
Combined Usage of MDK Inhibitor Augments Interferon-γ Anti-Tumor Activity in the SKOV3 Human Ovarian Cancer Cell Line
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture
2.2. Antibodies and Reagents
2.3. MDK Stable Overexpression
2.4. RNA Extraction and Real-Time Quantitative PCR (RT-qPCR)
2.5. Western Blotting Assay
2.6. Cell Cytotoxicity Assay
2.7. Real Time Cell Analysis (RTCA)
2.8. Cell Migration and Invasion Assay
2.9. Statistical Analysis
3. Results
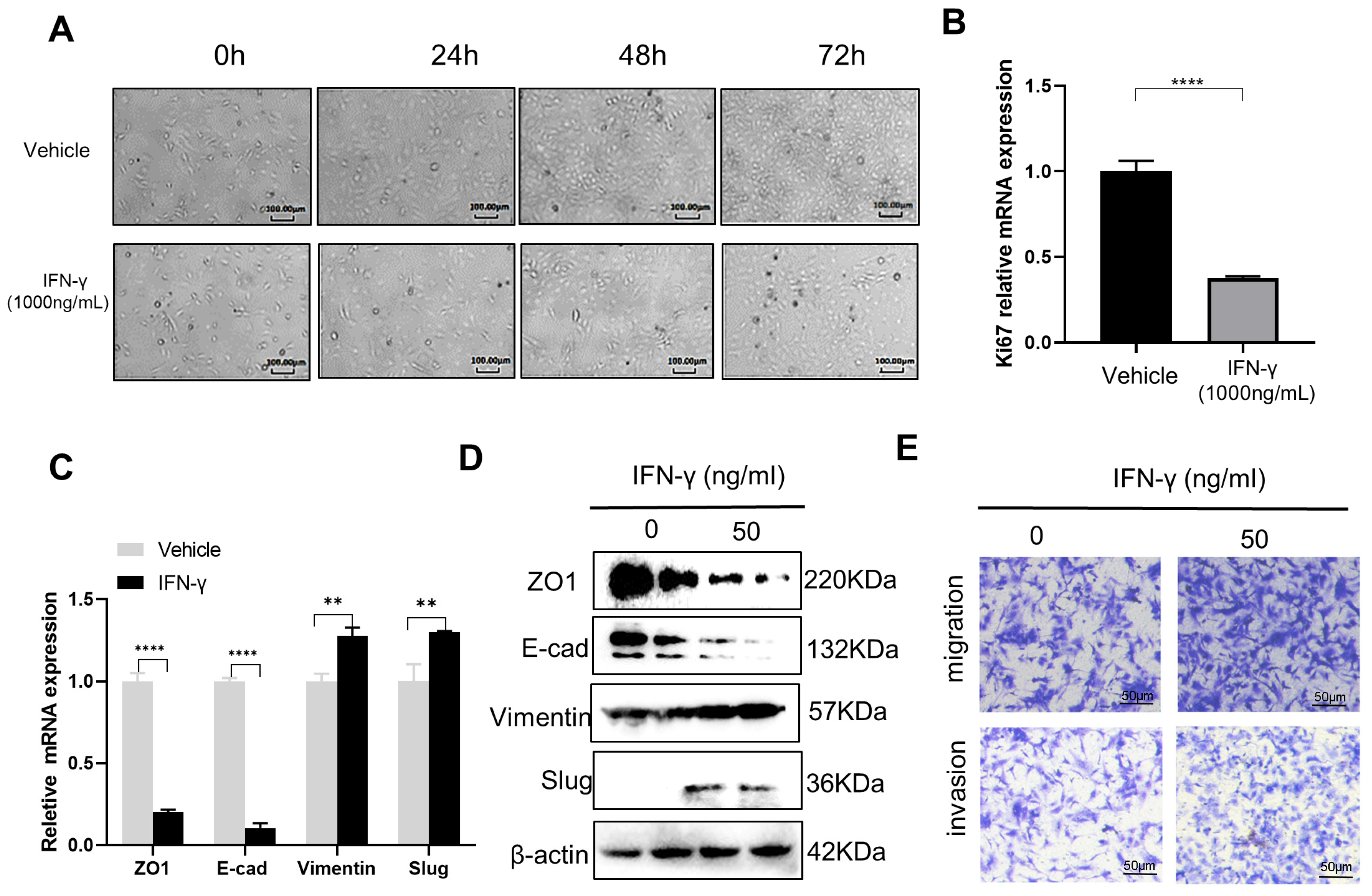
3.1. High-Dose IFN-γ Inhibits Cancer Growth while Low-Dose IFN-γ Promotes Aggressiveness in SKOV3 Cells
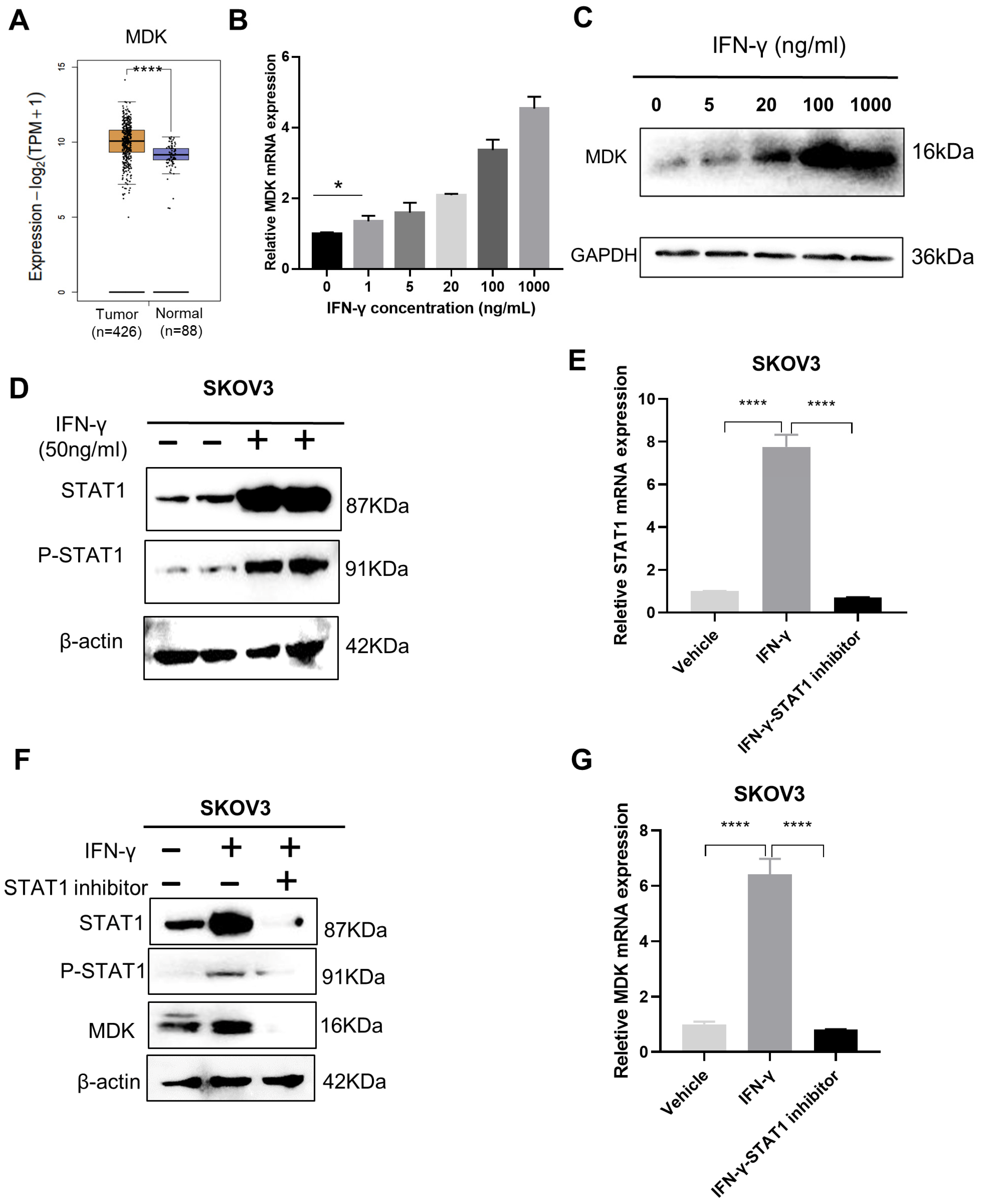
3.2. IFN-γ Activates MDK via STAT1 in SKOV3 Cells
3.3. MDK Promotes Cell Proliferation, Migration, and Invasion in SKOV3 Cells
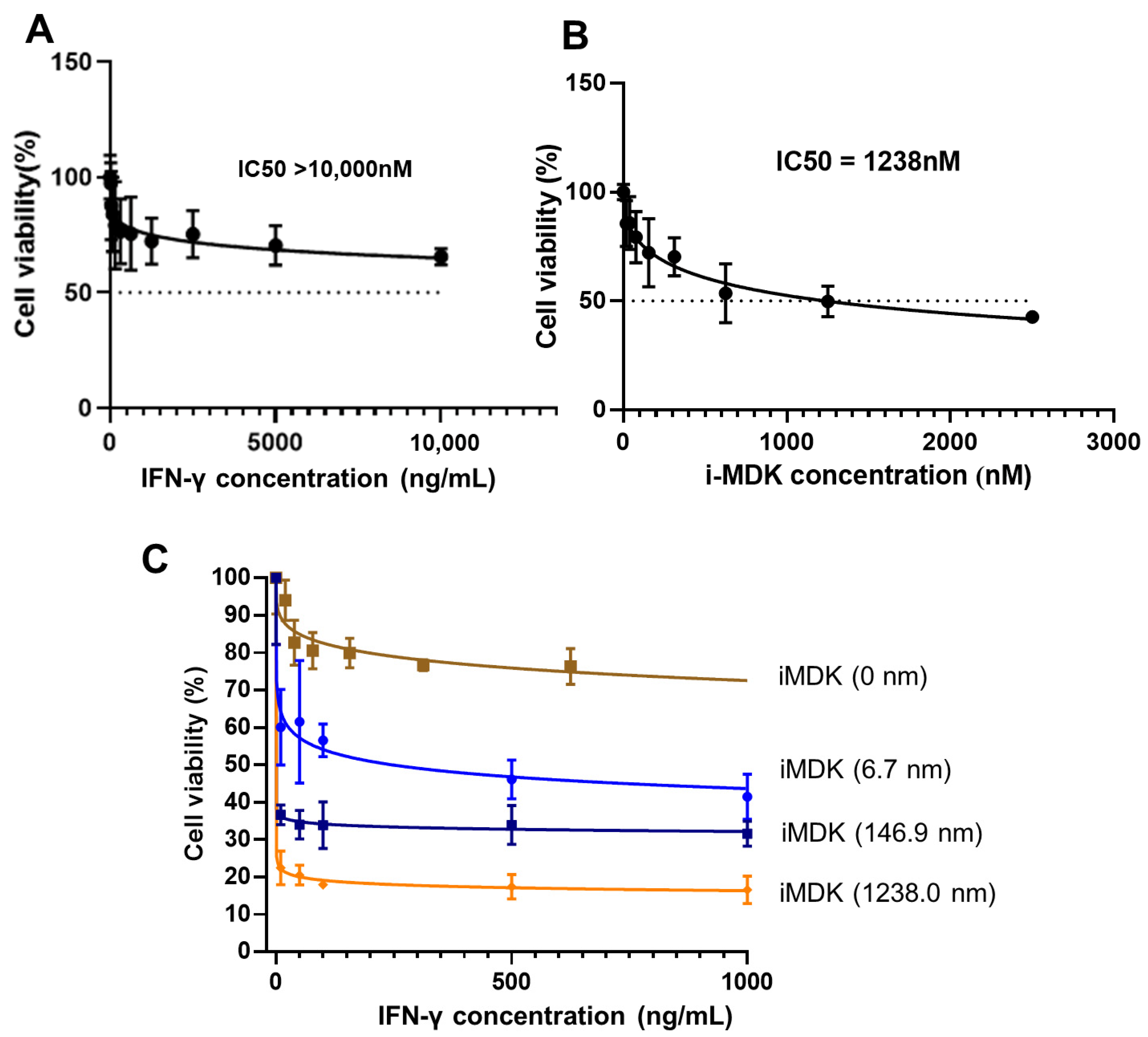
3.4. MDK Inhibition Potentiates the Tumoricidal Effect of IFN-γ in SKOV3 Cells
3.5. MDK Inhibition Attenuates the Pro-Metastatic Adverse Effect of IFN-γ
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Matulonis, U.A.; Sood, A.K.; Fallowfield, L.; Howitt, B.E.; Sehouli, J.; Karlan, B.Y. Ovarian cancer. Nat. Rev. Dis. Primers 2016, 2, 16061. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Pastor, R.; Goedegebuure, P.S.; Curiel, D.T. Understanding and addressing barriers to successful adenovirus-based virotherapy for ovarian cancer. Cancer Gene Ther. 2021, 28, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Savage, S.R.; Calinawan, A.P.; Lin, C.; Zhang, B.; Wang, P.; Starr, T.K.; Birrer, M.J.; Paulovich, A.G. A highly annotated database of genes associated with platinum resistance in cancer. Oncogene 2021, 40, 6395–6405. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, R.; Sonego, M.; Pucci, B.; Addi, L.; Iannelli, F.; Capone, F.; Alfano, L.; Roca, M.S.; Milone, M.R.; Moccia, T.; et al. HSP90 identified by a proteomic approach as druggable target to reverse platinum resistance in ovarian cancer. Mol. Oncol. 2021, 15, 1005–1023. [Google Scholar] [CrossRef]
- Li, S.S.; Ma, J.; Wong, A. Chemoresistance in ovarian cancer: Exploiting cancer stem cell metabolism. J. Gynecol. Oncol. 2018, 29, e32. [Google Scholar] [CrossRef] [Green Version]
- Bartoletti, M.; Musacchio, L.; Giannone, G.; Tuninetti, V.; Bergamini, A.; Scambia, G.; Pignata, S. Emerging molecular alterations leading to histology-specific targeted therapies in ovarian cancer beyond PARP inhibitors. Cancer Treat. Rev. 2021, 101, 102298. [Google Scholar] [CrossRef]
- Pietilä, E.A.; Gonzalez-Molina, J.; Moyano-Galceran, L.; Jamalzadeh, S.; Zhang, K.; Lehtinen, L.; Lehti, K. Co-evolution of matrisome and adaptive adhesion dynamics drives ovarian cancer chemoresistance. Nat. Commun. 2021, 12, 3904. [Google Scholar] [CrossRef]
- Miller, C.H.; Maher, S.G.; Young, H.A. Clinical Use of Interferon-gamma. Ann. N. Y. Acad. Sci. 2009, 1182, 69–79. [Google Scholar] [CrossRef]
- Zaidi, M.R. The Interferon-Gamma Paradox in Cancer. J. Interferon Cytokine Res. 2019, 39, 30–38. [Google Scholar] [CrossRef]
- Castro, F.; Cardoso, A.P.; Gonçalves, R.M.; Serre, K.; Oliveira, M.J. Interferon-Gamma at the Crossroads of Tumor Immune Surveillance or Evasion. Front. Immunol. 2018, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Wall, L.; Burke, F.; Barton, C.; Smyth, J.; Balkwill, F. IFN-gamma induces apoptosis in ovarian cancer cells in vivo and in vitro. Clin. Cancer Res. 2003, 9, 2487–2496. [Google Scholar]
- Wall, L.; Burke, F.; Smyth, J.F.; Balkwill, F. The anti-proliferative activity of interferon-gamma on ovarian cancer: In vitro and in vivo. Gynecol. Oncol. 2003, 88 Pt 2, S149–S151. [Google Scholar] [CrossRef]
- Pujade-Lauraine, E.; Guastalla, J.P.; Colombo, N.; Francois, E.; Fumoleau, P.; Monnier, A.; Brandely, M. Intraperitoneal recombinant interferon gamma in ovarian cancer patients with residual disease at second-look laparotomy. J. Clin. Oncol. 1996, 14, 343–350. [Google Scholar] [CrossRef]
- Windbichler, G.H.; Hausmaninger, H.; Stummvoll, W.; Graf, A.H.; Kainz, C.; Lahodny, J. Interferon-gamma in the first-line therapy of ovarian cancer: A randomized phase III trial. Br. J. Cancer. 2000, 82, 1138–1144. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, M.R.; Merlino, G. The two faces of interferon-γ in cancer. Clin. Cancer Res. 2011, 17, 6118–6124. [Google Scholar] [CrossRef] [Green Version]
- Devane, J.G.; Martin, M.L.; Matson, M.A. A short 2 week dose titration regimen reduces the severity of flu-like symptoms with initial interferon gamma-1b treatment. Curr. Med. Res. Opin. 2014, 30, 1179–1187. [Google Scholar] [CrossRef]
- Song, M.; Ping, Y.; Zhang, K.; Yang, L.; Li, F.; Zhang, C.; Zhang, Y. Low-Dose IFNγ Induces Tumor Cell Stemness in Tumor Microenvironment of Non-Small Cell Lung Cancer. Cancer Res. 2019, 79, 3737–3748. [Google Scholar] [CrossRef]
- Lo, U.G.; Bao, J.; Cen, J.; Yeh, H.C.; Luo, J.; Tan, W.; Hsieh, J.T. Interferon-induced IFIT5 promotes epithelial-to-mesenchymal transition leading to renal cancer invasion. Am. J. Clin. Exp. Urol. 2019, 7, 31–45. [Google Scholar]
- Gong, W.; Zhang, G.M.; Liu, Y.; Lei, Z.; Li, D.; Yuan, Y.; Feng, Z.H. IFN-gamma withdrawal after immunotherapy potentiates B16 melanoma invasion and metastasis by intensifying tumor integrin alphavbeta3 signaling. Int. J. Cancer 2008, 123, 702–708. [Google Scholar] [CrossRef]
- Kelly, S.A.; Gschmeissner, S.; East, N.; Balkwill, F.R. Enhancement of metastatic potential by gamma-interferon. Cancer Res. 1991, 51, 4020–4027. [Google Scholar] [PubMed]
- Zheng, L.; Liu, Q.; Li, R.; Chen, S.; Tan, J.; Li, L.; Liu, J. Targeting MDK Abrogates IFN-γ-Elicited Metastasis inCancers of Various Origins. Front. Oncol. 2022, 12, 885656. [Google Scholar] [CrossRef] [PubMed]
- Alberts, D.S.; Marth, C.; Alvarez, R.D.; Johnson, G.; Bidzinski, M.; Kardatzke, D.R. Randomized phase 3 trial of interferon gamma-1b plus standard carboplatin/paclitaxel versus carboplatin/paclitaxel alone for first-line treatment of advanced ovarian and primary peritoneal carcinomas: Results from a prospectively designed analysis of progression-free survival. Gynecol. Oncol. 2008, 109, 174–181. [Google Scholar] [PubMed]
- Jorgovanovic, D.; Song, M.; Wang, L.; Zhang, Y. Roles of IFN-γ in tumor progression and regression: A review. Biomark. Res. 2020, 8, 49. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Zhang, H.; Wu, J.; Guan, Y. Effect of plasmid-mediated stable interferon-γ expression on proliferation and cell death in the SKOV-3 human ovarian cancer cell line. Immunopharmacol. Immunotoxicol. 2011, 33, 498–503. [Google Scholar] [CrossRef]
- Lo, U.G.; Pong, R.C.; Yang, D.; Gandee, L.; Hernandez, E.; Dang, A.; Hsieh, J.T. IFNγ-Induced IFIT5 Promotes Epithelial-to-Mesenchymal Transition in Prostate Cancer via miRNA Processing. Cancer Res. 2019, 79, 1098–1112. [Google Scholar] [CrossRef]
- Chow, K.T.; Gale, M., Jr. SnapShot: Interferon Signaling. Cell 2015, 163, 1808.e1. [Google Scholar] [CrossRef]
- Liu, C.; Gao, A.C. IFNγ, a Double-Edged Sword in Cancer Immunity and Metastasis. Cancer Res. 2019, 79, 1032–1033. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Liu, J.; Xue, H.; Li, C.; Liu, Q.; Zhou, Y.; Wen, T. A TGF-β-MTA1-SOX4-EZH2 signaling axis drives epithelial-mesenchymal transition in tumor metastasis. Oncogene 2020, 39, 2125–2139. [Google Scholar] [CrossRef]
- Menon, U.; Karpinskyj, C.; Gentry-Maharaj, A. Ovarian Cancer Prevention and Screening. Obstet. Gynecol. 2018, 131, 909–927. [Google Scholar] [CrossRef] [Green Version]
- Lisio, M.A.; Fu, L.; Goyeneche, A.; Gao, Z.H.; Telleria, C. High-Grade Serous Ovarian Cancer: Basic Sciences, Clinical and Therapeutic Standpoints. Int. J. Mol. Sci. 2019, 20, 952. [Google Scholar] [CrossRef] [Green Version]
- Govindarajan, M.; Wohlmuth, C.; Waas, M.; Bernardini, M.Q.; Kislinger, T. High-throughput approaches for precision medicine in high-grade serous ovarian cancer. J. Hematol. Oncol. 2020, 13, 134. [Google Scholar] [CrossRef]
- Malik, S.T.; Knowles, R.G.; East, N.; Lando, D.; Stamp, G.; Balkwill, F.R. Antitumor activity of gamma-interferon in ascitic and solid tumor models of human ovarian cancer. Cancer Res. 1991, 51, 6643–6649. [Google Scholar]
- Razaghi, A.; Villacrés, C.; Jung, V.; Mashkour, N.; Butler, M.; Owens, L.; Heimann, K. Improved therapeutic efficacy of mammalian expressed-recombinant interferon gamma against ovarian cancer cells. Exp. Cell Res. 2017, 359, 20–29. [Google Scholar] [CrossRef]
- Burke, F.; East, N.; Upton, C.; Patel, K.; Balkwill, F.R. Interferon gamma induces cell cycle arrest and apoptosis in a model of ovarian cancer: Enhancement of effect by batimastat. Eur. J. Cancer 1997, 33, 1114–1121. [Google Scholar] [CrossRef]
- Marth, C.; Windbichler, G.H.; Hausmaninger, H.; Petru, E.; Estermann, K.; Pelzer, A.; Mueller-Holzner, E. Interferon-gamma in combination with carboplatin and paclitaxel as a safe and effective first-line treatment option for advanced ovarian cancer: Results of a phase I/II study. Int. J. Gynecol. Cancer 2006, 16, 1522–1528. [Google Scholar] [CrossRef]
- Filippou, P.S.; Karagiannis, G.S.; Constantinidou, A. Midkine (MDK) growth factor: A key player in cancer progression and a promising therapeutic target. Oncogene 2020, 39, 2040–2054. [Google Scholar] [CrossRef]
- Zhang, X.; Karim, M.; Hasan, M.M.; Hooper, J.; Wahab, R.; Roy, S.; Al-Hilal, T.A. Cancer-on-a-Chip: Models for Studying Metastasis. Cancers 2022, 14, 648. [Google Scholar] [CrossRef]
- Kavand, H.; Nasiri, R.; Herland, A. Advanced Materials and Sensors for Microphysiological Systems: Focus on Electronic and Electrooptical Interfaces. Adv Mater. 2022, 34, e2107876. [Google Scholar] [CrossRef]
- Cerezo-Wallis, D.; Contreras-Alcalde, M.; Troulé, K.; Catena, X.; Mucientes, C.; Calvo, T.G.; Soengas, M.S. Midkine rewires the melanoma microenvironment toward a tolerogenic and immune-resistant state. Nat. Med. 2020, 26, 1865–1877. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | IFN-γ (ng/mL) | iMDK (μM) | FA | CI |
|---|---|---|---|---|
| 1 | 10 | 6.662 | 0.3996 | 0.01426 |
| 2 | 50 | 6.662 | 0.3851 | 0.02502 |
| 3 | 100 | 6.662 | 0.4344 | 0.02188 |
| 4 | 500 | 6.662 | 0.5388 | 0.02493 |
| 5 | 1000 | 6.662 | 0.5852 | 0.02835 |
| 1 | 10 | 146.896 | 0.6339 | 0.03549 |
| 2 | 50 | 146.896 | 0.6599 | 0.02825 |
| 3 | 100 | 146.896 | 0.6613 | 0.02843 |
| 4 | 500 | 146.896 | 0.6608 | 0.03300 |
| 5 | 1000 | 146.896 | 0.6845 | 0.03017 |
| 1 | 10 | 1238 | 0.6339 | 0.06794 |
| 2 | 50 | 1238 | 0.6599 | 0.05325 |
| 3 | 100 | 1238 | 0.6613 | 0.03741 |
| 4 | 500 | 1238 | 0.6608 | 0.03482 |
| 5 | 1000 | 1238 | 0.6845 | 0.03114 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.; Tan, J.; Zhao, Z.; Li, R.; Zheng, L.; Chen, X.; Li, L.; Dong, X.; Wen, T.; Liu, J. Combined Usage of MDK Inhibitor Augments Interferon-γ Anti-Tumor Activity in the SKOV3 Human Ovarian Cancer Cell Line. Biomedicines 2023, 11, 8. https://doi.org/10.3390/biomedicines11010008
Liu Q, Tan J, Zhao Z, Li R, Zheng L, Chen X, Li L, Dong X, Wen T, Liu J. Combined Usage of MDK Inhibitor Augments Interferon-γ Anti-Tumor Activity in the SKOV3 Human Ovarian Cancer Cell Line. Biomedicines. 2023; 11(1):8. https://doi.org/10.3390/biomedicines11010008
Chicago/Turabian StyleLiu, Qun, Jingyu Tan, Zhenguo Zhao, Ruijun Li, Luyu Zheng, Xiangyu Chen, Lina Li, Xichen Dong, Tao Wen, and Jian Liu. 2023. "Combined Usage of MDK Inhibitor Augments Interferon-γ Anti-Tumor Activity in the SKOV3 Human Ovarian Cancer Cell Line" Biomedicines 11, no. 1: 8. https://doi.org/10.3390/biomedicines11010008