
Clinical Relevance of Gut Microbiota Alterations under the Influence of Selected Drugs—Updated Review
 , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
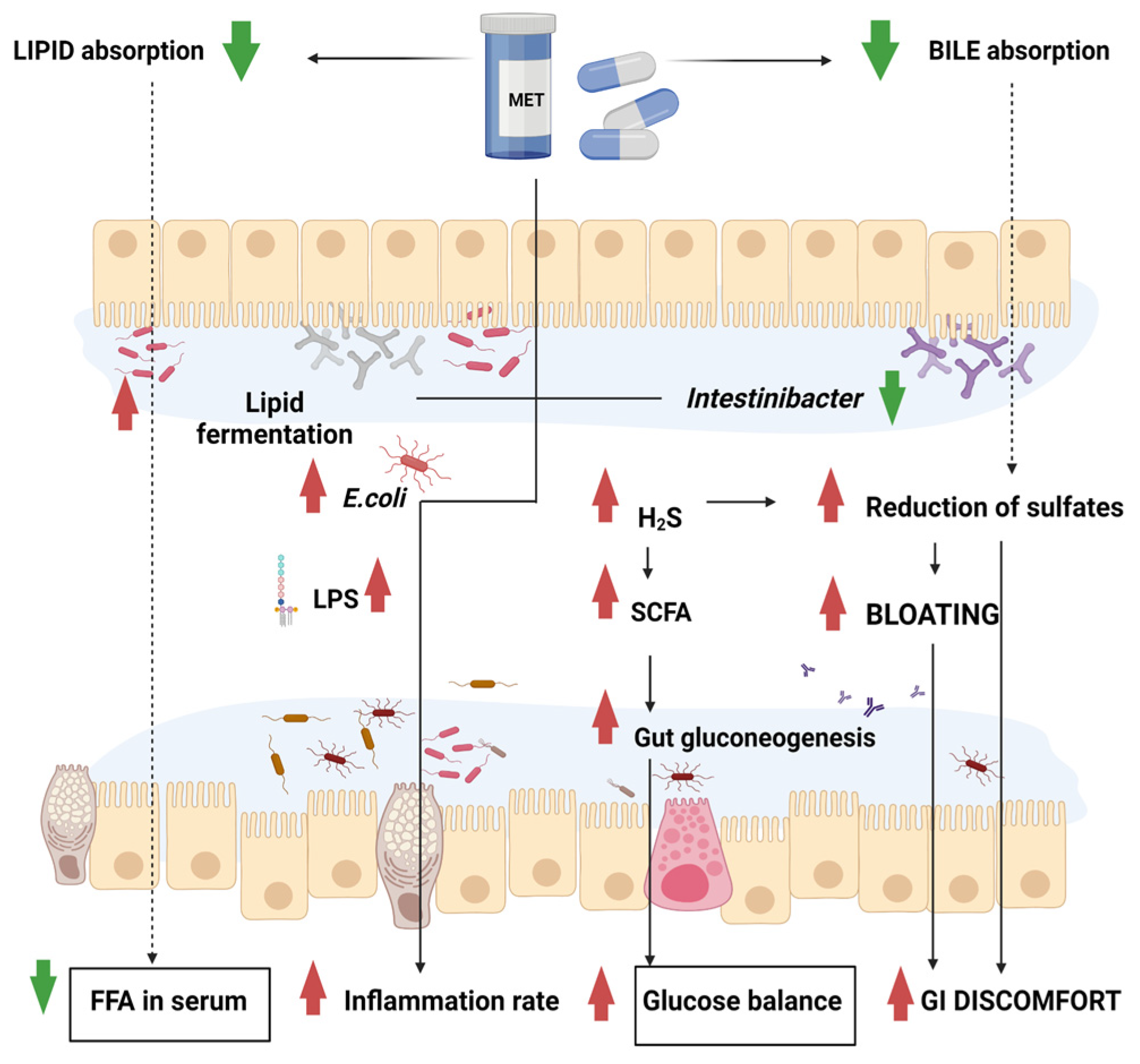
2. Metformin
3. Non-Steroidal Anti-Inflammatory Drugs
4. Proton Pump Inhibitors (PPIs)
5. Statins
6. Laxatives
7. Phytochemicals
8. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blaser, M.J. The Microbiome Revolution. J. Clin. Investig. 2014, 124, 4162–4165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, I.D.; Nicholson, J.K. Gut Microbiome Interactions with Drug Metabolism, Efficacy, and Toxicity. Transl. Res. 2017, 179, 204–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, T.; Paterson, R.; Moore, V.; Carlsson, A.; Abrahamsson, B.; Basit, A.W. The Gastrointestinal Microbiota as a Site for the Biotransformation of Drugs. Int. J. Pharm. 2008, 363, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Jourova, L.; Anzenbacher, P.; Anzenbacherova, E. Human Gut Microbiota Plays a Role in the Metabolism of Drugs. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech Repub. 2016, 160, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Jourova, L.; Anzenbacher, P.; Matuskova, Z.; Vecera, R.; Strojil, J.; Kolar, M.; Nobilis, M.; Hermanova, P.; Hudcovic, T.; Kozakova, H.; et al. Gut Microbiota Metabolizes Nabumetone in Vitro: Consequences for Its Bioavailability in Vivo in the Rodents with Altered Gut Microbiome. Xenobiotica 2019, 49, 1296–1302. [Google Scholar] [CrossRef]
- McCabe, M.; Sane, R.S.; Keith-Luzzi, M.; Xu, J.; King, I.; Whitcher-Johnstone, A.; Johnstone, N.; Tweedie, D.J.; Li, Y. Defining the Role of Gut Bacteria in the Metabolism of Deleobuvir: In Vitro and In Vivo Studies. Drug Metab. Dispos. 2015, 43, 1612–1618. [Google Scholar] [CrossRef]
- Boer, J.; Young-Sciame, R.; Lee, F.; Bowman, K.J.; Yang, X.; Shi, J.G.; Nedza, F.M.; Frietze, W.; Galya, L.; Combs, A.P.; et al. Roles of UGT, P450, and Gut Microbiota in the Metabolism of Epacadostat in Humans. Drug Metab. Dispos. 2016, 44, 1668–1674. [Google Scholar] [CrossRef] [Green Version]
- Misera, A.; Łoniewski, I.; Palma, J.; Kulaszyńska, M.; Czarnecka, W.; Kaczmarczyk, M.; Liśkiewicz, P.; Samochowiec, J.; Skonieczna-Żydecka, K. Clinical Significance of Microbiota Changes under the Influence of Psychotropic Drugs. An Updated Narrative Review. Front. Microbiol. 2023, 14, 548. [Google Scholar] [CrossRef]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillère, R.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, M.J.; et al. The Intestinal Microbiota Modulates the Anticancer Immune Effects of Cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef] [Green Version]
- Vich Vila, A.; Collij, V.; Sanna, S.; Sinha, T.; Imhann, F.; Bourgonje, A.R.; Mujagic, Z.; Jonkers, D.M.A.E.; Masclee, A.A.M.; Fu, J.; et al. Impact of Commonly Used Drugs on the Composition and Metabolic Function of the Gut Microbiota. Nat. Commun. 2020, 11, 362. [Google Scholar] [CrossRef] [Green Version]
- Feinglos, M.N.; Bethel, M.A. Treatment of Type 2 Diabetes Mellitus. Med. Clin. North Am. 1998, 82, 757–790. [Google Scholar] [CrossRef] [PubMed]
- Nabrdalik, K.; Skonieczna-Żydecka, K.; Irlik, K.; Hendel, M.; Kwiendacz, H.; Łoniewski, I.; Januszkiewicz, K.; Gumprecht, J.; Lip, G.Y.H. Gastrointestinal Adverse Events of Metformin Treatment in Patients with Type 2 Diabetes Mellitus: A Systematic Review, Meta-Analysis and Meta-Regression of Randomized Controlled Trials. Front. Endocrinol. 2022, 13, 975912. [Google Scholar] [CrossRef] [PubMed]
- Bryrup, T.; Thomsen, C.W.; Kern, T.; Allin, K.H.; Brandslund, I.; Jørgensen, N.R.; Vestergaard, H.; Hansen, T.; Hansen, T.H.; Pedersen, O.; et al. Metformin-Induced Changes of the Gut Microbiota in Healthy Young Men: Results of a Non-Blinded, One-Armed Intervention Study. Diabetologia 2019, 62, 1024–1035. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Sheng, L.; Li, H. Akkermansia muciniphila: Is It the Holy Grail for Ameliorating Metabolic Diseases? Gut Microbes 2021, 13, 1984104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ni, Y.; Qian, L.; Fang, Q.; Zheng, T.; Zhang, M.; Gao, Q.; Zhang, Y.; Ni, J.; Hou, X.; et al. Decreased Abundance of Akkermansia muciniphila Leads to the Impairment of Insulin Secretion and Glucose Homeostasis in Lean Type 2 Diabetes. Adv. Sci. 2021, 8, 2100536. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in Overweight and Obese Human Volunteers: A Proof-of-Concept Exploratory Study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Shin, N.-R.; Lee, J.-C.; Lee, H.-Y.; Kim, M.-S.; Whon, T.W.; Lee, M.-S.; Bae, J.-W. An Increase in the Akkermansia Spp. Population Induced by Metformin Treatment Improves Glucose Homeostasis in Diet-Induced Obese Mice. Gut 2014, 63, 727–735. [Google Scholar] [CrossRef] [Green Version]
- Forslund, K.; Hildebrand, F.; Nielsen, T.; Falony, G.; Le Chatelier, E.; Sunagawa, S.; Prifti, E.; Vieira-Silva, S.; Gudmundsdottir, V.; Krogh Pedersen, H.; et al. Disentangling Type 2 Diabetes and Metformin Treatment Signatures in the Human Gut Microbiota. Nature 2015, 528, 262–266. [Google Scholar] [CrossRef] [Green Version]
- Messori, S.; Trevisi, P.; Simongiovanni, A.; Priori, D.; Bosi, P. Effect of Susceptibility to Enterotoxigenic Escherichia Coli F4 and of Dietary Tryptophan on Gut Microbiota Diversity Observed in Healthy Young Pigs. Vet. Microbiol. 2013, 162, 173–179. [Google Scholar] [CrossRef]
- Winter, S.E.; Winter, M.G.; Xavier, M.N.; Thiennimitr, P.; Poon, V.; Keestra, A.M.; Laughlin, R.C.; Gomez, G.; Wu, J.; Lawhon, S.D.; et al. Host-Derived Nitrate Boosts Growth of E. Coli in the Inflamed Gut. Science 2013, 339, 708–711. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergström, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Bäckhed, F. Gut Metagenome in European Women with Normal, Impaired and Diabetic Glucose Control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef] [PubMed]
- de la Cuesta-Zuluaga, J.; Mueller, N.T.; Corrales-Agudelo, V.; Velásquez-Mejía, E.P.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Metformin Is Associated With Higher Relative Abundance of Mucin-Degrading Akkermansia muciniphila and Several Short-Chain Fatty Acid-Producing Microbiota in the Gut. Diabetes Care 2017, 40, 54–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastor-Villaescusa, B.; Plaza-Díaz, J.; Egea-Zorrilla, A.; Leis, R.; Bueno, G.; Hoyos, R.; Vázquez-Cobela, R.; Latorre, M.; Cañete, M.D.; Caballero-Villarraso, J.; et al. Evaluation of the Gut Microbiota after Metformin Intervention in Children with Obesity: A Metagenomic Study of a Randomized Controlled Trial. Biomed. Pharmacother. 2021, 134, 111117. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Wu, Q.; Osada, H.; Yoshida, M.; Pan, W.; Qi, J. Peanut Skin Extract Ameliorates the Symptoms of Type 2 Diabetes Mellitus in Mice by Alleviating Inflammation and Maintaining Gut Microbiota Homeostasis. Aging 2020, 12, 13991–14018. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.A.; Shapiro, J.A.; Church, T.R.; Miller, G.; Trinh-Shevrin, C.; Yuen, E.; Friedlander, C.; Hayes, R.B.; Ahn, J. A Taxonomic Signature of Obesity in a Large Study of American Adults. Sci. Rep. 2018, 8, 9749. [Google Scholar] [CrossRef] [Green Version]
- Sroka-Oleksiak, A.; Młodzińska, A.; Bulanda, M.; Salamon, D.; Major, P.; Stanek, M.; Gosiewski, T. Metagenomic Analysis of Duodenal Microbiota Reveals a Potential Biomarker of Dysbiosis in the Course of Obesity and Type 2 Diabetes: A Pilot Study. J. Clin. Med. 2020, 9, 369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbere, I.; Kalnina, I.; Silamikelis, I.; Konrade, I.; Zaharenko, L.; Sekace, K.; Radovica-Spalvina, I.; Fridmanis, D.; Gudra, D.; Pirags, V.; et al. Association of Metformin Administration with Gut Microbiome Dysbiosis in Healthy Volunteers. PLoS ONE 2018, 13, e0204317. [Google Scholar] [CrossRef] [Green Version]
- Cernea, S.; Dima, L.; Correll, C.U.; Manu, P. Pharmacological Management of Glucose Dysregulation in Patients Treated with Second-Generation Antipsychotics. Drugs 2020, 80, 1763–1781. [Google Scholar] [CrossRef]
- Skonieczna-Żydecka, K.; Łoniewski, I.; Misera, A.; Stachowska, E.; Maciejewska, D.; Marlicz, W.; Galling, B. Second-Generation Antipsychotics and Metabolism Alterations: A Systematic Review of the Role of the Gut Microbiome. Psychopharmacology 2019, 236, 1491–1512. [Google Scholar] [CrossRef] [Green Version]
- Rogers, M.A.M.; Aronoff, D.M. The Influence of Nonsteroidal Anti-Inflammatory Drugs on the Gut Microbiome. Clin. Microbiol. Infect. 2016, 22, 178.e1–178.e9. [Google Scholar] [CrossRef] [Green Version]
- Mäkivuokko, H.; Tiihonen, K.; Tynkkynen, S.; Paulin, L.; Rautonen, N. The Effect of Age and Non-Steroidal Anti-Inflammatory Drugs on Human Intestinal Microbiota Composition. Br. J. Nutr. 2010, 103, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Battaglia, T.; Aleman, J.O.; Walker, J.M.; Blaser, M.J.; Holt, P.R. Celecoxib Does Not Alter Intestinal Microbiome in a Longitudinal Diet-Controlled Study. Clin. Microbiol. Infect. 2016, 22, 464–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edogawa, S.; Peters, S.A.; Jenkins, G.D.; Gurunathan, S.V.; Sundt, W.J.; Johnson, S.; Lennon, R.J.; Dyer, R.B.; Camilleri, M.; Kashyap, P.C.; et al. Sex Differences in NSAID-Induced Perturbation of Human Intestinal Barrier Function and Microbiota. FASEB J. 2018, 32, 6615–6625. [Google Scholar] [CrossRef]
- Prizment, A.E.; Staley, C.; Onyeaghala, G.C.; Vivek, S.; Thyagarajan, B.; Straka, R.J.; Demmer, R.T.; Knights, D.; Meyer, K.A.; Shaukat, A.; et al. Randomised Clinical Study: Oral Aspirin 325 Mg Daily vs Placebo Alters Gut Microbial Composition and Bacterial Taxa Associated with Colorectal Cancer Risk. Aliment. Pharmacol. Ther. 2020, 52, 976–987. [Google Scholar] [CrossRef] [PubMed]
- Zhernakova, A.; Kurilshikov, A.; Bonder, M.J.; Tigchelaar, E.F.; Schirmer, M.; Vatanen, T.; Mujagic, Z.; Vila, A.V.; Falony, G.; Vieira-Silva, S.; et al. Population-Based Metagenomics Analysis Reveals Markers for Gut Microbiome Composition and Diversity. Science 2016, 352, 565–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Sun, Y.; Wang, R.; Zhang, J. Gut Microbiota-Mediated Drug-Drug Interaction between Amoxicillin and Aspirin. Sci. Rep. 2019, 9, 16194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Bittinger, K.; Li, X.; Abernethy, D.R.; Bushman, F.D.; FitzGerald, G.A. Bidirectional Interactions between Indomethacin and the Murine Intestinal Microbiota. Elife 2015, 4, e08973. [Google Scholar] [CrossRef]
- Martin, R.M.; Dunn, N.R.; Freemantle, S.; Shakir, S. The Rates of Common Adverse Events Reported during Treatment with Proton Pump Inhibitors Used in General Practice in England: Cohort Studies. Br. J. Clin. Pharmacol. 2000, 50, 366–372. [Google Scholar] [CrossRef] [Green Version]
- Bruno, G.; Zaccari, P.; Rocco, G.; Scalese, G.; Panetta, C.; Porowska, B.; Pontone, S.; Severi, C. Proton Pump Inhibitors and Dysbiosis: Current Knowledge and Aspects to Be Clarified. World J. Gastroenterol. 2019, 25, 2706–2719. [Google Scholar] [CrossRef]
- Amir, I.; Konikoff, F.M.; Oppenheim, M.; Gophna, U.; Half, E.E. Gastric Microbiota Is Altered in Oesophagitis and Barrett’s Oesophagus and Further Modified by Proton Pump Inhibitors. Environ. Microbiol. 2014, 16, 2905–2914. [Google Scholar] [CrossRef]
- Scarpignato, C.; Gatta, L.; Zullo, A.; Blandizzi, C. Effective and Safe Proton Pump Inhibitor Therapy in Acid-Related Diseases—A Position Paper Addressing Benefits and Potential Harms of Acid Suppression. BMC Med. 2016, 14, 179. [Google Scholar] [CrossRef] [PubMed]
- Nardelli, S.; Gioia, S.; Ridola, L.; Farcomeni, A.; Merli, M.; Riggio, O. Proton Pump Inhibitors Are Associated With Minimal and Overt Hepatic Encephalopathy and Increased Mortality in Patients with Cirrhosis. Hepatology 2019, 70, 640–649. [Google Scholar] [CrossRef] [PubMed]
- Marlicz, W.; Loniewski, I.; Grimes, D.S.; Quigley, E.M. Nonsteroidal Anti-Inflammatory Drugs, Proton Pump Inhibitors, and Gastrointestinal Injury: Contrasting Interactions in the Stomach and Small Intestine. Mayo Clin. Proc. 2014, 89, 1699–1709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marlicz, W.; Loniewski, I.; Koulaouzidis, G. Proton Pump Inhibitors, Dual Antiplatelet Therapy, and the Risk of Gastrointestinal Bleeding. Mayo Clin. Proc. 2022, 97, 648–651. [Google Scholar] [CrossRef]
- Janarthanan, S.; Ditah, I.; Adler, D.G.; Ehrinpreis, M.N. Clostridium Difficile-Associated Diarrhea and Proton Pump Inhibitor Therapy: A Meta-Analysis. Am. J. Gastroenterol. 2012, 107, 1001–1010. [Google Scholar] [CrossRef]
- Schmulson, M.J.; Frati-Munari, A.C. Bowel Symptoms in Patients That Receive Proton Pump Inhibitors. Results of a Multicenter Survey in Mexico. Rev. Gastroenterol. Mex. 2019, 84, 44–51. [Google Scholar] [CrossRef]
- Yang, Y.-X.; Hennessy, S.; Propert, K.; Hwang, W.-T.; Sedarat, A.; Lewis, J.D. Chronic Proton Pump Inhibitor Therapy and the Risk of Colorectal Cancer. Gastroenterology 2007, 133, 748–754. [Google Scholar] [CrossRef]
- Davis, J.A.; Collier, F.; Mohebbi, M.; Stuart, A.L.; Loughman, A.; Pasco, J.A.; Jacka, F.N. Obesity, Akkermansia Muciniphila, and Proton Pump Inhibitors: Is There a Link? Obes. Res. Clin. Pract. 2020, 14, 524–530. [Google Scholar] [CrossRef]
- Imhann, F.; Bonder, M.J.; Vich Vila, A.; Fu, J.; Mujagic, Z.; Vork, L.; Tigchelaar, E.F.; Jankipersadsing, S.A.; Cenit, M.C.; Harmsen, H.J.M.; et al. Proton Pump Inhibitors Affect the Gut Microbiome. Gut 2016, 65, 740–748. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.A.; Goodrich, J.K.; Maxan, M.-E.; Freedberg, D.E.; Abrams, J.A.; Poole, A.C.; Sutter, J.L.; Welter, D.; Ley, R.E.; Bell, J.T.; et al. Proton Pump Inhibitors Alter the Composition of the Gut Microbiota. Gut 2016, 65, 749–756. [Google Scholar] [CrossRef] [Green Version]
- Cai, T.; Abel, L.; Langford, O.; Monaghan, G.; Aronson, J.K.; Stevens, R.J.; Lay-Flurrie, S.; Koshiaris, C.; McManus, R.J.; Hobbs, F.D.R.; et al. Associations between Statins and Adverse Events in Primary Prevention of Cardiovascular Disease: Systematic Review with Pairwise, Network, and Dose-Response Meta-Analyses. BMJ 2021, 374, n1537. [Google Scholar] [CrossRef] [PubMed]
- Newman, C.B.; Preiss, D.; Tobert, J.A.; Jacobson, T.A.; Page, R.L.; Goldstein, L.B.; Chin, C.; Tannock, L.R.; Miller, M.; Raghuveer, G.; et al. Statin Safety and Associated Adverse Events: A Scientific Statement from the American Heart Association. Arterioscler. Thromb. Vasc. Biol. 2019, 39, e38–e81. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Li, H.; Zhao, X.; Zhou, R.; Liu, H.; Sun, Y.; Fan, Y.; Shi, Y.; Qiao, S.; Liu, S.; et al. Multi-Omics Study Reveals That Statin Therapy Is Associated with Restoration of Gut Microbiota Homeostasis and Improvement in Outcomes in Patients with Acute Coronary Syndrome. Theranostics 2021, 11, 5778–5793. [Google Scholar] [CrossRef]
- Ko, H.H.T.; Lareu, R.R.; Dix, B.R.; Hughes, J.D. Statins: Antimicrobial Resistance Breakers or Makers? PeerJ 2017, 5, e3952. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Lee, H.; An, J.; Song, Y.; Lee, C.-K.; Kim, K.; Kong, H. Alterations in Gut Microbiota by Statin Therapy and Possible Intermediate Effects on Hyperglycemia and Hyperlipidemia. Front. Microbiol. 2019, 10, 1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Li, H.; Yuan, L.; Zhang, J.; Han, L.; Liu, R.; Wu, R.; Shi, Y.; Mushtaq, N.; Ullah, S.; et al. Molecular Characterization of Gut Microbiota in High-lipid Diet-induced Hyperlipidemic Rats Treated with Simvastatin. Int. J. Mol. Med. 2020, 45, 1601–1615. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Garcia, U.; Jebari, S.; Larrea-Sebal, A.; Uribe, K.B.; Siddiqi, H.; Ostolaza, H.; Benito-Vicente, A.; Martín, C. Statin Treatment-Induced Development of Type 2 Diabetes: From Clinical Evidence to Mechanistic Insights. Int. J. Mol. Sci. 2020, 21, 4725. [Google Scholar] [CrossRef]
- Climent, E.; Benaiges, D.; Pedro-Botet, J. Statin Treatment and Increased Diabetes Risk. Possible Mechanisms. Clin. Investig. Arterioscler. 2019, 31, 228–232. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, W.; Guo, M.; Hua, Y.; Zhou, B.; Li, X.; Zhang, X.; Dong, J.; Yang, X.; Wang, Y.; et al. The Gut Microbiota Is Associated with Clinical Response to Statin Treatment in Patients with Coronary Artery Disease. Atherosclerosis 2021, 325, 16–23. [Google Scholar] [CrossRef]
- Zhao, C.; Hu, Y.; Chen, H.; Li, B.; Cao, L.; Xia, J.; Yin, Y. An in Vitro Evaluation of the Effects of Different Statins on the Structure and Function of Human Gut Bacterial Community. PLoS ONE 2020, 15, e0230200. [Google Scholar] [CrossRef]
- Nolan, J.A.; Skuse, P.; Govindarajan, K.; Patterson, E.; Konstantinidou, N.; Casey, P.G.; MacSharry, J.; Shanahan, F.; Stanton, C.; Hill, C.; et al. The Influence of Rosuvastatin on the Gastrointestinal Microbiota and Host Gene Expression Profiles. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G488–G497. [Google Scholar] [CrossRef]
- Tian, H.; Ge, X.; Nie, Y.; Yang, L.; Ding, C.; McFarland, L.V.; Zhang, X.; Chen, Q.; Gong, J.; Li, N. Fecal Microbiota Transplantation in Patients with Slow-Transit Constipation: A Randomized, Clinical Trial. PLoS ONE 2017, 12, e0171308. [Google Scholar] [CrossRef] [Green Version]
- Chu, J.R.; Kang, S.-Y.; Kim, S.-E.; Lee, S.-J.; Lee, Y.-C.; Sung, M.-K. Prebiotic UG1601 Mitigates Constipation-Related Events in Association with Gut Microbiota: A Randomized Placebo-Controlled Intervention Study. World J. Gastroenterol. 2019, 25, 6129–6144. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Wu, S.; Ji, L.; Fan, Y.; Wang, X.; Yang, K. The Combined Therapy of Fecal Microbiota Transplantation and Laxatives for Functional Constipation in Adults: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Medicine 2021, 100, e25390. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.; Takahara, C.; Tabuchi, N.; Okamura, N. Daiokanzoto (Da-Huang-Gan-Cao-Tang) Is an Effective Laxative in Gut Microbiota Associated with Constipation. Sci. Rep. 2019, 9, 3833. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zheng, J.; Jiang, N.; Sun, G.; Bao, X.; Kong, M.; Cheng, X.; Lin, A.; Liu, H. Modulation of Gut Microbiota and Intestinal Metabolites by Lactulose Improves Loperamide-Induced Constipation in Mice. Eur. J. Pharm. Sci. 2021, 158, 105676. [Google Scholar] [CrossRef] [PubMed]
- Tomkovich, S.; Taylor, A.; King, J.; Colovas, J.; Bishop, L.; McBride, K.; Royzenblat, S.; Lesniak, N.A.; Bergin, I.L.; Schloss, P.D. An Osmotic Laxative Renders Mice Susceptible to Prolonged Clostridioides Difficile Colonization and Hinders Clearance. mSphere 2019, 6, e00629-21. [Google Scholar] [CrossRef]
- Matsuura, T.; Kyokane, K.; Yamada, S.; Kuno, Y. The Development of the Cure of the Functional Intestinal Disorder Based on the Differences of Gut Microbiota in Aged Patients. Medicine 2021, 100, e27696. [Google Scholar] [CrossRef]
- Cheung, M.K.; Yue, G.G.L.; Chiu, P.W.Y.; Lau, C.B.S. A Review of the Effects of Natural Compounds, Medicinal Plants, and Mushrooms on the Gut Microbiota in Colitis and Cancer. Front. Pharmacol. 2020, 11, 744. [Google Scholar] [CrossRef]
- Lyu, M.; Wang, Y.; Fan, G.; Wang, X.; Xu, S.; Zhu, Y. Balancing Herbal Medicine and Functional Food for Prevention and Treatment of Cardiometabolic Diseases through Modulating Gut Microbiota. Front. Microbiol. 2017, 8, 2146. [Google Scholar] [CrossRef] [Green Version]
- Pferschy-Wenzig, E.-M.; Pausan, M.R.; Ardjomand-Woelkart, K.; Röck, S.; Ammar, R.M.; Kelber, O.; Moissl-Eichinger, C.; Bauer, R. Medicinal Plants and Their Impact on the Gut Microbiome in Mental Health: A Systematic Review. Nutrients 2022, 14, 2111. [Google Scholar] [CrossRef] [PubMed]
- Chu, N.; Chan, J.C.N.; Chow, E. Pharmacomicrobiomics in Western Medicine and Traditional Chinese Medicine in Type 2 Diabetes. Front. Endocrinol. 2022, 13, 857090. [Google Scholar] [CrossRef] [PubMed]
- Panyod, S.; Wu, W.-K.; Chen, C.-C.; Wu, M.-S.; Ho, C.-T.; Sheen, L.-Y. Modulation of Gut Microbiota by Foods and Herbs to Prevent Cardiovascular Diseases. J. Tradit. Complement. Med. 2021, 13, 107–118. [Google Scholar] [CrossRef]
- Chen, Z.; Lv, Y.; Xu, H.; Deng, L. Herbal Medicine, Gut Microbiota, and COVID-19. Front. Pharmacol. 2021, 12, 646560. [Google Scholar] [CrossRef] [PubMed]
- Martínez, Y.; Más, D. Role of Herbs and Medicinal Spices as Modulators of Gut Microbiota. In Herbs and Spices; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef] [Green Version]
- An, X.; Bao, Q.; Di, S.; Zhao, Y.; Zhao, S.; Zhang, H.; Lian, F.; Tong, X. The Interaction between the Gut Microbiota and Herbal Medicines. Biomed. Pharmacother. 2019, 118, 109252. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Kuo, H.-C.; Hudlikar, R.; Sargsyan, D.; Li, S.; Wang, L.; Wu, R.; Kong, A.-N. Gut Microbiota, Dietary Phytochemicals and Benefits to Human Health. Curr. Pharmacol. Rep. 2019, 5, 332–344. [Google Scholar] [CrossRef]
- Dingeo, G.; Brito, A.; Samouda, H.; Iddir, M.; La Frano, M.R.; Bohn, T. Phytochemicals as Modifiers of Gut Microbial Communities. Food Funct. 2020, 11, 8444–8471. [Google Scholar] [CrossRef]
- Kumar Singh, A.; Cabral, C.; Kumar, R.; Ganguly, R.; Kumar Rana, H.; Gupta, A.; Rosaria Lauro, M.; Carbone, C.; Reis, F.; Pandey, A.K. Beneficial Effects of Dietary Polyphenols on Gut Microbiota and Strategies to Improve Delivery Efficiency. Nutrients 2019, 11, 2216. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Gao, M.; Kang, G.; Huang, H. The Potential Role of Phytonutrients Flavonoids Influencing Gut Microbiota in the Prophylaxis and Treatment of Inflammatory Bowel Disease. Front. Nutr. 2021, 8, 798038. [Google Scholar] [CrossRef]
- Medhi, J.; Kalita, M.C. Nut Phytonutrients for Healthy Gut: Prebiotic Potential; IntechOpen: London, UK, 2020; ISBN 978-1-78985-511-1. [Google Scholar]


{kind=link}
| Phytochemicals | Source | Part of Plant | Research Model | Effect on Bacterial Counts | |
|---|---|---|---|---|---|
| Increase | Decrease | ||||
| phenolic acids, stilbenes, flavonols, dihydroflavonols, anthocyanins | grapevine (Vitis vinifera) | fruits | Human | Enterococcus spp., Prevotella spp., Bifidobacterium spp., Bacteroides uniformis, Eggerthella lenta, Blautia coccoides, Eubacterium rectale | |
| chlorogenic acid | coffee (Coffea arabica) | green beans | animal (mice) | Bacteroides, Bifidobacterium, Lactobacillus, Ruminococcaceae | Desulfovibrionaceae, Lachnospiraceae, Erysipelotrichaceae |
| caffeine | coffee (Coffea arabica) | roasted beans | human | Bifidobacterium spp. Faecalibacterium, Roseburia | Erysipelatoclostridium |
| phenolic compounds | garlic (Allium sativum) | whole plant | in vitro | Staphylloccocus epidermidis, Klebsiella pneumoniae | |
| allicin | garlic (Allium sativum) | bulbs | animal (mice) | Akkermansia spp. | |
| Tannins | moringa (Moringa oleifera) | leaves | animal (mice) | Bacteroides | Clostridium leptum |
| anthocyanidins, flavonoids | pomegranate (Punica granatum) | fruits | animal (mice) | Lactobacillus spp., Bifidobacterium | Escherichia coli, Salmonella spp. |
| Anthocyanidins | shrubby blackberry (Rubus fruticosus), raspberry (Rubus occidentalis) | fruits | in vitro | Helicobacter pylori, Staphylococcus epidermis, Klebsiella pneumoniae, Escherichia coli, Salmonella spp. | |
| Garden snapdragon (Antirrhinum majus) | flowers | animal (mice) | Bifidobacterium spp., Lactobacillus spp. | ||
| Catechins, flavanols | green tea (Camelia sinensis) | leaves | human | Lactobacillus spp. | Streptococcus mutans, Shigella, Vibrio cholera, Clostridium histolyticum, Clostridium coccoides, Eubacterium rectale |
| procyanidins, catechins, epicatechins | apple (Pyrus malus) | fruits | animal (mice) | Bifidobacterium spp. | |
| Coumarins | cashew (Anacardium occidentale) | leaves | in vitro | Staphylococcus aureus | |
| sulforaphane | brassicaceous vegetables (e.g., radish, cabbages, cauliflower, broccoli) | edible parts | animal (mice) | Bacteroides fragilis, Clostridium cluster | |
| polyphenols | black currant (Ribes nigrum) | leaves and fruits | human | Lactobacilli, Bifidobacteria | Clostridium spp., Bacteroides spp. |
| Quercetin, rutin, (or buckwheat supplementation) | buckwheat (Fagopyrum esculentum) | seeds | human | Eubacterium ramulus | |
| flavanol | cocoa (Theobroma cacao) | fruits | human | Bifidobacterium spp., Lactobacillus spp. | Clostridium spp. |
| Polyphenols | red wine | grapes | Enterococcus, Prevotella, Bacteroides, Bifidobacterium, Bacteroides uniformis, Eggerthella lenta, Blautia coccoides, Eubacterium rectale | ||
| β-carotene | isolated pure chemical compound | human | Firmicutes | Bacteroides | |
| lycopene | isolated pure chemical compound | human | Bifidobacterium adolescentis, Bifidobacterium longum | Bacillus subtilis | |
| terpenes, aroma active compounds | rosemary (Rosmarinus officinalis) | herbaceous | animal (rats) | Bifidobacterium spp., Blautia coccoides, Bacteroides spp. | Lactobacillus, Leuconostoc, Pediococcus, Clostridium spp. |
| Source | Research Model | Effect on Bacterial Counts | |
|---|---|---|---|
| Increase | Decrease | ||
| defatted almonds | in vitro | Bifidobacteria, Eubacterium rectale | |
| almonds (raw and roasted) | in vitro | Lactobacillus acidophilus, Bifidobacterium breve | Escherichia coli |
| animal (rats) | Lactobacillus spp., Bifidobacterium ssp. | Enterococcus spp. Escherichia coli | |
| chestnut | in vitro | Lactobacillus paracasei GG, Lactobacillus rhamnosus, Lactobacillus casei | Streptococcus macedonicus Streptococcus thermophilus |
| walnuts | animal (rats) | Lactobacillus, Ruminococcaceae, Roseburia | Bacteroides Anaerotruncus Alphaproteobacteria |
| human | butyrate-producing bacteria, Ruminococcaceae, Bifidobacteria | Clostridium spp. | |
| roasted almond, almond skin | human | Lactobacillus spp., Bifidobacterium spp. | Clostridium perfringens |
| whole almonds, roasted almonds, roasted chopped almonds, almond butter | human | Lachnospira Roseburia Oscillospira Dialister | |
| Drug Class | Substance | Increase | Decrease |
|---|---|---|---|
| Hypoglycaemic drugs | metformin | Akkermansia muciniphila, Butyrivibrio, Bifidobacterium bifidum, Megasphaera, Escherichia coli, Shigella spp. Klebsiella, Citrobacter, Enterobacter cloacae, Salmonella enterica, Prevotella, Megasphaera, | Intestinibacter, Clostridium spp., Clostridium bartlettii, Mobiluncus mulieris, Peptoniphilus duerdenii, Eubacterium siraeum, Oscillospira, Barnesiellaceae Actinobacteria, Bacillus |
| NSAIDs | acetylsalicylic acid | Prevotella, Bacteroides, Barnesiella, Ruminococcaceae | |
| celecoxib, ibuprofen | Acidaminococcaceae, Enterobacteriaceae Propionibacteriaceae, Pseudomonadaceae, Puniceicoccaceae, Rikenellaceae, Enterococcaceae and Erysipelotrichaceae | ||
| celecoxib | Enterococcaceae and Erysipelotrichaceae | Firmicutes, Ruminococcus | |
| PPIs | Fusobacterium, Leptotrichia, Neisseria, Veillonella, Firmicutes, Streptococcaceae, Prevotellaceae, Campylobacteraceae, Enterococcus, Streptococcus, Staphylococcus | Bacteroidetes, Proteobacteria | |
| Statins | Atorvastatin, rosuvastatin | Bifidobacterium longum subsp. longum, Anaerostipes hadrus and Ruminococcus obeum Bacteroides, Butyricimonas and Mucispirillum, Holdemanella, Faecalibacterium | Akkermansia muciniphila, Lactobacillus |
| fluvastatin | Escherichia/Shigella, Ruminococcaceae UCG 014, Sutterella | ||
| Laxatives | 15% polyethylene glycol (PEG) | C. difficile |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mruk-Mazurkiewicz, H.; Kulaszyńska, M.; Jakubczyk, K.; Janda-Milczarek, K.; Czarnecka, W.; Rębacz-Maron, E.; Zacha, S.; Sieńko, J.; Zeair, S.; Dalewski, B.; et al. Clinical Relevance of Gut Microbiota Alterations under the Influence of Selected Drugs—Updated Review. Biomedicines 2023, 11, 952. https://doi.org/10.3390/biomedicines11030952
Mruk-Mazurkiewicz H, Kulaszyńska M, Jakubczyk K, Janda-Milczarek K, Czarnecka W, Rębacz-Maron E, Zacha S, Sieńko J, Zeair S, Dalewski B, et al. Clinical Relevance of Gut Microbiota Alterations under the Influence of Selected Drugs—Updated Review. Biomedicines. 2023; 11(3):952. https://doi.org/10.3390/biomedicines11030952
Chicago/Turabian StyleMruk-Mazurkiewicz, Honorata, Monika Kulaszyńska, Karolina Jakubczyk, Katarzyna Janda-Milczarek, Wiktoria Czarnecka, Ewa Rębacz-Maron, Sławomir Zacha, Jerzy Sieńko, Samir Zeair, Bartosz Dalewski, and et al. 2023. "Clinical Relevance of Gut Microbiota Alterations under the Influence of Selected Drugs—Updated Review" Biomedicines 11, no. 3: 952. https://doi.org/10.3390/biomedicines11030952