M.; Shapiro, C.L.; et al. MicroRNA Expression Profiling Identifies a Four MicroRNA Signature as a Novel Diagnostic and Prognostic Biomarker in Triple Negative Breast Cancers. Oncotarget 2014, 5, 1174–1184. [Google Scholar] [CrossRef]
- Cascione, L.; Gasparini, P.; Lovat, F.; Carasi, S.; Pulvirenti, A.; Ferro, A.; Alder, H.; He, G.; Vecchione, A.; Croce, C.M.; et al. Integrated MicroRNA and MRNA Signatures Associated with Survival in Triple Negative Breast Cancer. PLoS ONE 2013, 8, e55910. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Yan, B.; Lu, Q.; Lin, Y.; Ma, L. Reciprocal Regulation of Hsa-MiR-1 and Long Noncoding RNA MALAT1 Promotes Triple-Negative Breast Cancer Development. Tumor Biol. 2015, 37, 7383–7394. [Google Scholar] [CrossRef] [PubMed]
- Kołacińska, A.; Morawiec, J.; Fendler, W.; Małachowska, B.; Morawiec, Z.; Szemraj, J.; Pawłowska, Z.; Chowdhury, D.; Choi, Y.E.; Kubiak, R.; et al. Association of MicroRNAs and Pathologic Response to Preoperative Chemotherapy in Triple Negative Breast Cancer: Preliminary Report. Mol. Biol. Rep. 2014, 41, 2851–2857. [Google Scholar] [CrossRef]
- Chen, P.S.; Su, J.L.; Hung, M. Dysregulation of MicroRNAs in Cancer. J. Biomed. Sci. 2012, 19, 90. [Google Scholar] [CrossRef] [PubMed]
- Ml, S.; Zhu, S.; Wu, H.; Zhang, L.; Wu, F.; Mo, Y. MiR-21-Mediated Tumor Growth. Oncogene 2006, 26, 2799–2803. [Google Scholar] [CrossRef]
- Kovalchuk, O.; Filkowski, J.; Meservy, J.; Ilnytskyy, Y.; Tryndyak, V.; Pogribny, I.P. Involvement of MicroRNA-451 in Resistance of the MCF-7 Breast Cancer Cells to Chemotherapeutic Drug Doxorubicin. Mol. Cancer Ther. 2008, 7, 2152–2159. [Google Scholar] [CrossRef]
- Safaei, S.; Amini, M.; Najjary, S.; Mokhtarzadeh, A.; Bolandi, N.; Saeedi, H.; Alizadeh, N.; Javadrashid, D.; Baradaran, B. MiR-200c Increases the Sensitivity of Breast Cancer Cells to Doxorubicin through Downregulating MDR1 Gene. Exp. Mol. Pathol. 2022, 125, 104753. [Google Scholar] [CrossRef] [PubMed]
- Salter, K.H.; Acharya, C.R.; Walters, K.S.; Redman, R.C.; Anguiano, A.; Garman, K.S.; Anders, C.K.; Mukherjee, S.; Dressman, H.K.; Barry, W.T.; et al. An Integrated Approach to the Prediction of Chemotherapeutic Response in Patients with Breast Cancer. PLoS ONE 2008, 3, e1908. [Google Scholar] [CrossRef] [PubMed]
- Szczepanek, J.; Skorupa, M.; Tretyn, A. MicroRNA as a Potential Therapeutic Molecule in Cancer. Cells 2022, 11, 1008. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zhao, Z.; Zhou, H.; Liu, Y.; Yang, H. Systematic Analysis of MicroRNA Involved in Resistance of the MCF-7 Human Breast Cancer Cell to Doxorubicin. Med. Oncol. 2009, 27, 406–415. [Google Scholar] [CrossRef]
- Xu, C.; Du, Z.; Ren, S.; Pian, Y. Downregulation of GSK3B by MIR-132-3P enhances Etoposide-Induced Breast Cancer cell apoptosis. Ann. Clin. Lab. Sci. 2021, 51, 285–294. [Google Scholar]
- Hu, Q.; Chen, W.; Zhong, S.; Zhang, J.; Ma, T.; Jiang, H.; Lv, M.; Tang, J.; Zhao, J. MicroRNA-452 Contributes to the Docetaxel Resistance of Breast Cancer Cells. Tumor Biol. 2014, 35, 6327–6334. [Google Scholar] [CrossRef]
- Xin, F.; Li, M.; Balch, C.; Thomson, M.J.; Fan, M.; Liu, Y.; Hammond, S.M.; Kim, S.; Nephew, K.P. Computational Analysis of MicroRNA Profiles and Their Target Genes Suggests Significant Involvement in Breast Cancer Antiestrogen Resistance. Bioinformatics 2008, 25, 430–434. [Google Scholar] [CrossRef]
- Brown, C.; Mantzaris, M.D.; Nicolaou, E.; Karanasiou, G.S.; Papageorgiou, E.; Curigliano, G.; Cardinale, D.; Filippatos, G.; Memos, N.; Naka, K.K.; et al. A Systematic Review of MiRNAs as Biomarkers for Chemotherapy-Induced Cardiotoxicity in Breast Cancer Patients Reveals Potentially Clinically Informative Panels as Well as Key Challenges in MiRNA Research. Cardio-Oncol. 2022, 8, 16. [Google Scholar] [CrossRef]
- Davey, M.G.; Abbas, R.; Kerin, E.; Casey, M.C.; McGuire, A.; Waldron, R.; Heneghan, H.; Newell, J.; McDermott, A.M.; Keane, M.M.; et al. Circulating MicroRNAs Can Predict Chemotherapy-Induced Toxicities in Patients Being Treated for Primary Breast Cancer. Breast Cancer Res. Treat. 2023, 202, 73–81. [Google Scholar] [CrossRef]
- Schmid, P.; Cortés, J.; Pusztai, L.; McArthur, H.L.; Kümmel, S.; Bergh, J.; Denkert, C.; Park, Y.H.; Hui, R.; Harbeck, N.; et al. Pembrolizumab for Early Triple-Negative Breast Cancer. N. Engl. J. Med. 2020, 382, 810–821. [Google Scholar] [CrossRef]
- Ding, L.; Gu, H.; Xiong, X.; Ao, H.; Cao, J.; Lin, W.T.; Yu, M.; Lin, J.; Cui, Q. MicroRNAs Involved in Carcinogenesis, Prognosis, Therapeutic Resistance, and Applications in Human Triple-Negative Breast Cancer. Cells 2019, 8, 1492. [Google Scholar] [CrossRef]
- Ma, L.; Reinhardt, F.; Pan, E.; Soutschek, J.; Bhat, B.; Marcusson, E.G.; Teruya-Feldstein, J.; Bell, G.W.; Weinberg, R.A. Therapeutic Silencing of MiR-10b Inhibits Metastasis in a Mouse Mammary Tumor Model. Nat. Biotechnol. 2010, 28, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, Z.; Chen, C.; Liu, Y.; Si, Q.; Chuang, T.H.; Gómez-Cabrero, A.; Ra, R.; Xiang, R.; Luo, Y. MicroRNA-19a-3p Inhibits Breast Cancer Progression and Metastasis by Inducing Macrophage Polarization through Downregulated Expression of Fra-1 Proto-Oncogene. Oncogene 2013, 33, 3014–3023. [Google Scholar] [CrossRef]
- Zhu, H.; Wu, H.; Liu, X.; Evans, B.R.; Medina, D.; Liu, C.G.; Yang, J.M. Role of MicroRNA MiR-27a and MiR-451 in the Regulation of MDR1/P-Glycoprotein Expression in Human Cancer Cells. Biochem. Pharmacol. 2008, 76, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xie, X.; Luo, J.; Liu, M.; Xi, S.; Guo, J.; Kong, Y.; Wu, P.; Gao, J.; Xie, Z.; et al. Targeted Expression of MIR-34A Using the T-VISA System Suppresses Breast Cancer Cell Growth and Invasion. Mol. Ther. 2012, 20, 2326–2334. [Google Scholar] [CrossRef]
- Kim, S.-J.; Oh, J.; Shin, J.; Lee, K.-D.; Sung, K.W.; Nam, S.J.; Chun, K. Development of MicroRNA-145 for Therapeutic Application in Breast Cancer. J. Control Release 2011, 155, 427–434. [Google Scholar] [CrossRef]
- Sachdeva, M.; Zhu, S.; Wu, F.; Wu, H.; Walia, V.; Kumar, S.; Elble, R.C.; Watabe, K.; Mo, Y. P53 Represses C-Myc through Induction of the Tumor Suppressor MiR-145. Proc. Natl. Acad. Sci. USA 2009, 106, 3207–3212. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Wu, H.; Xia, J.; Li, Y.; Zhang, Y.; Huang, K.; Wagar, N.; Yoon, Y.; Cho, H.T.; Scala, S.; et al. Involvement of MiR-326 in Chemotherapy Resistance of Breast Cancer through Modulating Expression of Multidrug Resistance-Associated Protein 1. Biochem. Pharmacol. 2010, 79, 817–824. [Google Scholar] [CrossRef]
- Bao, L.; Hazari, S.; Mehra, S.; Kaushal, D.; Moroz, K.; Dash, S. Increased Expression of P-Glycoprotein and Doxorubicin Chemoresistance of Metastatic Breast Cancer Is Regulated by MIR-298. Am. J. Pathol. 2012, 180, 2490–2503. [Google Scholar] [CrossRef]
- Saliminejad, K.; Khorshid, H.R.K.; Ghaffari, S.H. Why Have microRNA Biomarkers Not Been Translated from Bench to Clinic? Future Oncol. 2019, 15, 801–803. [Google Scholar] [CrossRef]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating microRNAs as Potential Cancer Biomarkers: The Advantage and Disadvantage. Clin. Epigenetics 2018, 10, 59. [Google Scholar] [CrossRef] [PubMed]
- Siddika, T.; Heinemann, I.U. Bringing MicroRNAs to Light: Methods for MicroRNA Quantification and Visualization in Live Cells. Front. Bioeng. Biotechnol. 2021, 8, 619583. [Google Scholar] [CrossRef]
- Sayed, S.R.E.; Cristante, J.; Guyon, L.; Denis, J.; Chabre, O.; Cherradi, N. MicroRNA Therapeutics in Cancer: Current Advances and Challenges. Cancers 2021, 13, 2680. [Google Scholar] [CrossRef] [PubMed]
- Orellana, E.A.; Kasinski, A.L. MicroRNAs in Cancer: A Historical Perspective on the Path from Discovery to Therapy. Cancers 2015, 7, 1388–1405. [Google Scholar] [CrossRef] [PubMed]
- Silvestro, S.; Mazzon, E. MIRNAs as Promising Translational Strategies for Neuronal Repair and Regeneration in Spinal Cord Injury. Cells 2022, 11, 2177. [Google Scholar] [CrossRef]
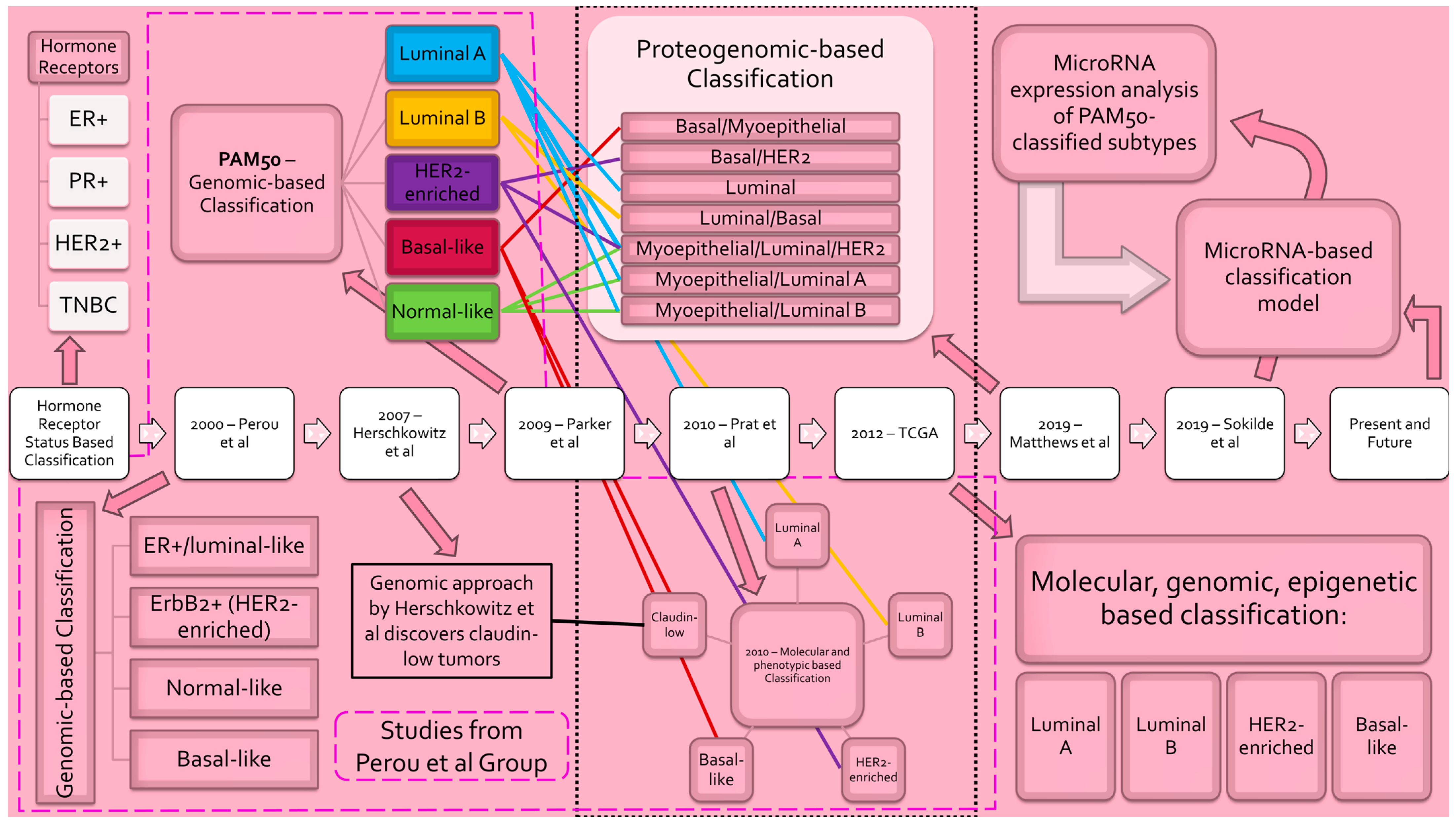
Figure 1.
Evolution of breast cancer subclassification. Progression of breast cancer molecular subtyping schema [12,17,19,29,30,33,34]. Classification models are listed in chronological order from left to right. Blue lines denote correspondence to PAM50 luminal A subtype. Yellow lines denote correspondence to PAM50 luminal B subtype. Purple lines denote correspondence to PAM50 HER2-enriched subtype. Red lines denote correspondence to PAM50 basal-like subtype. Green lines denote correspondence to PAM50 normal-like subtype. _____ denotes correspondence to Herschkowitz et al.’s claudin-low subtype. - - - - encompasses studies completed by the Perou et al. group. ......... denotes classification model incorporating PAM50. [Abbreviations: ER—estrogen receptor; PR—progesterone receptor; HER2—human epidermal growth factor receptor 2; TNBC—triple-negative breast cancer; ErbB2—avian erythroblastic leukemia viral oncogene homolog 2; PAM50—prediction analysis of microarray 50].
Figure 1.
Evolution of breast cancer subclassification. Progression of breast cancer molecular subtyping schema [12,17,19,29,30,33,34]. Classification models are listed in chronological order from left to right. Blue lines denote correspondence to PAM50 luminal A subtype. Yellow lines denote correspondence to PAM50 luminal B subtype. Purple lines denote correspondence to PAM50 HER2-enriched subtype. Red lines denote correspondence to PAM50 basal-like subtype. Green lines denote correspondence to PAM50 normal-like subtype. _____ denotes correspondence to Herschkowitz et al.’s claudin-low subtype. - - - - encompasses studies completed by the Perou et al. group. ......... denotes classification model incorporating PAM50. [Abbreviations: ER—estrogen receptor; PR—progesterone receptor; HER2—human epidermal growth factor receptor 2; TNBC—triple-negative breast cancer; ErbB2—avian erythroblastic leukemia viral oncogene homolog 2; PAM50—prediction analysis of microarray 50].


{kind=link}
Table 1.
Summary of the dual functional roles of microRNAs in breast cancer.
| List of MicroRNA | Functional Role in Breast Cancer | Regulated Expression in Breast Cancer | Target(s) | Reference(s) |
|---|---|---|---|---|
| miR-10b | OncomiR | Upregulated | HOXD10 | [70,71] |
| miR-21 | OncomiR | Upregulated | PDCD4, PTEN | [72] |
| miR-155 | OncomiR | Upregulated | SOCS1, TP53INP1, FOXO3, RhoA | [73] |
| miR-182-5p | OncomiR | Upregulated | CASP9 | [74] |
| miR-191 | OncomiR | Upregulated | DICER1 | [61] |
| miR-375 | OncomiR | Upregulated | RASD1 | [75,76] |
| miR-425 | OncomiR | Upregulated | DICER1 | [61] |
| miR-125 | Tumor suppressor | Downregulated | HuR, HER2, ETS1, Cyclin J, MEGF9 | [73,76,77] |
| miR-145 | Tumor suppressor | Downregulated | ARF6, fascin, JAM-A, C-MYC, ROCK1, CCNE2, RTKN, OCT4, MUC1, FSCN1, Rab27a | [78,79] |
| miR-200 family | Tumor suppressor | Differential | ZEB1, ZEB2 | [73,76,80,81] |
| miR-205 | Tumor suppressor | Downregulated | ZEB1, ZEB2, E2F1, LAMC1, KLF12, NFIB, AMOT, ERp29, HER3, CLCN3, SIP1, Ubc13, HMGB1/3, RunX2, ITGA5, VEGF-A, FGF2 | [73,76,82] |
Abbreviations: HOXD10—homeobox D10; PDCD4—programmed cell death 4; TPM1—tropomyosin-1; PTEN—phosphatase and TENsin homolog deleted; CDC25A—cell division cycle 25 A; RECK—reversion-inducing-cysteine-rich protein with kazal motifs; MASPIN—mammary serine protease inhibitor; TIMP3—tissue inhibitor of metalloproteinase-3; SOCS1—suppressor of cytokine signaling 1; TP53INP1—tumor protein 53-induced nuclear protein 1; FOXO3—forkhead box protein O3; RhoA—Ras homolog family member A; CASP9—caspase 9; DICER1—dicer 1, ribonuclease III; RASD1—dexamethasone-induced Ras-related protein 1; HuR—human antigen R; HER2—human epidermal growth factor receptor 2; ETS1—E26 transformation specific proto-oncogene 1; MEGF9—multiple epidermal growth factor like-domains 9; ARF6—adenosine diphosphate-ribosylation factor 6; JAM-A—junctional adhesion molecule A; C-MYC—cellular myelocytomatosis; ROCK1—rho-associated coiled-coil-containing protein kinase 1; CCNE2—cycline E2; RTKN—rhotekin; OCT4—octamer-binding transcription factor 4; MUC1—mucin 1; FSCN1—fascin actin-binding protein 1; ZEB1—zinc finger E-box binding homeobox 1; ZEB2—zinc finger E-box binding homeobox 2; E2F1—E2 promoter binding factor 1; LAMC1—laminin γ1 chain; KLF12—Krueppel-like factor 12; NFIB—nuclear factor 1 B-type; AMOT—angiomotin; ERp29—endoplasmic reticulum protein 29; HER3—human epidermal growth factor receptor 3; CLCN3—chloride voltage-gated channel 3; SIP1—Smad-interacting protein 1; Ubc13—ubiquitin-conjugating enzyme 13; HMGB1—high mobility group box 1; HMGB3—high mobility group box 3, RunX2—runt-related transcription factor 2; ITGA5—integrin alpha-5; VEGF-A—vascular endothelial growth factor A; FGF2—fibroblast growth factor 2.
Table 2.
MicroRNA signature panels that distinguish BLBC/TNBC from other subtypes.
| List of MicroRNA(s) | Sample Subtype | Significance of miRNA Expression | Reference(s) |
|---|---|---|---|
| miR-21 | BLBC | Increased expression is highly associated with BLBC. | [91] |
| miR-101-3p miR-126-3p miR-126-5p miR-144-3p miR-144-5p miR-301a-3p miR-664b-5p | Basal-like TNBC | MiRNA panel differentiated healthy women from patients with basal-like TNBC with a sensitivity and specificity of 84% and 74%, respectively. | [84] |
| miR-205-5p miR-224-5p miR-375 | BLBC | BLBC differentiated from non-basal TNBCs. | [92] |
| miR-146a miR-26a | TNBC | Significantly upregulated in TNBC relative to non-TNBC. | [93] |
| miR-146b miR-148a miR-200a miR-200b | TNBC | Downregulated specifically in TNBC. | [94] |
| ER status signature miRNAs: miR-135b miR-190 miR-217 miR-218 miR-299 miR-342 PR status signature miRNAs: miR-377 miR-520f-520c miR-520g miR-527-518a HER2 status signature miRNAs: miR-30e miR-181c miR-302c miR-376b miR-520d | TNBC | The miRNA signatures for ER, PR, and HER2 status were able to predict hormone receptor positivity/negativity with 100% accuracy. Incorporating all three panels together can allow for delineation of TNBC from other subtypes. | [95] |
| miR-210-3p miR-105-5p miR-767-5p miR-5683 | TNBC | Upregulated in TNBC. Downregulated in TNBC. | [90] |
Abbreviation: BLBC—basal-like breast cancer; TNBC—triple negative breast cancer; ER—estrogen receptor; PR—progesterone receptor; HER2—human epidermal growth factor receptor 2.
Table 3.
MicroRNAs discerned as oncomiRs or tumor suppressors in BLBC/TNBC.
| MicroRNA | Functional Role in BLBC | BLBC/TNBC | Differential Expression | Validated Target(s) | Reference(s) |
|---|---|---|---|---|---|
| miR-20a-5p | OncomiR | TNBC | Upregulated | RUNX3 | [97] |
| miR-29a | OncomiR | TNBC | Upregulated | SUV420H2 | [47,68] |
| miR-29b-3p | OncomiR | TNBC | Upregulated | TRAF3 | [67] |
| miR-93 | OncomiR | TNBC | Upregulated | LATS2, JAK1, STAT3, FBXL5, SOX4, EZH1, HMGA2 | [87] |
| miR-105-5p | OncomiR | TNBC | Upregulated | AKT1, GRB2 | [90,105] |
| miR-135b | OncomiR | BLBC | Upregulated | LATS2 | [98,106] |
| miR-136 | Tumor suppressor | TNBC | Downregulated | RASAL2 | [107] |
| miR-140 | Tumor suppressor | BLBC | Downregulated | SOX9, ALDH1, TRIM28 | [102,108,109] |
| miR-146a | OncomiR | TNBC | Upregulated | BRCA1, NRP2 | [107,110] |
| miR-146a-5p | Tumor suppressor | TNBC | Downregulated | SOX5 | [111] |
| miR-181a | OncomiR | TNBC | Upregulated | BIM, ATM, BAX | [107] |
| miR-181b | OncomiR | TNBC | Upregulated | BIM, ATM | [107] |
| miR-182 | OncomiR | TNBC | Upregulated | PFN1, RECK, FOXF2 | [107,112,113] |
| miR-182-3p | Tumor suppressor | TNBC | Downregulated | TRF2 | [104] |
| miR-200a | Tumor suppressor | BLBC and TNBC | Downregulated | EPHA2, ZEB1, ZEB2 | [94,107] |
| miR-200b | Tumor suppressor | BLBC and TNBC | Downregulated | PKCα, ZEB1, ZEB2 | [94,107,114] |
| miR-210-3p | OncomiR | TNBC | Upregulated | GPD1L, CYGB | [90,115] |
| miR-221 | OncomiR | BLBC and TNBC | Upregulated | ANXA3, PTEN, TRPS1 | [107,116,117,118] |
| miR-222 | OncomiR | BLBC | Upregulated | ANXA3, PTEN, TRPS1 | [116,117,118] |
| miR-296-5p | Tumor suppressor | BLBC | Downregulated | hTERT | [119] |
| miR-342 | OncomiR | BLBC and TNBC | Upregulated | ID4 | [107,120] |
| miR-512-5p | Tumor suppressor | BLBC | Downregulated | hTERT | [119] |
| miR-767-5p | OncomiR | TNBC | Upregulated | SOCS2 | [90,121] |
| miR-3940-3p | OncomiR | TNBC | Upregulated | KLLN | [122] |
Abbreviations: BLBC—basal-like breast cancer; TNBC—triple-negative breast cancer; RUNX3—runt-related transcription factor 3; SUV420H2—suppressor of variegation 4–20 homolog 2; TRAF3—tumor necrosis factor receptor associated factor 3; LATS2—large tumor suppressor kinase 2; JAK1—Janus kinase 1; STAT3—signal transducer and activator of transcription 3; FBXL5—F-box and leucine-rich repeat protein 5; SOX4—sex-determining region Y-related high-mobility group box 4; EZH1—enhancer of zeste 1; HMGA2—High-mobility group AT-hook 2; AKT1—Ak strain transforming; GRB2—Growth Factor Receptor-bound protein 2; RASAL2—RAS protein activator like 2; SOX9—sex-determining region Y-related high-mobility group box 9; ALDH1—aldehyde dehydrogenase 1; TRIM28—tripartite motif-containing 28; BRCA1—breast cancer gene 1; NRP2—neuropilin 2; SOX5—sex-determining region Y-related high-mobility group box 5; BIM—Bcl-2-like protein 11; ATM-ataxia telangiectasia–mutated; BAX—Bcl-2-associated X protein; PFN1—profilin 1; RECK—reversion-inducing-cysteine-rich protein with kazal motifs; FOXF2—forkhead box F2; TRF2—telomeric repeat-binding factor 2; EPHA2—ephrin type-A receptor 2; ZEB1—zinc finger E-box binding homeobox 1; ZEB2—zinc finger E-box binding homeobox 2; PKCα—protein kinase C alpha; GPD1L—glycerol-3-phosphate dehydrogenase 1-like; CYGB—cytoglobin; ANXA3—annexin A3; PTEN-phosphatase and TENsin homolog deleted; TRPS1—transcriptional repressor GATA binding 1; hTERT—human telomerase reverse transcriptase; ID4—Inhibitor Of DNA Binding 4; SOCS2—suppressor of cytokine signaling 2; KLLN—Killin, p53-regulated DNA replication inhibitor.
Table 4.
MiRNAs signature panels and their prognostic implication in BLBC/TNBC.
| MicroRNAs in Panel | BLBC/TNBC | Prognostic Implication | Reference(s) |
|---|---|---|---|
| miR-155 | TNBC | Significantly associated with overall survival (p < 0.01). | [123,125,126] |
| miR-30a-5p miR-30a-3p miR-30c-5p | Basal-like TNBC and TNBC | Dysregulation of miR-30 expression was seen to impede the recurrence free survival. | [92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123] |
| miR-30a-3p miR-30a-5p miR-199a-5p miR-203b-5p miR-324-5p let-7d-3p | Basal-like TNBC and TNBC | Panel signatures were significantly associated with decreased overall survival (OS). | [92] |
| miR-30a-3p miR-30a-5p miR-30c-5p miR-128-3p let-7d-3p | Basal-like TNBC and TNBC | Panel signatures were significantly associated with decreased relapse-free survival (RFS). | [92] |
| miR-95-3p | Basal-like TNBC and TNBC | Elevated expression of miR-95-3p in patients treated with anthracycline chemotherapeutics correlates with a diminished overall survival (p = 0.03) and relapse-free survival (p < 0.01). | [92] |
| miR-146a miR-26a miR-10b miR-153 | TNBC | Expression of miR-146a and miR-26a is greater in node-negative TNBC relative to node-positive TNBC (p = 0.04 and 0.01, respectively), whereas miR-10b showed the inverse relationship (p = 0.05). | [93] |
| miR-1 | TNBC | MALAT1, a long noncoding RNA upregulated in TNBC, possesses known oncogenic function and has a reciprocal relationship with miR-1 expression. Excessive MALAT1 expression is associated with poor overall survival relative to low. | [127] |
| miR-27a miR-30e miR-155 miR-493 | BLBC | Panel signatures were significantly associated with median overall survival (p < 0.01). | [125] |
| miR-10b miR-21 miR-31 miR-125b miR-130a-3p miR-155 miR-181a miR-181b miR-451a | TNBC | The listed miRNAs were all found to be dysregulated in TNBC and were found to be associated with pathways of chemotherapy resistance. | [89] |
| miR-190a miR-200b-3p miR-512-5p | TNBC | A panel incorporating expression levels of the three listed miRNAs was found to be associated with favorable chemotherapy activity (p = 0.06). | [128] |
| miR-125b miR-655 miR-421 miR-16 miR-374a miR-374b miR-497 | TNBC | Panel signatures were significantly associated with median distant disease-free survival (DFS) (p < 0.01). | [126] |
| miR-16 miR-155 miR-374a miR-125b | TNBC | Panel signatures were significantly associated with overall survival (p = 0.05). | [126] |
| miR-148a miR-200b | TNBC and BLBC | Both miRNAs modulate the E2F pathway, whose function is associated with poor prognosis, and both are significantly under expressed in TNBC. | [94] |
Abbreviations: BLBC—basal-like breast cancer; TNBC—triple-negative breast cancer; MALAT1—metastasis associated in lung adenocarcinoma transcript 1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tariq, M.; Richard, V.; Kerin, M.J. MicroRNAs as Molecular Biomarkers for the Characterization of Basal-like Breast Tumor Subtype. Biomedicines 2023, 11, 3007. https://doi.org/10.3390/biomedicines11113007
AMA Style
Tariq M, Richard V, Kerin MJ. MicroRNAs as Molecular Biomarkers for the Characterization of Basal-like Breast Tumor Subtype. Biomedicines. 2023; 11(11):3007. https://doi.org/10.3390/biomedicines11113007
Chicago/Turabian StyleTariq, Muhammad, Vinitha Richard, and Michael J. Kerin. 2023. "MicroRNAs as Molecular Biomarkers for the Characterization of Basal-like Breast Tumor Subtype" Biomedicines 11, no. 11: 3007. https://doi.org/10.3390/biomedicines11113007
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.