
OGG1 Inhibition Reduces Acinar Cell Injury in a Mouse Model of Acute Pancreatitis
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
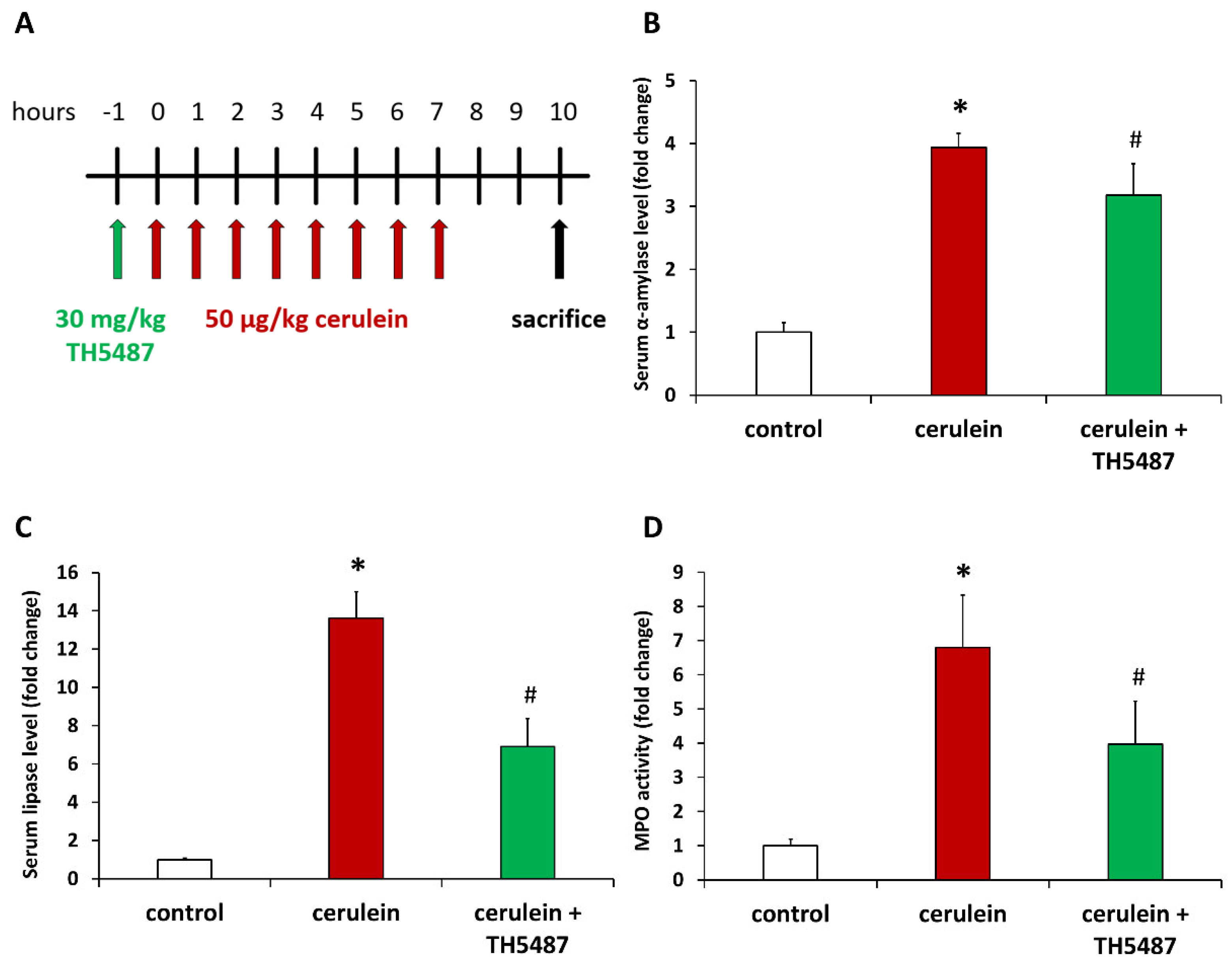
2.2. Acute Pancreatitis Model
2.3. Isolation and Culture of Pancreatic Acinar Cells
2.4. Serum α-Amylase and Lipase Determination
2.5. Myeloperoxidase Assay
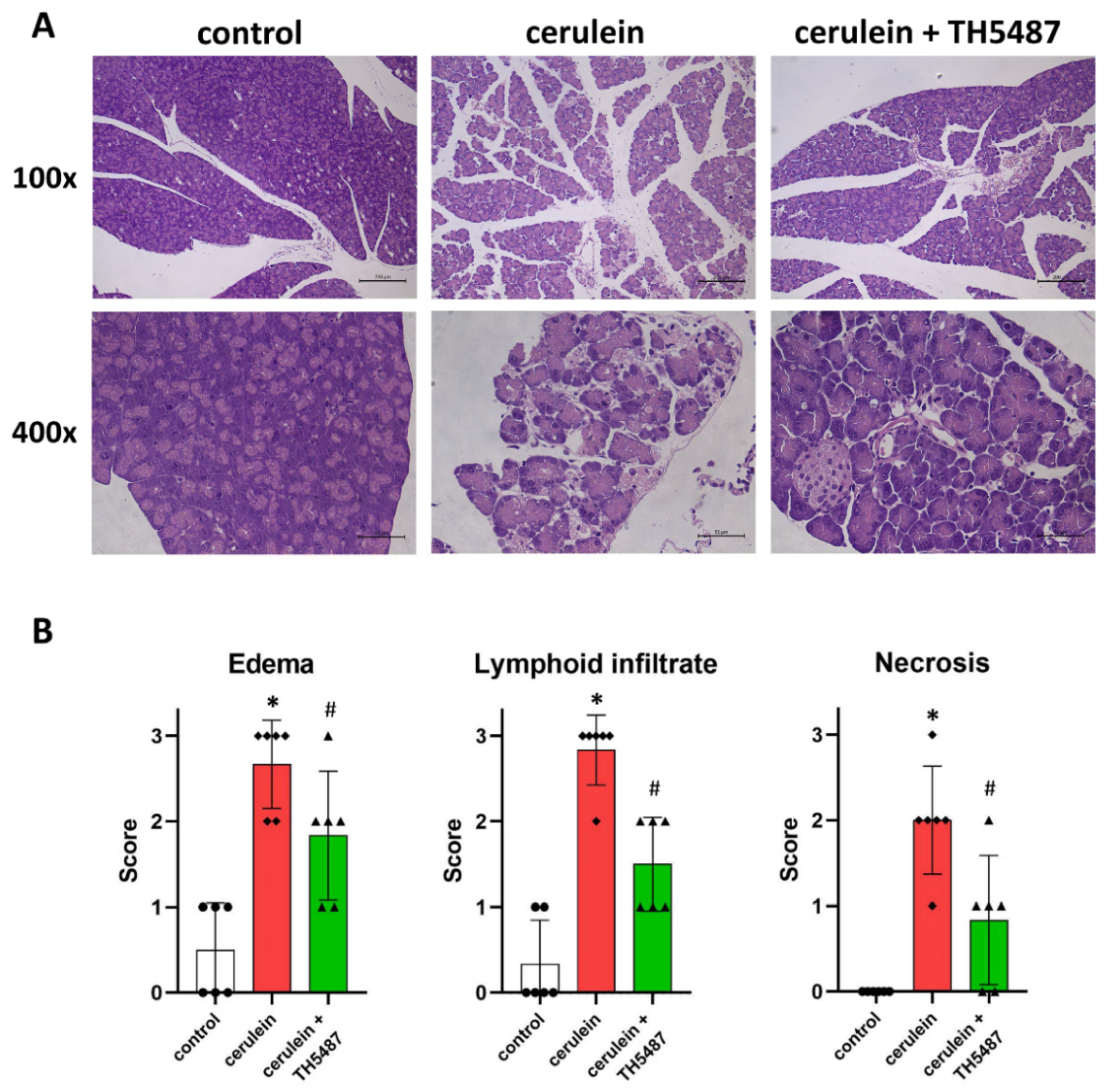
2.6. Histology
2.7. Immunohistochemistry
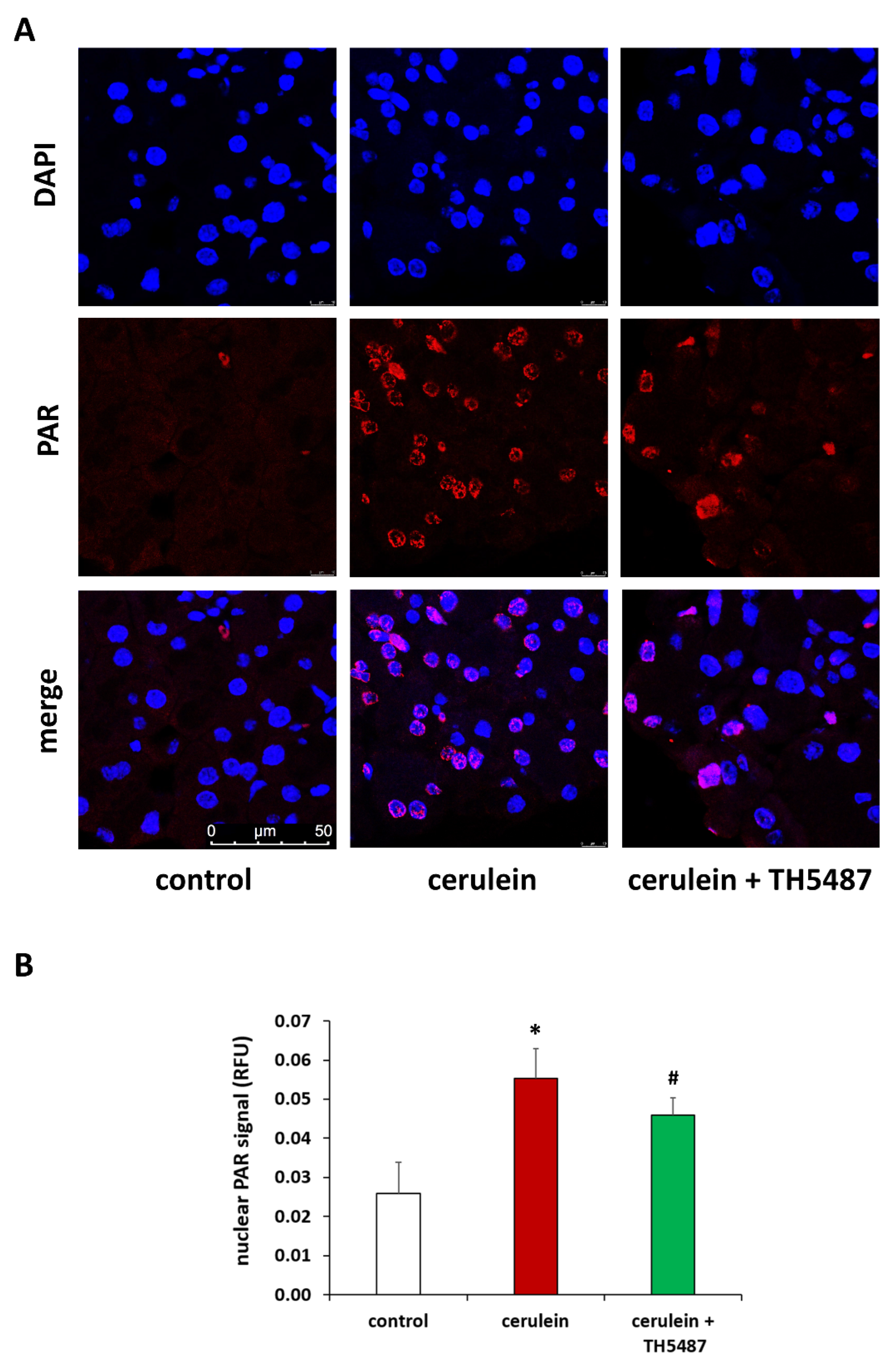
2.8. Immunofluorescent Staining
2.9. Cytokine Array
2.10. RNA Isolation and Quantitative Real-Time PCR
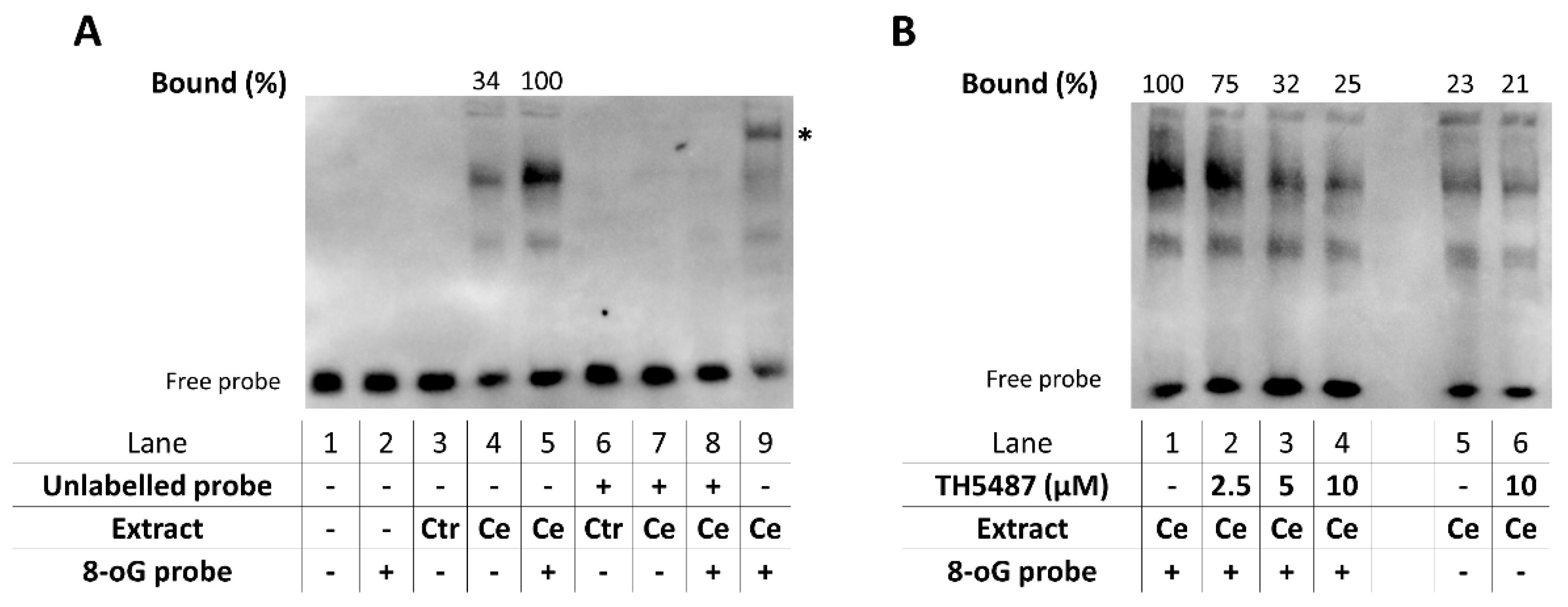
2.11. Electrophoretic Mobility Shift Assay (EMSA)
2.12. Apoptosis, Necrosis and Viability Assays
2.13. Comet Assay
2.14. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiao, A.Y.; Tan, M.L.; Wu, L.M.; Asrani, V.M.; Windsor, J.A.; Yadav, D.; Petrov, M.S. Global incidence and mortality of pancreatic diseases: A systematic review, meta-analysis, and meta-regression of population-based cohort studies. Lancet Gastroenterol. Hepatol. 2016, 1, 45–55. [Google Scholar] [CrossRef]
- Mergener, K.; Baillie, J. Acute pancreatitis. BMJ 1998, 316, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Criddle, D.N. Reactive oxygen species, Ca2+ stores and acute pancreatitis; a step closer to therapy? Cell Calcium 2016, 60, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Mossner, J. New advances in cell physiology and pathophysiology of the exocrine pancreas. Dig. Dis. 2010, 28, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, P.; Petersen, O.H. The exocrine pancreas: The acinar-ductal tango in physiology and pathophysiology. Rev. Physiol. Biochem. Pharmacol. 2013, 165, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, P.; Rakonczay, Z. Insufficiency of electrolyte and fluid secretion by pancreatic ductal cells leads to increased patient risk for pancreatitis. Am. J. Gastroenterol. 2010, 105, 2119–2120. [Google Scholar] [CrossRef]
- Hegyi, P.; Rakonczay, Z., Jr. The role of pancreatic ducts in the pathogenesis of acute pancreatitis. Pancreatology 2015, 15, S13–S17. [Google Scholar] [CrossRef]
- Padureanu, V.; Florescu, D.N.; Padureanu, R.; Ghenea, A.E.; Gheonea, D.I.; Oancea, C.N. Role of antioxidants and oxidative stress in the evolution of acute pancreatitis (Review). Exp. Ther. Med. 2022, 23, 197. [Google Scholar] [CrossRef]
- Yudkina, A.V.; Shilkin, E.S.; Endutkin, A.V.; Makarova, A.V.; Zharkov, D.O. Reading and Misreading 8-oxoguanine, a Paradigmatic Ambiguous Nucleobase. Crystals 2019, 9, 269. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Bosch, N.; Fernandez-Zapico, M.E.; Navarro, P.; Yelamos, J. Poly(ADP-Ribose) Polymerases: New Players in the Pathogenesis of Exocrine Pancreatic Diseases. Am. J. Pathol. 2016, 186, 234–241. [Google Scholar] [CrossRef]
- Mota, R.; Sanchez-Bueno, F.; Berenguer-Pina, J.J.; Hernandez-Espinosa, D.; Parrilla, P.; Yelamos, J. Therapeutic treatment with poly(ADP-ribose) polymerase inhibitors attenuates the severity of acute pancreatitis and associated liver and lung injury. Br. J. Pharmacol. 2007, 151, 998–1005. [Google Scholar] [CrossRef] [PubMed]
- Mota, R.A.; Sanchez-Bueno, F.; Saenz, L.; Hernandez-Espinosa, D.; Jimeno, J.; Tornel, P.L.; Martinez-Torrano, A.; Ramirez, P.; Parrilla, P.; Yelamos, J. Inhibition of poly(ADP-ribose) polymerase attenuates the severity of acute pancreatitis and associated lung injury. Lab. Investig. 2005, 85, 1250–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, J.W.; Hazra, T.K.; Izumi, T.; Mitra, S. Stimulation of human 8-oxoguanine-DNA glycosylase by AP-endonuclease: Potential coordination of the initial steps in base excision repair. Nucleic Acids Res. 2001, 29, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, C.; Qiao, P.; Xue, Y.; Zheng, X.; Chen, H.; Zeng, X.; Liu, W.; Boldogh, I.; Ba, X. OGG1-initiated base excision repair exacerbates oxidative stress-induced parthanatos. Cell Death Dis. 2018, 9, 628. [Google Scholar] [CrossRef]
- Gedik, C.M.; Collins, A.; ESCODD. Establishing the background level of base oxidation in human lymphocyte DNA: Results of an interlaboratory validation study. FASEB J. 2005, 19, 82–84. [Google Scholar] [CrossRef] [Green Version]
- Helbock, H.J.; Beckman, K.B.; Ames, B.N. 8-Hydroxydeoxyguanosine and 8-hydroxyguanine as biomarkers of oxidative DNA damage. Methods Enzymol. 1999, 300, 156–166. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2′-deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef] [Green Version]
- Mabley, J.G.; Pacher, P.; Deb, A.; Wallace, R.; Elder, R.H.; Szabo, C. Potential role for 8-oxoguanine DNA glycosylase in regulating inflammation. FASEB J. 2005, 19, 290–292. [Google Scholar] [CrossRef]
- Bacsi, A.; Aguilera-Aguirre, L.; Szczesny, B.; Radak, Z.; Hazra, T.K.; Sur, S.; Ba, X.; Boldogh, I. Down-regulation of 8-oxoguanine DNA glycosylase 1 expression in the airway epithelium ameliorates allergic lung inflammation. DNA Repair 2013, 12, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Aguilera-Aguirre, L.; Bacsi, A.; Radak, Z.; Hazra, T.K.; Mitra, S.; Sur, S.; Brasier, A.R.; Ba, X.; Boldogh, I. Innate inflammation induced by the 8-oxoguanine DNA glycosylase-1-KRAS-NF-kappaB pathway. J. Immunol. 2014, 193, 4643–4653. [Google Scholar] [CrossRef]
- Hegyi, P.; Venglovecz, V.; Pallagi, P.; Maleth, J.; Takacs, T.; Rakonczay, Z., Jr. Galanin, a potent inhibitor of pancreatic bicarbonate secretion, is involved in the induction and progression of cerulein-induced experimental acute pancreatitis. Pancreas 2011, 40, 155–156. [Google Scholar] [CrossRef] [PubMed]
- Nagy-Penzes, M.; Hajnady, Z.; Regdon, Z.; Demeny, M.A.; Kovacs, K.; El-Hamoly, T.; Maleth, J.; Hegyi, P.; Hegedus, C.; Virag, L. Tricetin Reduces Inflammation and Acinar Cell Injury in Cerulein-Induced Acute Pancreatitis: The Role of Oxidative Stress-Induced DNA Damage Signaling. Biomedicines 2022, 10, 1371. [Google Scholar] [CrossRef] [PubMed]
- Visnes, T.; Cazares-Korner, A.; Hao, W.; Wallner, O.; Masuyer, G.; Loseva, O.; Mortusewicz, O.; Wiita, E.; Sarno, A.; Manoilov, A.; et al. Small-molecule inhibitor of OGG1 suppresses proinflammatory gene expression and inflammation. Science 2018, 362, 834–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virag, L.; Bai, P.; Bak, I.; Pacher, P.; Mabley, J.G.; Liaudet, L.; Bakondi, E.; Gergely, P.; Kollai, M.; Szabo, C. Effects of poly(ADP-ribose) polymerase inhibition on inflammatory cell migration in a murine model of asthma. Med. Sci. Monit. 2004, 10, BR77–BR83. [Google Scholar]
- Histological and Histochemical Methods: Theory and Practice, 5th ed.; Scion Publishing: Banbury, UK, 2015.
- Hanna, B.M.F.; Helleday, T.; Mortusewicz, O. OGG1 Inhibitor TH5487 Alters OGG1 Chromatin Dynamics and Prevents Incisions. Biomolecules 2020, 10, 1483. [Google Scholar] [CrossRef]
- Regdon, Z.; Robaszkiewicz, A.; Kovacs, K.; Rygielska, Z.; Hegedus, C.; Bodoor, K.; Szabo, E.; Virag, L. LPS protects macrophages from AIF-independent parthanatos by downregulation of PARP1 expression, induction of SOD2 expression, and a metabolic shift to aerobic glycolysis. Free Radic. Biol. Med. 2019, 131, 184–196. [Google Scholar] [CrossRef]
- Regdon, Z.; Demeny, M.A.; Kovacs, K.; Hajnady, Z.; Nagy-Penzes, M.; Bakondi, E.; Kiss, A.; Hegedus, C.; Virag, L. High-content screening identifies inhibitors of oxidative stress-induced parthanatos: Cytoprotective and anti-inflammatory effects of ciclopirox. Br. J. Pharmacol. 2020, 178, 1095–1113. [Google Scholar] [CrossRef]
- Mazzon, E.; Genovese, T.; Di Paola, R.; Muia, C.; Crisafulli, C.; Malleo, G.; Esposito, E.; Meli, R.; Sessa, E.; Cuzzocrea, S. Effects of 3-aminobenzamide, an inhibitor of poly (ADP-ribose) polymerase, in a mouse model of acute pancreatitis induced by cerulein. Eur. J. Pharmacol. 2006, 549, 149–156. [Google Scholar] [CrossRef]
- Erdelyi, K.; Bakondi, E.; Gergely, P.; Szabo, C.; Virag, L. Pathophysiologic role of oxidative stress-induced poly(ADP-ribose) polymerase-1 activation: Focus on cell death and transcriptional regulation. Cell. Mol. Life Sci. 2005, 62, 751–759. [Google Scholar] [CrossRef]
- Virag, L.; Robaszkiewicz, A.; Rodriguez-Vargas, J.M.; Oliver, F.J. Poly(ADP-ribose) signaling in cell death. Mol. Aspects Med. 2013, 34, 1153–1167. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Bonham, M.J.; Buss, H.; Abu-Zidan, F.M.; Windsor, J.A. Elevated protein carbonyls as plasma markers of oxidative stress in acute pancreatitis. Pancreatology 2003, 3, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.; Perez-Gonzalez, N.; Llanillo, M.; Calvo, J.J.; Sanchez-Bernal, C. Acute pancreatitis decreases pancreas phospholipid levels and increases susceptibility to lipid peroxidation in rat pancreas. Lipids 2002, 37, 167–171. [Google Scholar] [CrossRef]
- Bakondi, E.; Singh, S.B.; Hajnady, Z.; Nagy-Penzes, M.; Regdon, Z.; Kovacs, K.; Hegedus, C.; Madacsy, T.; Maleth, J.; Hegyi, P.; et al. Spilanthol Inhibits Inflammatory Transcription Factors and iNOS Expression in Macrophages and Exerts Anti-inflammatory Effects in Dermatitis and Pancreatitis. Int. J. Mol. Sci. 2019, 20, 4308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Tang, X.; Ke, X.; Dai, Y.; Shi, J. Triptolide Suppresses NF-kappaB-Mediated Inflammatory Responses and Activates Expression of Nrf2-Mediated Antioxidant Genes to Alleviate Caerulein-Induced Acute Pancreatitis. Int. J. Mol. Sci. 2022, 23, 1252. [Google Scholar] [CrossRef] [PubMed]
- Bansod, S.; Godugu, C. Nimbolide ameliorates pancreatic inflammation and apoptosis by modulating NF-kappaB/SIRT1 and apoptosis signaling in acute pancreatitis model. Int. Immunopharmacol. 2021, 90, 107246. [Google Scholar] [CrossRef]
- Khurana, A.; Anchi, P.; Allawadhi, P.; Kumar, V.; Sayed, N.; Packirisamy, G.; Godugu, C. Superoxide dismutase mimetic nanoceria restrains cerulein induced acute pancreatitis. Nanomedicine 2019, 14, 1805–1825. [Google Scholar] [CrossRef]
- Lee, J.H.; An, C.S.; Yun, B.S.; Kang, K.S.; Lee, Y.A.; Won, S.M.; Gwag, B.J.; Cho, S.I.; Hahm, K.B. Prevention effects of ND-07, a novel drug candidate with a potent antioxidative action and anti-inflammatory action, in animal models of severe acute pancreatitis. Eur. J. Pharmacol. 2012, 687, 28–38. [Google Scholar] [CrossRef]
- Ohashi, S.; Nishio, A.; Nakamura, H.; Kido, M.; Ueno, S.; Uza, N.; Inoue, S.; Kitamura, H.; Kiriya, K.; Asada, M.; et al. Protective roles of redox-active protein thioredoxin-1 for severe acute pancreatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G772–G781. [Google Scholar] [CrossRef] [Green Version]
- Pervaiz, S.; Bellot, G.L.; Lemoine, A.; Brenner, C. Redox signaling in the pathogenesis of human disease and the regulatory role of autophagy. Int. Rev. Cell Mol. Biol. 2020, 352, 189–214. [Google Scholar] [CrossRef]
- Zuo, J.; Zhang, Z.; Luo, M.; Zhou, L.; Nice, E.C.; Zhang, W.; Wang, C.; Huang, C. Redox signaling at the crossroads of human health and disease. MedComm 2022, 3, e127. [Google Scholar] [CrossRef]
- Kino, K.; Hirao-Suzuki, M.; Morikawa, M.; Sakaga, A.; Miyazawa, H. Generation, repair and replication of guanine oxidation products. Genes Environ. 2017, 39, 21. [Google Scholar] [CrossRef] [PubMed]
- Steenken, S.; Jovanovic, S.V. How easily oxidizable is DNA? One-electron reduction potentials of adenosine and guanosine radicals in aqueous solution. J. Am. Chem. Soc. 1997, 119, 617–648. [Google Scholar] [CrossRef]
- Ahmad, A.; Haas De Mello, A.; Szczesny, B.; Toro, G.; Marcatti, M.; Druzhyna, N.; Liaudet, L.; Tarantini, S.; Salomao, R.; Garcia Soriano, F.; et al. Effects of the Poly(ADP-Ribose) Polymerase Inhibitor Olaparib in Cerulein-Induced Pancreatitis. Shock 2020, 53, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Virag, L. Structure and function of poly(ADP-ribose) polymerase-1: Role in oxidative stress-related pathologies. Curr. Vasc. Pharmacol. 2005, 3, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Yuan, K.; Yan, C.; Fox, J., 3rd; Gaid, M.; Breitwieser, W.; Bansal, A.K.; Zeng, H.; Gao, H.; Wu, M. 8-Oxoguanine-DNA glycosylase 1 deficiency modifies allergic airway inflammation by regulating STAT6 and IL-4 in cells and in mice. Free Radic. Biol. Med. 2012, 52, 392–401. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Lin, P.; Wu, Q.; Pu, Q.; Zhou, C.; Wang, B.; Gao, P.; Wang, Z.; Gao, A.; Overby, M.; et al. Small-Molecule Inhibitor of 8-Oxoguanine DNA Glycosylase 1 Regulates Inflammatory Responses during Pseudomonas aeruginosa Infection. J. Immunol. 2020, 205, 2231–2242. [Google Scholar] [CrossRef]
- Rognlien, A.G.W.; Wollen, E.J.; Atneosen-Asegg, M.; Suganthan, R.; Bjoras, M.; Saugstad, O.D. Neonatal Ogg1/Mutyh knockout mice have altered inflammatory gene response compared to wildtype mice in the brain and lung after hypoxia-reoxygenation. J. Perinat. Med. 2018, 47, 114–124. [Google Scholar] [CrossRef] [Green Version]
- Shibutani, S.; Takeshita, M.; Grollman, A.P. Insertion of specific bases during DNA synthesis past the oxidation-damaged base 8-oxodG. Nature 1991, 349, 431–434. [Google Scholar] [CrossRef]
- Larsen, E.; Reite, K.; Nesse, G.; Gran, C.; Seeberg, E.; Klungland, A. Repair and mutagenesis at oxidized DNA lesions in the developing brain of wild-type and Ogg1-/- mice. Oncogene 2006, 25, 2425–2432. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primers (5→3) | Reverse Primers (5→3) |
|---|---|---|
| CCL2 | CACTCACCTGCTGCTACTCA | GCTTGGTGACAAAAACTACAGC |
| CCL4 | CCCACTTCCTGCTGTTTCTCT | GCCTCTTTTGGTCAGGAATACCA |
| CCL12 | AAGCAGAAGATTCACGTCCG | ATCCAGTATGGTCCTGAAGATCAC |
| IL23 | CCAGCGGGACATATGAATCTAC | TGTCCTTGAGTCCTTGTGGG |
| IL16 | AACTCCTCTACTGACTCCGC | ACCCTGTTCTGTCCCTTTGA |
| IL10 | GCTGTCATCGATTTCTCCCCT | GACACCTTGGTCTTGGAGCTTAT |
| CSF1 | GCCCTTCTTCGACATGGCT | CCTTCAGGTGTCCATTCCCA |
| TREM1 | TGCCACTTCATACACTGGGT | ATTGTGGAGAAGTCGAGGCA |
| IL1-ra | CATTGCTGGGTACTTACAAGGA | AGTGATGTTAACTTCCTCCAGC |
| IL6 | TTCTTGGGACTGATGCTGGT | CCATTGCACAACTCTTTTCTCA |
| IL1β | TGCCACCTTTTGACAGTGATG | ATGTGCTGCTGCGAGATTTG |
| TNFα | GGATGAGAAGTTCCCAAATGGC | TGTCTTTGAGATCCATGCCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajnády, Z.; Nagy-Pénzes, M.; Demény, M.A.; Kovács, K.; El-Hamoly, T.; Maléth, J.; Hegyi, P.; Polgár, Z.; Hegedűs, C.; Virág, L. OGG1 Inhibition Reduces Acinar Cell Injury in a Mouse Model of Acute Pancreatitis. Biomedicines 2022, 10, 2543. https://doi.org/10.3390/biomedicines10102543
Hajnády Z, Nagy-Pénzes M, Demény MA, Kovács K, El-Hamoly T, Maléth J, Hegyi P, Polgár Z, Hegedűs C, Virág L. OGG1 Inhibition Reduces Acinar Cell Injury in a Mouse Model of Acute Pancreatitis. Biomedicines. 2022; 10(10):2543. https://doi.org/10.3390/biomedicines10102543
Chicago/Turabian StyleHajnády, Zoltán, Máté Nagy-Pénzes, Máté A. Demény, Katalin Kovács, Tarek El-Hamoly, József Maléth, Péter Hegyi, Zsuzsanna Polgár, Csaba Hegedűs, and László Virág. 2022. "OGG1 Inhibition Reduces Acinar Cell Injury in a Mouse Model of Acute Pancreatitis" Biomedicines 10, no. 10: 2543. https://doi.org/10.3390/biomedicines10102543