
Accumulation of Arachidonic Acid, Precursor of Pro-Inflammatory Eicosanoids, in Adipose Tissue of Obese Women: Association with Breast Cancer Aggressiveness Indicators

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Population and Samples
2.2. Fatty Acid Profile of Adipose Tissues
2.3. Statistical Analysis
3. Results
3.1. Characteristics of the Population
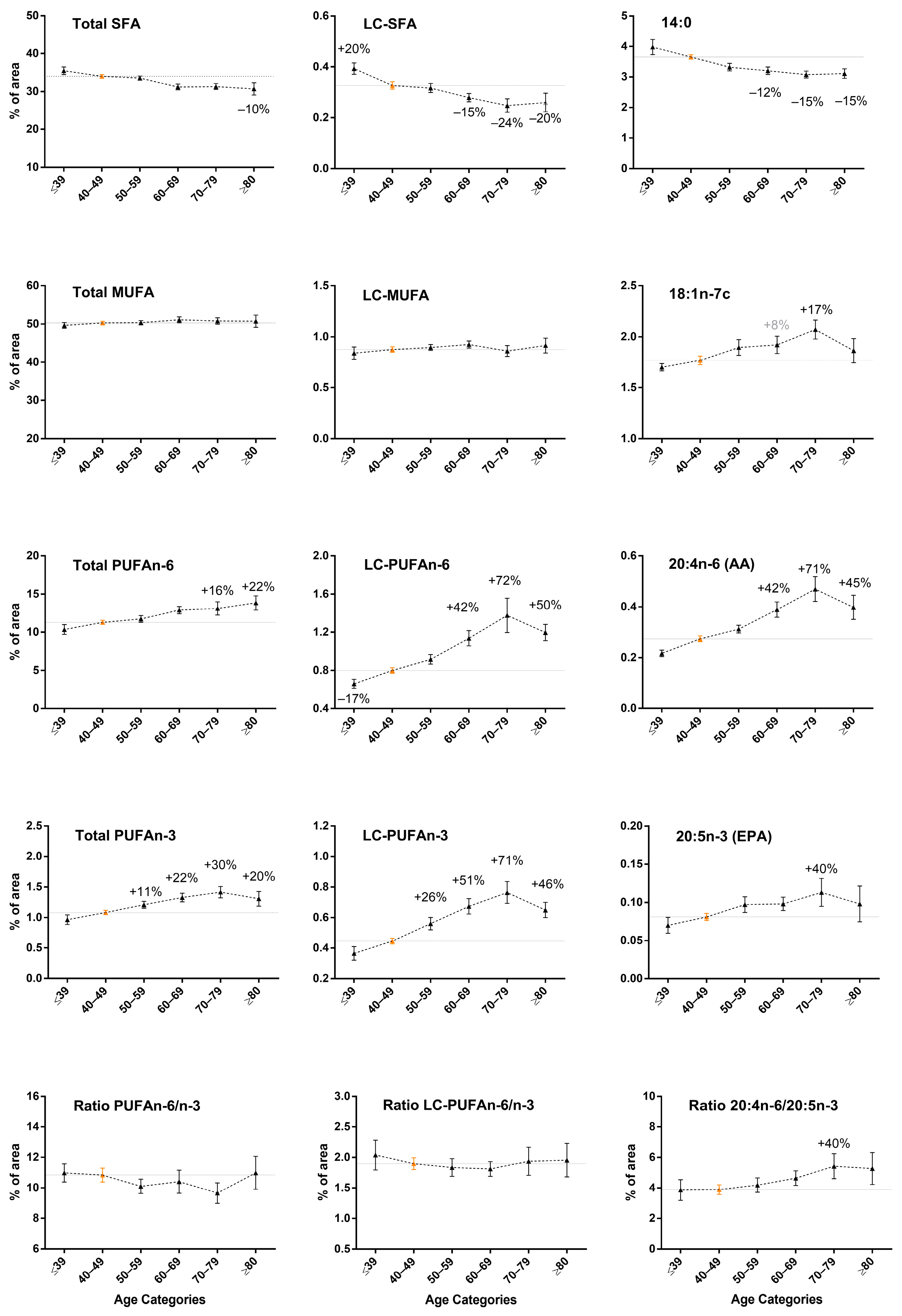
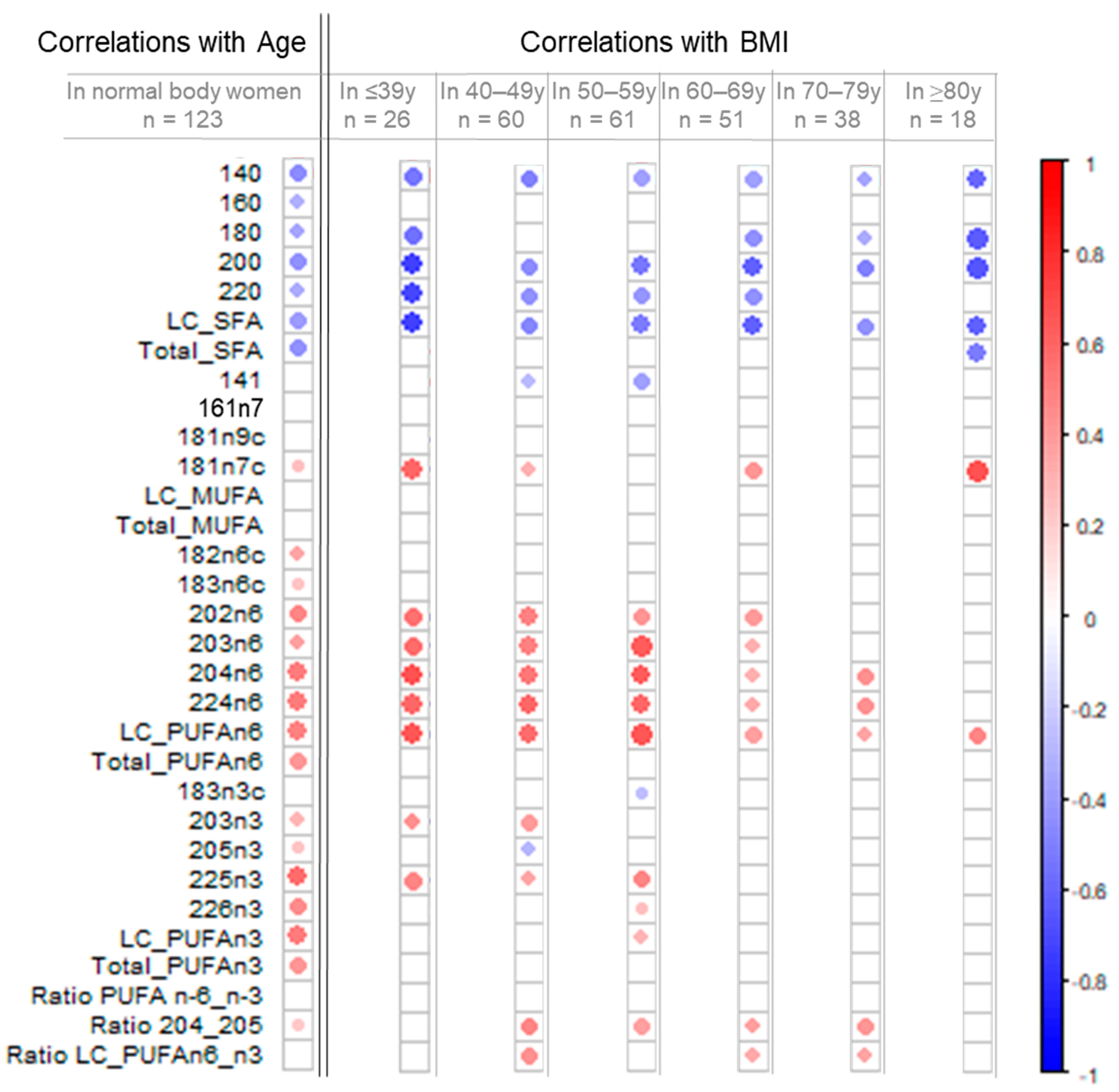
3.2. Increase in LC-PUFA n-6 and LC-PUFA n-3 Content in Breast Adipose Tissue by Age in Normal-Weight Women
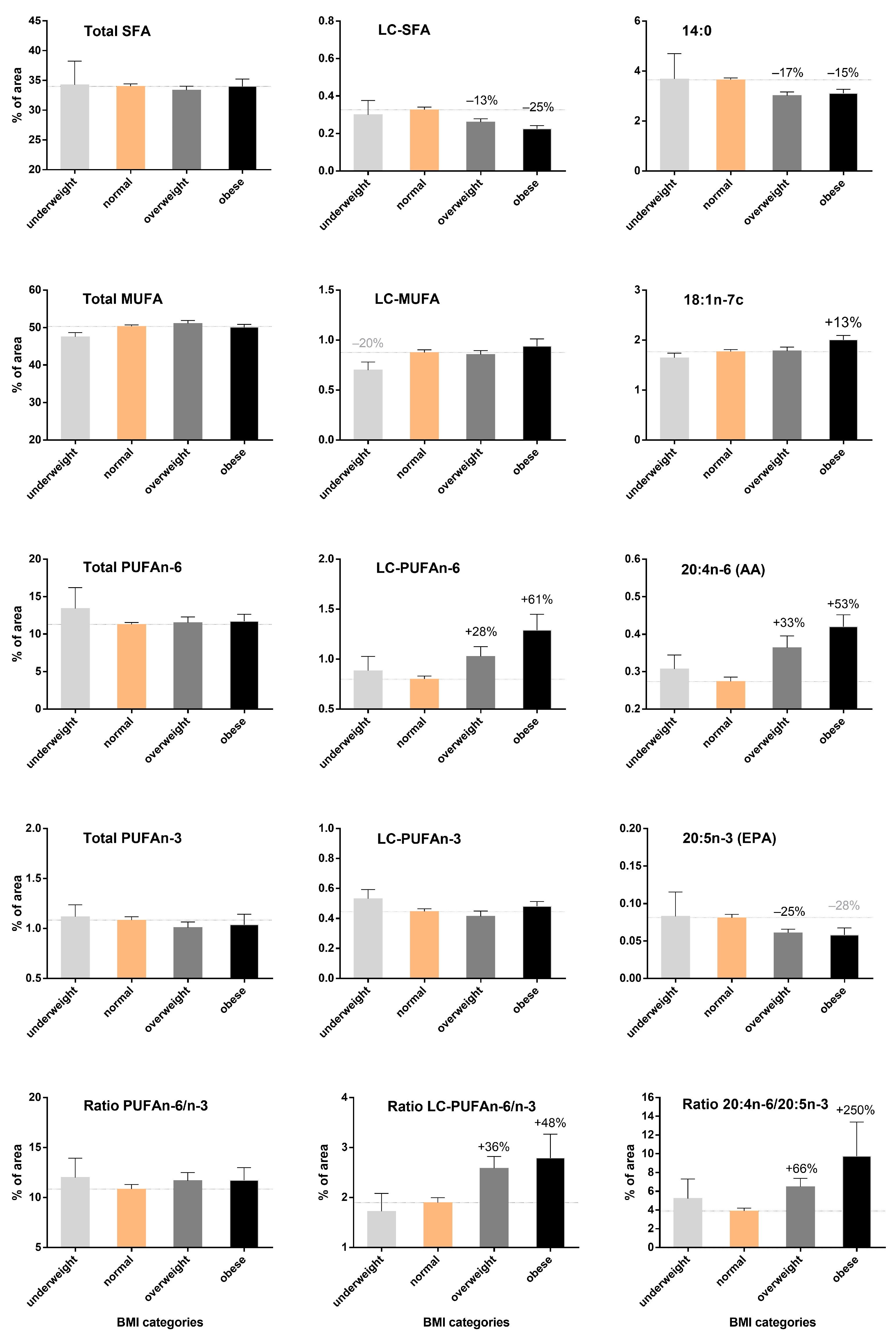
3.3. Increase in LC-PUFA n-6 According to BMI but Not LC-PUFA n-3 Content in Breast Adipose Tissue
3.4. Remodeling of the Fatty Acid Profile Induced by BMI Differs from Age-Induced Changes
3.5. Associations between Obesity and Indicators of Breast Tumor Aggressiveness
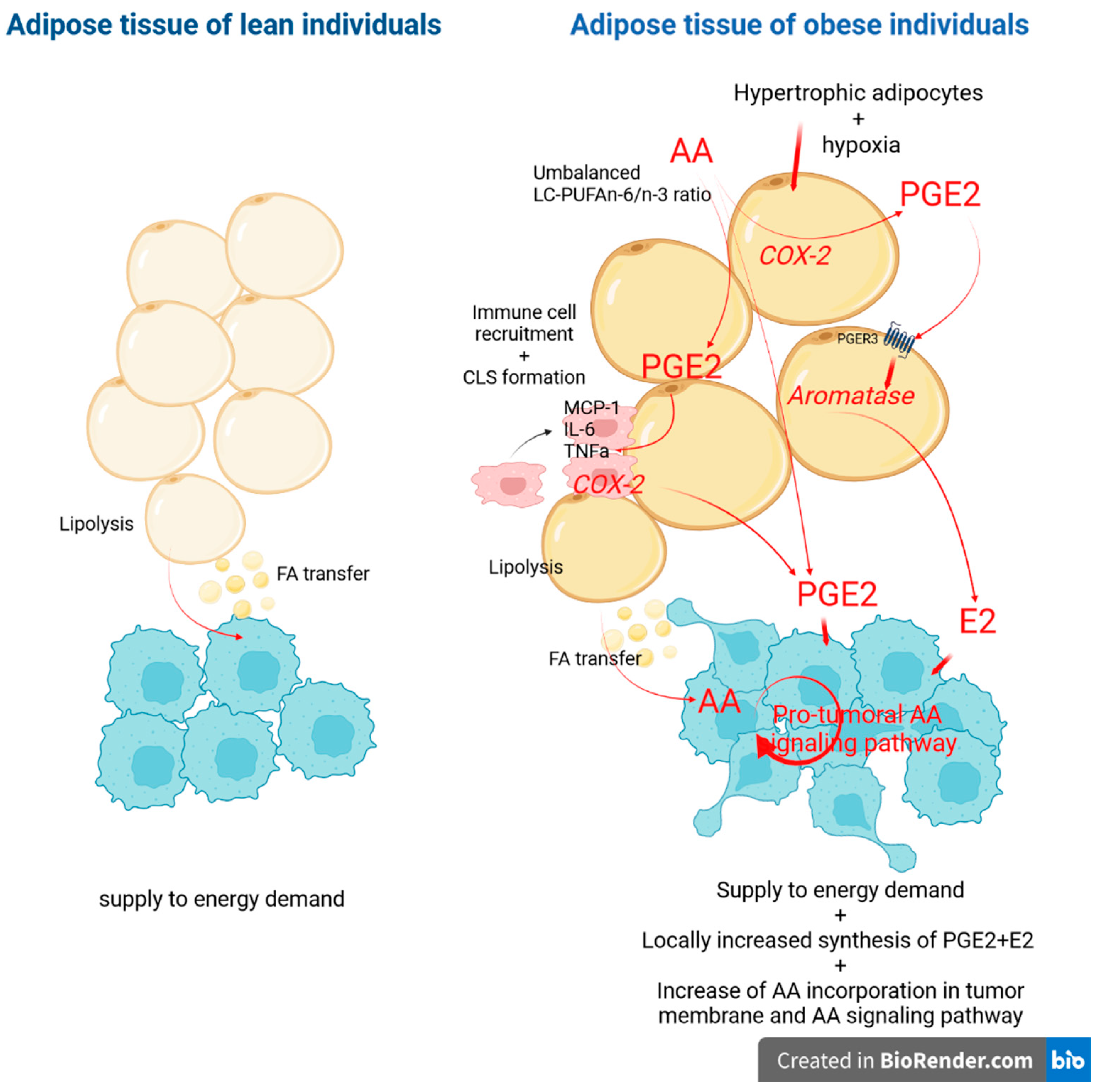
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Latino-Martel, P.; Cottet, V.; Druesne-Pecollo, N.; Pierre, F.H.F.; Touillaud, M.; Touvier, M.; Vasson, M.-P.; Deschasaux, M.; Le Merdy, J.; Barrandon, E.; et al. Alcoholic Beverages, Obesity, Physical Activity and Other Nutritional Factors, and Cancer Risk: A Review of the Evidence. Crit. Rev. Oncol. Hematol. 2016, 99, 308–323. [Google Scholar] [CrossRef]
- Bowers, L.W.; Maximo, I.X.F.; Brenner, A.J.; Beeram, M.; Hursting, S.D.; Price, R.S.; Tekmal, R.R.; Jolly, C.A.; deGraffenried, L.A. NSAID Use Reduces Breast Cancer Recurrence in Overweight and Obese Women: Role of Prostaglandin–Aromatase Interactions. Cancer Res. 2014, 74, 4446–4457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrelli, F.; Cortellini, A.; Indini, A.; Tomasello, G.; Ghidini, M.; Nigro, O.; Salati, M.; Dottorini, L.; Iaculli, A.; Varricchio, A.; et al. Association of Obesity with Survival Outcomes in Patients With Cancer: A Systematic Review and Meta-Analysis. JAMA Netw. Open 2021, 4, e213520. [Google Scholar] [CrossRef]
- Sun, L.; Zhu, Y.; Qian, Q.; Tang, L. Body Mass Index and Prognosis of Breast Cancer: An Analysis by Menstruation Status When Breast Cancer Diagnosis. Medicine 2018, 97, e11220. [Google Scholar] [CrossRef] [PubMed]
- Chlebowski, R.T. Nutrition and Physical Activity Influence on Breast Cancer Incidence and Outcome. Breast 2013, 22, S30–S37. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Avenell, A.; Bolland, M.; Hudson, J.; Stewart, F.; Robertson, C.; Sharma, P.; Fraser, C.; MacLennan, G. Effects of Weight Loss Interventions for Adults Who Are Obese on Mortality, Cardiovascular Disease, and Cancer: Systematic Review and Meta-Analysis. BMJ 2017, 359, j4849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avgerinos, K.I.; Spyrou, N.; Mantzoros, C.S.; Dalamaga, M. Obesity and Cancer Risk: Emerging Biological Mechanisms and Perspectives. Metabolism 2019, 92, 121–135. [Google Scholar] [CrossRef]
- Seiler, A.; Chen, M.A.; Brown, R.L.; Fagundes, C.P. Obesity, Dietary Factors, Nutrition, and Breast Cancer Risk. Curr Breast Cancer Rep. 2018, 10, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Zanoaga, O.; Jurj, A.; Raduly, L.; Cojocneanu-Petric, R.; Fuentes-Mattei, E.; Wu, O.; Braicu, C.; Gherman, C.; Berindan-Neagoe, I. Implications of Dietary Ω-3 and Ω-6 Polyunsaturated Fatty Acids in Breast Cancer (Review). Exp. Ther. Med. 2018, 15, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Djuricic, I.; Calder, P.C. Beneficial Outcomes of Omega-6 and Omega-3 Polyunsaturated Fatty Acids on Human Health: An Update for 2021. Nutrients 2021, 13, 2421. [Google Scholar] [CrossRef]
- Zheng, J.-S.; Hu, X.-J.; Zhao, Y.-M.; Yang, J.; Li, D. Intake of Fish and Marine N-3 Polyunsaturated Fatty Acids and Risk of Breast Cancer: Meta-Analysis of Data from 21 Independent Prospective Cohort Studies. BMJ 2013, 346, f3706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bougnoux, P.; Giraudeau, B.; Couet, C. Diet, Cancer, and the Lipidome. Cancer Epidemiol. Biomark. Prev. 2006, 15, 416–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouldamer, L.; Goupille, C.; Vildé, A.; Arbion, F.; Guimaraes, C.; Jourdan, M.; Bougnoux, P.; Body, G.; Chevalier, S. Total Long-Chain Polyunsaturated n-3 Fatty Acids Level Is an Independent Predictive Factor of Breast Cancer Multifocality in Women with Positive Hormone-Receptors Tumors. Surg. Oncol. 2021, 38, 101597. [Google Scholar] [CrossRef] [PubMed]
- Goupille, C.; Frank, P.G.; Arbion, F.; Jourdan, M.-L.; Guimaraes, C.; Pinault, M.; Body, G.; Chevalier, S.; Bougnoux, P.; Ouldamer, L. Low Levels of Omega-3 Long-Chain Polyunsaturated Fatty Acids Are Associated with Bone Metastasis Formation in Premenopausal Women with Breast Cancer: A Retrospective Study. Nutrients 2020, 12, 3832. [Google Scholar] [CrossRef]
- Atkinson, R.L.; El-Zein, R.; Valero, V.; Lucci, A.; Bevers, T.B.; Fouad, T.; Liao, W.; Ueno, N.T.; Woodward, W.A.; Brewster, A.M. Epidemiological Risk Factors Associated with Inflammatory Breast Cancer Subtypes. Cancer Causes Control 2016, 27, 359–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chas, M.; Goupille, C.; Arbion, F.; Bougnoux, P.; Pinault, M.; Jourdan, M.L.; Chevalier, S.; Ouldamer, L. Low Eicosapentaenoic Acid and Gamma-Linolenic Acid Levels in Breast Adipose Tissue Are Associated with Inflammatory Breast Cancer. Breast 2019, 45, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Schairer, C.; Laurent, C.A.; Moy, L.M.; Gierach, G.L.; Caporaso, N.E.; Pfeiffer, R.M.; Kushi, L.H. Obesity and Related Conditions and Risk of Inflammatory Breast Cancer: A Nested Case–Control Study. Breast Cancer Res. Treat. 2020, 183, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Strawford, A.; Antelo, F.; Christiansen, M.; Hellerstein, M.K. Adipose Tissue Triglyceride Turnover, de Novo Lipogenesis, and Cell Proliferation in Humans Measured with 2H2O. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E577–E588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolton-Smith, C.; Woodward, M.; Tavendale, R. Evidence for Age-Related Differences in the Fatty Acid Composition of Human Adipose Tissue, Independent of Diet. Eur. J. Clin. Nutr. 1997, 51, 619–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, A.K.; Kirkland, J.L. Aging and Adipose Tissue: Potential Interventions for Diabetes and Regenerative Medicine. Exp. Gerontol. 2016, 86, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Sun, K.; Kusminski, C.M.; Scherer, P.E. Adipose Tissue Remodeling and Obesity. J. Clin. Investig. 2011, 121, 2094–2101. [Google Scholar] [CrossRef] [Green Version]
- Martyniak, K.; Masternak, M.M. Changes in Adipose Tissue Cellular Composition during Obesity and Aging as a Cause of Metabolic Dysregulation. Exp. Gerontol. 2017, 94, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Ouldamer, L.; Goupille, C.; Vildé, A.; Arbion, F.; Body, G.; Chevalier, S.; Cottier, J.P.; Bougnoux, P. N-3 Polyunsaturated Fatty Acids of Marine Origin and Multifocality in Human Breast Cancer. PLoS ONE 2016, 11, e0147148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fekete, K.; Györei, E.; Lohner, S.; Verduci, E.; Agostoni, C.; Decsi, T. Long-Chain Polyunsaturated Fatty Acid Status in Obesity: A Systematic Review and Meta-Analysis: Meta-Analysis of Fatty Acid Status in Obesity. Obes Rev. 2015, 16, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Savva, S.C.; Chadjigeorgiou, C.; Hatzis, C.; Kyriakakis, M.; Tsimbinos, G.; Tornaritis, M.; Kafatos, A. Association of Adipose Tissue Arachidonic Acid Content with BMI and Overweight Status in Children from Cyprus and Crete. Br. J. Nutr. 2004, 91, 643–649. [Google Scholar] [CrossRef]
- Williams, E.S.; Baylin, A.; Campos, H. Adipose Tissue Arachidonic Acid and the Metabolic Syndrome in Costa Rican Adults. Clin. Nutr. 2007, 26, 474–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodson, L.; Skeaff, C.M.; Fielding, B.A. Fatty Acid Composition of Adipose Tissue and Blood in Humans and Its Use as a Biomarker of Dietary Intake. Prog. Lipid Res. 2008, 47, 348–380. [Google Scholar] [CrossRef] [PubMed]
- Yee, L.D.; Lester, J.L.; Cole, R.M.; Richardson, J.R.; Hsu, J.C.; Li, Y.; Lehman, A.; Belury, M.A.; Clinton, S.K. Omega-3 Fatty Acid Supplements in Women at High Risk of Breast Cancer Have Dose-Dependent Effects on Breast Adipose Tissue Fatty Acid Composition. Am. J. Clin. Nutr. 2010, 91, 1185–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baylin, A.; Kim, M.K.; Donovan-Palmer, A.; Siles, X.; Dougherty, L.; Tocco, P.; Campos, H. Fasting Whole Blood as a Biomarker of Essential Fatty Acid Intake in Epidemiologic Studies: Comparison with Adipose Tissue and Plasma. Am. J. Epidemiol. 2005, 162, 373–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baylin, A.; Kabagambe, E.K.; Siles, X.; Campos, H. Adipose Tissue Biomarkers of Fatty Acid Intake. Am. J. Clin. Nutr. 2002, 76, 750–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plourde, M.; Cunnane, S.C. Extremely Limited Synthesis of Long Chain Polyunsaturates in Adults: Implications for Their Dietary Essentiality and Use as Supplements. Appl. Physiol. Nutr. Metab. 2007, 32, 619–634. [Google Scholar] [CrossRef] [Green Version]
- Giltay, E.; Duschek, E.; Katan, M.; Zock, P.; Neele, S.; Netelenbos, J. Raloxifene and Hormone Replacement Therapy Increase Arachidonic Acid and Docosahexaenoic Acid Levels in Postmenopausal Women. J. Endocrinol. 2004, 182, 399–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Soto, M.; Mutch, D.M. Diet Regulation of Long-Chain PUFA Synthesis: Role of Macronutrients, Micronutrients, and Polyphenols on Δ-5/Δ-6 Desaturases and Elongases 2/5. Adv. Nutr. 2021, 12, 980–994. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, L.M.; Dutta, R.; Seeds, M.C.; Lake, K.N.; Hallmark, B.; Mathias, R.A.; Howard, T.D.; Chilton, F.H. FADS Genetic and Metabolomic Analyses Identify the ∆5 Desaturase (FADS1) Step as a Critical Control Point in the Formation of Biologically Important Lipids. Sci. Rep. 2020, 10, 15873. [Google Scholar] [CrossRef] [PubMed]
- de Meneses Fujii, T.M.; Norde, M.M.; Fisberg, R.M.; Marchioni, D.M.L.; Ordovás, J.M.; Rogero, M.M. FADS1 and ELOVL2 Polymorphisms Reveal Associations for Differences in Lipid Metabolism in a Cross-Sectional Population-Based Survey of Brazilian Men and Women. Nutr. Res. 2020, 78, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Metherel, A.H.; Irfan, M.; Klingel, S.L.; Mutch, D.M.; Bazinet, R.P. Higher Increase in Plasma DHA in Females Compared to Males Following EPA Supplementation May Be Influenced by a Polymorphism in ELOVL2: An Exploratory Study. Lipids 2021, 56, 211–228. [Google Scholar] [CrossRef] [PubMed]
- Macášek, J.; Zeman, M.; Žák, A.; Staňková, B.; Vecka, M. Altered Indices of Fatty Acid Elongases ELOVL6, ELOVL5, and ELOVL2 Activities in Patients with Impaired Fasting Glycemia. Metab. Syndr. Relat. Disord. 2021, 19, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Plourde, M.; Chouinard-Watkins, R.; Vandal, M.; Zhang, Y.; Lawrence, P.; Brenna, J.T.; Cunnane, S.C. Plasma Incorporation, Apparent Retroconversion and β-Oxidation of 13C-Docosahexaenoic Acid in the Elderly. Nutr. Metab. 2011, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- Misso, M.L.; Jang, C.; Adams, J.; Tran, J.; Murata, Y.; Bell, R.; Boon, W.C.; Simpson, E.R.; Davis, S.R. Differential Expression of Factors Involved in Fat Metabolism with Age and the Menopause Transition. Maturitas 2005, 51, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.-H.; Kim, H.-S. Menopause-Associated Lipid Metabolic Disorders and Foods Beneficial for Postmenopausal Women. Nutrients 2020, 12, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrara, C.M.; Lynch, N.A.; Nicklas, B.J.; Ryan, A.S.; Berman, D.M. Differences in Adipose Tissue Metabolism between Postmenopausal and Perimenopausal Women. J. Clin. Endocrinol. Metab. 2002, 87, 4166–4170. [Google Scholar] [CrossRef]
- Abbadia, Z.; Vericél, E.; Mathevet, P.; Bertin, N.; Panaye, G.; Frappart, L. Fatty Acid Composition and CD36 Expression in Breast Adipose Tissue of Premenopausal and Postmenopausal Women. Anticancer Res. 1997, 17, 1217–1221. [Google Scholar] [PubMed]
- McQuaid, S.E.; Hodson, L.; Neville, M.J.; Dennis, A.L.; Cheeseman, J.; Humphreys, S.M.; Ruge, T.; Gilbert, M.; Fielding, B.A.; Frayn, K.N.; et al. Downregulation of Adipose Tissue Fatty Acid Trafficking in Obesity: A Driver for Ectopic Fat Deposition? Diabetes 2011, 60, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Spalding, K.L.; Bernard, S.; Näslund, E.; Salehpour, M.; Possnert, G.; Appelsved, L.; Fu, K.-Y.; Alkass, K.; Druid, H.; Thorell, A.; et al. Impact of Fat Mass and Distribution on Lipid Turnover in Human Adipose Tissue. Nat. Commun. 2017, 8, 15253. [Google Scholar] [CrossRef]
- Summers, L.K.; Barnes, S.C.; Fielding, B.A.; Beysen, C.; Ilic, V.; Humphreys, S.M.; Frayn, K.N. Uptake of Individual Fatty Acids into Adipose Tissue in Relation to Their Presence in the Diet. Am. J. Clin. Nutr. 2000, 71, 1470–1477. [Google Scholar] [CrossRef]
- Raclot, T.; Langin, D.; Lafontan, M.; Groscolas, R. Selective Release of Human Adipocyte Fatty Acids According to Molecular Structure. Biochem J. 1997, 324 Pt 3, 911–915. [Google Scholar] [CrossRef] [Green Version]
- Boivin, L.; Diguisto, C.; Chas, M.; Bédouet, L.; Etienne, C.; Arbion, F.; Body, G.; Ouldamer, L. Indice de masse corporelle et facteurs pronostiques du cancer du sein. Gynécologie Obs. Fertil. Sénologie 2017, 45, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Gomes, R.; Costa, S.; Colquhoun, A. Eicosanoids and Cancer. Clinics 2018, 73, e530s. [Google Scholar] [CrossRef] [PubMed]
- Markosyan, N.; Smyth, E.M.; Patrignani, P.; Ricciotti, E. Editorial: Eicosanoids in Cancer. Front. Pharmacol. 2021, 12, 765214. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.; Hsiao, F.; Chang, H.; Wabitsch, M.; Hsieh, P.S. Importance of Adipocyte Cyclooxygenase-2 and Prostaglandin E 2 -prostaglandin E Receptor 3 Signaling in the Development of Obesity-induced Adipose Tissue Inflammation and Insulin Resistance. FASEB J. 2016, 30, 2282–2297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, R.M.; Delgobo, M.; Agnes, J.P.; das Neves, R.N.; Falchetti, M.; Casagrande, T.; Garcia, A.P.V.; Vieira, T.C.; Somensi, N.; Bruxel, M.A.; et al. COX-2 Promotes Mammary Adipose Tissue Inflammation, Local Estrogen Biosynthesis, and Carcinogenesis in High-Sugar/Fat Diet Treated Mice. Cancer Lett. 2021, 502, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Fain, J.N.; Kanu, A.; Bahouth, S.W.; Cowan, G.S.M.; Hiler, M.L.; Leffler, C.W. Comparison of PGE2, Prostacyclin and Leptin Release by Human Adipocytes versus Explants of Adipose Tissue in Primary Culture. Prostaglandins Leukot. Essent. Fat. Acids 2002, 67, 467–473. [Google Scholar] [CrossRef]
- Yarla, N.S.; Bishayee, A.; Sethi, G.; Reddanna, P.; Kalle, A.M.; Dhananjaya, B.L.; Dowluru, K.S.V.G.K.; Chintala, R.; Duddukuri, G.R. Targeting Arachidonic Acid Pathway by Natural Products for Cancer Prevention and Therapy. Semin. Cancer Biol. 2016, 40–41, 48–81. [Google Scholar] [CrossRef]
- Unamuno, X.; Gómez-Ambrosi, J.; Rodríguez, A.; Becerril, S.; Frühbeck, G.; Catalán, V. Adipokine Dysregulation and Adipose Tissue Inflammation in Human Obesity. Eur. J. Clin. Investig. 2018, 48, e12997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, G.I.; Mittendorfer, B.; Klein, S. Metabolically Healthy Obesity: Facts and Fantasies. J. Clin. Investig. 2019, 129, 3978–3989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raclot, T. Selective Mobilization of Fatty Acids from Adipose Tissue Triacylglycerols. Prog. Lipid Res. 2003, 42, 257–288. [Google Scholar] [CrossRef]
- Conner, W.E.; Lin, D.S.; Colvis, C. Differential Mobilization of Fatty Acids from Adipose Tissue. J. Lipid Res. 1996, 37, 290–298. [Google Scholar] [CrossRef]
- Martínez-Fernández, L.; Laiglesia, L.M.; Huerta, A.E.; Martínez, J.A.; Moreno-Aliaga, M.J. Omega-3 Fatty Acids and Adipose Tissue Function in Obesity and Metabolic Syndrome. Prostaglandins Other Lipid Mediat. 2015, 121, 24–41. [Google Scholar] [CrossRef]
- Liddle, D.; Hutchinson, A.; Wellings, H.; Power, K.; Robinson, L.; Monk, J. Integrated Immunomodulatory Mechanisms through Which Long-Chain n-3 Polyunsaturated Fatty Acids Attenuate Obese Adipose Tissue Dysfunction. Nutrients 2017, 9, 1289. [Google Scholar] [CrossRef] [Green Version]
- Fabian, C.J.; Kimler, B.F.; Hursting, S.D. Omega-3 Fatty Acids for Breast Cancer Prevention and Survivorship. Breast Cancer Res. 2015, 17, 62. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Ma, D. The Role of N-3 Polyunsaturated Fatty Acids in the Prevention and Treatment of Breast Cancer. Nutrients 2014, 6, 5184–5223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, C.; Plastina, P.; Barone, I.; Catalano, S.; Bonofiglio, D. N–3 Polyunsaturated Fatty Acid Amides: New Avenues in the Prevention and Treatment of Breast Cancer. IJMS 2020, 21, 2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BMI Category | |||||
|---|---|---|---|---|---|
| Underweight BMI < 18.5 kg/m2 n = 12 | Normal Weight BMI 18.5–24.9 kg/m2 n = 123 | Overweight BMI 25–29.9 kg/m2 n = 76 | Obese BMI ≥ 30 kg/m2 n = 50 | p Value | |
| Age (years), mean (SD) | 47.7 (14.2) | 55.7 (13.2) | 58.6 (13.2) | 59.4 (15.1) | 0.01 |
| BMI, mean (SD) | 17.3 (1.5) | 21.9 (1.6) | 26.8 (1.4) | 33.5 (3.4) | <0.0001 |
| Menopause | 5 (41.6%) | 68 (55.2%) | 53 (69.7%) | 37 (74.0%) | 0.02 |
| High blood pressure | 2 (16.7%) | 32 (26.0%) | 20 (26.0%) | 24 (48.0%) | 0.01 |
| Diabetes | 0 (0%) | 2 (1.6%) | 5 (6.6%) | 9 (18.0%) | 0.0006 |
| Dyslipidemia | 1 (8.3%) | 30 (24.4%) | 24 (31.6%) | 19 (38.0%) | 0.09 |
| Non-Obese n = 211 | Obese n = 50 | p Values | |
|---|---|---|---|
| Age (years) | 56.3 (13.5) | 59.4 (15.1) | 0.15 |
| BMI (kg/m²) | 23.4 (3.2) | 33.5 (3.4) | <0.0001 |
| Fatty acids content | |||
| LC-SFA | 0.28 (0.09) | 0.19 (0.06) | <0.0001 |
| LC-MUFA | 0.88 (0.17) | 0.90 (0.17) | 0.57 |
| 20:4n-6 (arachidonic acid) | 0.37 (0.14) | 0.50 (0.16) | <0.0001 |
| LC-PUFA n-6 | 1.07 (0.42) | 1.48 (0.41) | <0.0001 |
| LC-PUFA n-3 | 0.59 (0.25) | 0.66 (0.22) | 0.05 |
| Ratios | |||
| PUFA n-6/n-3 | 10.75 (3.4) | 10.87 (3.2) | 0.82 |
| 20:4n-6/20:5n-3 | 5.02 (2.93) | 7.30 (4.20) | <0.0001 |
| LC-PUFA n-6/n-3 | 1.99 (0.76) | 2.36 (0.73) | 0.002 |
| Tumor phenotype | 0.94 | ||
| Luminal A | 72 (34.1%) | 16 (32%) | |
| Luminal B | 59 (45.5%) | 16 (32%) | |
| HER2 | 28 (9.1%) | 7 (14%) | |
| Triple negative | 51 (9.1%) | 11 (22%) | |
| UK | 1 (0.5%) | - | |
| Tumor grade | 0.54 | ||
| 1 | 17 (8.1%) | 6 (12.0%) | |
| 2 | 95 (45.0%) | 23 (46.0%) | |
| 3 | 96 (45.5%) | 19 (38.0%) | |
| UK | 3 (1.4%) | 2 (4.0%) | |
| Tumor size | 28.2 (18.7) | 31.7 (19.3) | 0.23 |
| Multifocal tumor | 50 (23.7%) | 11 (22%) | 0.79 |
| Axillary positive lymph node | 94 (44.5%) | 31 (62.0%) | 0.02 |
| Inflammatory breast cancer | 19 (9.0%) | 9 (18.0%) | 0.08 |
| Carcinoma in situ | 122 (57.8%) | 20 (40%) | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouldamer, L.; Jourdan, M.-L.; Pinault, M.; Arbion, F.; Goupille, C. Accumulation of Arachidonic Acid, Precursor of Pro-Inflammatory Eicosanoids, in Adipose Tissue of Obese Women: Association with Breast Cancer Aggressiveness Indicators. Biomedicines 2022, 10, 995. https://doi.org/10.3390/biomedicines10050995
Ouldamer L, Jourdan M-L, Pinault M, Arbion F, Goupille C. Accumulation of Arachidonic Acid, Precursor of Pro-Inflammatory Eicosanoids, in Adipose Tissue of Obese Women: Association with Breast Cancer Aggressiveness Indicators. Biomedicines. 2022; 10(5):995. https://doi.org/10.3390/biomedicines10050995
Chicago/Turabian StyleOuldamer, Lobna, Marie-Lise Jourdan, Michelle Pinault, Flavie Arbion, and Caroline Goupille. 2022. "Accumulation of Arachidonic Acid, Precursor of Pro-Inflammatory Eicosanoids, in Adipose Tissue of Obese Women: Association with Breast Cancer Aggressiveness Indicators" Biomedicines 10, no. 5: 995. https://doi.org/10.3390/biomedicines10050995