
Prevention of Anti-HMGCR Immune-Mediated Necrotising Myopathy by C5 Complement Inhibition in a Humanised Mouse Model

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Samples
2.2. Mice
2.3. IgG Purification
2.4. Passive Transfer of Human IgG into Mice
2.5. Human Complement Supplementation
2.6. In Vitro Classical Pathway Complement-Mediated Hemolysis Assay and ELISA for Complement Components
2.7. Zilucoplan Administration
2.8. Mouse Muscle Strength Evaluation
2.9. Histological Analysis
2.10. Statistical Analysis
3. Results
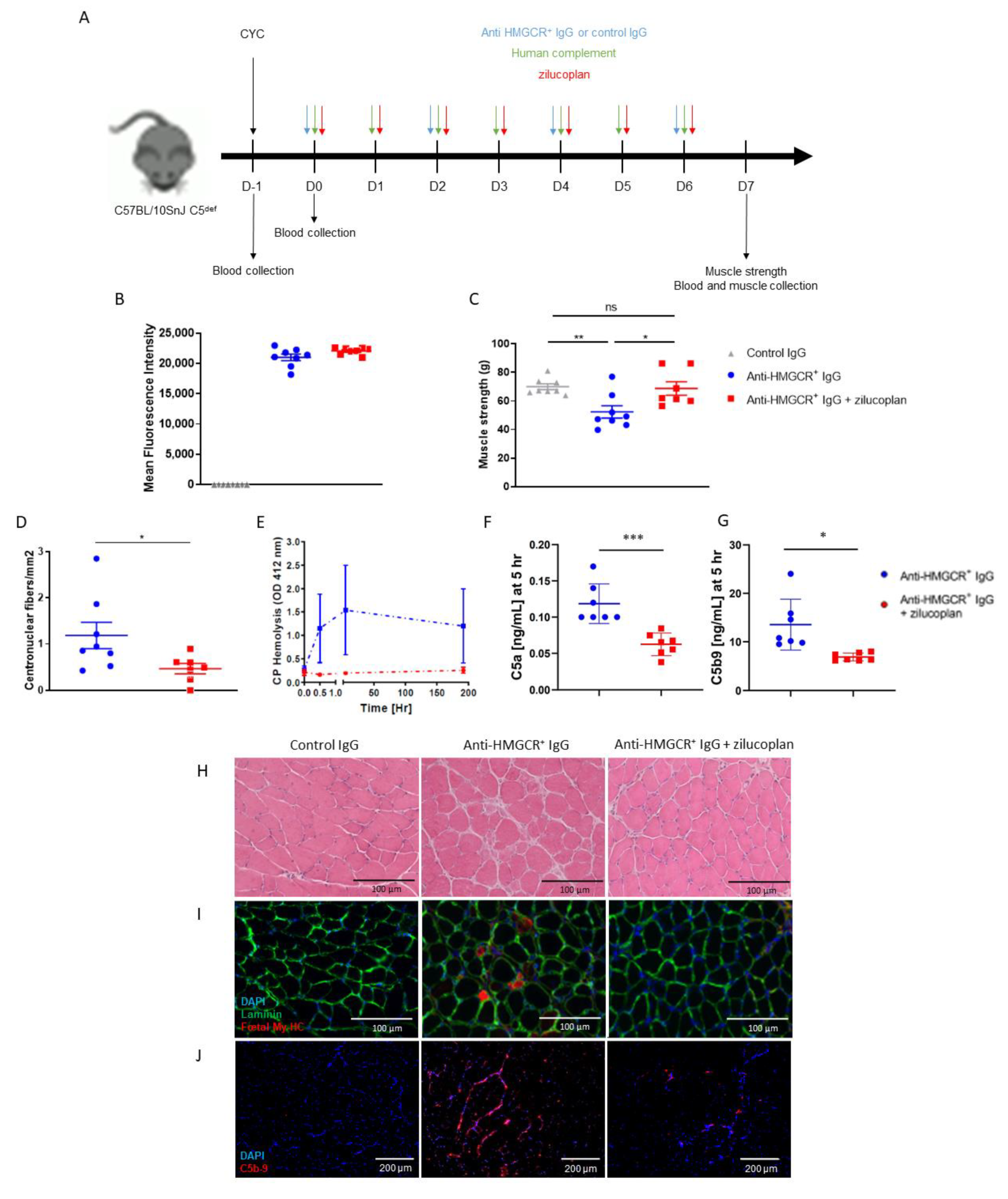
3.1. Zilucoplan Prevents Onset of the Disease in C5-Deficient Mice Supplemented with Human Complement
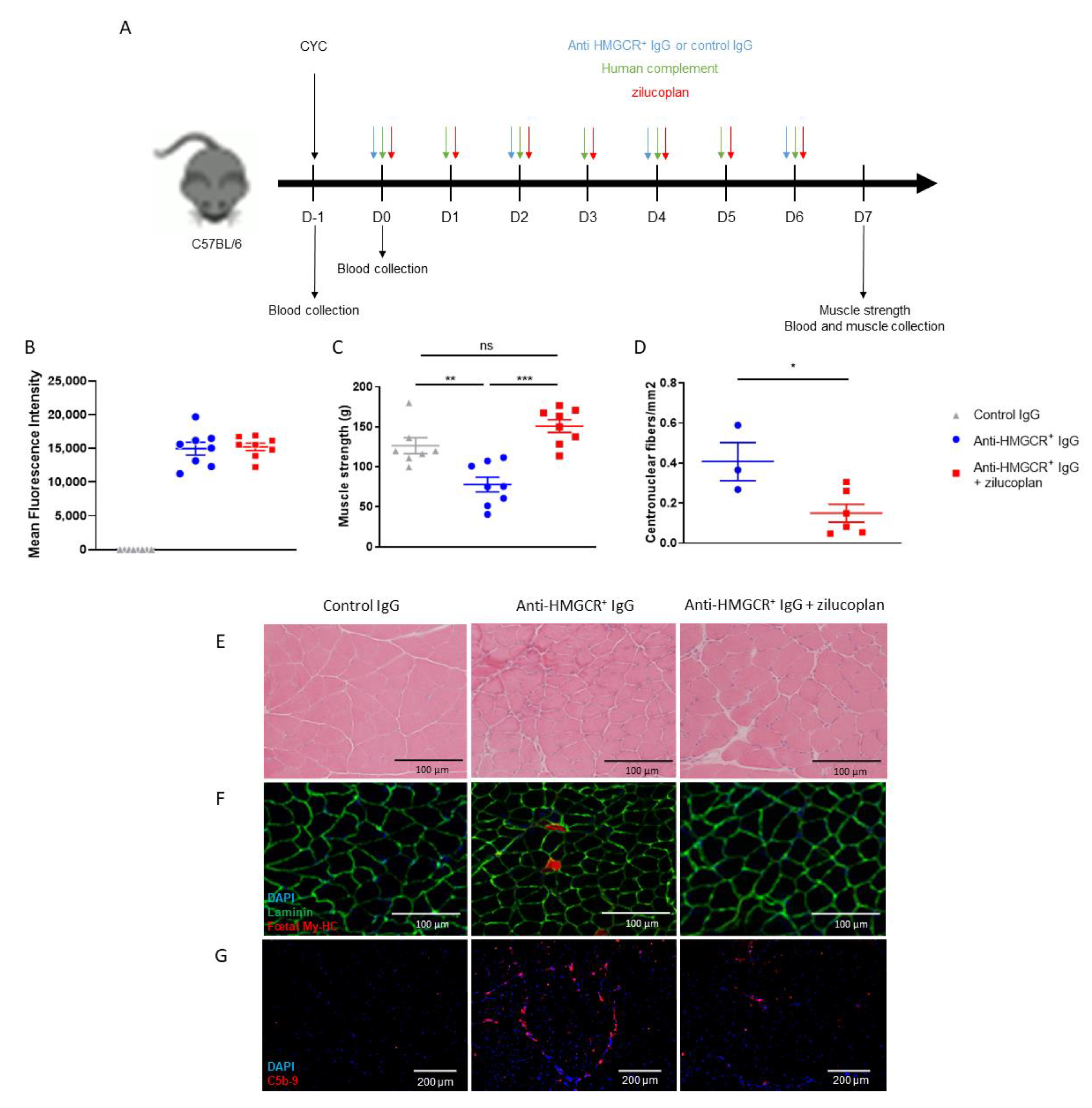
3.2. C57BL/6 Mice Prophylactically Treated with Zilucoplan Are Protected from Myopathy
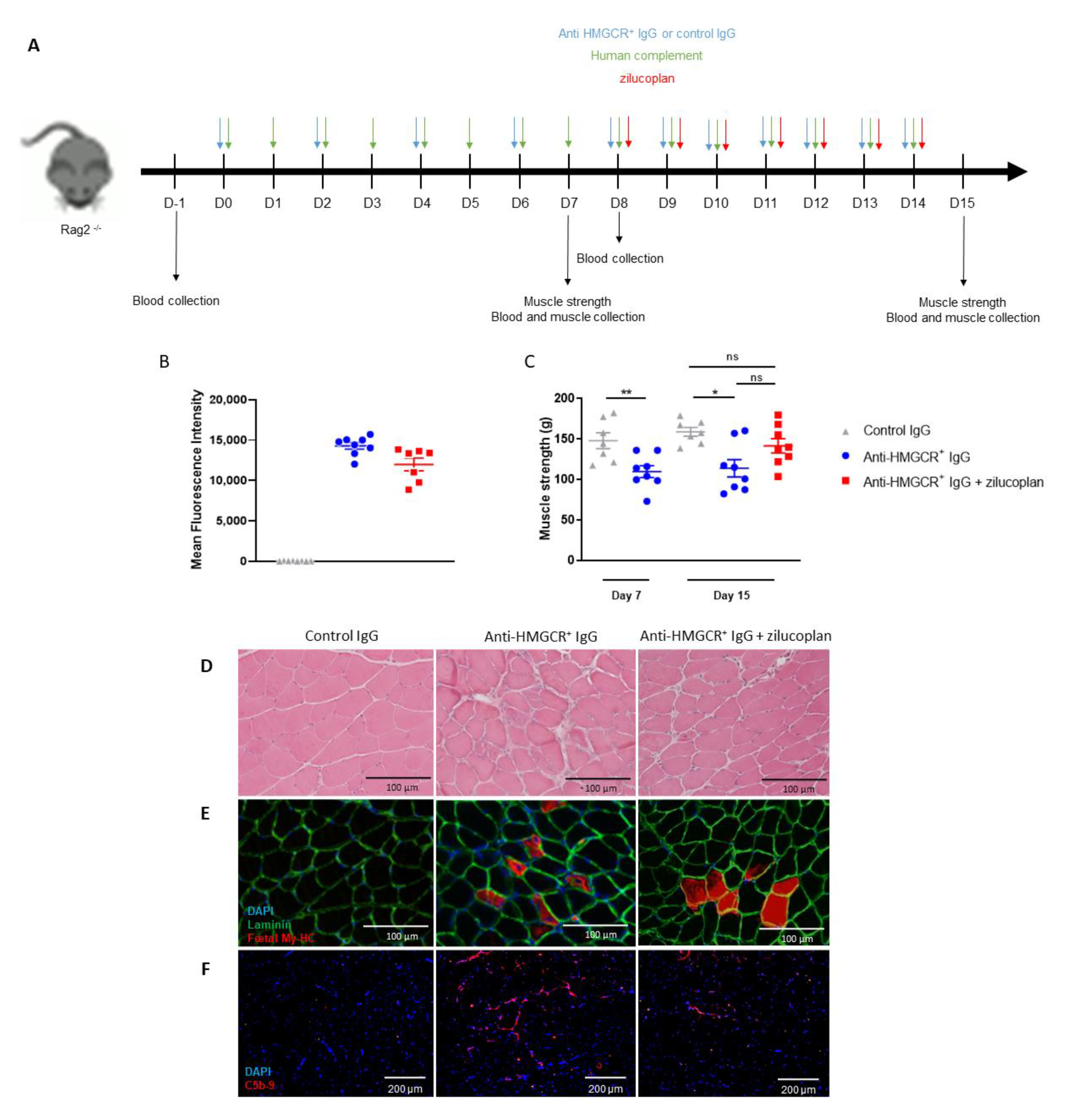
3.3. Intervention with Zilucoplan Does Not Fully Normalize Muscle Strength Loss in Animals with Established IMNM Disease
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allenbach, Y.; Benveniste, O.; Stenzel, W.; Boyer, O. Immune-Mediated Necrotizing Myopathy: Clinical Features and Pathogenesis. Nat. Rev. Rheumatol. 2020, 16, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Christopher-Stine, L.; Casciola-Rosen, L.A.; Hong, G.; Chung, T.; Corse, A.M.; Mammen, A.L. A Novel Autoantibody Recognizing 200-Kd and 100-Kd Proteins Is Associated with an Immune-Mediated Necrotizing Myopathy. Arthritis Rheum. 2010, 62, 2757–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, Y.; Uruha, A.; Suzuki, S.; Nakahara, J.; Hamanaka, K.; Takayama, K.; Suzuki, N.; Nishino, I. Clinical Features and Prognosis in Anti-SRP and Anti-HMGCR Necrotising Myopathy. J. Neurol. Neurosurg. Psychiatry 2016, 87, 1038–1044. [Google Scholar] [CrossRef] [PubMed]
- Pinal-Fernandez, I.; Parks, C.; Werner, J.L.; Albayda, J.; Paik, J.; Danoff, S.K.; Casciola-Rosen, L.; Christopher-Stine, L.; Mammen, A.L. Longitudinal Course of Disease in a Large Cohort of Myositis Patients With Autoantibodies Recognizing the Signal Recognition Particle. Arthritis Care Res. 2017, 69, 263–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, W.H.; Nigam, S.K.; Blobel, G. Human Autoantibodies Reactive with the Signal-Recognition Particle. Proc. Natl. Acad. Sci. USA 1986, 83, 9507–9511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mammen, A.L.; Chung, T.; Christopher-Stine, L.; Rosen, P.; Rosen, A.; Doering, K.R.; Casciola-Rosen, L.A. Autoantibodies against 3-Hydroxy-3-Methylglutaryl-Coenzyme A Reductase in Patients with Statin-Associated Autoimmune Myopathy. Arthritis Rheum. 2011, 63, 713–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benveniste, O.; Drouot, L.; Jouen, F.; Charuel, J.-L.; Bloch-Queyrat, C.; Behin, A.; Amoura, Z.; Marie, I.; Guiguet, M.; Eymard, B.; et al. Correlation of Anti-Signal Recognition Particle Autoantibody Levels with Creatine Kinase Activity in Patients with Necrotizing Myopathy. Arthritis Rheum. 2011, 63, 1961–1971. [Google Scholar] [CrossRef] [PubMed]
- Werner, J.L.; Christopher-Stine, L.; Ghazarian, S.R.; Pak, K.S.; Kus, J.E.; Daya, N.R.; Lloyd, T.E.; Mammen, A.L. Antibody Levels Correlate with Creatine Kinase Levels and Strength in Anti-3-Hydroxy-3-Methylglutaryl-Coenzyme A Reductase-Associated Autoimmune Myopathy. Arthritis Rheum. 2012, 64, 4087–4093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allenbach, Y.; Arouche-Delaperche, L.; Preusse, C.; Radbruch, H.; Butler-Browne, G.; Champtiaux, N.; Mariampillai, K.; Rigolet, A.; Hufnagl, P.; Zerbe, N.; et al. Necrosis in Anti-SRP+ and Anti-HMGCR+myopathies: Role of Autoantibodies and Complement. Neurology 2018, 90, e507–e517. [Google Scholar] [CrossRef] [PubMed]
- Arouche-Delaperche, L.; Allenbach, Y.; Amelin, D.; Preusse, C.; Mouly, V.; Mauhin, W.; Tchoupou, G.D.; Drouot, L.; Boyer, O.; Stenzel, W.; et al. Pathogenic Role of Anti-Signal Recognition Protein and Anti-3-Hydroxy-3-Methylglutaryl-CoA Reductase Antibodies in Necrotizing Myopathies: Myofiber Atrophy and Impairment of Muscle Regeneration in Necrotizing Autoimmune Myopathies. Ann. Neurol. 2017, 81, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Bergua, C.; Chiavelli, H.; Allenbach, Y.; Arouche-Delaperche, L.; Arnoult, C.; Bourdenet, G.; Jean, L.; Zoubairi, R.; Guerout, N.; Mahler, M.; et al. In Vivo Pathogenicity of IgG from Patients with Anti-SRP or Anti-HMGCR Autoantibodies in Immune-Mediated Necrotising Myopathy. Ann. Rheum. Dis. 2019, 78, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Allenbach, Y.; Mammen, A.L.; Benveniste, O.; Stenzel, W.; Immune-Mediated Necrotizing Myopathies Working Group. 224th ENMC International Workshop: Clinico-Sero-Pathological Classification of Immune-Mediated Necrotizing Myopathies Zandvoort, The Netherlands, 14–16 October 2016. Neuromuscul. Disord. NMD 2018, 28, 87–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamirou, F.; Arnaud, L.; Talarico, R.; Scirè, C.A.; Alexander, T.; Amoura, Z.; Avcin, T.; Bortoluzzi, A.; Cervera, R.; Conti, F.; et al. Systemic Lupus Erythematosus: State of the Art on Clinical Practice Guidelines. RMD Open 2018, 4, e000793. [Google Scholar] [CrossRef] [PubMed]
- Pickering, M.C.; Ismajli, M.; Condon, M.B.; McKenna, N.; Hall, A.E.; Lightstone, L.; Terence Cook, H.; Cairns, T.D. Eculizumab as Rescue Therapy in Severe Resistant Lupus Nephritis. Rheumatol. Oxf. Engl. 2015, 54, 2286–2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pittock, S.J.; Berthele, A.; Fujihara, K.; Kim, H.J.; Levy, M.; Palace, J.; Nakashima, I.; Terzi, M.; Totolyan, N.; Viswanathan, S.; et al. Eculizumab in Aquaporin-4-Positive Neuromyelitis Optica Spectrum Disorder. N. Engl. J. Med. 2019, 381, 614–625. [Google Scholar] [CrossRef] [PubMed]
- Hoogendijk, J.E.; Amato, A.A.; Lecky, B.R.; Choy, E.H.; Lundberg, I.E.; Rose, M.R.; Vencovsky, J.; de Visser, M.; Hughes, R.A. 119th ENMC International Workshop: Trial Design in Adult Idiopathic Inflammatory Myopathies, with the Exception of Inclusion Body Myositis, 10–12 October 2003, Naarden, The Netherlands. Neuromuscul. Disord. NMD 2004, 14, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Drouot, L.; Allenbach, Y.; Jouen, F.; Charuel, J.-L.; Martinet, J.; Meyer, A.; Hinschberger, O.; Bader-Meunier, B.; Kone-Paut, I.; Campana-Salort, E.; et al. Exploring Necrotizing Autoimmune Myopathies with a Novel Immunoassay for Anti-3-Hydroxy-3-Methyl-Glutaryl-CoA Reductase Autoantibodies. Arthritis Res. Ther. 2014, 16, R39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, J.F.; Nowak, R.J.; Wolfe, G.I.; Freimer, M.L.; Vu, T.H.; Hinton, J.L.; Benatar, M.; Duda, P.W.; MacDougall, J.E.; Farzaneh-Far, R.; et al. Clinical Effects of the Self-Administered Subcutaneous Complement Inhibitor Zilucoplan in Patients With Moderate to Severe Generalized Myasthenia Gravis: Results of a Phase 2 Randomized, Double-Blind, Placebo-Controlled, Multicenter Clinical Trial. JAMA Neurol. 2020, 77, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Rødgaard, A.; Nielsen, F.C.; Djurup, R.; Somnier, F.; Gammeltoft, S. Acetylcholine Receptor Antibody in Myasthenia Gravis: Predominance of IgG Subclasses 1 and 3. Clin. Exp. Immunol. 1987, 67, 82–88. [Google Scholar] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Julien, S.; Vadysirisack, D.; Sayegh, C.; Ragunathan, S.; Tang, Y.; Briand, E.; Carrette, M.; Jean, L.; Zoubairi, R.; Gondé, H.; et al. Prevention of Anti-HMGCR Immune-Mediated Necrotising Myopathy by C5 Complement Inhibition in a Humanised Mouse Model. Biomedicines 2022, 10, 2036. https://doi.org/10.3390/biomedicines10082036
Julien S, Vadysirisack D, Sayegh C, Ragunathan S, Tang Y, Briand E, Carrette M, Jean L, Zoubairi R, Gondé H, et al. Prevention of Anti-HMGCR Immune-Mediated Necrotising Myopathy by C5 Complement Inhibition in a Humanised Mouse Model. Biomedicines. 2022; 10(8):2036. https://doi.org/10.3390/biomedicines10082036
Chicago/Turabian StyleJulien, Sarah, Douangsone Vadysirisack, Camil Sayegh, Sharan Ragunathan, Yalan Tang, Emma Briand, Marion Carrette, Laetitia Jean, Rachid Zoubairi, Henri Gondé, and et al. 2022. "Prevention of Anti-HMGCR Immune-Mediated Necrotising Myopathy by C5 Complement Inhibition in a Humanised Mouse Model" Biomedicines 10, no. 8: 2036. https://doi.org/10.3390/biomedicines10082036