
Aberrant Expression and Prognostic Potential of IL-37 in Human Lung Adenocarcinoma
 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Study of the Expression Levels of IL-37 and SIGIRR in Various Human Cancers
2.3. Investigation of IL-37 and SIGIRR Expression Levels in LUAD versus Non-LUAD Lung Tissue
2.4. Exploration of Associations between IL-37 or SIGIRR Expression Levels with Certain Pathological Characteristics of the LUAD Tumor
2.5. Assessment of the Effect of IL-37 and SIGIRR Expression Levels on Survival Rates of Patients with LUAD
2.6. Development of a List of Positively/Negatively Correlated Genes of IL-37 in LUAD Biopsies
2.7. Investigation of the IL-37 and SIGIRR Protein Expression Levels
2.8. Blood and Immune Single-Cell Analysis in Lung Tissue
2.9. Analysis of Possible Correlations between IL-37 Gene Alterations or Expression Levels with Immune Cell Infiltration Patterns
2.10. Analysis of Possible Correlations between IL-37 Gene Alterations or Expression Levels with Immune Cell Infiltration Patterns
2.11. Statistical Analysis
3. Results
3.1. Among Human Cancers, IL-37 Exhibits the Highest Expression Levels in Lung Adenocarcinoma (LUAD)
3.2. IL-37 Levels Are Increased in LUAD versus Non-LUAD Lung Tissues
3.3. IL-37 and SIGIRR Levels Are Associated with Histological Type and Tumor Grade of LUAD Tumors

3.4. Increased IL-37 Expression Is a Favorable Prognostic Factor for Overall Survival in LUAD Patients
3.5. Correlation of IL-37 Expression with Cancer-Associated Genes in LUAD Tumors
3.6. IL-37 Protein Expression Correlates with the Grade of LUAD Tumor
3.7. T-Lymphocytes and Macrophages of the Lung Express IL-37 and SIGIRR Genes
3.8. IL37 Gene Alterations Correlate with Differential Immune Cell Infiltration of the Lung Tumor
3.9. IL37 Expression Levels Correlate with Infiltration Levels of Certain Immune Cell Subsets
3.10. IL-37 Signaling Shares Common Nodes with PD-1/PDL-1 and CTLA-4 Immune Checkpoint Pathways
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Su, Z.; Tao, X. Current Understanding of IL-37 in Human Health and Disease. Front. Immunol. 2021, 12, 696605. [Google Scholar] [CrossRef] [PubMed]
- Cavalli, G.; Dinarello, C.A. Suppression of inflammation and acquired immunity by IL-37. Immunol. Rev. 2018, 281, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Sharaf, N.; Nicklin, M.J.; di Giovine, F.S. Long-range DNA interactions at the IL-1/IL-36/IL-37 gene cluster (2q13) are induced by activation of monocytes. Cytokine 2014, 68, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Bufler, P.; Gamboni-Robertson, F.; Azam, T.; Kim, S.H.; Dinarello, C.A. Interleukin-1 homologues IL-1F7b and IL-18 contain functional mRNA instability elements within the coding region responsive to lipopolysaccharide. Biochem. J. 2004, 381 Pt 2, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Sims, J.E.; Smith, D.E. The IL-1 family: Regulators of immunity. Nat. Rev. Immunol. 2010, 10, 89–102. [Google Scholar] [CrossRef]
- Rudloff, I.; Cho, S.X.; Lao, J.C.; Ngo, D.; McKenzie, M.; Nold-Petry, C.A.; Nold, M.F. Monocytes and dendritic cells are the primary sources of interleukin 37 in human immune cells. J. Leukoc. Biol. 2017, 101, 901–911. [Google Scholar] [CrossRef]
- Nold, M.F.; Nold-Petry, C.A.; Zepp, J.A.; Palmer, B.E.; Bufler, P.; Dinarello, C.A. IL-37 is a fundamental inhibitor of innate immunity. Nat. Immunol. 2010, 11, 1014–1022. [Google Scholar] [CrossRef] [Green Version]
- Cavalli, G.; Justice, J.N.; Boyle, K.E.; D’Alessandro, A.; Eisenmesser, E.Z.; Herrera, J.J.; Hansen, K.C.; Nemkov, T.; Stienstra, R.; Garlanda, C.; et al. Interleukin 37 reverses the metabolic cost of inflammation, increases oxidative respiration, and improves exercise tolerance. Proc. Natl. Acad. Sci. USA 2017, 114, 2313–2318. [Google Scholar] [CrossRef] [Green Version]
- Cavalli, G.; Koenders, M.; Kalabokis, V.; Kim, J.; Choon Tan, A.; Garlanda, C.; Mantovani, A.; Dagna, L.; Joosten, L.A.B.; Dinarello, C.A. Treating experimental arthritis with the innate immune inhibitor interleukin-37 reduces joint and systemic inflammation. Rheumatology 2017, 56, 2256. [Google Scholar] [CrossRef] [Green Version]
- Moretti, S.; Bozza, S.; Oikonomou, V.; Renga, G.; Casagrande, A.; Iannitti, R.G.; Puccetti, M.; Garlanda, C.; Kim, S.; Li, S.; et al. IL-37 inhibits inflammasome activation and disease severity in murine aspergillosis. PLoS Pathog. 2014, 10, e1004462. [Google Scholar] [CrossRef]
- Wald, D.; Qin, J.; Zhao, Z.; Qian, Y.; Naramura, M.; Tian, L.; Towne, J.; Sims, J.E.; Stark, G.R.; Li, X. SIGIRR, a negative regulator of Toll-like receptor-interleukin 1 receptor signaling. Nat. Immunol. 2003, 4, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Nold-Petry, C.A.; Lo, C.Y.; Rudloff, I.; Elgass, K.D.; Li, S.; Gantier, M.P.; Lotz-Havla, A.S.; Gersting, S.W.; Cho, S.X.; Lao, J.C.; et al. IL-37 requires the receptors IL-18Ralpha and IL-1R8 (SIGIRR) to carry out its multifaceted anti-inflammatory program upon innate signal transduction. Nat. Immunol. 2015, 16, 354–365. [Google Scholar] [CrossRef] [PubMed]
- Bulau, A.M.; Nold, M.F.; Li, S.; Nold-Petry, C.A.; Fink, M.; Mansell, A.; Schwerd, T.; Hong, J.; Rubartelli, A.; Dinarello, C.A.; et al. Role of caspase-1 in nuclear translocation of IL-37, release of the cytokine, and IL-37 inhibition of innate immune responses. Proc. Natl. Acad. Sci. USA 2014, 111, 2650–2655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimsby, S.; Jaensson, H.; Dubrovska, A.; Lomnytska, M.; Hellman, U.; Souchelnytskyi, S. Proteomics-based identification of proteins interacting with Smad3: SREBP-2 forms a complex with Smad3 and inhibits its transcriptional activity. FEBS Lett. 2004, 577, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Kulk, N.; Nold, M.F.; Graf, R.; Kim, S.H.; Reinhardt, D.; Dinarello, C.A.; Bufler, P. The IL-1 family member 7b translocates to the nucleus and down-regulates proinflammatory cytokines. J. Immunol. 2008, 180, 5477–5482. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Li, Y.; Guo, C.; Wang, L.; Chu, H.; Zhu, F.; Li, Y.; Wang, X.; Wang, Q.; Zhao, W.; et al. IL-37 isoform D downregulates pro-inflammatory cytokines expression in a Smad3-dependent manner. Cell Death Dis. 2018, 9, 582. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, C.; Ferreira, F. Tumor microenvironment of human breast cancer, and feline mammary carcinoma as a potential study model. Biochim. Biophys. Acta Rev. Cancer. 2021, 1876, 188587. [Google Scholar] [CrossRef]
- Christodoulou, M.I.; Zaravinos, A. New Clinical Approaches and Emerging Evidence on Immune-Checkpoint Inhibitors as Anti-Cancer Therapeutics: CTLA-4 and PD-1 Pathways and Beyond. Crit. Rev. Immunol. 2019, 39, 379–408. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, J.; He, P.; Han, J.; Sun, C. Interleukin-37 suppresses hepatocellular carcinoma growth through inhibiting M2 polarization of tumor-associated macrophages. Mol. Immunol. 2020, 122, 13–20. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, J.J.; Zhou, Z.Q.; Pan, Q.Z.; Zhu, Q.; Tang, Y.; Xia, J.C.; Weng, D.S. IL-37 induces anti-tumor immunity by indirectly promoting dendritic cell recruitment and activation in hepatocellular carcinoma. Cancer Manag. Res. 2019, 11, 6691–6702. [Google Scholar] [CrossRef]
- Wang, Z.; Zeng, F.L.; Hu, Y.W.; Wang, X.Y.; Zhao, F.L.; Zhou, P.; Hu, J.; Xiao, Y.Y.; Hu, Z.L.; Guo, M.F.; et al. Interleukin-37 promotes colitis-associated carcinogenesis via SIGIRR-mediated cytotoxic T cells dysfunction. Signal Transduct. Target. Ther. 2022, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.; Zhu, Y.; Teo, H.Y.; Liu, Y.; Song, Y.; Lim, H.Y.; Binte Hanafi, Z.; Angeli, V.; Liu, H. The indirect antiangiogenic effect of IL-37 in the tumor microenvironment. J. Leukoc. Biol. 2020, 107, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, M.; Guo, C.; Chu, H.; Li, W.; Chen, X.; Wang, X.; Li, Y.; Jia, Y.; Koussatidjoa, S.; et al. Intracellular mature IL-37 suppresses tumor metastasis via inhibiting Rac1 activation. Oncogene 2018, 37, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Zhao, Q.; Chen, W.; Zhao, Y.; Yan, Q.; Peng, R.; Zhu, J.; Yang, C.; Lan, K.; Gu, X.; et al. IL-37 Confers Anti-Tumor Activity by Regulation of m6A Methylation. Front. Oncol. 2020, 10, 526866. [Google Scholar] [CrossRef]
- Zhu, B.; Luo, J.; Jiang, Y.; Yu, L.; Liu, M.; Fu, J. Prognostic significance of nomograms integrating IL-37 expression, neutrophil level, and MMR status in patients with colorectal cancer. Cancer Med. 2018, 7, 3682–3694. [Google Scholar] [CrossRef]
- Haghshenas, M.R.; Hosseini, S.R.; Fattahi, M.J.; Malekzadeh, M.; Ariafar, A.; Ghaderi, A. Elevated IL-37 Serum Levels in Patients with Transitional Cell Carcinoma of Bladder. Iran. J. Immunol. 2021, 18, 346–353. [Google Scholar]
- Osborne, D.G.; Domenico, J.; Luo, Y.; Reid, A.L.; Amato, C.; Zhai, Z.; Gao, D.; Ziman, M.; Dinarello, C.A.; Robinson, W.A.; et al. Interleukin-37 is highly expressed in regulatory T cells of melanoma patients and enhanced by melanoma cell secretome. Mol. Carcinog. 2019, 58, 1670–1679. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Zhao, X.; Zhu, N.; Zhao, M.; Hu, Q.; Ni, Y. The balance of serum IL-18/IL-37 levels is disrupted during the development of oral squamous cell carcinoma. Surg. Oncol. 2020, 32, 99–107. [Google Scholar] [CrossRef]
- Wei, X.; Li, Y.; Zhang, G.; Wang, N.; Mi, M.; Xin, Y.; Jiang, H.; Sun, C. IL-37 Was Involved in Progress of Acute Myeloid Leukemia Through Regulating IL-6 Expression. Cancer Manag. Res. 2021, 13, 3393–3402. [Google Scholar] [CrossRef]
- Farahani, N.; Mohagheghi, F.; Mosayebi, G.; Ghazavi, A.; Ganji, A. Reduced IL-37 gene expression and CD8 T lymphocytes in patients with metastatic breast cancer. Breast Dis. 2021, 40, 235–240. [Google Scholar] [CrossRef]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjostedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A pathology atlas of the human cancer transcriptome. Science 2017, 357, 2507. [Google Scholar] [CrossRef] [PubMed]
- Bartha, A.; Gyorffy, B. TNMplot.com: A Web Tool for the Comparison of Gene Expression in Normal, Tumor and Metastatic Tissues. Int. J. Mol. Sci. 2021, 22, 2622. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.; Munkacsy, G.; Gyorffy, B. Pancancer survival analysis of cancer hallmark genes. Sci. Rep. 2021, 11, 6047. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Elzakra, N.; Kim, Y. HIF-1alpha Metabolic Pathways in Human Cancer. Adv. Exp. Med. Biol. 2021, 1280, 243–260. [Google Scholar] [PubMed]
- Rashid, M.; Zadeh, L.R.; Baradaran, B.; Molavi, O.; Ghesmati, Z.; Sabzichi, M.; Ramezani, F. Up-down regulation of HIF-1alpha in cancer progression. Gene 2021, 798, 145796. [Google Scholar] [CrossRef]
- Thul, P.J.; Akesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Ait Blal, H.; Alm, T.; Asplund, A.; Bjork, L.; Breckels, L.M.; et al. A subcellular map of the human proteome. Science 2017, 356, eaal3321. [Google Scholar] [CrossRef]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef]
- Barta, J.A.; Powell, C.A.; Wisnivesky, J.P. Global Epidemiology of Lung Cancer. Ann. Glob. Health 2019, 85, 8. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Society, A.C. Cancer Facts and Figures 2022. Available online: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2022.html (accessed on 24 August 2022).
- Jones, G.S.; Baldwin, D.R. Recent advances in the management of lung cancer. Clin. Med. 2018, 18 (Suppl. 2), s41–s46. [Google Scholar] [CrossRef] [PubMed]
- Myers, D.J.; Wallen, J.M. Lung adenocarcinoma. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Sun, B.; Zhao, H. A Study on Immune Cell Infiltration in Lung Adenocarcinoma. Comb. Chem. High Throughput Screen 2022, 25, 2082–2088. [Google Scholar] [PubMed]
- Chen, G.; Dong, Z.; Wu, D.; Chen, Y. Profiles of immune infiltration in lung adenocarcinoma and their clinical significant: A gene-expression-based retrospective study. J. Cell Biochem. 2020, 121, 4431–4439. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Li, M.; Chen, Z.; Bian, Y.; Zheng, Y.; Hu, Z.; Liang, J.; Huang, Y.; Yin, J.; Zhan, C.; et al. Identification of immune-related gene signature predicting survival in the tumor microenvironment of lung adenocarcinoma. Immunogenetics 2020, 72, 455–465. [Google Scholar] [CrossRef]
- Zuo, S.; Wei, M.; Wang, S.; Dong, J.; Wei, J. Pan-Cancer Analysis of Immune Cell Infiltration Identifies a Prognostic Immune-Cell Characteristic Score (ICCS) in Lung Adenocarcinoma. Front. Immunol. 2020, 11, 1218. [Google Scholar] [CrossRef]
- Insitute, N.C. The Cancer Genome Atlas. Available online: https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga (accessed on 24 August 2022).
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Zheng, M.; Shi, M.; Wang, J.; Huang, Z.; Zhang, H.; Zhou, Y.; Shi, J. Characteristic of molecular subtypes in lung adenocarcinoma based on m6A RNA methylation modification and immune microenvironment. BMC Cancer 2021, 21, 938. [Google Scholar] [CrossRef]
- Wu, C.; Rao, X.; Lin, W. Immune landscape and a promising immune prognostic model associated with TP53 in early-stage lung adenocarcinoma. Cancer Med. 2021, 10, 806–823. [Google Scholar] [CrossRef]
- Zhao, Z.; He, B.; Cai, Q.; Zhang, P.; Peng, X.; Zhang, Y.; Xie, H.; Wang, X. Combination of tumor mutation burden and immune infiltrates for the prognosis of lung adenocarcinoma. Int. Immunopharmacol. 2021, 98, 107807. [Google Scholar] [CrossRef] [PubMed]
- Sata, Y.; Nakajima, T.; Fukuyo, M.; Matsusaka, K.; Hata, A.; Morimoto, J.; Rahmutulla, B.; Ito, Y.; Suzuki, H.; Yoshino, I.; et al. High expression of CXCL14 is a biomarker of lung adenocarcinoma with micropapillary pattern. Cancer Sci. 2020, 111, 2588–2597. [Google Scholar] [CrossRef] [PubMed]
- Shaykhiev, R.; Sackrowitz, R.; Fukui, T.; Zuo, W.L.; Chao, I.W.; Strulovici-Barel, Y.; Downey, R.J.; Crystal, R.G. Smoking-induced CXCL14 expression in the human airway epithelium links chronic obstructive pulmonary disease to lung cancer. Am. J. Respir. Cell Mol. Biol. 2013, 49, 418–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sang, L.; He, Y.J.; Kang, J.; Ye, H.; Bai, W.; Luo, X.D.; Sun, J. Mitochondrial Deoxyguanosine Kinase Regulates NAD(+) Biogenesis Independent of Mitochondria Complex I Activity. Front. Oncol. 2020, 10, 570656. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, Q.; Shen, J.; Chen, X.; Ma, S. Multi-omics analysis identifies potential mechanisms of AURKB in mediating poor outcome of lung adenocarcinoma. Aging 2021, 13, 5946–5966. [Google Scholar] [CrossRef]
- Asada, K.; Kobayashi, K.; Joutard, S.; Tubaki, M.; Takahashi, S.; Takasawa, K.; Komatsu, M.; Kaneko, S.; Sese, J.; Hamamoto, R. Uncovering Prognosis-Related Genes and Pathways by Multi-Omics Analysis in Lung Cancer. Biomolecules 2020, 10, 524. [Google Scholar] [CrossRef] [Green Version]
- Kato, T.; Ujiie, H.; Hatanaka, K.C.; Nange, A.; Okumura, A.; Tsubame, K.; Naruchi, K.; Sato, M.; Kaga, K.; Matsuno, Y.; et al. A novel Tn antigen epitope-recognizing antibody for MUC1 predicts clinical outcome in patients with primary lung adenocarcinoma. Oncol. Lett. 2021, 21, 202. [Google Scholar] [CrossRef]
- Ren, J.; Yang, Y.; Li, C.; Xie, L.; Hu, R.; Qin, X.; Zhang, M. A Novel Prognostic Model of Early-Stage Lung Adenocarcinoma Integrating Methylation and Immune Biomarkers. Front. Genet. 2020, 11, 634634. [Google Scholar] [CrossRef]
- Wang, Z.; Li, X.; Chen, H.; Han, L.; Ji, X.; Wang, Q.; Wei, L.; Miao, Y.; Wang, J.; Mao, J.; et al. Decreased HLF Expression Predicts Poor Survival in Lung Adenocarcinoma. Med. Sci. Monit. 2021, 27, e929333. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, L.; Zhang, L.; Zheng, X.; Xu, H.; Wang, K.; Weng, X. Identification of a Four-Gene Signature Associated with the Prognosis Prediction of Lung Adenocarcinoma Based on Integrated Bioinformatics Analysis. Genes 2022, 13, 238. [Google Scholar] [CrossRef]
- Yang, D.; He, Y.; Wu, B.; Liu, R.; Wang, N.; Wang, T.; Luo, Y.; Li, Y.; Liu, Y. Predictions of the dysregulated competing endogenous RNA signature involved in the progression of human lung adenocarcinoma. Cancer Biomark. 2020, 29, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.K.; Abdollah, N.A.; Shafie, N.H.; Yusof, N.M.; Razak, S.R.A. Dual-specificity phosphatase 6 (DUSP6): A review of its molecular characteristics and clinical relevance in cancer. Cancer Biol. Med. 2018, 15, 14–28. [Google Scholar] [PubMed] [Green Version]
- Amelung, J.T.; Buhrens, R.; Beshay, M.; Reymond, M.A. Key genes in lung cancer translational research: A meta-analysis. Pathobiology 2010, 77, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Garcia, C.V.; Agudo-Lopez, A.; Perez, C.; Prieto-Garcia, E.; Iglesias, L.; Ponce, S.; Rodriguez Garzotto, A.; Rodriguez-Peralto, J.L.; Cortes-Funes, H.; Lopez-Martin, J.A.; et al. Prognostic value of dual-specificity phosphatase 6 expression in non-small cell lung cancer. Tumour. Biol. 2015, 36, 1199–1206. [Google Scholar] [CrossRef]
- Wang, T.L.; Ren, Y.W.; Wang, H.T.; Yu, H.; Zhao, Y.X. Association of Topoisomerase II (TOP2A) and Dual-Specificity Phosphatase 6 (DUSP6) Single Nucleotide Polymorphisms with Radiation Treatment Response and Prognosis of Lung Cancer in Han Chinese. Med. Sci. Monit. 2017, 23, 984–993. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.L.; Song, Y.Q.; Ren, Y.W.; Zhou, B.S.; Wang, H.T.; Bai, L.; Zhang, H.B.; Yu, H.; Zhao, Y.X. Dual Specificity Phosphatase 6 (DUSP6) Polymorphism Predicts Prognosis of Inoperable Non-Small Cell Lung Cancer after Chemoradiotherapy. Clin. Lab. 2016, 62, 301–310. [Google Scholar] [CrossRef]
- Alam, S.K.; Zhang, Y.; Wang, L.; Zhu, Z.; Hernandez, C.E.; Zhou, Y.; Yang, N.; Lei, J.; Chen, X.; Zeng, L.; et al. DARPP-32 promotes ERBB3-mediated resistance to molecular targeted therapy in EGFR-mutated lung adenocarcinoma. Oncogene 2022, 41, 83–98. [Google Scholar] [CrossRef]
- Pudelko, L.; Jaehrling, F.; Reusch, C.; Vitri, S.; Stroh, C.; Linde, N.; Sanderson, M.P.; Musch, D.; Lebrun, C.J.; Keil, M.; et al. SHP2 Inhibition Influences Therapeutic Response to Tepotinib in Tumors with MET Alterations. iScience 2020, 23, 101832. [Google Scholar] [CrossRef]
- Yan, M.; Wang, W.; Zhou, J.; Chang, M.; Peng, W.; Zhang, G.; Li, J.; Li, H.; Bai, C. Knockdown of PLAT enhances the anticancer effect of gefitinib in non-small cell lung cancer. J. Thorac. Dis. 2020, 12, 712–723. [Google Scholar] [CrossRef]
- Elia, I.; Broekaert, D.; Christen, S.; Boon, R.; Radaelli, E.; Orth, M.F.; Verfaillie, C.; Grunewald, T.G.P.; Fendt, S.M. Proline metabolism supports metastasis formation and could be inhibited to selectively target metastasizing cancer cells. Nat. Commun. 2017, 8, 15267. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Mao, C.; Wang, M.; Liu, N.; Ouyang, L.; Liu, S.; Tang, H.; Cao, Y.; Liu, S.; Wang, X.; et al. Cancer progression is mediated by proline catabolism in non-small cell lung cancer. Oncogene 2020, 39, 2358–2376. [Google Scholar] [CrossRef] [PubMed]
- Le, X.; Negrao, M.V.; Reuben, A.; Federico, L.; Diao, L.; McGrail, D.; Nilsson, M.; Robichaux, J.; Munoz, I.G.; Patel, S.; et al. Characterization of the Immune Landscape of EGFR-Mutant NSCLC Identifies CD73/Adenosine Pathway as a Potential Therapeutic Target. J. Thorac. Oncol. 2021, 16, 583–600. [Google Scholar] [CrossRef]
- Liu, H.; Kuang, X.; Zhang, Y.; Ye, Y.; Li, J.; Liang, L.; Xie, Z.; Weng, L.; Guo, J.; Li, H.; et al. ADORA1 Inhibition Promotes Tumor Immune Evasion by Regulating the ATF3-PD-L1 Axis. Cancer Cell 2020, 37, 324–339 e8. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Zhu, H.; Jiang, H.; Cui, Y.; Wang, M.; Ni, X.; Ma, C. The effects of aberrant expression of LncRNA DGCR5/miR-873-5p/TUSC3 in lung cancer cell progression. Cancer Med. 2018, 7, 3331–3341. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Shi, J.; Li, B.; Luo, M.; Xu, S.; Liu, X. LncRNA DGCR5 regulates the non-small cell lung cancer cell growth, migration, and invasion through regulating miR-211-5p/EPHB6 axis. Biofactors 2019, 45, 788–794. [Google Scholar] [CrossRef]
- Chen, E.G.; Zhang, J.S.; Xu, S.; Zhu, X.J.; Hu, H.H. Long non-coding RNA DGCR5 is involved in the regulation of proliferation, migration and invasion of lung cancer by targeting miR-1180. Am. J. Cancer Res. 2017, 7, 1463–1475. [Google Scholar] [PubMed]
- Dong, H.X.; Wang, R.; Jin, X.Y.; Zeng, J.; Pan, J. LncRNA DGCR5 promotes lung adenocarcinoma (LUAD) progression via inhibiting hsa-mir-22-3p. J. Cell Physiol. 2018, 233, 4126–4136. [Google Scholar] [CrossRef]
- Ren, W.; Mi, D.; Yang, K.; Cao, N.; Tian, J.; Li, Z.; Ma, B. The expression of hypoxia-inducible factor-1alpha and its clinical significance in lung cancer: A systematic review and meta-analysis. Swiss Med. Wkly. 2013, 143, w13855. [Google Scholar]
- Yang, S.L.; Ren, Q.G.; Wen, L.; Hu, J.L. Clinicopathological and prognostic significance of hypoxia-inducible factor-1 alpha in lung cancer: A systematic review with meta-analysis. J. Huazhong Univ. Sci. Technol. Med. Sci. 2016, 36, 321–327. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, N.; Liu, R.; Lv, H.; Li, Z.; Zhang, F.; Gai, C.; Tian, Z. Epigenetic Study of Esophageal Carcinoma Based on Methylation, Gene Integration and Weighted Correlation Network Analysis. OncoTargets Ther. 2021, 14, 3133–3149. [Google Scholar] [CrossRef]
- Xiang, R.; Ma, L.; Yang, M.; Zheng, Z.; Chen, X.; Jia, F.; Xie, F.; Zhou, Y.; Li, F.; Wu, K.; et al. Increased expression of peptides from non-coding genes in cancer proteomics datasets suggests potential tumor neoantigens. Commun. Biol. 2021, 4, 496. [Google Scholar] [CrossRef] [PubMed]
- Ni, C.; Yang, P.; Guo, J.; Ye, M. Role of DiGeorge syndrome critical region gene 9, a long noncoding RNA, in gastric cancer. OncoTargets Ther. 2018, 11, 2259–2267. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhang, X.; Wang, O.; Bi, Y.; Xing, X.; Cui, M.; Wang, M.; Tao, W.; Liao, Q.; Zhao, Y. The genomic profile of parathyroid carcinoma based on whole-genome sequencing. Int. J. Cancer 2020, 147, 2446–2457. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.R.; Chen, Z.; Faden, D.L.; Agrawal, N.; Li, R.J.; Hanna, G.J.; Iyer, N.G.; Boot, A.; Rozen, S.G.; Vettore, A.L.; et al. The mutational landscape of early- and typical-onset oral tongue squamous cell carcinoma. Cancer 2021, 127, 544–553. [Google Scholar] [CrossRef]
- Zou, H.; Zhu, N.; Li, S. The emerging role of dipeptidyl-peptidase-4 as a therapeutic target in lung disease. Expert Opin. Ther. Targets 2020, 24, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Ban, H.S.; Uto, Y.; Nakamura, H. Hypoxia-inducible factor (HIF) inhibitors: A patent survey (2016–2020). Expert Opin. Ther. Pat. 2021, 31, 387–397. [Google Scholar] [CrossRef]
- Zhang, T.; Tong, X.; Zhang, S.; Wang, D.; Wang, L.; Wang, Q.; Fan, H. The Roles of Dipeptidyl Peptidase 4 (DPP4) and DPP4 Inhibitors in Different Lung Diseases: New Evidence. Front. Pharmacol. 2021, 12, 731453. [Google Scholar] [CrossRef]
- Sangaletti, S.; Ferrara, R.; Tripodo, C.; Garassino, M.C.; Colombo, M.P. Myeloid cell heterogeneity in lung cancer: Implication for immunotherapy. Cancer Immunol. Immunother. 2021, 70, 2429–2438. [Google Scholar] [CrossRef]
- Wang, J.B.; Huang, X.; Li, F.R. Impaired dendritic cell functions in lung cancer: A review of recent advances and future perspectives. Cancer Commun. 2019, 39, 43. [Google Scholar] [CrossRef] [Green Version]
- Gardner, A.; de Mingo Pulido, A.; Ruffell, B. Dendritic Cells and Their Role in Immunotherapy. Front. Immunol. 2020, 11, 924. [Google Scholar] [CrossRef]
- Kiss, M.; Caro, A.A.; Raes, G.; Laoui, D. Systemic Reprogramming of Monocytes in Cancer. Front. Oncol. 2020, 10, 1399. [Google Scholar] [CrossRef] [PubMed]
- Leveque, E.; Rouch, A.; Syrykh, C.; Mazieres, J.; Brouchet, L.; Valitutti, S.; Espinosa, E.; Lafouresse, F. Phenotypic and Histological Distribution Analysis Identify Mast Cell Heterogeneity in Non-Small Cell Lung Cancer. Cancers 2022, 14, 1394. [Google Scholar] [CrossRef] [PubMed]
- Sheida, F.; Razi, S.; Keshavarz-Fathi, M.; Rezaei, N. The role of myeloid-derived suppressor cells in lung cancer and targeted immunotherapies. Expert Rev. Anticancer Ther. 2022, 22, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Noyes, D.; Bag, A.; Oseni, S.; Semidey-Hurtado, J.; Cen, L.; Sarnaik, A.A.; Sondak, V.K.; Adeegbe, D. Tumor-associated Tregs obstruct antitumor immunity by promoting T cell dysfunction and restricting clonal diversity in tumor-infiltrating CD8+ T cells. J. Immunother. Cancer 2022, 10, e004605. [Google Scholar] [CrossRef] [PubMed]
- Yuan, A.; Hsiao, Y.J.; Chen, H.Y.; Chen, H.W.; Ho, C.C.; Chen, Y.Y.; Liu, Y.C.; Hong, T.H.; Yu, S.L.; Chen, J.J.; et al. Opposite Effects of M1 and M2 Macrophage Subtypes on Lung Cancer Progression. Sci. Rep. 2015, 5, 14273. [Google Scholar] [CrossRef] [Green Version]
- La Fleur, L.; Botling, J.; He, F.; Pelicano, C.; Zhou, C.; He, C.; Palano, G.; Mezheyeuski, A.; Micke, P.; Ravetch, J.V.; et al. Targeting MARCO and IL37R on Immunosuppressive Macrophages in Lung Cancer Blocks Regulatory T Cells and Supports Cytotoxic Lymphocyte Function. Cancer Res. 2021, 81, 956–967. [Google Scholar] [CrossRef]
- Mei, Y.; Liu, H. IL-37: An anti-inflammatory cytokine with antitumor functions. Cancer Rep. 2019, 2, e1151. [Google Scholar] [CrossRef]
- Ge, G.; Wang, A.; Yang, J.; Chen, Y.; Yang, J.; Li, Y.; Xue, Y. Interleukin-37 suppresses tumor growth through inhibition of angiogenesis in non-small cell lung cancer. J. Exp. Clin. Cancer Res. 2016, 35, 13. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.H.; Zhou, B.Y.; Wu, X.J.; Xu, J.F.; Zhang, J.A.; Chen, Y.H.; Liang, S.S. [Corrigendum] CCL22 and IL37 inhibit the proliferation and epithelialmesenchymal transition process of NSCLC A549 cells. Oncol. Rep. 2021, 45, 786. [Google Scholar] [CrossRef]
- Jiang, M.; Wang, Y.; Zhang, H.; Ji, Y.; Zhao, P.; Sun, R.; Zhang, C. IL-37 inhibits invasion and metastasis in non-small cell lung cancer by suppressing the IL-6/STAT3 signaling pathway. Thorac. Cancer 2018, 9, 621–629. [Google Scholar] [CrossRef]
- Kim, M.S.; Baek, A.R.; Lee, J.H.; Jang, A.S.; Kim, D.J.; Chin, S.S.; Park, S.W. IL-37 Attenuates Lung Fibrosis by Inducing Autophagy and Regulating TGF-beta1 Production in Mice. J. Immunol. 2019, 203, 2265–2275. [Google Scholar] [CrossRef] [PubMed]
- Christodoulou, M.I.; Kapsogeorgou, E.K.; Moutsopoulos, H.M. Characteristics of the minor salivary gland infiltrates in Sjogren’s syndrome. J. Autoimmun. 2010, 34, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Christodoulou, M.I.; Kapsogeorgou, E.K.; Moutsopoulos, N.M.; Moutsopoulos, H.M. Foxp3+ T-regulatory cells in Sjogren’s syndrome: Correlation with the grade of the autoimmune lesion and certain adverse prognostic factors. Am. J. Pathol. 2008, 173, 1389–1396. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
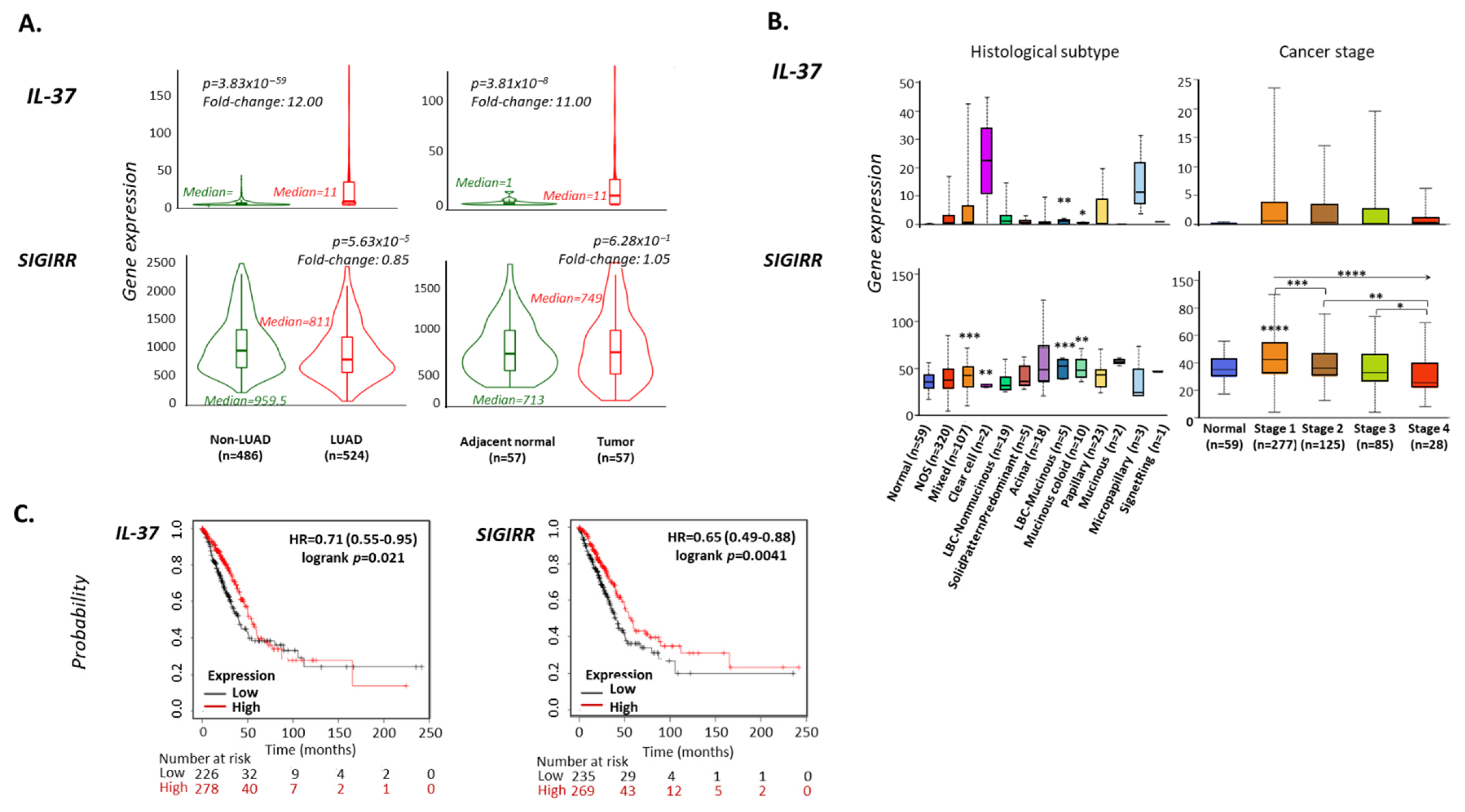
| Human Cancer Type | No of Patients | Gene Expression Levels | |||
|---|---|---|---|---|---|
| IL-37 | SIGIRR | ||||
| Mean FPKM ± SD | p-Value Compared to Lung/LUAD | Mean FPKM ± SD | p-Value Compared to Lung/LUAD | ||
| Breast | 1075 | 0.07 ± 0.58 | <0.0001/<0.0001 | 14.04 ± 10.16 | <0.0001/NS |
| Cervical | 291 | 0.17 ± 0.19 | <0.0001/<0.0001 | 12.77 ± 8.29 | <0.0001/0.0006 |
| Colorectal | 597 | 0.68 ± 2.38 | 0.09/<0.0001 | 17.64 ± 7.15 | <0.0001/<0.0001 |
| Endometrial | 541 | 0.19 ± 1.45 | <0.0001/<0.0001 | 22.52 ± 10.64 | <0.0001/<0.0001 |
| Glioma | 153 | 0.05 ± 0.07 | <0.0001/<0.0001 | 2.56 ± 1.68 | <0.0001/<0.0001 |
| Head-and-neck | 499 | 0.18 ± 0.49 | <0.0001/<0.0001 | 5.04 ± 4.21 | <0.0001/<0.0001 |
| Liver | 365 | 0.03 ± 0.07 | <0.0001/<0.0001 | 14.02 ± 12.76 | <0.0001/0.0008 |
| Lung | 994 | 6.59 ± 31.88 | NA/NA | 10.30 ± 6.78 | NA/NA |
| LUAD | 500 | 12.85 ± 44.05 | NA/NA | 13.64 ± 7.03 | NA/NA |
| LUSC | 494 | 2.56 ± 17.89 | NA/0.0001 | 6.92 ± 4.46 | NA/<0.0001 |
| Melanoma | 102 | 0.83 ± 2.11 | 0.0002/<0.0001 | 10.96 ± 6.44 | NS/<0.0001 |
| Ovarian | 373 | 0.10 ± 0.15 | <0.0001/<0.0001 | 16.70 ± 8.41 | <0.0001/<0.0001 |
| Pancreatic | 176 | 0.34 ± 0.54 | NS/<0.0001 | 15.54 ± 7.74 | <0.0001/0.005 |
| Prostate | 494 | 0.01 ± 0.05 | <0.0001/<0.0001 | 15.57 ± 6.26 | <0.0001/<0.0001 |
| Renal | 877 | 0.02 ± 0.06 | <0.0001/<0.0001 | 9.94 ± 6.93 | 0.05/<0.0001 |
| Stomach | 354 | 0.35 ± 1.21 | 0.0001/<0.0001 | 15.17 ± 5.94 | <0.0001/<0.0001 |
| Testis | 134 | 1.08 ± 2.27 | NS/NS | 12.25 ± 5.31 | <0.0001/0.08 |
| Thyroid | 501 | 0.25 ± 1.24 | <0.0001/<0.0001 | 11.02 ± 4.66 | <0.0001/<0.0001 |
| Urothelial | 406 | 0.67 ± 8.58 | <0.0001/<0.0001 | 11.40 ± 7.3 | 0.006/<0.0001 |
| IL-37 | SIGIRR | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Non-LUAD vs. LUAD | |||||||||
| n | Levels (Median; Range) | Fold-Change | p-Value | n | Levels (Median; Range) | Fold-Change | p-Value | Platform | |
| Non-LUAD | 486 | 1 | 486 | 959.5 | TNMplot | ||||
| LUAD | 524 | 11 | 12 | 3.83 × 10−59 | 524 | 811 | 0.85 | 5.63 × 10−5 | TNMplot |
| Normal | 59 | 0.08 (0–0.38) | 59 | 35.06 (17.01–55.55) | UACLAN | ||||
| Primary tumor | 515 | 0.43 (0–19.6) | 5.38 | 5.97 × 10−11 | 515 | 38.71 (3.84–84.81) | 1.11 | 9.20 × 10−7 | UACLAN |
| Tumors vs. adjacent Normal | |||||||||
| Adjacent normal | 57 | 1 | 57 | 713 | TNMplot | ||||
| Tumor | 57 | 11 | 11 | 3.81 × 10−8 | 57 | 749 | 1.05 | 6.28 × 10−1 | TNMplot |
| LUAD histological subtype | |||||||||
| Normal | 59 | 0.08 (0–0.38) | 59 | 35.06 (17.01–55.55) | UACLAN | ||||
| NOS | 320 | 0.42 (0–16.99) | 5.25 | NS | 320 | 37.08 (3.84–84.81) | 1.06 | NS | UACLAN |
| Mixed | 107 | 0.7 (0–42.55) | 5.6 | NS | 107 | 42.19 (9.77–71.23) | 1.20 | 0.0003 | UACLAN |
| Clear cell | 2 | 22.40 (0–44.79) | 280 | NS | 2 | 30.54 (29.11–31.98) | 0.87 | 0.0022 | UACLAN |
| LBC-Nonmucinous | 19 | 1.08 (0–14.61) | 13.5 | NS | 19 | 31.81 (24.68–59.76) | 0.90 | NS | UACLAN |
| Solid pattern predominant | 5 | 0.10 (0–3.13) | 1.25 | NS | 5 | 35.81 (27.55–62.29) | 1.02 | NS | UACLAN |
| Acinar | 18 | 0.34 (0–9.66) | 4.25 | NS | 18 | 48.39 (20.35–122.59) | 1.38 | NS | UACLAN |
| LBC-Mucinous | 5 | 1.07 (0.20–1.96) | 13.38 | 0.0072 | 5 | 52.32 (37.87–60.69) | 1.49 | 0.0008 | UACLAN |
| Mucinous colloid | 10 | 0.26 (0–0.76) | 3.25 | 0.0153 | 10 | 47.72 (35.68–70.80) | 1.36 | 0.0011 | UACLAN |
| Papillary | 23 | 0.30 (0–19.55) | 3.75 | NS | 23 | 43.01 (23.36–70.09) | 1.23 | NS | UACLAN |
| Mucinous | 2 | 0.09 (0–0.17) | 1.13 | NS | 2 | 56.86 (52.91–60.81) | 1.62 | NS | UACLAN |
| Micropapillary | 3 | 11.24 (3.76–31.37) | 140.05 | NS | 3 | 23.67 (21.42–73.27) | 0.67 | NS | UACLAN |
| Signet ring | 1 | 0.82 (0.82–0.82) | 10.25 | NA | 1 | 45.86 (45.86–45.86) | 1.30 | NS | UACLAN |
| Cancer stage | |||||||||
| Normal | 59 | 0.08 (0–0.38) | 59 | 35.06 (17.01–55.55) | UACLAN | ||||
| Stage 1 | 277 | 28 (0–23.62) | 7.63 | NS | 277 | 42.53 (3.97–89.73) | 1.21 | <0.0001 | UACLAN |
| Stage 2 | 125 | 0.37 (0–13.55) | 4.63 | NS | 125 | 36.16 (12.59–75.62) | 1.03 | NS | UACLAN |
| Stage 3 | 85 | 0.17 (0–19.55) | 2.13 | NS | 85 | 33.03 (3.84–74.01) | 0.94 | NS | UACLAN |
| Stage 4 | 28 | 0.35 (0–6.17) | 4.38 | NS | 28 | 25.65 (7.76–69.02) | 0.73 | NS | UACLAN |
| Linear trend Stage 1→4 | NS | <0.0001 | |||||||
| IL-37 | SIGIRR | ||||||
|---|---|---|---|---|---|---|---|
| Non-LUAD vs. LUAD | |||||||
| n | Levels (Median; Range) | p-Value | n | Levels (Median; Range) | p-Value | Platform | |
| Normal | 111 | 0.11 (−1.02–1.16) | 111 | −0.25 (−1.14–0.38) | UACLAN | ||
| Primary tumor | 111 | −0.043 (−1.02–1.16) | NS | 111 | 0 (−1.91–2.26) | 2.82 × 10−2 | UACLAN |
| Grade | |||||||
| Normal | 111 | 0.11 (−1.02–1.16) | 111 | −0.25 (−1.14–0.38) | UACLAN | ||
| Grade 1 | 7 | 0.22 (0.21–0.24) | 0.0313 | 7 | 0.93 (−0.23–1.35) | 0.001 | UACLAN |
| Grade 2 | 59 | −0.28 (−0.75–1.07) | 0.0351 | 59 | 0.35 (−1.55–2.26) | NS | UACLAN |
| Grade 3 | 39 | 0.66 (−0.68–1.90) | <0.0001 | 39 | −0.48 (−1.57–1.26) | <0.0001 | UACLAN |
| Linear trend Grade 1 → 3 | NS | 0.0001 | |||||
| EPIC | TIMER | MCP-COUNTER | QUANTISEQ | CIBERSORT | CIBERSORT-ABS | XCELL | TIDE | |
|---|---|---|---|---|---|---|---|---|
| CD8+ T cell | −0.007; NS | 0.032; NS | −0.029; NS | −0.002; NS | −0.187; 3 × 10−5 | −0.051; NS | 0.026; NS | NA |
| Naïve | NA | NA | NA | NA | NA | NA | −0.03 | NA |
| Memory | NA | NA | NA | NA | NA | NA | 0.069 | NA |
| Central memory | NA | NA | NA | NA | NA | NA | −0.034; NS | NA |
| CD4+ T cell | 0.006; NS | 0.06; NS | −0.168 *; 2 × 10−4 | NA | NA | 0.088 *; NS | NA | |
| Naïve | NA | NA | NA | NA | 0.019; NS | 0.019; NS | 0.126; 5 × 10−3 | NA |
| Memory | NA | NA | NA | NA | NA | NA | −0.019; NS | NA |
| Central memory | NA | NA | NA | NA | NA | NA | 0.04; NS | NA |
| Effector memory | NA | NA | NA | NA | NA | NA | 0.384; 8 × 10−20 | NA |
| Memory activated | NA | NA | NA | NA | −0.163; 3 × 10−4 | −0.158; 4 × 10−4 | NA | NA |
| Memory resting | NA | NA | NA | NA | 0.208; 3 × 10−6 | 0.259; 5 × 10−9 | NA | NA |
| Th1 | NA | NA | NA | NA | NA | NA | −0.103; 3 × 10−2 | NA |
| Th2 | NA | NA | NA | NA | NA | NA | −0.323; 2 × 10−13 | NA |
| Tregs | NA | NA | NA | 0.241; 6 × 10−8 | 0.119; 6 × 10−3 | 0.167; 2 × 10−4 | NA | NA |
| B cell | −0.017; NS | 0.126; 5 × 10−3 | 0.052; NS | −0.026; NS | NA | NA | 0.066; NS | NA |
| Naïve | NA | NA | NA | NA | −0.103; 2 × 10−2 | −0.061; NS | 0.047; NS | NA |
| Memory | 0.26; NS | NA | NA | NA | 0.202; 6 × 10−6 | 0.204; 5 × 10−6 | −0.031; NS | NA |
| Neutrophil | NA | 0.033; NS | 0.145; 1 × 10−3 | 0.311; 2 × 10−12 | −0.032; NS | −0.011; NS | −0.146; 1 × 10−3 | NA |
| Monocyte | NA | NA | −0.056; NS | −0.029; NS | 0.207; 3 × 10−6 | 0.226; 4 × 10−7 | 0.26; 4 × 10−9 | NA |
| Macrophage | 0.137; 2 × 10−3 | −0.01; NS | −0.056; NS | NA | −0.147; 1 × 10−3 | −0.113; 1 × 10−2 | 0.12; 8 × 10−3 | NA |
| M1 | NA | NA | NA | 0.388; 4 × 10−19 | −0.109; 2 × 10−2 | −0.018; NS | −0.014; NS | NA |
| M2 | NA | NA | NA | 0.279; 3 × 10−10 | 0.21; 3 × 10−6 | 0.251; 2 × 10−8 | 0.166; 2 × 10−4 | 0.047; NS |
| Dendritic cell | ||||||||
| Myeloid | NA | NA | 0.406; 5 × 10−21 | −0.141; NS | NA | NA | 0.42; 2 × 10−22 | 0.307; 4 × 10−12 |
| Myeloid activated | NA | NA | NA | NA | 0.101; 2 × 10−2 | 0.138; 2 × 10−3 | 0.316; 7 × 10−13 | NA |
| Myeloid resting | NA | NA | NA | NA | NA | NA | NA | NA |
| Plasmacytoid | NA | NA | NA | NA | NA | NA | −0.124; 6 × 10−3 | NA |
| Natural killer | −0.086; NS | NA | −0.239; 8 × 10−8 | 0.16; 7 × 10−4 | NA | NA | −0.018; NS | NA |
| Activated | NA | NA | NA | NA | −0.188; 3 × 10−5 | −0.064; NS | NA | NA |
| Resting | NA | NA | NA | NA | −0.078; NS | −0.077; NS | NA | NA |
| Mast cell | NA | NA | NA | NA | NA | NA | 0.201; 7 × 10−6 | NA |
| Activated | NA | NA | NA | NA | 0.272; 9 × 10−10 | 0.303; 6 × 10−12 | NA | NA |
| Resting | NA | NA | NA | NA | −0.159; 4 × 10−4 | −0.138; 2 × 10−3 | NA | NA |
| Cancer-associated fibroblast | 0.065; NS | NA | 0.045; NS | NA | NA | NA | 0.211; 2 × 10−6 | 0.063; NS |
| Common lymphoid progenitor | NA | NA | NA | NA | NA | NA | −0.14; 2 × 10−3 | NA |
| Common myeloid progenitor | NA | NA | NA | NA | NA | NA | 0.092; NS | NA |
| Endothelial cell | −0.007 | NA | 0.109; 2 × 10−3 | NA | NA | NA | −0.062; NS | NA |
| Eosinophil | NA | NA | NA | NA | 0.067 | NA | 0.067; NS | 0.005; NS |
| Granulocyte-lymphocyte progenitor | NA | NA | NA | NA | NA | NA | 0.323; 2 × 10−13 | NA |
| Hematopoieitic stem cell | NA | NA | NA | NA | NA | NA | 0.159; 4 × 10−4 | NA |
| follicular helper T cell | NA | NA | NA | NA | −0.187; NS | NA | NA | NA |
| γδ T cell | NA | NA | NA | NA | −0.021; NS | −0.02; NS | 0.025; NS | NA |
| NK Τ cell | NA | NA | NA | NA | NA | NA | 0.43; NS | NA |
| MDSCs | NA | NA | NA | NA | NA | NA | NA | −0.292; 4 × 10−11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Christodoulou, P.; Kyriakou, T.-C.; Boutsikos, P.; Andreou, M.; Ji, Y.; Xu, D.; Papageorgis, P.; Christodoulou, M.-I. Aberrant Expression and Prognostic Potential of IL-37 in Human Lung Adenocarcinoma. Biomedicines 2022, 10, 3037. https://doi.org/10.3390/biomedicines10123037
Christodoulou P, Kyriakou T-C, Boutsikos P, Andreou M, Ji Y, Xu D, Papageorgis P, Christodoulou M-I. Aberrant Expression and Prognostic Potential of IL-37 in Human Lung Adenocarcinoma. Biomedicines. 2022; 10(12):3037. https://doi.org/10.3390/biomedicines10123037
Chicago/Turabian StyleChristodoulou, Panayiota, Theodora-Christina Kyriakou, Panagiotis Boutsikos, Maria Andreou, Yuan Ji, Damo Xu, Panagiotis Papageorgis, and Maria-Ioanna Christodoulou. 2022. "Aberrant Expression and Prognostic Potential of IL-37 in Human Lung Adenocarcinoma" Biomedicines 10, no. 12: 3037. https://doi.org/10.3390/biomedicines10123037