
Chlorogenic Acid Restores Ovarian Functions in Mice with Letrozole-Induced Polycystic Ovarian Syndrome Via Modulation of Adiponectin Receptor

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Methods and Materials
2.1. Chemicals
2.2. Animals
2.3. Biochemical Studies
Antioxidant Assay
2.4. Plasma Adiponectin
2.5. Inflammatory Cytokines
2.6. Reverse Transcription and Real-Time PCR
2.7. Histological Assessment of Ovaries
2.8. Statistical Analysis
2.9. Bioinformatics Analysis of Adiponectin
2.9.1. Protein–Protein Interaction of Adiponectin
2.9.2. Gene Ontology and Pathway Analysis
3. Results
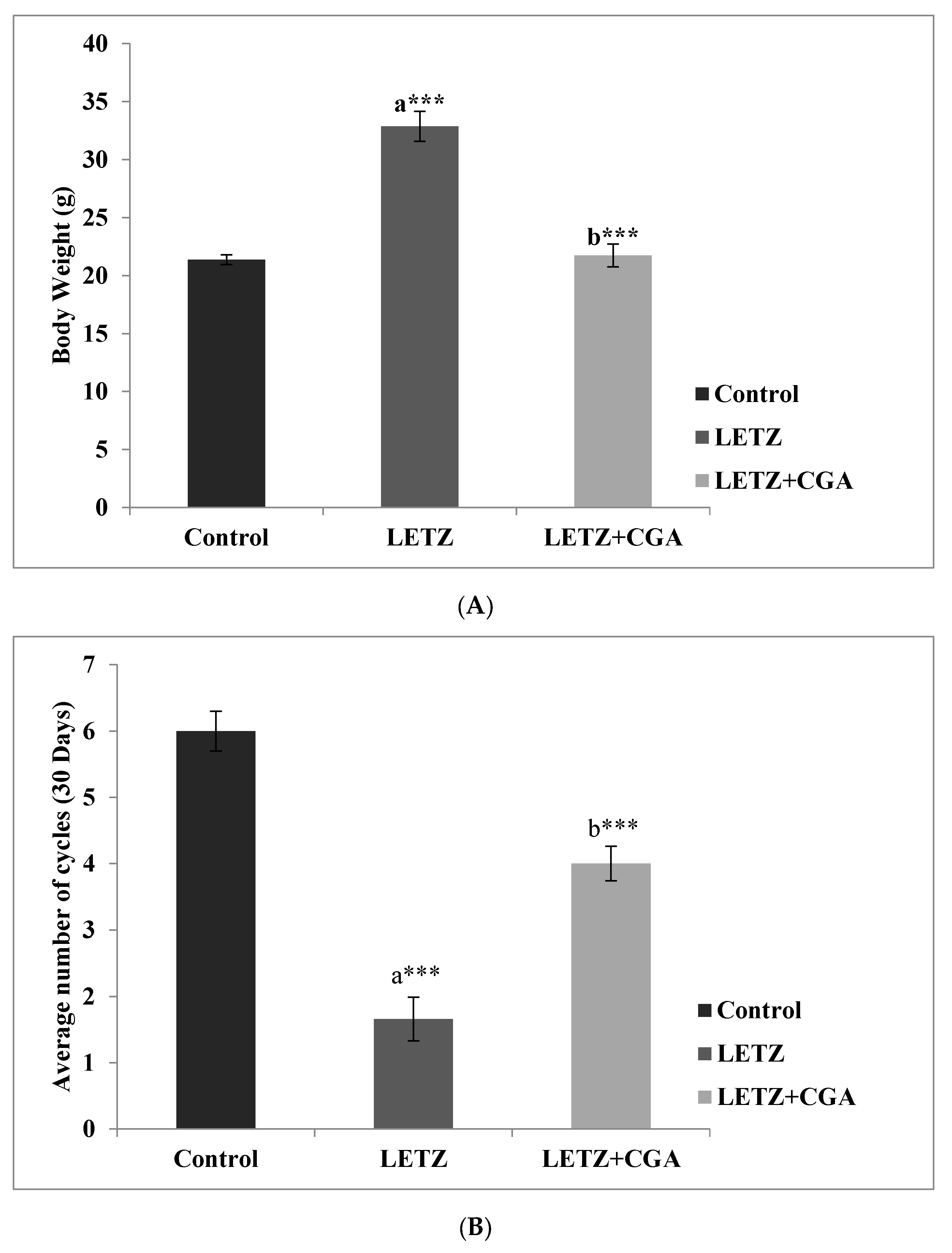
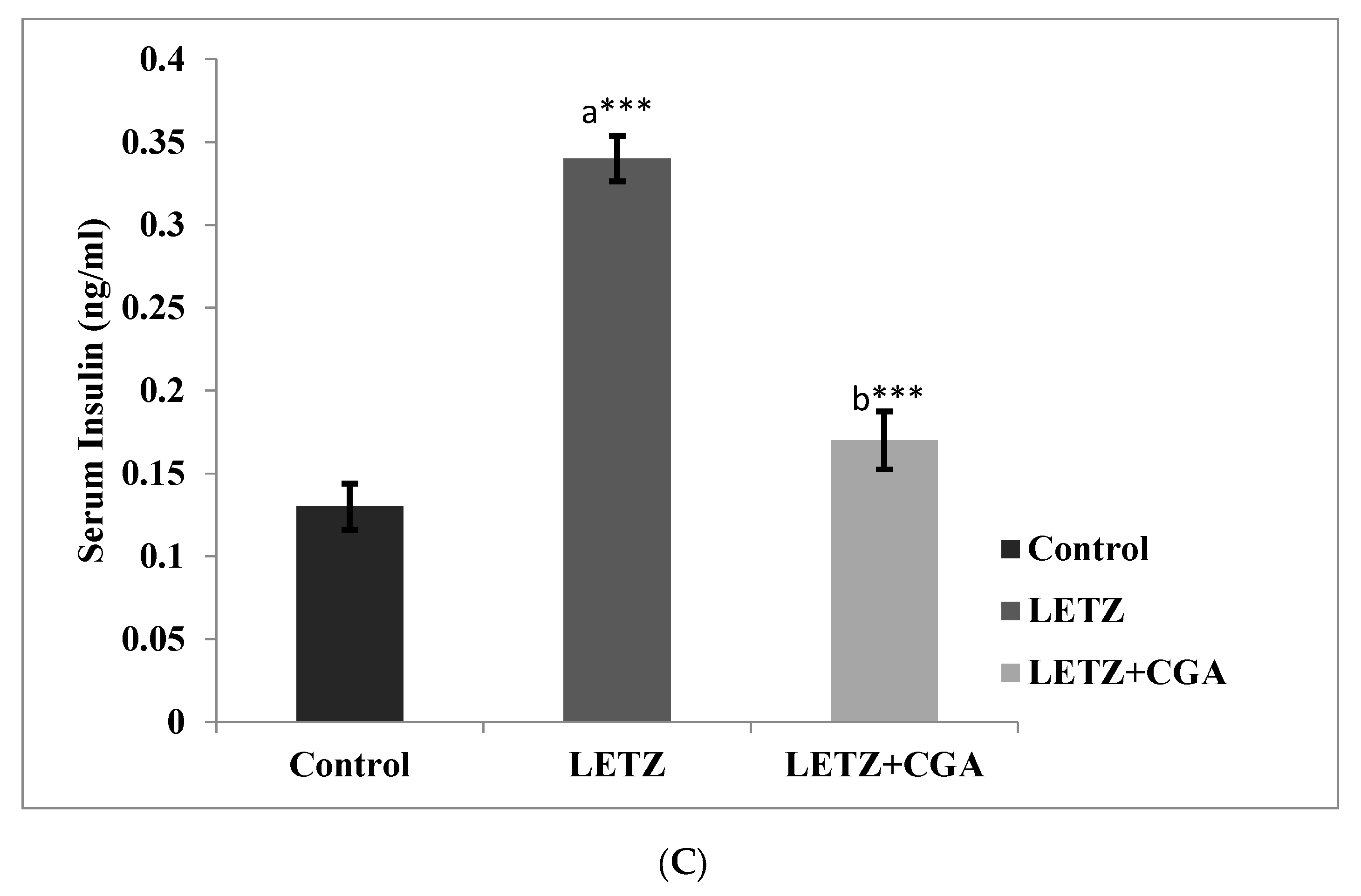
3.1. Effect of Chlorogenic Acid (CGA) on Body Weight, Fasting Blood Glucose, Insulin Concentration, and Changes in Estrous Cyclicity in LETZ-Induced PCOS Mice
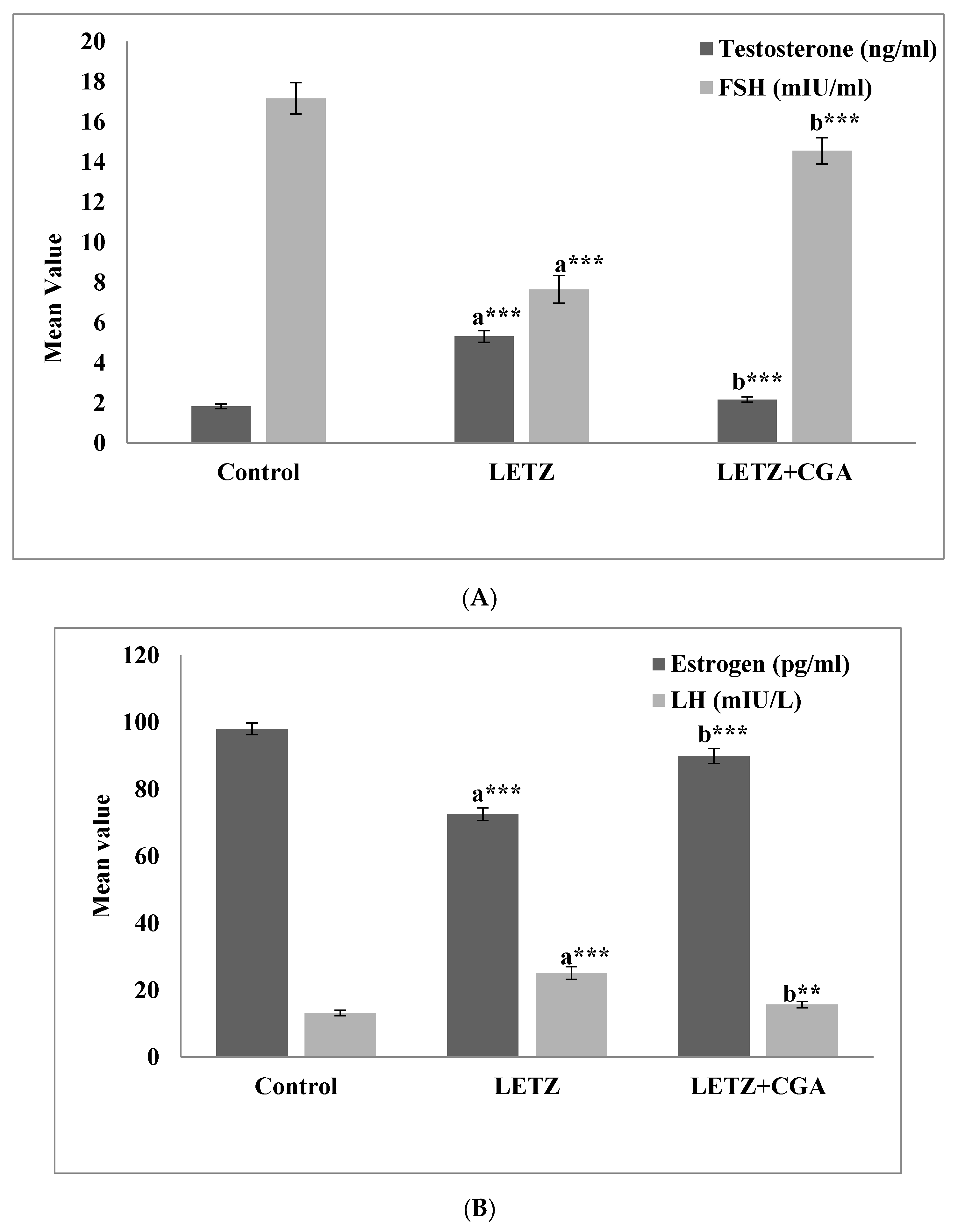
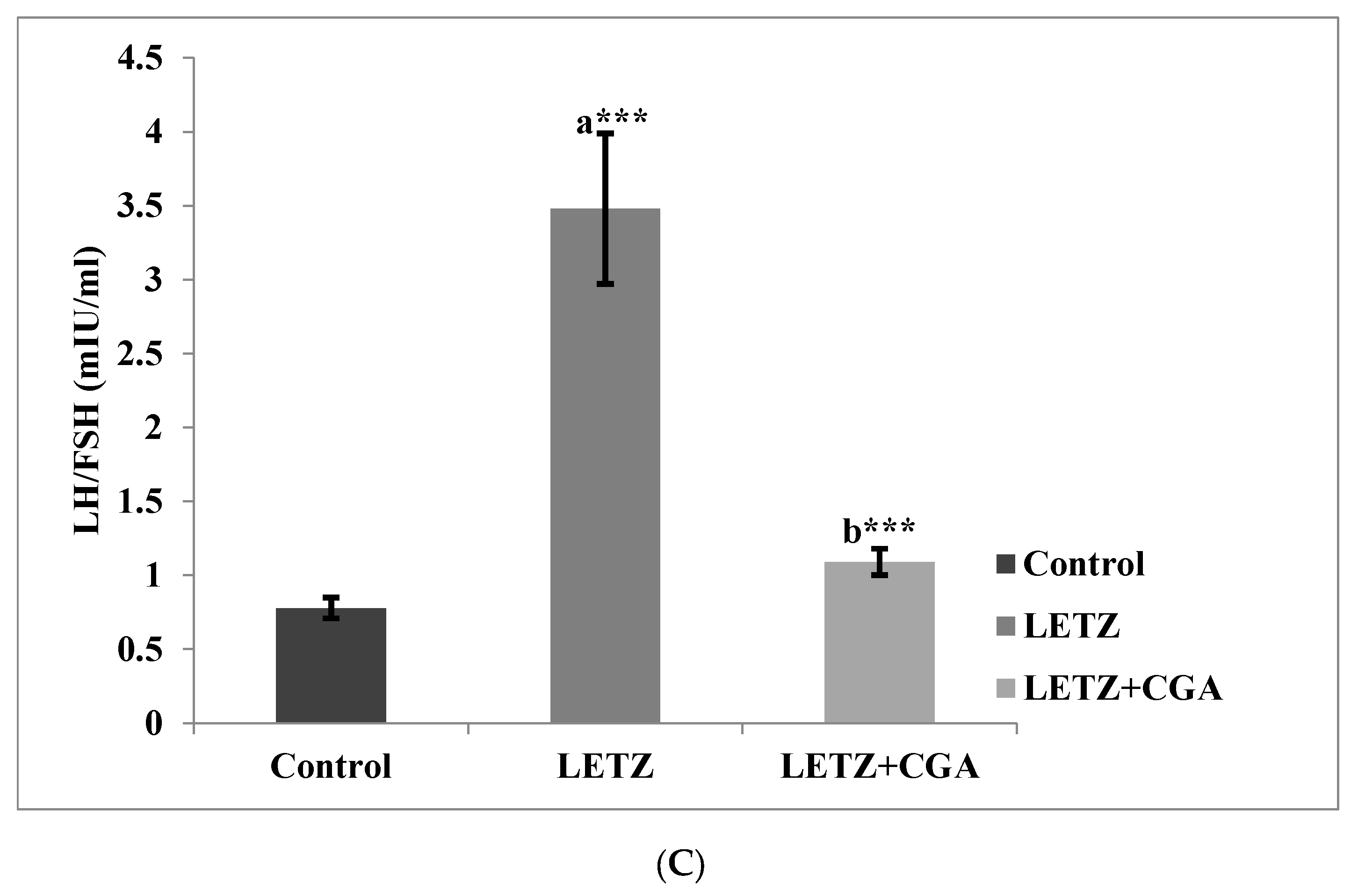
3.2. Effect of CGA on Hormone Levels in Mice with PCOS
3.3. Impact of CGA Treatment on Lipid Profiles in PCOS Mice
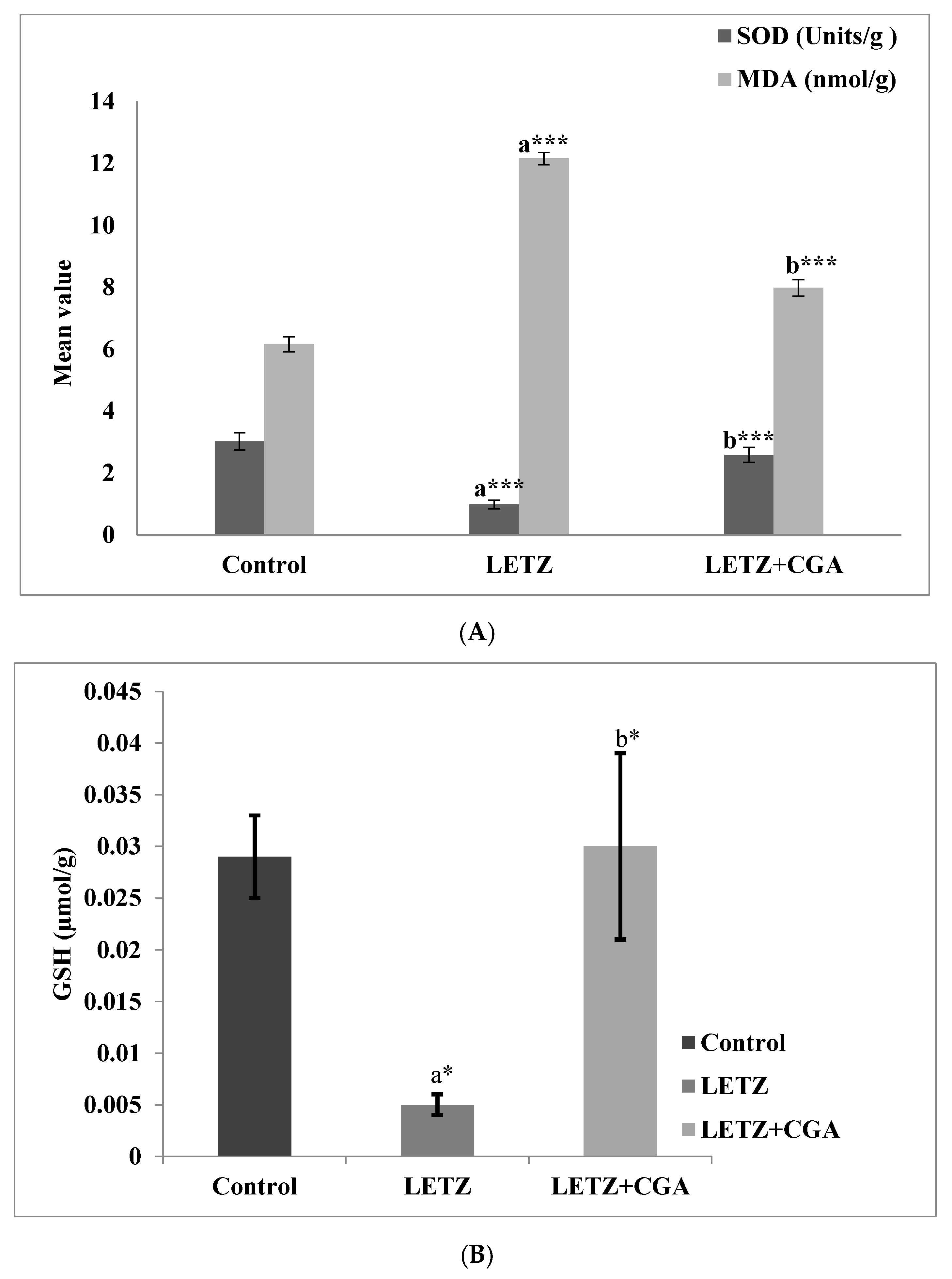
3.4. Impact of CGA on Lipid Peroxidation of Antioxidants
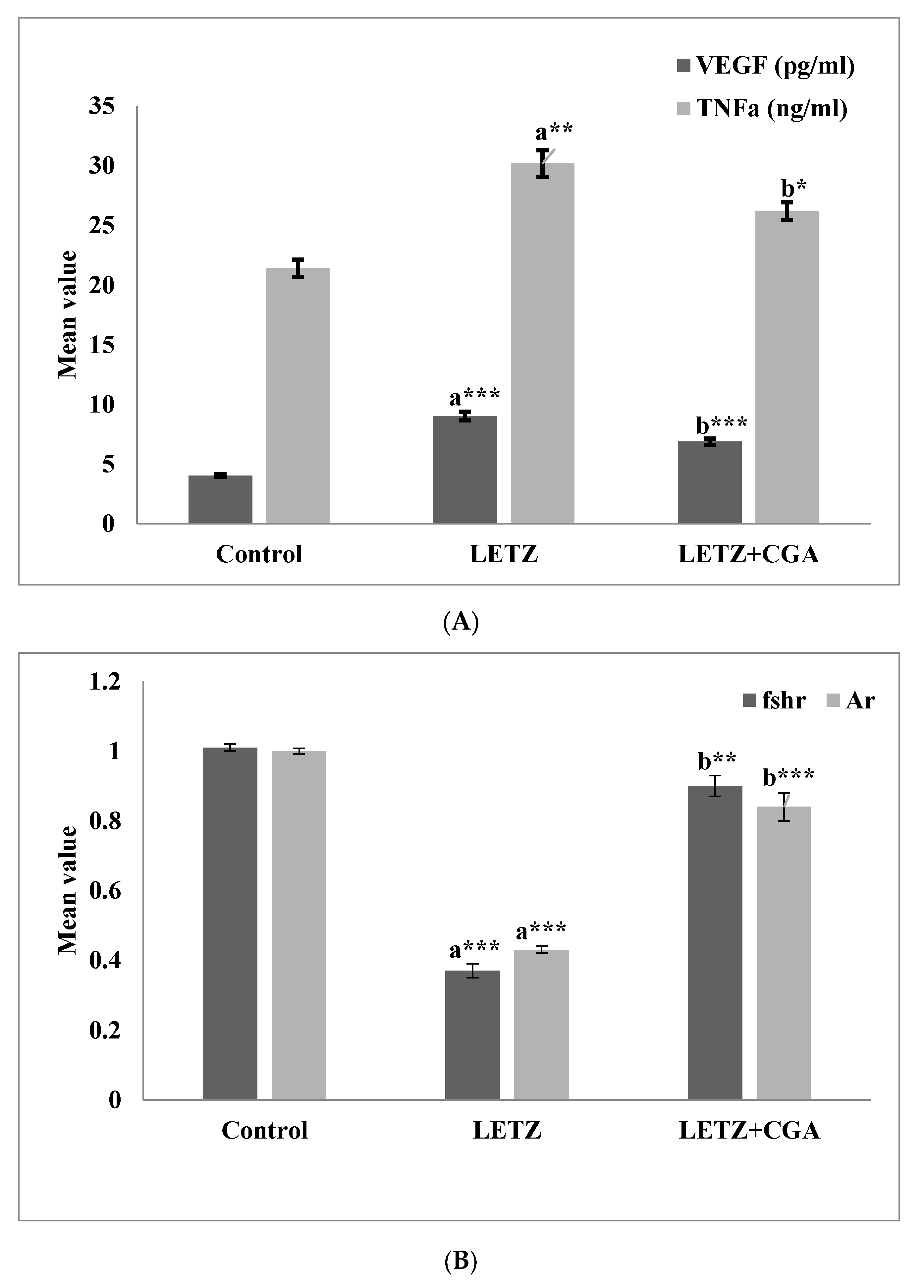
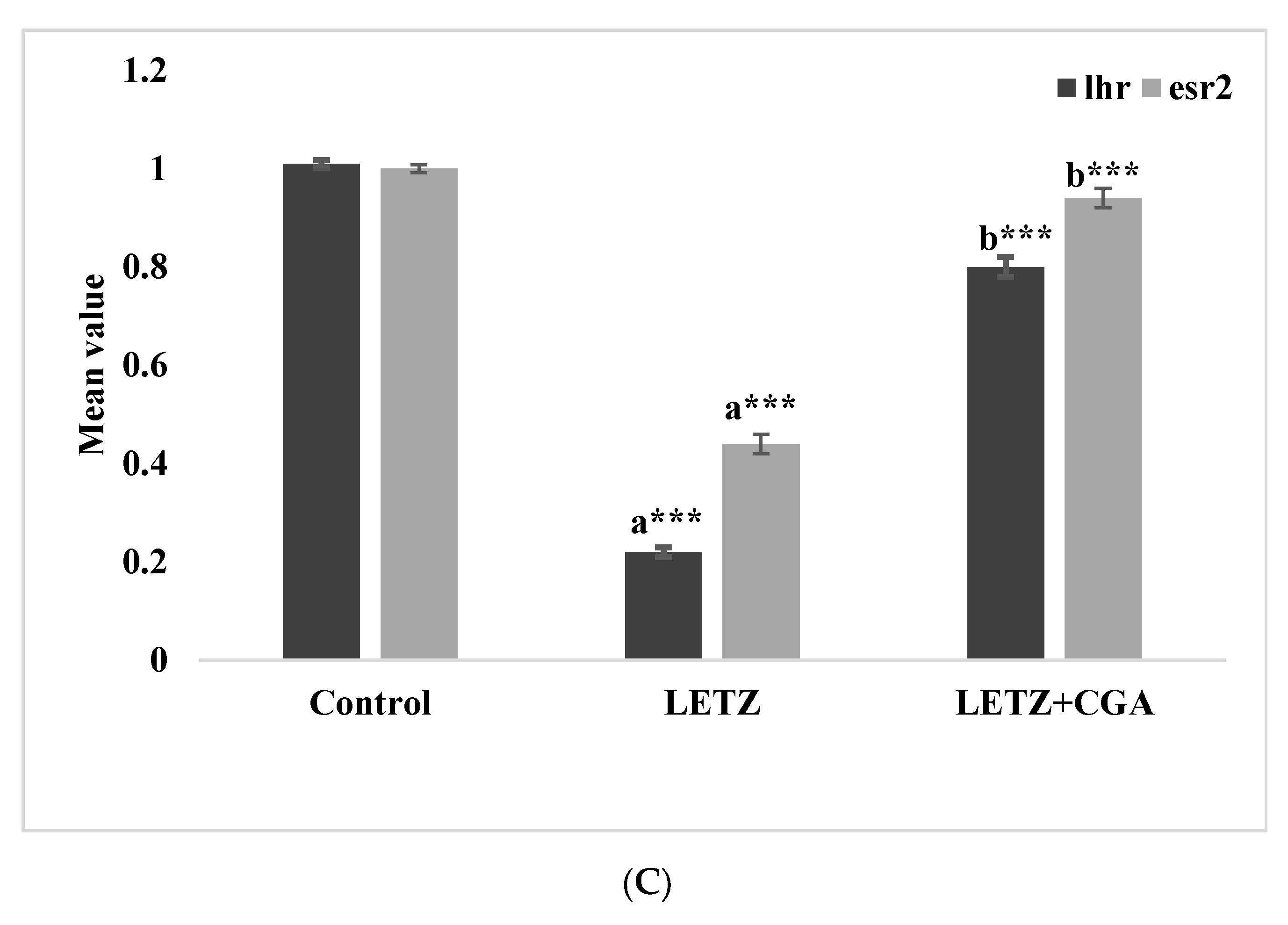
3.5. CGA Administration Treatment Reduces Inflammatory Cytokines in PCOS Mice and Effects the Expressions of Genes Related to Hormones
3.6. CGA Treatment Increases the Levels of Adiponectin and Adipo-R1 Expression in PCOS Mice
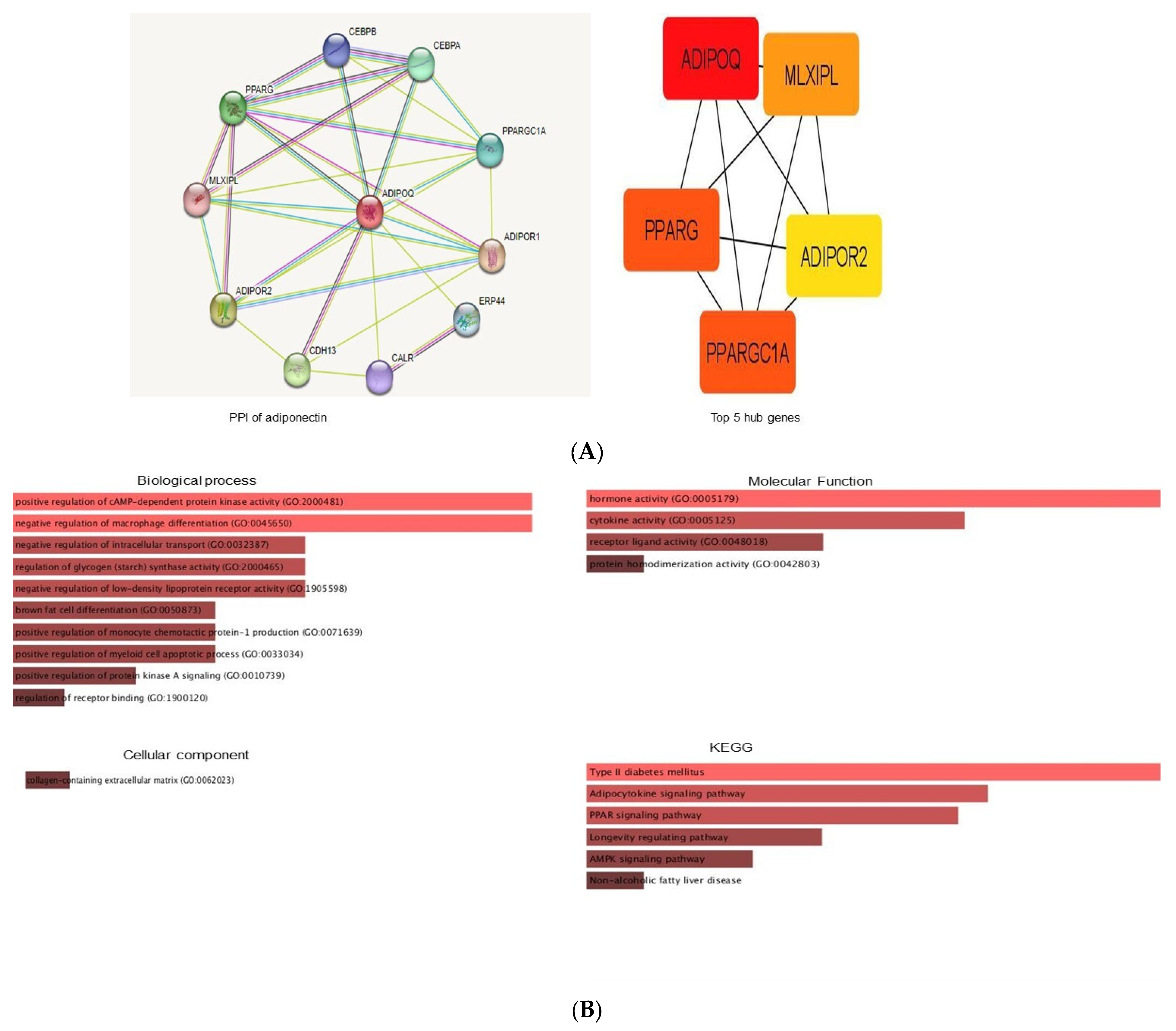
3.7. Protein–Protein Interactions of Adiponectin
3.8. Gene Ontology and KEGG Pathway Analysis
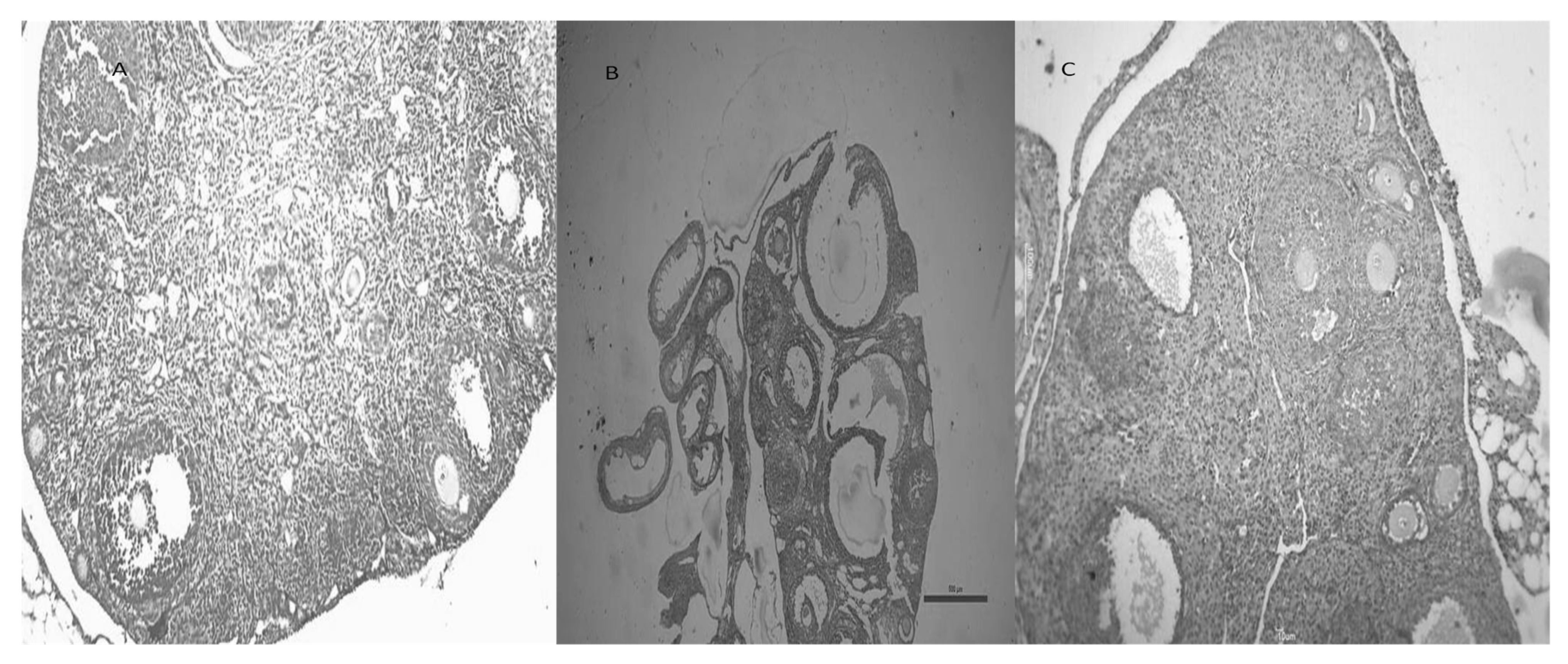
3.9. CGA Improves Ovarian Morphology in Mice with Letrozole-Induced PCOS
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brakta, S.; Lizneva, D.; Mykhalchenko, K.; Imam, A.; Walker, W.; Diamond, M.P.; Azziz, R. Perspectives on polycystic ovary syndrome: Is polycystic ovary syndrome research underfunded? J. Clin. Endocrinol. Metab. 2017, 102, 4421–4427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosiek, A.; Frąckowiak Maciejewska, N.; Leksowski, K.; Rosiek-Kryszewska, A.; Leksowski, Ł. Effect of television on obesity and excess of weight and consequences of health. Int. J. Environ. Res. Public Health 2015, 12, 9408–9426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmina, E. Diagnosis of polycystic ovary syndrome: From NIH criteria to ESHRE-ASRM guidelines. Minerva Ginecol. 2004, 56, 1–6. [Google Scholar] [PubMed]
- Zafar, U.; Memon, Z.; Moin, K.; Agha, S.; Hassan, J.A.; Zehra, D. Prevalence of PCOS with associated symptoms and complications at tertiary care hospital of Karachi. J. Adv. Med. Med. Res. 2019, 30, 1–9. [Google Scholar] [CrossRef]
- Gambineri, A.; Pelusi, C.; Vicennati, V.; Pagotto, U.; Pasquali, R. Obesity and the polycystic ovary syndrome. Int. J. Obes. 2002, 26, 883–896. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.S.; Davies, M.J.; Norman, R.J.; Moran, L.J. Overweight, obesity and central obesity in women with polycystic ovary syndrome: A systematic review and meta-analysis. Hum. Reprod. Update 2012, 18, 618–637. [Google Scholar] [CrossRef]
- Ul Haq Shah, M.Z.; Soni, M.; Shrivastava, V.K.; Mir, M.A.; Muzamil, S. Gallic acid reverses ovarian disturbances in mice with letrozole-induced PCOS via modulating Adipo R1 expression. Toxicol. Rep. 2022, 9, 1938–1949. [Google Scholar] [CrossRef]
- Spritzer, P.M.; Lecke, S.B.; Satler, F.; Morsch, D.M. Adipose tissue dysfunction, adipokines, and low-grade chronic inflammation in polycystic ovary syndrome. Reproduction 2015, 149, R219–R227. [Google Scholar] [CrossRef] [Green Version]
- Benrick, A.; Chanclón, B.; Micallef, P.; Wu, Y.; Hadi, L.; Shelton, J.M.; Stener-Victorin, E.; Wernstedt Asterholm, I. Adiponectin protects against development of metabolic disturbances in a PCOS mouse model. Proc. Natl. Acad. Sci. USA 2017, 114, E7187–E7196. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Chanclón, B.; Micallef, P.; Stener-Victorin, E.; Wernstedt Asterholm, I.; Benrick, A. Maternal adiponectin prevents visceral adiposity and adipocyte hypertrophy in prenatal androgenized female mice. FASEB J. 2021, 35, e21299. [Google Scholar] [CrossRef]
- Shah, M.Z.; Shrivastava, V.K. Ameliorative effects of quercetin on endocrine and metabolic abnormalities associated with experimentally induced polycystic ovary syndrome in mice. Comp. Clin. Pathol. 2023, 32-1. [Google Scholar] [CrossRef]
- Li, H.; Chen, Y.; Li, Y.; Huang, J.; Zhao, X.; Chen, X.; Yang, D. A case-control study of correlation between serum adiponectin levels and polycystic ovary syndrome. Zhonghua Fu Chan Ke Za Zhi 2015, 50, 814–818. [Google Scholar]
- Biljan, M.M.; Mahutte, N.G.; Tulandi, T.; Tan, S.L. Prospective randomized double-blind trial of the correlation between time of administration and antiestrogenic effects of clomiphene citrate on reproductive end organs. Fertil. Steril. 1999, 71, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Mitwally, M.F.M.; Casper, R.F. Aromatase inhibitors in ovulation induction. Semin. Reprod. Med. 2004, 22, 61–78. [Google Scholar] [PubMed]
- Donangelo, F.A. Phenolic compounds in coffee. Braz. J. Plant Physiol. 2006, 18, 23–36. [Google Scholar]
- Clifford, M.N. Chlorogenic acids and other cinnamates–nature, occurrence and dietary burden. J. Sci. Food Agric. 1999, 79, 362–372. [Google Scholar] [CrossRef]
- Cropley, V.; Croft, R.; Silber, B.; Neale, C.; Scholey, A.; Stough, C.; Schmitt, J. Does coffee enriched with chlorogenic acids improve mood and cognition after acute administration in healthy elderly? A pilot study. Psychopharmacology 2012, 219, 737–749. [Google Scholar] [CrossRef]
- Feng, R.; Lu, Y.; Bowman, L.L.; Qian, Y.; Castranova, V.; Ding, M. Inhibition of activator protein-1, NF-κB, and MAPKs and induction of phase 2 detoxifying enzyme activity by chlorogenic acid. J. Biol. Chem. 2005, 280, 27888–27895. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.J.; Jun, S.H.; Park, Y.; Cha, S.-H.; Yoon, M.; Cho, S.; Park, Y. Green synthesis of gold nanoparticles using chlorogenic acid and their enhanced performance for inflammation. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 1677–1688. [Google Scholar] [CrossRef]
- McCarty, M.F. A chlorogenic acid-induced increase in GLP-1 production may mediate the impact of heavy coffee consumption on diabetes risk. Med. Hypotheses 2005, 64, 848–853. [Google Scholar] [CrossRef]
- Wan, C.W.; Wong, C.N.Y.; Pin, W.K.; Wong, M.H.Y.; Kwok, C.Y.; Chan, R.Y.K.; Chan, S.W. Chlorogenic acid exhibits cholesterol lowering and fatty liver attenuating properties by up-regulating the gene expression of PPAR-α in hypercholesterolemic rats induced with a high-cholesterol diet. Phytother. Res. 2013, 27, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Haidari, F.; Omidian, K.; Rafiei, H.; Zarei, M.; Shahi, M.M. Green tea (Camellia sinensis) supplementation to diabetic rats improves serum and hepatic oxidative stress markers. Iran. J. Pharm. Res. 2013, 12, 109. [Google Scholar] [PubMed]
- ul haq Shah, M.Z.; Shrivastava, V.; Mir, M.A. Metformin treatment ameliorates endocrine-metabolic disturbances in letrozole-induced PCOS mice model by modulating adiponectin status. Obes. Med. Res. 2022, 31, 100392. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.-L.; Ideker, T. Cytoscape 2.8: New features for data integration and network visualization. Bioinformatics 2011, 27, 431–432. [Google Scholar] [CrossRef] [Green Version]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Lachmann, A. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Soumya, V.; Muzib, Y.I.; Venkatesh, P. A novel method of extraction of bamboo seed oil (Bambusa bambos Druce) and its promising effect on metabolic symptoms of experimentally induced polycystic ovarian disease. Indian J. Pharmacol. 2016, 48, 162. [Google Scholar] [CrossRef]
- Rosenfield, R.L.; Ehrmann, D.A. The pathogenesis of polycystic ovary syndrome (PCOS): The hypothesis of PCOS as functional ovarian hyperandrogenism revisited. Endocr. Rev. 2016, 37, 467–520. [Google Scholar] [CrossRef]
- Joham, A.E.; Teede, H.J.; Ranasinha, S.; Zoungas, S.; Boyle, J. Prevalence of infertility and use of fertility treatment in women with polycystic ovary syndrome: Data from a large community-based cohort study. J. Women’s Health 2015, 24, 299–307. [Google Scholar] [CrossRef]
- Cussons, A.J.; Watts, G.F.; Burke, V.; Shaw, J.E.; Zimmet, P.Z.; Stuckey, B.G.A. Cardiometabolic risk in polycystic ovary syndrome: A comparison of different approaches to defining the metabolic syndrome. Hum. Reprod. 2008, 23, 2352–2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olaniyi, K.S.; Olatunji, L.A. L-glutamine ameliorates adipose-hepatic dysmetabolism in OC-treated female rats. J. Endocrinol. 2020, 246, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dunaif, A.; Mandeli, J.; Fluhr, H.; Dobrjansky, A. The impact of obesity and chronic hyperinsulinemia on gonadotropin release and gonadal steroid secretion in the polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 1988, 66, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Carmina, E.; Bucchieri, S.; Esposito, A.; Del Puente, A.; Mansueto, P.; Orio, F.; Rini, G. Abdominal fat quantity and distribution in women with polycystic ovary syndrome and extent of its relation to insulin resistance. J. Clin. Endocrinol. Metab. 2007, 92, 2500–2505. [Google Scholar] [CrossRef] [Green Version]
- Bhandarkar, N.S.; Brown, L.; Panchal, S.K. Chlorogenic acid attenuates high-carbohydrate, high-fat diet–induced cardiovascular, liver, and metabolic changes in rats. Nutr. Res. 2019, 62, 78–88. [Google Scholar] [CrossRef]
- Groth, S.W. Adiponectin and polycystic ovary syndrome Jul. Biol. Res. Nurs. 2010, 12, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Gibb, F.W.; Homer, N.Z.M.; Faqehi, A.M.M.; Upreti, R.; Livingstone, D.E.; McInnes, K.J.; Walker, B.R. Aromatase inhibition reduces insulin sensitivity in healthy men. J. Clin. Endocrinol. Metab. 2016, 101, 2040–2046. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Kim, J.Y.; Xue, K.; Liu, J.Y.; Leader, A.; Tsang, B.K. Chemerin, a novel regulator of follicular steroidogenesis and its potential involvement in polycystic ovarian syndrome. Endocrinology 2012, 153, 5600–5611. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.; Shrivastava, V.K. Turmeric extract alleviates endocrine-metabolic disturbances in letrozole-induced PCOS by increasing adiponectin circulation: A comparison with Metformin. Metab. Open 2021, 13, 100160. [Google Scholar] [CrossRef]
- Kafali, H.; Iriadam, M.; Ozardali, I.; Demir, N. Letrozole-induced polycystic ovaries in the rat: A new model for cystic ovarian disease. Arch. Med. Res. 2004, 35, 103–108. [Google Scholar] [CrossRef]
- Abbott, D.H.; Barnett, D.K.; Bruns, C.M.; Dumesic, D.A. Androgen excess fetal programming of female reproduction: A developmental aetiology for polycystic ovary syndrome Hum. Reprod. Update 2005, 11, 357–374. [Google Scholar] [CrossRef] [Green Version]
- Dewailly, D.; Robin, G.; Peigne, M.; Decanter, C.; Pigny, P.; Catteau-Jonard, S. Interactions between androgens, FSH, anti-Müllerian hormone and estradiol during folliculogenesis in the human normal and polycystic ovary. Hum. Reprod. Update 2016, 22, 709–724. [Google Scholar] [CrossRef] [Green Version]
- Hammes, S.R. Androgens regulate ovarian follicular development by increasing follicle stimulating hormone receptor and microRNA-125b expression. Proc. Natl. Acad. Sci. USA 2014, 111, 3008–3013. [Google Scholar] [CrossRef] [Green Version]
- Chauvin, S.; Cohen-Tannoudji, J.; Guigon, C.J. Estradiol Signaling at the Heart of Folliculogenesis: Its Potential Deregulation in Human Ovarian Pathologies. Int. J. Mol. Sci. 2022, 23, 512. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Kamada, Y.; Hosoya, T.; Fujita, S.; Nishiyama, Y.; Iwata, N.; Hiramatsu, Y.; Otsuka, F. A regulatory role of androgen in ovarian steroidogenesis by rat granulosa cells. J. Steroid Biochem. Mol. Biol. 2017, 172, 160–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, J.W.; Dille, E.A.; Heckert, L.L. Current concepts of follicle-stimulating hormone receptor gene regulation. Biol. Reprod. 2011, 84, 7–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edson, M.A.; Nagaraja, A.K.; Matzuk, M.M. The mammalian ovary from genesis to revelation. Endocr. Rev. 2009, 30, 624–712. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Kim, H.J.; Pyun, B.J.; Lee, H.W. Licorice ethanol extract improves symptoms of polycytic ovary syndrome in Letrozole-induced female rats. Integr. Med. Res. 2018, 7, 264–270. [Google Scholar] [CrossRef]
- Hidaka, T.; Yonezawa, R.; Saito, S. Kami-shoyo-san, Kampo (Japanese traditional medicine), is effective for climacteric syndrome, especially in hormone-replacement-therapy-resistant patients who strongly complain of psychological symptoms. J. Obstet. Gynaecol. Res. 2013, 39, 223–228. [Google Scholar] [CrossRef]
- Baravalle, C.; Salvetti, N.R.; Mira, G.A.; Pezzone, N.; Ortega, H.H. Microscopic characterization of follicular structures in letrozoleinduced polycystic ovarian syndrome in the rat. Arch. Med. Res. 2006, 37, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Gomez, B.; Ramos, S.; Goya, L.; Mesa, M.D.; del Castillo, M.D.; Martín, M.Á. Coffee silverskin extract improves glucose-stimulated insulin secretion and protects against streptozotocin-induced damage in pancreatic INS-1E beta cells. Food Res. Int. 2016, 89, 1015–1022. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Zhao, Y. MiR-185-5p protects against angiogenesis in polycystic ovary syndrome by targeting VEGFA. Front. Pharmacol. 2020, 11, 1030. [Google Scholar] [CrossRef]
- El-Bahya, A.A.Z.; Radwanb, R.A.; Gadc, M.Z.; Abdel, S.M. A closer insight into the role of vitamin D in polycystic ovary syndrome (Pcos). Glob. J. Pharm. Pharm. Sci. 2018, 6, 79–87. [Google Scholar]
- Artini, P.G.; Monti, M.; Matteucci, C.; Valentino, V.; Cristello, F.; Genazzani, A.R. Vascular endothelial growth factor and basic fibroblast growth factor in polycystic ovary syndrome during controlled ovarian hyperstimulation. Gynecol. Endocrinol. 2006, 22, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Pittas, A.G.; Joseph, N.A.; Greenberg, A.S. Adipocytokines and insulin resistance. J Clin Endocrinol Metab 2004, 89, 447–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Frankenberg, A.D.; do Nascimento, F.V.; Gatelli, L.E.; Nedel, B.L.; Garcia, S.P.; de Oliveira, C.S.V.; Gerchman, F. Major components of metabolic syndrome and adiponectin levels: A cross-sectional study. Diabetol. Metab. Syndr. 2014, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- Rak, A.; Mellouk, N.; Froment, P.; Dupont, J. Adiponectin and resistin: Potential metabolic signals affecting hypothalamo-pituitary gonadal axis in females and males of different species. Reproduction 2017, 153, R215–R226. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Hu, T.; Zhao, H.; Huang, Y.; Ye, R.; Lin, J.; Zhang, C.; Zhang, H.; Wei, G.; Zhou, H.; et al. Brown adipose tissue transplantation ameliorates polycystic ovary syndrome. Proc. Natl. Acad. Sci. USA 2016, 113, 2708–2713. [Google Scholar] [CrossRef] [Green Version]
- Chabrolle, C.; Tosca, L.; Ramé, C.; Lecomte, P.; Royère, D.; Dupont, J. Adiponectin increases insulin-like growth factor I-induced progesterone and estradiol secretion in human granulosa cells. Fertil. Steril. 2009, 92, 1988–1996. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No | Gene | Forward Primer (5′ –3′) | Reverse Primer (3′ –5′) |
|---|---|---|---|
| 1 | Ar | CTGGGAAGGGTCTACCCAC | GGTGCTATGTTAGCGGCCTC |
| 2 | Lhr | ACACTGCCCTCCAAAGAAAA | CCTCAAAGATGGCGGAATAA |
| 3 | Fshr | CTCATCAAGCGACACCAAGA | GGAAAGGATTGGCACAAGAA |
| 4 | Esr2 | AGTAGCCGGAAGCTGACACA | CATGCTGAGCAGATGTTCCA |
| 5 | AdipoR1 | CGC TTT CTG CGT ATC GTC TG | CCA ACC TGC ACA AGT TCC CTT |
| 6 | Actin | TACGTCGCCCTGGATTTT | ATGAAAGAGGGCTGGAAGAG |
| Groups | Fasting Glucose (mg/dL) | Total Cholesterol (TC) (mg/dL) | Triglycerides (mg/dL) | HDL (mg/dL) | LDL (mg/dL) | VLDL (mg/dL) |
|---|---|---|---|---|---|---|
| Control | 109.96 ± 1.52 | 118.56 ± 1.56 | 121.23 ± 0.99 | 78.63 ± 0.99 | 15.68 ± 2.21 | 24.25 ± 0.9 |
| LETZ | 153.52 ± 2.48 a *** | 155.18 ± 2.91 a *** | 153.93 ± 1.51 a *** | 48.46 ± 1.87 a *** | 75.93 ± 4.11 a *** | 30.78 ± 0.3 a *** |
| LETZ with CGA | 117.86 ± 3.41 b *** | 130.78 ± 2.24 b *** | 128.86 ± 1.65 b *** | 69.16 ± 1.15 b *** | 35.84 ± 3.4 b *** | 25.77 ± 0.3 b *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, M.Z.u.H.; Shrivastava, V.K.; Sofi, S.; Jamous, Y.F.; Khan, M.F.; Alkholifi, F.K.; Ahmad, W.; Mir, M.A. Chlorogenic Acid Restores Ovarian Functions in Mice with Letrozole-Induced Polycystic Ovarian Syndrome Via Modulation of Adiponectin Receptor. Biomedicines 2023, 11, 900. https://doi.org/10.3390/biomedicines11030900
Shah MZuH, Shrivastava VK, Sofi S, Jamous YF, Khan MF, Alkholifi FK, Ahmad W, Mir MA. Chlorogenic Acid Restores Ovarian Functions in Mice with Letrozole-Induced Polycystic Ovarian Syndrome Via Modulation of Adiponectin Receptor. Biomedicines. 2023; 11(3):900. https://doi.org/10.3390/biomedicines11030900
Chicago/Turabian StyleShah, Mohd Zahoor ul Haq, Vinoy Kumar Shrivastava, Shazia Sofi, Yahya F. Jamous, Mohd Faiyaz Khan, Faisal K. Alkholifi, Wasim Ahmad, and Manzoor Ahmad Mir. 2023. "Chlorogenic Acid Restores Ovarian Functions in Mice with Letrozole-Induced Polycystic Ovarian Syndrome Via Modulation of Adiponectin Receptor" Biomedicines 11, no. 3: 900. https://doi.org/10.3390/biomedicines11030900