
Comparative Proteomic Assessment of Normal vs. Polyhydramnios Amniotic Fluid Based on Computational Analysis

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Gel Electrophoresis
2.3. In-Gel Digestion and MALDI-TOF MS Analysis
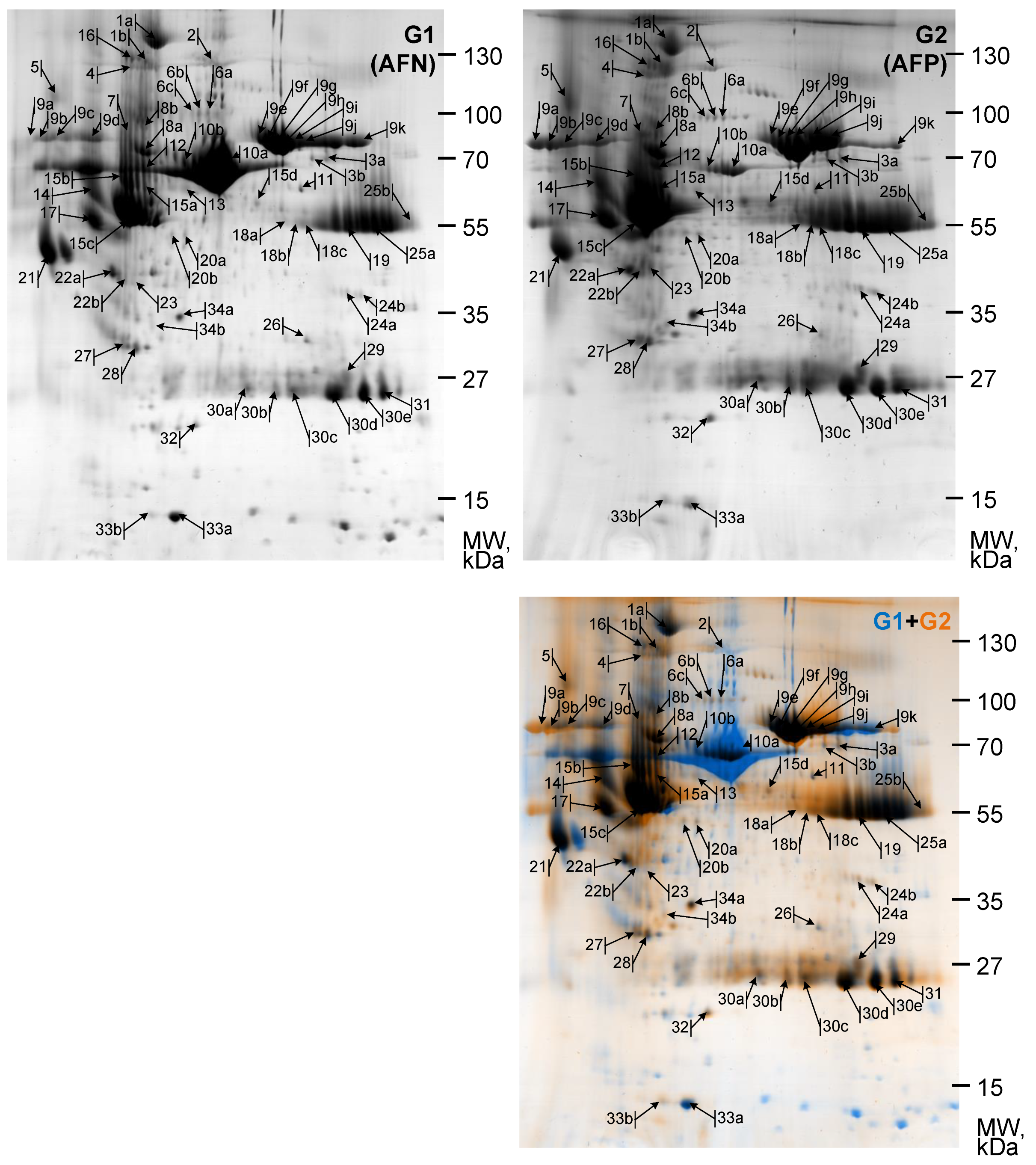
2.4. Gel Image Computational Analysis
- Image preparation tools: image cropping (to remove excess areas), spot labelling, master gel selection (to align the group of gels), molecular mass markers’ calibration (to delineate area of MM marker and input of MM values), pI calibration (to input positions of known pI values);
- Image preprocessing related tools: image smoothing (to eliminate impulse noise), background elimination (to remove variations of background staining), individual image warping (to straighten protein migration paths);
- Image segmentation tools: 2DE image splitting (to split image into primary segments), segmented area evaluation (to highlight uncertain segmentations of protein spots for the user), editing of segments (to manually edit protein spots in order to remove false negatives and positives of segmentation by merging, splitting, adding, or removing areas);
- Image alignment tools: initial registration (to automatically detect some high confident control points for initial image registration), spot pairing and concluding image alignment (to find correspondences between spots), manual editing of alignment vectors (to remove mismatches and add new matches between images);
- Quantitative analysis tools: spot quantification (to measure normalized quantities of spots), changes evaluation (to calculate change ratios);
- Visualization tools: 3D viewer (to display small area of image as surface), image fusion (to display overlay of two images using pseudocolors).
2.5. Image Preprocessing
2.6. Gel Image Alignment
Protein Spot Detection and Segmentation
- 1.
- Mode of each region of the 2DE image;
- 2.
- Weighted difference of maximum and minimum value in contour pixels of each region of the 2DE image;
- 3.
- Weighted difference of maximum and minimum value in contour pixels of each region of the symmetry map;
- 4.
- Maximum value of Kovesi phase symmetry feature detector [22] in each region of the 2DE image;
- 5.
- Maximum Laplacian value of Gaussian in each region of the 2DE image.
3. Results
3.1. Image Preparation and Preprocessing
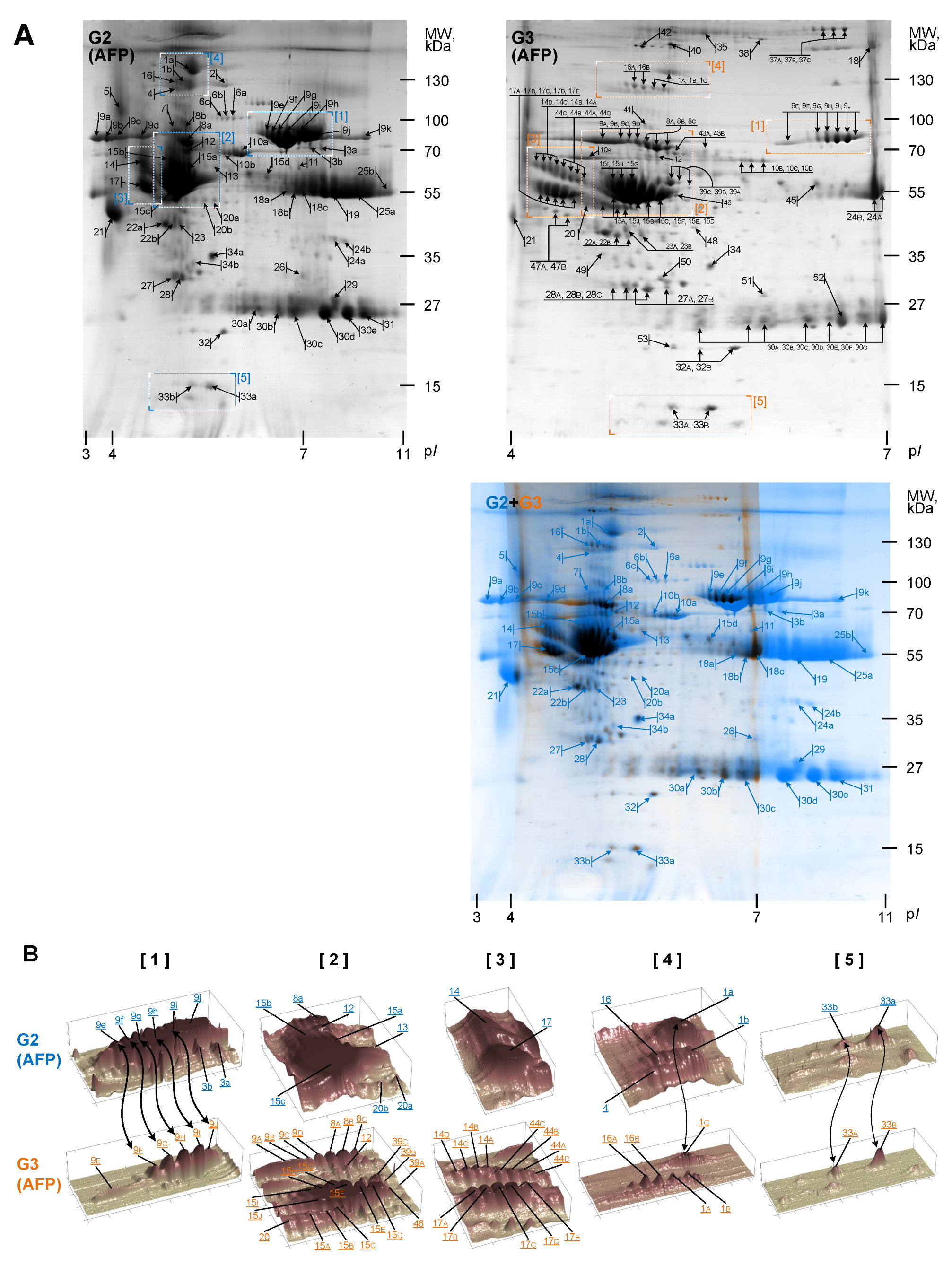
3.2. Gel Image Alignment and Segmentation
3.3. Quantitative Analysis
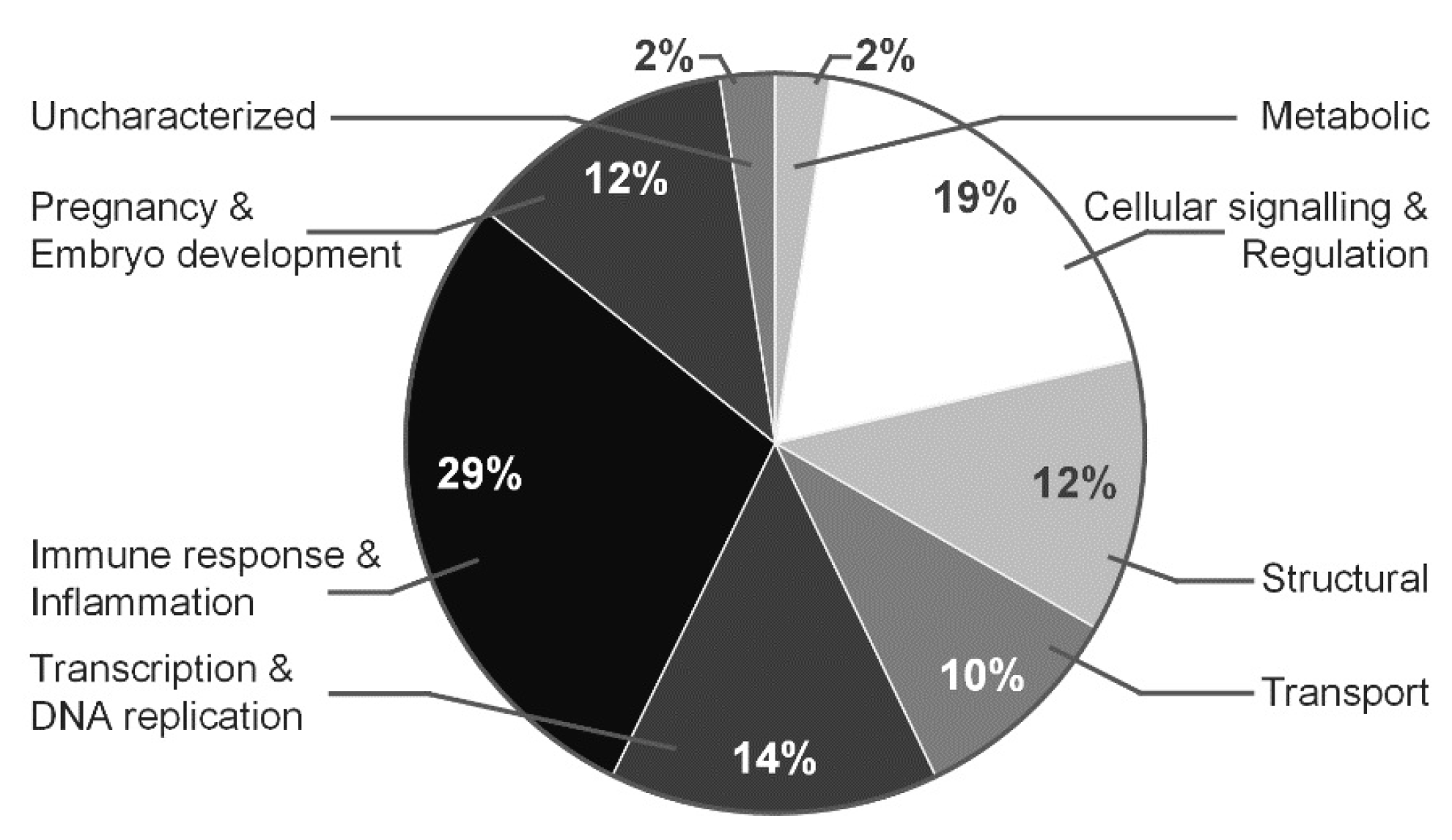
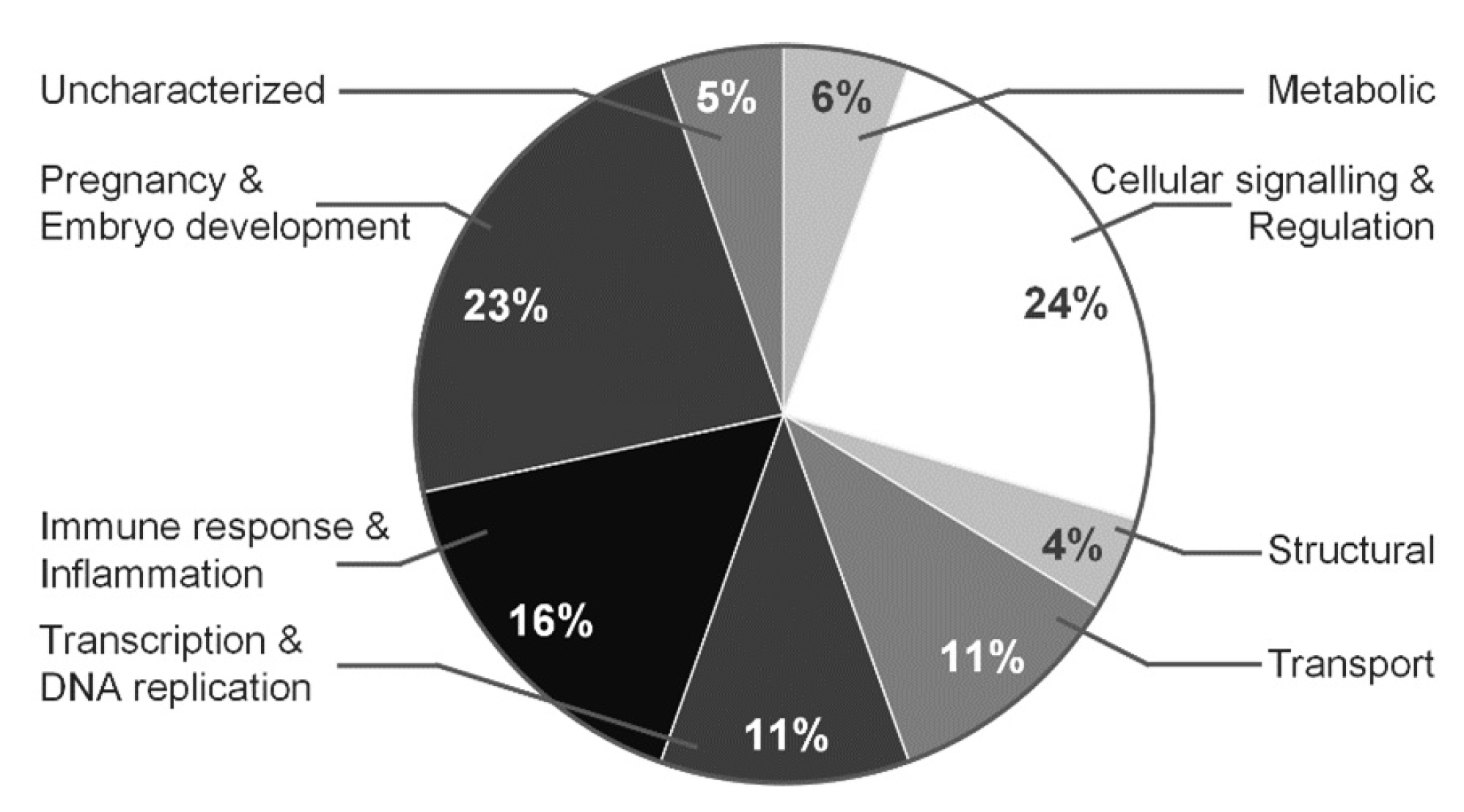
3.4. Proteomic Profile Characteristics for Normal and Polyhydramnios Amniotic Fluid
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clapp, J.F., 3rd; Schmidt, S.; Paranjape, A.; Lopez, B. Maternal insulin-like growth factor-I levels (IGF-I) reflect placental mass and neonatal fat mass. Am. J. Obstet. Gynecol. 2004, 190, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Angel, T.E.; Aryal, U.K.; Hengel, S.M.; Baker, E.S.; Kelly, R.T.; Robinson, E.W.; Smith, R.D. Mass spectrometry-based proteomics: Existing capabilities and future directions. Chem. Soc. Rev. 2012, 41, 3912–3928. [Google Scholar] [CrossRef] [PubMed]
- Kurdi, W. Non-immune fetal hydrops: Are we doing the appropriate tests each time? J. Prenat. Med. 2007, 1, 26. [Google Scholar] [PubMed]
- Henrich, W.; Heeger, J.; Schmider, A.; Dudenhausen, J. Complete spontaneous resolution of severe nonimmunological hydrops fetalis with unknown etiology in the second trimester—A case report. J. Perinat. Med. 2002, 30, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, N.; Ryan, G. Fetal primary pleural effusions: Prenatal diagnosis and management. Best Pract. Res. Clin. Obstet. Gynaecol. 2019, 58, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Bellini, C.; Hennekam, R.C.; Fulcheri, E.; Rutigliani, M.; Morcaldi, G.; Boccardo, F.; Bonioli, E. Etiology of nonimmune hydrops fetalis: A systematic review. Am. J. Med. Genet. Part A 2009, 149, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Kolialexi, A.; Mavrou, A.; Spyrou, G.; Tsangaris, G.T. Mass spectrometry-based proteomics in reproductive medicine. Mass Spectrom. Rev. 2008, 27, 624–634. [Google Scholar] [CrossRef]
- Okeke, T.; Ezenyeaku, C.; Ikeako, L. Recent Perspective of Amniotic Fluid Embolism: A Review Article. Br. J. Med. Med. Res. 2015, 8, 408–418. [Google Scholar] [CrossRef]
- Cho, C.K.J.; Shan, S.J.; Winsor, E.J.; Diamandis, E.P. Proteomics analysis of human amniotic fluid. Mol. Cell. Proteom. 2007, 6, 1406–1415. [Google Scholar] [CrossRef]
- Huang, W.; Gu, H.; Yuan, Z. Identifying biomarkers for prenatal diagnosis of neural tube defects based on “omics”. Clin. Genet. 2022, 101, 381–389. [Google Scholar] [CrossRef]
- Navakauskiene, R.; Baronaite, S.; Matuzevicius, D.; Zaikova, I.; Arlauskiene, A.; Navakauskas, D.; Treigyte, G. Identification and Characterization of Amniotic Fluid Proteins Incident to Normal, Preeclampsia and Polyhydramnios Pregnancies. Curr. Proteom. 2016, 13, 206–217. [Google Scholar] [CrossRef]
- Matuzevičius, D.; Žurauskas, E.; Navakauskienė, R.; Navakauskas, D. Improved proteomic characterization of human myocardium and heart conduction system by computational methods. Biologija 2008, 54, 283–289. [Google Scholar] [CrossRef]
- Navakauskienė, R.; Navakauskas, D.; Borutinskaitė, V.; Matuzevičius, D. Epigenetics and Proteomics of Leukemia—A Synergy of Experimental Biology and Computational Informatics; Springer: Cham, Switzerland, 2021; p. 406. [Google Scholar]
- Matuzevičius, D. Analysis of 2D Electrophoresis Gel Images Using Intelligent Techniques. Ph.D. Thesis, Vilnius Gediminas Technical University, Vilnius, Lithuania, 2010. [Google Scholar]
- Mateika, D.; Martavičius, R. Large image formation using Harris-Plessey corner detection algorithm. Elektron. Elektrotech. 2008, 5, 21–24. [Google Scholar]
- Bookstein, F.L. Principal warps: Thin-plate splines and the decomposition of deformations. IEEE Trans. Pattern Anal. Mach. Intell. 1989, 11, 567–585. [Google Scholar] [CrossRef]
- Lowe, D.G. Distinctive image features from scale-invariant keypoints. Int. J. Comput. Vis. 2004, 60, 91–110. [Google Scholar] [CrossRef]
- Johansson, B. Low Level Operations and Learning in Computer Vision. Ph.D. Thesis, Linköping University Electronic Press, Linköping, Sweden, 2004. [Google Scholar]
- Vincent, L.; Soille, P. Watersheds in digital spaces: An efficient algorithm based on immersion simulations. IEEE Trans. Pattern Anal. Mach. Intell. 1991, 13, 583–598. [Google Scholar] [CrossRef]
- Dowsey, A.W.; Dunn, M.J.; Yang, G.Z. The role of bioinformatics in two-dimensional gel electrophoresis. Proteom. Int. Ed. 2003, 3, 1567–1596. [Google Scholar] [CrossRef]
- Matuzevičius, D.; Serackis, A.; Navakauskas, D. Mathematical models of oversaturated protein spots. Elektron. Elektrotech. 2007, 73, 63–68. [Google Scholar]
- Kovesi, P. Image features from phase congruency. Videre J. Comput. Vis. Res. 1999, 1, 1–26. [Google Scholar]
- Yetgin, S.; Aytac, S.; Gurakan, F.; Yurdakok, M. Nonimmune hydrops fetalis in two cases of consanguineous parents and associated with hereditary spherocytosis and hemophagocytic hystiocytosis. J. Perinatol. 2007, 27, 252–254. [Google Scholar] [CrossRef]
- Yurdakök, M. Non-immune hydrops fetalis. J. Pediatr. Neonatal Individ. Med. (JPNIM) 2014, 3, e030214. [Google Scholar]
- Bellini, C.; Donarini, G.; Paladini, D.; Calevo, M.G.; Bellini, T.; Ramenghi, L.A.; Hennekam, R.C. Etiology of non-immune hydrops fetalis: An update. Am. J. Med. Genet. Part A 2015, 167, 1082–1088. [Google Scholar] [CrossRef]
- Chaiworapongsa, T.; Espinoza, J.; Yoshimatsu, J.; Kim, Y.; Bujold, E.; Edwin, S.; Yoon, B.; Romero, R. Activation of coagulation system in preterm labor and preterm premature rupture of membranes. J. Matern. -Fetal Neonatal Med. 2002, 11, 368–373. [Google Scholar] [CrossRef]
- Elovitz, M.A.; Baron, J.; Phillippe, M. The role of thrombin in preterm parturition. Am. J. Obstet. Gynecol. 2001, 185, 1059–1063. [Google Scholar] [CrossRef]
- Padda, J.; Khalid, K.; Mohan, A.; Pokhriyal, S.; Batra, N.; Hitawala, G.; Cooper, A.; Jean-Charles, G. Factor V Leiden G1691A and Prothrombin Gene G20210A Mutations on Pregnancy Outcome. Cureus 2021, 13, e17185. [Google Scholar] [CrossRef]
- Schjetlein, R.; Abdelnoor, M.; Haugen, G.; Husby, H.; Sandset, P.; Wisløff, F. Hemostatic variables as independent predictors for fetal growth retardation in preeclampsia. Acta Obstet. Gynecol. Scand. 1999, 78, 191–197. [Google Scholar]
- Hayashi, M.; Ohkura, T. Elevated levels of serum macrophage colony-stimulating factor in normotensive pregnancies complicated by intrauterine fetal growth restriction. Exp. Hematol. 2002, 30, 388–393. [Google Scholar] [CrossRef]
- Erez, O.; Othman, M.; Rabinovich, A.; Leron, E.; Gotsch, F.; Thachil, J. DIC in Pregnancy–Pathophysiology, Clinical Characteristics, Diagnostic Scores, and Treatments. J. Blood Med. 2022, 13, 21–44. [Google Scholar] [CrossRef]
- Mastrolia, S.A.; Mazor, M.; Loverro, G.; Klaitman, V.; Erez, O. Placental vascular pathology and increased thrombin generation as mechanisms of disease in obstetrical syndromes. PeerJ 2014, 2, e653. [Google Scholar] [CrossRef]
- Karamessinis, P.M.; Malamitsi-Puchner, A.; Boutsikou, T.; Makridakis, M.; Vougas, K.; Fountoulakis, M.; Vlahou, A.; Chrousos, G. Marked defects in the expression and glycosylation of α2-HS glycoprotein/fetuin-A in plasma from neonates with intrauterine growth restriction: Proteomics screening and potential clinical implications. Mol. Cell. Proteom. 2008, 7, 591–599. [Google Scholar] [CrossRef]
- Hutchison, A.A.; Drew, J.H.; Yu, V.; Williams, M.L.; Fortune, D.W.; Beischer, N.A. Nonimmunologic hydrops fetalis: A review of 61 cases. Obstet. Gynecol. 1982, 59, 347–352. [Google Scholar] [PubMed]
- Krishnaswamy, S. Exosite-driven substrate specificity and function in coagulation. J. Thromb. Haemost. 2005, 3, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Erez, O.; Romer, R.; Vaisbuch, E.; Chaiworapongsa, T.; Kusanovic, J.P.; Mazaki-Tovi, S.; Gotsch, F.; Gomez, R.; Maymon, E.; Pacora, P.; et al. Changes in amniotic fluid concentration of thrombin–antithrombin III complexes in patients with preterm labor: Evidence of an increased thrombin generation. J. Matern. -Fetal Neonatal Med. 2009, 22, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Staretz-Chacham, O.; Lang, T.C.; LaMarca, M.E.; Krasnewich, D.; Sidransky, E. Lysosomal storage disorders in the newborn. Pediatrics 2009, 123, 1191–1207. [Google Scholar] [CrossRef]
- Anderson, S. Congenital Nephrotic Syndrome of the Finnish Type in a Dominican Newborn: An Overview and Case Report. Neonatal Netw. 2022, 41, 83–88. [Google Scholar] [CrossRef]
- Nicolaides, K.; Warenski, J.; Rodeck, C. The relationship of fetal plasma protein concentration and hemoglobin level to the development of hydrops in rhesus isoimmunization. Am. J. Obstet. Gynecol. 1985, 152, 341–344. [Google Scholar] [CrossRef]
- Jones, S.E.; Jomary, C. Clusterin. Int. J. Biochem. Cell Biol. 2002, 34, 427–431. [Google Scholar] [CrossRef]
- Blumenstein, M.; McCowan, L.M.; Wu, S.; Cooper, G.J.; North, R.A. Plasma clusterin increased prior to small for gestational age (SGA) associated with preeclampsia and decreased prior to SGA in normotensive pregnancies. Reprod. Sci. 2012, 19, 650–657. [Google Scholar] [CrossRef]
- Zeng, S.; Pan, Y.; Liu, F.; Yin, J.; Jiang, M.; Long, Y.; Zhao, X.; Lash, G.E.; Yang, H. Role of clusterin in the regulation of trophoblast development and preeclampsia. Biochem. Biophys. Res. Commun. 2021, 583, 128–134. [Google Scholar] [CrossRef]
- Han, B.H.; DeMattos, R.B.; Dugan, L.L.; Kim-Han, J.S.; Brendza, R.P.; Fryer, J.D.; Kierson, M.; Cirrito, J.; Quick, K.; Harmony, J.A.; et al. Clusterin contributes to caspase-3–independent brain injury following neonatal hypoxia-ischemia. Nat. Med. 2001, 7, 338–343. [Google Scholar] [CrossRef]
- Choolani, M.; Narasimhan, K.; Kolla, V.; Hahn, S. Proteomic technologies for prenatal diagnostics: Advances and challenges ahead. Expert Rev. Proteom. 2009, 6, 87–101. [Google Scholar] [CrossRef]
- Mak, C.M.; Lam, C.W.; Tam, S. Diagnostic accuracy of serum ceruloplasmin in Wilson disease: Determination of sensitivity and specificity by ROC curve analysis among ATP7B-genotyped subjects. Clin. Chem. 2008, 54, 1356–1362. [Google Scholar] [CrossRef]
- Sanchez-Monteagudo, A.; Ripolles, E.; Berenguer, M.; Espinos, C. Wilson’s Disease: Facing the Challenge of Diagnosing a Rare Disease. Biomedicines 2021, 9, 1100. [Google Scholar] [CrossRef]
- Chan, W.Y.; Richichi, J.; Griesmann, G.E.; Cushing, W.; Kling, O.R.; Rennert, O.M. Copper and ceruloplasmin activity in human amniotic fluid. Am. J. Obstet. Gynecol. 1980, 138, 257–259. [Google Scholar] [CrossRef]
- Bellos, I.; Papantoniou, N.; Pergialiotis, V. Serum ceruloplasmin levels in preeclampsia: A meta-analysis. J. Matern. -Fetal Neonatal Med. 2018, 31, 2342–2348. [Google Scholar] [CrossRef]
- Anagnostopoulos, A.K.; Kolialexi, A.; Mavrou, A.; Vougas, K.; Papantoniou, N.; Antsaklis, A.; Kanavakis, E.; Fountoulakis, M.; Tsangaris, G.T. Proteomic analysis of amniotic fluid in pregnancies with Klinefelter syndrome foetuses. J. Proteom. 2010, 73, 943–950. [Google Scholar] [CrossRef]
- Knuppel, R.A.; Hassan, M.I.; McDermott, J.J.; Tucker, M.; Morrison, J.C. Oxidative stress and antioxidants: Preterm birth and preterm infants. In Preterm Birth—Mother and Child; InTech: Rijeka, Croatia, 2012; pp. 125–150. [Google Scholar]
- Hitti, J.; Lapidus, J.A.; Lu, X.; Reddy, A.P.; Jacob, T.; Dasari, S.; Eschenbach, D.A.; Gravett, M.G.; Nagalla, S.R. Noninvasive diagnosis of intraamniotic infection: Proteomic biomarkers in vaginal fluid. Am. J. Obstet. Gynecol. 2010, 203, 32-e1. [Google Scholar] [CrossRef]
- Kalsheker, N.A. α1-antichymotrypsin. Int. J. Biochem. Cell Biol. 1996, 28, 961–964. [Google Scholar] [CrossRef]
- Legge, M. Amniotic fluid fibrinolytic system in fetal neural tube defects. Prenat. Diagn. 1983, 3, 145–149. [Google Scholar] [CrossRef]
- Iliodromiti, S.; Vrachnis, N.; Samoli, E.; Iliodromiti, Z.; Pangalos, C.; Drakoulis, N.; Creatsas, G.; Botsis, D. Fetuin A concentration in the second trimester amniotic fluid of fetuses with trisomy 21 appears to be lower: Phenotypic considerations. Mediat. Inflamm. 2012, 2012, 4. [Google Scholar] [CrossRef]
- Jenkins, N.T.; McKenzie, J.A.; Hagberg, J.M.; Witkowski, S. Plasma fetuin-A concentrations in young and older high-and low-active men. Metabolism 2011, 60, 265–271. [Google Scholar] [CrossRef]
- Chakravarti, S.; Magnuson, T.; Lass, J.H.; Jepsen, K.J.; LaMantia, C.; Carroll, H. Lumican regulates collagen fibril assembly: Skin fragility and corneal opacity in the absence of lumican. J. Cell Biol. 1998, 141, 1277–1286. [Google Scholar] [CrossRef]
- Mavrou, A.; Anagnostopoulos, A.K.; Kolialexi, A.; Vougas, K.; Papantoniou, N.; Antsaklis, A.; Fountoulakis, M.; Tsangaris, G.T. Proteomic analysis of amniotic fluid in pregnancies with Turner syndrome fetuses. J. Proteome Res. 2008, 7, 1862–1866. [Google Scholar] [CrossRef]
- Jirka, M.; Blanickỳ, P.; Šarajer, J.; Zwinger, A.; Jirasek, J. Human serum Zn-α2-glycoprotein in amniotic fluid. Clin. Chim. Acta 1978, 85, 107–110. [Google Scholar] [CrossRef]
- Opichka, M.A.; Rappelt, M.W.; Gutterman, D.D.; Grobe, J.L.; McIntosh, J.J. Vascular Dysfunction in Preeclampsia. Cells 2021, 10, 3055. [Google Scholar] [CrossRef]
- Chevallier, B.; Lagarde, A.; Degrelle, H.; Belaisch-Allart, J.; Giraudet, P.; Gallet, J.P. Insulin-like growth factor binding protein 1 level in amniotic fluid: Correlation with birth weight. Neonatology 1998, 73, 404–406. [Google Scholar] [CrossRef]
- Kratzer, I.; Bernhart, E.; Wintersperger, A.; Hammer, A.; Waltl, S.; Malle, E.; Sperk, G.; Wietzorrek, G.; Dieplinger, H.; Sattler, W. Afamin is synthesized by cerebrovascular endothelial cells and mediates α-tocopherol transport across an in vitro model of the blood–brain barrier. J. Neurochem. 2009, 108, 707–718. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nmb. | AC | Entry | Protein Description/Name | Score | Match | TP | DP | Theoretical | Experimental | FC | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mw, kDa | pI | Mw, kDa | pI | G1/G2 | ||||||||
| 1a | P00450 | CERU_HUMAN | Ceruloplasmin OS = Homo sapiens GN = CP PE = 1 SV = 1 | 100 | 20 | 107 | 21 | 122.128 | 5.44 | 155.47 | 5.00 | −1.57 |
| 1b | 100 | 12 | 13 | 137.48 | 4.93 | −2.11 | ||||||
| 2 | P01024 | CO3_HUMAN | Complement C3 OS = Homo sapiens GN = C3 PE = 1 SV = 2 | 99.95 | 11 | 196 | 21 | 187.030 | 6.02 | 138.40 | 5.40 | −1.05 |
| 3a | P01024 | CO3_HUMAN | Complement C3 OS = Homo sapiens GN = C3 PE = 1 SV = 2 | 100 | 15 | 196 | 29 | 187.030 | 6.02 | 77.14 | 6.51 | 1.29 |
| 3b | 98.66 | 14 | 28 | 76.93 | 6.38 | 1.05 | ||||||
| 4 | Q14624 | ITIH4_HUMAN | Inter-alpha-trypsin inhibitor heavy chain H4 OS = Homo sapiens GN = ITIH4 PE = 1 SV = 4 | 100 | 18 | 98 | 18 | 103.293 | 6.51 | 132.41 | 4.83 | −5.11 |
| 5 | P05155 | IC1_HUMAN | Plasma protease C1 inhibitor OS = Homo sapiens GN = SERPING1 PE = 1 SV = 2 | 100 | 7 | 44 | 3 | 55.119 | 6.09 | 110.98 | 4.16 | −2.93 |
| 6a | P06396 | GELS_HUMAN | Gelsolin OS = Homo sapiens GN = GSN PE = 1 SV = 1 | 100 | 13 | 88 | 11 | 85.644 | 5.9 | 100.95 | 5.49 | 1.08 |
| 6b | 94.55 | 2 | 2 | 100.68 | 5.41 | −1.12 | ||||||
| 6c | 100 | 8 | 7 | 100.47 | 5.35 | 1.12 | ||||||
| 7 | P00734 | THRB_HUMAN | Prothrombin OS = Homo sapiens GN = F2 PE = 1 SV = 2 | 99.99 | 5 | 74 | 4 | 69.992 | 5.64 | 89.00 | 4.80 | −12.51 |
| 8a | P04217 | A1BG_HUMAN | Alpha-1B-glycoprotein OS = Homo sapiens GN = A1BG PE = 1 SV = 4 | 100 | 16 | 45 | 7 | 54.220 | 5.56 | 79.91 | 4.95 | −2.36 |
| 8b | 99.89 | 11 | 5 | 92.19 | 4.93 | −4.39 | ||||||
| 9a | P02787 | TRFE_HUMAN | Serotransferrin OS = Homo sapiens GN = TF PE = 1 SV = 3 | 100 | 22 | 86 | 19 | 77.014 | 6.81 | 85.76 | 3.89 | −3.49 |
| 9b | 100 | 13 | 11 | 85.99 | 4.00 | −1.85 | ||||||
| 9c | 99.99 | 17 | 15 | 86.26 | 4.13 | −2.53 | ||||||
| 9d | 99.99 | 20 | 17 | 85.52 | 4.42 | −3.20 | ||||||
| 9e | 100 | 19 | 16 | 86.63 | 5.88 | 2.28 | ||||||
| 9f | 100 | 23 | 20 | 85.03 | 5.94 | 3.04 | ||||||
| 9g | 100 | 14 | 12 | 82.63 | 6.01 | 2.41 | ||||||
| 9h | 100 | 15 | 13 | 82.25 | 6.11 | 1.59 | ||||||
| 9i | 100 | 23 | 20 | 83.99 | 6.25 | 1.11 | ||||||
| 9j | 100 | 5 | 4 | 84.30 | 6.39 | −1.01 | ||||||
| 9k | 100 | 19 | 16 | 84.56 | 7.03 | 2.99 | ||||||
| 10a | P02768 | ALBU_HUMAN | Serum albumin OS = Homo sapiens GN = ALB PE = 1 SV = 2 | 100 | 24 | 88 | 21 | 69.321 | 5.92 | 70.29 | 5.59 | 5.45 |
| 10b | 100 | 19 | 17 | 75.28 | 5.38 | −1.37 | ||||||
| 11 | P02768 | ALBU_HUMAN | Serum albumin OS = Homo sapiens GN = ALB PE = 1 SV = 2 | 100 | 15 | 88 | 13 | 69.321 | 5.92 | 64.25 | 6.26 | 1.09 |
| 12 | P02748 | CO9_HUMAN | Complement component C9 OS = Homo sapiens GN = C9 PE = 1 SV = 2 | 100 | 17 | 70 | 12 | 63.133 | 5.43 | 72.20 | 4.94 | −3.02 |
| 13 | P01876 | IGHA1_HUMAN | Ig alpha-1 chain C region OS = Homo sapiens GN = IGHA1 PE = 1 SV = 2 | 99.96 | 23 | 26 | 6 | 37.631 | 6.08 | 64.70 | 5.26 | −1.32 |
| 14 | P01011 | AACT_HUMAN | Alpha-1-antichymotrypsin OS = Homo sapiens GN = SERPINA3 PE = 1 SV = 2 | 100 | 26 | 43 | 11 | 47.621 | 5.33 | 63.68 | 4.43 | −2.11 |
| 15a | P01009 | A1AT_HUMAN | Alpha-1-antitrypsin OS = Homo sapiens GN = SERPINA1 PE = 1 SV = 3 | 100 | 46 | 41 | 19 | 46.707 | 5.37 | 65.21 | 4.97 | −2.11 |
| 15b | 100 | 54 | 22 | 68.91 | 4.74 | −4.52 | ||||||
| 15c | 100 | 17 | 7 | 55.64 | 4.82 | −3.24 | ||||||
| 15d | 94.67 | 29 | 12 | 59.76 | 5.88 | 2.12 | ||||||
| 16 | P01009 | A1AT_HUMAN | Alpha-1-antitrypsin OS = Homo sapiens GN = SERPINA1 PE = 1 SV = 3 | 100 | 39 | 41 | 16 | 46.707 | 5.37 | 139.43 | 4.82 | −5.12 |
| 17 | P02765 | FETUA_HUMAN | Alpha-2-HS-glycoprotein OS = Homo sapiens GN = AHSG PE = 1 SV = 1 | 94.42 | 21 | 29 | 6 | 39.300 | 5.43 | 54.46 | 4.47 | −2.18 |
| 18a | P01859 | IGHG2_HUMAN | Ig gamma-2 chain C region OS = Homo sapiens GN = IGHG2 PE = 1 SV = 2 | 99.49 | 19 | 32 | 6 | 35.878 | 7.66 | 54.11 | 6.12 | 2.05 |
| 18b | 99.31 | 16 | 5 | 53.51 | 6.22 | 1.45 | ||||||
| 18c | 99.49 | 19 | 6 | 53.38 | 6.30 | 1.19 | ||||||
| 19 | P01860 | IGHG3_HUMAN | Ig gamma-3 chain C region OS = Homo sapiens GN = IGHG3 PE = 1 SV = 2 | 99.5 | 3 | 40 | 1 | 41.260 | 8.23 | 52.99 | 6.67 | −1.08 |
| 20a | P51884 | LUM_HUMAN | Lumican OS = Homo sapiens GN = LUM PE = 1 SV = 2 | 100 | 23 | 35 | 8 | 38.405 | 6.16 | 50.27 | 5.26 | −1.80 |
| 20b | 100 | 17 | 6 | 50.64 | 5.16 | −2.26 | ||||||
| 21 | P02763 | A1AG1_HUMAN | Alpha-1-acid glycoprotein 1 OS = Homo sapiens GN = ORM1 PE = 1 SV = 1 | 100 | 13 | 23 | 3 | 23.497 | 4.93 | 46.02 | 4.10 | −1.39 |
| 22a | P25311 | ZA2G_HUMAN | Zinc-alpha-2-glycoprotein OS = Homo sapiens GN = AZGP1 PE = 1 SV = 2 | 100 | 41 | 34 | 14 | 34.237 | 5.71 | 41.51 | 4.68 | −4.38 |
| 22b | 98.89 | 32 | 11 | 39.71 | 4.79 | −4.48 | ||||||
| 23 | P00738 | HPT_HUMAN | Haptoglobin OS = Homo sapiens GN = HP PE = 1 SV = 1 | 45.47 | 18 | 45 | 8 | 45.177 | 6.13 | 39.41 | 4.86 | −3.54 |
| 24a | P01857 | IGHG1_HUMAN | Ig gamma-1 chain C region OS = Homo sapiens GN = IGHG1 PE = 1 SV = 1 | 99.98 | 18 | 34 | 6 | 36.083 | 8.46 | 37.54 | 6.67 | 1.86 |
| 24b | 34.45 | 9 | 3 | 37.01 | 6.80 | −1.63 | ||||||
| 25a | P01857 | IGHG1_HUMAN | Ig gamma-1 chain C region OS = Homo sapiens GN = IGHG1 PE = 1 SV = 1 | 100 | 24 | 34 | 8 | 36.083 | 8.46 | 53.65 | 6.93 | 1.14 |
| 25b | 100 | 21 | 7 | 53.23 | 7.30 | −1.13 | ||||||
| 26 | P0C0L5 | CO4B_HUMAN | Complement C4-B OS = Homo sapiens GN = C4B PE = 1 SV = 2 | 99.36 | 6 | 187 | 12 | 192.631 | 6.89 | 29.70 | 6.29 | 1.25 |
| 27 | P08833 | IBP1_HUMAN | Insulin-like growth factor-binding protein 1 OS = Homo sapiens GN = IGFBP1 PE = 1 SV = 1 | 99.97 | 24 | 21 | 5 | 27.885 | 5.11 | 28.94 | 4.80 | −3.41 |
| 28 | P02760 | AMBP_HUMAN | Protein AMBP OS = Homo sapiens GN = AMBP PE = 1 SV = 1 | 100 | 8 | 38 | 3 | 38.974 | 5.95 | 28.51 | 4.89 | −1.41 |
| 29 | A0M8Q6 | LAC7_HUMAN | Ig lambda-7 chain C region OS = Homo sapiens GN = IGLC7 PE = 1 SV = 2 | 99.45 | 25 | 12 | 3 | 11.296 | 8.49 | 25.21 | 6.67 | 1.07 |
| 30a | P01834 | IGKC_HUMAN | Ig kappa chain C region OS = Homo sapiens GN = IGKC PE = 1 SV = 1 | 54.8 | 27 | 11 | 3 | 11.602 | 5.58 | 23.89 | 5.77 | 3.89 |
| 30b | 54.65 | 27 | 3 | 23.47 | 6.03 | 2.58 | ||||||
| 30c | 96.48 | 18 | 2 | 23.32 | 6.18 | 1.71 | ||||||
| 30d | 99.66 | 27 | 3 | 23.30 | 6.56 | 1.27 | ||||||
| 30e | 99.93 | 18 | 2 | 23.43 | 6.81 | 1.33 | ||||||
| 31 | P01622 | KV304_HUMAN | Ig kappa chain V-III region Ti OS = Homo sapiens PE = 1 SV = 1 | 54.65 | 20 | 10 | 2 | 11.781 | 8.72 | 23.39 | 7.02 | −1.38 |
| 32 | P98160 | PGBM_HUMAN | Basement membrane-specific heparan sulfate proteoglycan core protein OS = Homo sapiens GN = HSPG2 PE = 1 SV = 4 | 100 | 2 | 329 | 8 | 468.532 | 6.06 | 20.25 | 5.37 | −2.76 |
| 33a | P02766 | TTHY_HUMAN | Transthyretin OS = Homo sapiens GN = TTR PE = 1 SV = 1 | 100 | 21 | 14 | 3 | 15.877 | 5.52 | 14.10 | 5.21 | −2.08 |
| 33b | 47.6 | 36 | 5 | 14.22 | 5.00 | −2.48 | ||||||
| 34a | P02766 | TTHY_HUMAN | Transthyretin OS = Homo sapiens GN = TTR PE = 1 SV = 1 | 100 | 57 | 14 | 8 | 15.877 | 5.52 | 32.99 | 5.24 | −1.82 |
| 34b | 43.3 | 36 | 5 | 31.48 | 5.01 | −2.61 | ||||||
| Nmb. | AC | Entry | Protein Description/Name | Score | Match | TP | DP | Theoretical | Experimental | FC | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mw, kDa | pI | Mw, kDa | pI | G1/G2 | ||||||||
| 1A | P00450 | CERU_HUMAN | Ceruloplasmin OS = Homo sapiens GN = CP PE = 1 SV = 1 | 100 | 7 | 107 | 6 | 122.128 | 5.44 | 125.413 | 5.11 | 13.8 |
| 1B | 38.76 | 13 | 11 | 124.394 | 5.2 | 15.9 | ||||||
| 1C | 100 | 7 | 6 | 160.438 | 5.22 | 70.3 | ||||||
| 8A | P04217 | A1BG_HUMAN | Alpha-1B-glycoprotein OS = Homo sapiens GN = A1BG PE = 1 SV = 4 | 95.3 | 20 | 45 | 7 | 54.220 | 5.56 | 78.913 | 5.1 | 30.7 |
| 8B | 100 | 31 | 11 | 73.612 | 5.19 | 23.6 | ||||||
| 8C | 91.35 | 20 | 8 | 73.460 | 5.22 | 45.7 | ||||||
| 9A | P02787 | TRFE_HUMAN | Serotransferrin OS = Homo sapiens GN = TF PE = 1 SV = 3 | 100 | 24 | 86 | 17 | 77.014 | 6.81 | 82.460 | 4.72 | 27.6 |
| 9B | 99.99 | 21 | 16 | 82.617 | 4.85 | 5.0 | ||||||
| 9C | 99.99 | 17 | 13 | 82.309 | 4.97 | 7.2 | ||||||
| 9D | 100 | 20 | 13 | 82.416 | 5.04 | 13.1 | ||||||
| 9E | 92.46 | 15 | 11 | 81.032 | 6.1 | 12.3 | ||||||
| 9F | 100 | 16 | 12 | 87.691 | 6.33 | 5.1 | ||||||
| 9G | 100 | 23 | 17 | 86.703 | 6.37 | 7.7 | ||||||
| 9H | 100 | 25 | 17 | 86.995 | 6.46 | 7.5 | ||||||
| 9I | 100 | 24 | 16 | 87.640 | 6.51 | 6.4 | ||||||
| 9J | 99.99 | 22 | 16 | 88.003 | 6.6 | 8.0 | ||||||
| 10A | P02768 | ALBU_HUMAN | Serum albumin OS = Homo sapiens GN = ALB PE = 1 SV = 2 | 100 | 26 | 88 | 17 | 69.321 | 5.92 | 68.406 | 4.63 | 82.4 |
| 10B | 99.81 | 15 | 10 | 66.982 | 5.83 | 4.6 | ||||||
| 10C | 99.75 | 19 | 13 | 66.903 | 5.87 | 8.3 | ||||||
| 10D | 97.61 | 24 | 14 | 66.783 | 5.96 | 4.7 | ||||||
| 12 | P02748 | CO9_HUMAN | Complement component C9 OS = Homo sapiens GN = C9 PE = 1 SV = 2 | 99.64 | 21 | 70 | 12 | 63.133 | 5.43 | 67.943 | 5.18 | |
| 14A | P01011 | AACT_HUMAN | Alpha-1-antichymotrypsin OS = Homo sapiens GN = SERPINA3 PE = 1 SV = 2 | 99.99 | 36 | 43 | 12 | 47.621 | 5.33 | 63.391 | 4.42 | 28.4 |
| 14B | 100 | 37 | 13 | 64.130 | 4.4 | 21.1 | ||||||
| 14C | 100 | 40 | 14 | 64.980 | 4.37 | 31.8 | ||||||
| 14D | 100 | 36 | 12 | 65.103 | 4.32 | 18.7 | ||||||
| 15A | P01009 | A1AT_HUMAN | Alpha-1-antitrypsin OS = Homo sapiens GN = SERPINA1 PE = 1 SV = 3 | 95.46 | 25 | 41 | 12 | 46.707 | 5.37 | 52.167 | 4.81 | 2.8 |
| 15B | 100 | 36 | 15 | 52.340 | 4.9 | 5.3 | ||||||
| 15C | 99.48 | 34 | 13 | 51.942 | 4.98 | 1.9 | ||||||
| 15D | 100 | 43 | 20 | 55.594 | 5.12 | 16.9 | ||||||
| 15E | 100 | 43 | 18 | 56.410 | 5.06 | 10.0 | ||||||
| 15F | 100 | 16 | 8 | 57.418 | 5.04 | 16.4 | ||||||
| 15G | 100 | 45 | 16 | 58.708 | 5.02 | 19.8 | ||||||
| 15H | 100 | 51 | 21 | 57.694 | 4.99 | 10.5 | ||||||
| 15I | 100 | 53 | 22 | 58.746 | 4.93 | 8.3 | ||||||
| 15J | 100 | 38 | 15 | 54.340 | 4.87 | 8.0 | ||||||
| 16A | P01009 | A1AT_HUMAN | Alpha-1-antitrypsin OS = Homo sapiens GN = SERPINA1 PE = 1 SV = 3 | 99.99 | 35 | 41 | 15 | 46.707 | 5.37 | 125.779 | 4.98 | 65.1 |
| 16B | 77.76 | 36 | 41 | 14 | 124.943 | 5.05 | 34.9 | |||||
| 17A | P02765 | FETUA_HUMAN | Alpha-2-HS-glycoprotein OS = Homo sapiens GN = AHSG PE = 1 SV = 1 | 99.4 | 15 | 29 | 4 | 39.300 | 5.43 | 57.132 | 4.31 | 28.2 |
| 17B | 99.99 | 13 | 3 | 57.942 | 4.39 | 19.9 | ||||||
| 17C | 98.94 | 14 | 4 | 57.136 | 4.43 | 18.9 | ||||||
| 17D | 99.33 | 15 | 4 | 57.461 | 4.51 | 15.8 | ||||||
| 17E | 100 | 15 | 4 | 56.984 | 4.58 | 17.2 | ||||||
| 18 | P01859 | IGHG2_HUMAN | Ig gamma-2 chain C region OS = Homo sapiens GN = IGHG2 PE = 1 SV = 2 | 99.86 | 14 | 32 | 5 | 35.878 | 7.66 | 246.031 | 6.62 | |
| 20 | P51884 | LUM_HUMAN | Lumican OS = Homo sapiens GN = LUM PE = 1 SV = 2 | 99.99 | 15 | 35 | 6 | 38.405 | 6.16 | 51.234 | 4.69 | |
| 21 | P02763 | A1AG1_HUMAN | Alpha-1-acid glycoprotein 1 OS = Homo sapiens GN = ORM1 PE = 1 SV = 1 | 97.2 | 17 | 23 | 4 | 23.497 | 4.93 | 48.113 | 4.16 | |
| 22A | P25311 | ZA2G_HUMAN | Zinc-alpha-2-glycoprotein OS = Homo sapiens GN = AZGP1 PE = 1 SV = 2 | 100 | 35 | 34 | 12 | 34.237 | 5.71 | 42.866 | 4.92 | 40.5 |
| 22B | 99.99 | 26 | 9 | 42.745 | 5.02 | 59.5 | ||||||
| 23A | P00738 | HPT_HUMAN | Haptoglobin OS = Homo sapiens GN = HP PE = 1 SV = 1 | 95.86 | 18 | 45 | 8 | 45.177 | 6.13 | 44.988 | 4.97 | 68.2 |
| 23B | 95.86 | 18 | 45 | 8 | 43.946 | 5.11 | 31.8 | |||||
| 24A | P01857 | IGHG1_HUMAN | Ig gamma-1 chain C region OS = Homo sapiens GN = IGHG1 PE = 1 SV = 1 | 100 | 12 | 34 | 3 | 36.083 | 8.46 | 58.164 | 6.82 | 33.7 |
| 24B | 100 | 41 | 10 | 57.942 | 6.75 | 66.3 | ||||||
| 27A | P08833 | IBP1_HUMAN | Insulin-like growth factor-binding protein 1 OS = Homo sapiens GN = IGFBP1 PE = 1 SV = 1 | 75.3 | 29 | 21 | 6 | 27.885 | 5.11 | 31.024 | 5.07 | 55.4 |
| 27B | 24.73 | 19 | 4 | 30.460 | 5.3 | 44.6 | ||||||
| 28A | P02760 | AMBP_HUMAN | Protein AMBP OS = Homo sapiens GN = AMBP PE = 1 SV = 1 | 98.5 | 8 | 38 | 3 | 38.974 | 5.95 | 30.648 | 4.9 | 22.4 |
| 28B | 98.06 | 11 | 4 | 30.712 | 5.02 | 46.0 | ||||||
| 28C | 100 | 21 | 8 | 29.843 | 5.17 | 31.5 | ||||||
| 30A | P01834 | IGKC_HUMAN | Ig kappa chain C region OS = Homo sapiens GN = IGKC PE = 1 SV = 1 | 76.74 | 18 | 11 | 2 | 11.602 | 5.58 | 24.106 | 5.59 | 18.3 |
| 30B | 92.81 | 18 | 2 | 25.013 | 5.9 | 6.9 | ||||||
| 30C | 73.4 | 45 | 5 | 25.647 | 6.01 | 7.1 | ||||||
| 30D | 99.97 | 36 | 4 | 25.687 | 6.32 | 8.2 | ||||||
| 30E | 95.96 | 36 | 4 | 25.946 | 6.49 | 4.5 | ||||||
| 30F | 54.8 | 27 | 3 | 26.768 | 6.7 | 24.8 | ||||||
| 30G | 85.98 | 18 | 2 | 26.416 | 6.9 | 30.3 | ||||||
| 32A | P98160 | PGBM_HUMAN | Basement membrane-specific heparan sulfate proteoglycan core protein OS = Homo sapiens GN = HSPG2 PE = 1 SV = 4 | 97.93 | 3 | 329 | 9 | 468.532 | 6.06 | 21.035 | 5.59 | 57.8 |
| 32B | 100 | 5 | 15 | 21.546 | 5.82 | 42.2 | ||||||
| 33A | P02766 | TTHY_HUMAN | Transthyretin OS = Homo sapiens GN = TTR PE = 1 SV = 1 | 100 | 36 | 14 | 5 | 15.877 | 5.52 | 13.645 | 5.38 | 51.0 |
| 33B | 100 | 50 | 7 | 13.681 | 5.68 | 49.0 | ||||||
| 34 | P02766 | TTHY_HUMAN | Transthyretin OS = Homo sapiens GN = TTR PE = 1 SV = 1 | 100 | 36 | 14 | 5 | 15.877 | 5.52 | 34.100 | 5.63 | |
| 35 | P02751 | FINC_HUMAN | Fibronectin OS = Homo sapiens GN = FN1 PE = 1 SV = 4 | 34.38 | 16 | 204 | 28 | 262.460 | 5.46 | 281.640 | 5.43 | |
| 36 | P00450 | CERU_HUMAN | Ceruloplasmin OS = Homo sapiens GN = CP PE = 1 SV = 1 | 98.8 | 19 | 107 | 16 | 122.128 | 5.44 | 301.394 | 5.17 | |
| 37A | P02787 | TRFE_HUMAN | Serotransferrin OS = Homo sapiens GN = TF PE = 1 SV = 3 | 99.95 | 17 | 86 | 12 | 77.014 | 6.81 | 282.942 | 6.31 | 35.9 |
| 37B | 99.95 | 17 | 12 | 279.480 | 6.38 | 32.5 | ||||||
| 37C | 99.99 | 17 | 7 | 280.342 | 6.44 | 31.5 | ||||||
| 38 | P02768 | ALBU_HUMAN | Serum albumin OS = Homo sapiens GN = ALB PE = 1 SV = 2 | 89.83 | 24 | 88 | 14 | 69.321 | 5.92 | 250.992 | 5.88 | |
| 39A | P01008 | ANT3_HUMAN | Antithrombin-III OS = Homo sapiens GN = SERPINC1 PE = 1 SV = 1 | 90.29 | 33 | 60 | 13 | 52.569 | 6.32 | 58.761 | 5.43 | 18.5 |
| 39B | 100 | 31 | 12 | 60.463 | 5.38 | 13.7 | ||||||
| 39C | 100 | 38 | 16 | 59.465 | 5.34 | 67.8 | ||||||
| 40 | P01008 | ANT3_HUMAN | Antithrombin-III OS = Homo sapiens GN = SERPINC1 PE = 1 SV = 1 | 100 | 36 | 60 | 17 | 52.569 | 6.32 | 246.108 | 5.18 | |
| 41 | P43652 | AFAM_HUMAN | Afamin OS = Homo sapiens GN = AFM PE = 1 SV = 1 | 90.07 | 22 | 76 | 14 | 69.024 | 5.64 | 95.405 | 5.08 | |
| 42 | P01009 | A1AT_HUMAN | Alpha-1-antitrypsin OS = Homo sapiens GN = SERPINA1 PE = 1 SV = 3 | 100 | 17 | 41 | 7 | 46.707 | 5.37 | 232.691 | 5.03 | |
| 43A | P02790 | HEMO_HUMAN | Hemopexin OS = Homo sapiens GN = HPX PE = 1 SV = 2 | 100 | 24 | 46 | 10 | 51.643 | 6.55 | 68.403 | 5.31 | 47.2 |
| 43B | 98.19 | 16 | 6 | 69.730 | 5.52 | 52.8 | ||||||
| 44A | P01042 | KNG1_HUMAN | Kininogen-1 OS = Homo sapiens GN = KNG1 PE = 1 SV = 2 | 100 | 15 | 73 | 10 | 71.912 | 6.34 | 60.418 | 4.62 | 24.2 |
| 44B | 100 | 16 | 9 | 61.470 | 4.57 | 28.5 | ||||||
| 44C | 100 | 13 | 8 | 63.011 | 4.51 | 36.8 | ||||||
| 44D | 100 | 9 | 6 | 59.137 | 4.64 | 10.5 | ||||||
| 45 | P02749 | APOH_HUMAN | Beta-2-glycoprotein 1 OS = Homo sapiens GN = APOH PE = 1 SV = 3 | 19.26 | 8 | 41 | 3 | 38.273 | 8.34 | 60.134 | 6.33 | |
| 46 | P01019 | ANGT_HUMAN | Angiotensinogen OS = Homo sapiens GN = AGT PE = 1 SV = 1 | 71.35 | 22 | 39 | 7 | 53.121 | 5.87 | 55.993 | 5.37 | |
| 47A | P02750 | A2GL_HUMAN | Leucine-rich alpha-2-glycoprotein OS = Homo sapiens GN = LRG1 PE = 1 SV = 2 | 99.85 | 13 | 32 | 4 | 38.154 | 6.45 | 50.640 | 4.41 | 56.3 |
| 47B | 99.85 | 13 | 4 | 48.940 | 4.52 | 43.7 | ||||||
| 48 | P63261 | ACTG_HUMAN | Actin. cytoplasmic 2 OS = Homo sapiens GN = ACTG1 PE = 1 SV = 1 | 81.07 | 17 | 38 | 5 | 41.766 | 5.31 | 44.097 | 5.32 | |
| 49 | P10909 | CLUS_HUMAN | Clusterin OS = Homo sapiens GN = CLU PE = 1 SV = 1 | 76.74 | 7 | 59 | 4 | 52.461 | 5.89 | 37.061 | 4.78 | |
| 50 | P02452 | CO1A1_HUMAN | Collagen alpha-1(I) chain OS = Homo sapiens GN = COL1A1 PE = 1 SV = 5 | 99.7 | 3 | 129 | 4 | 138.857 | 5.6 | 32.106 | 5.25 | |
| 51 | P08123 | CO1A2_HUMAN | Collagen alpha-2(I) chain OS = Homo sapiens GN = COL1A2 PE = 1 SV = 7 | 99.98 | 5 | 122 | 6 | 129.235 | 9.08 | 29.514 | 6.02 | |
| 52 | P18136 | KV313_HUMAN | Ig kappa chain V-III region HIC OS = Homo sapiens PE = 2 SV = 2 | 91.75 | 20 | 10 | 2 | 14.080 | 6.18 | 26.543 | 6.57 | |
| 53 | P02753 | RET4_HUMAN | Retinol-binding protein 4 OS = Homo sapiens GN = RBP4 PE = 1 SV = 3 | 49.11 | 22 | 27 | 6 | 22.995 | 5.76 | 21.842 | 5.37 | |
| Proteins Corresponding Different Spots in AFP (pI 4–7) | Proteins with Higher Expression Level in AFP (pI 3–11) |
|---|---|
| Serotransferrin | Prothrombin |
| Kininogen-1 | Alpha-1-antitrypsin |
| Alpha-antichymotrypsin | Inter-alpha-trypsin inhibitor heavy chain H4 |
| Insulin-like growth factor-binding protein 1 | Ig kappa chain C region |
| Zinc-alpha-2-glycoprotein | Zinc-alpha-2-glycoprotein |
| Protein AMBP | Alpha-1B-glycoprotein |
| Ceruloplasmin | Serotransferrin |
| Transthyretin | Insulin-like growth factor-binding protein 1 |
| Haptoglobin | Complement component C9 |
| Alpha-1-antitrypsin | Plasma protease C1 inhibitor |
| Kininogen | Protein AMBP |
| Antithrombin-III | Transthyretin |
| Alpha-1B-glycoprotein | |
| Ceruloplasmin | |
| Alpha-1-antichymotrypsin | |
| Basement membrane-specific heparan sulfate proteoglycan core protein Alpha-1B-glycoprotein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navakauskienė, R.; Baronaitė, S.; Matuzevičius, D.; Krasovskaja, N.; Treigytė, G.; Arlauskienė, A.; Navakauskas, D. Comparative Proteomic Assessment of Normal vs. Polyhydramnios Amniotic Fluid Based on Computational Analysis. Biomedicines 2022, 10, 1821. https://doi.org/10.3390/biomedicines10081821
Navakauskienė R, Baronaitė S, Matuzevičius D, Krasovskaja N, Treigytė G, Arlauskienė A, Navakauskas D. Comparative Proteomic Assessment of Normal vs. Polyhydramnios Amniotic Fluid Based on Computational Analysis. Biomedicines. 2022; 10(8):1821. https://doi.org/10.3390/biomedicines10081821
Chicago/Turabian StyleNavakauskienė, Rūta, Sandra Baronaitė, Dalius Matuzevičius, Natalija Krasovskaja, Gražina Treigytė, Audronė Arlauskienė, and Dalius Navakauskas. 2022. "Comparative Proteomic Assessment of Normal vs. Polyhydramnios Amniotic Fluid Based on Computational Analysis" Biomedicines 10, no. 8: 1821. https://doi.org/10.3390/biomedicines10081821