
Inorganic Polymeric Materials for Injured Tissue Repair: Biocatalytic Formation and Exploitation

 , , ,
, , , 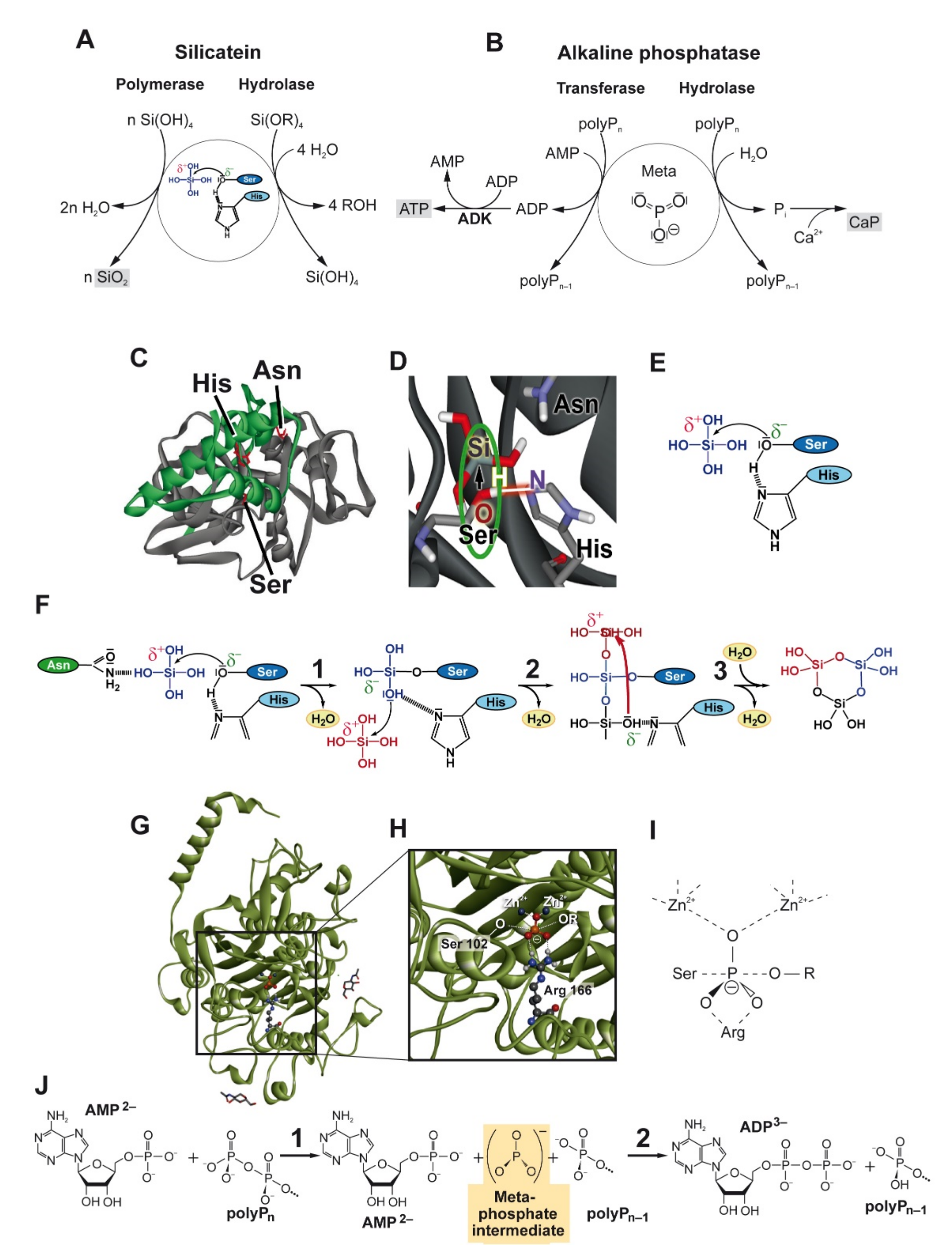{kind=link}
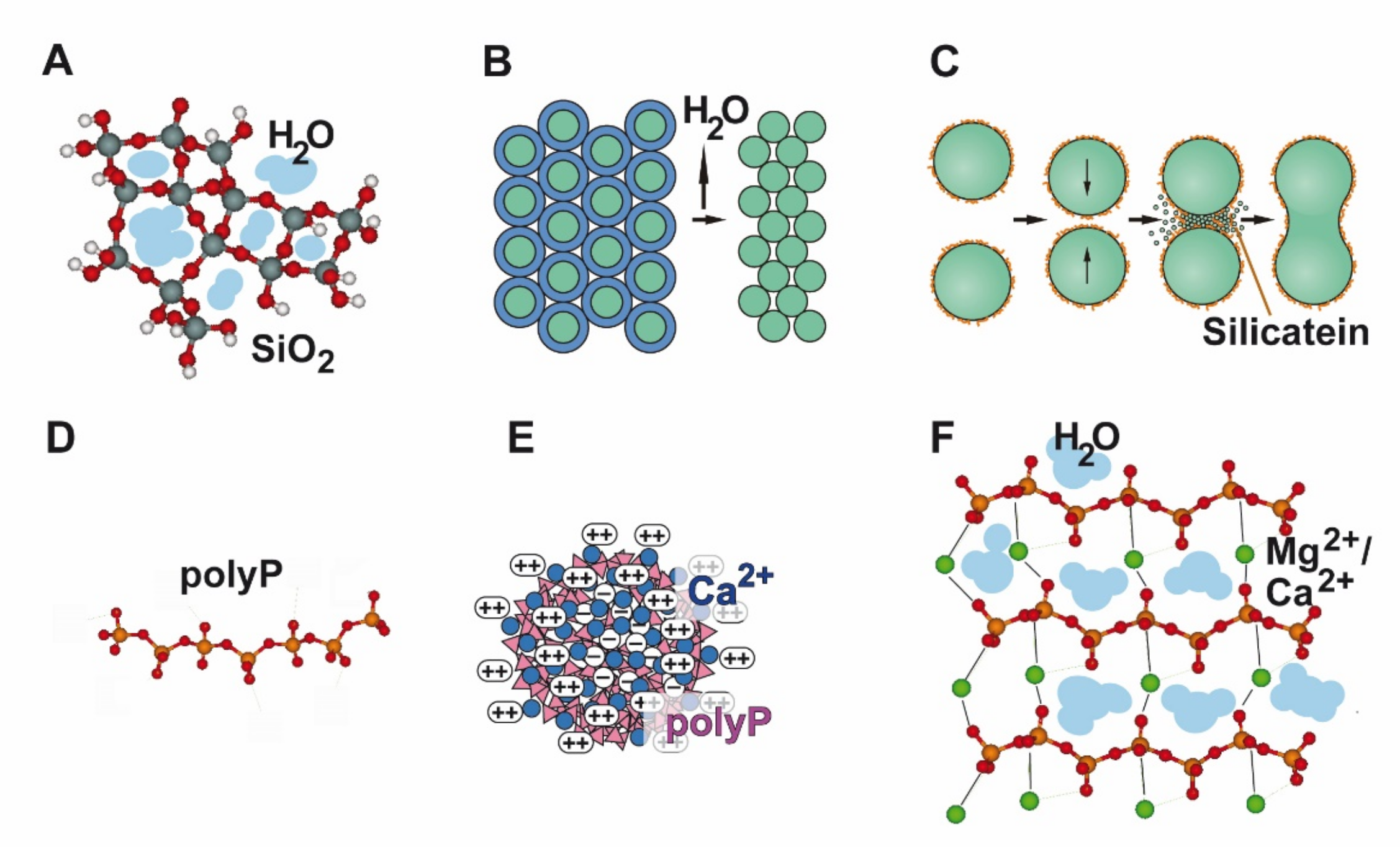{kind=link}
{kind=link}
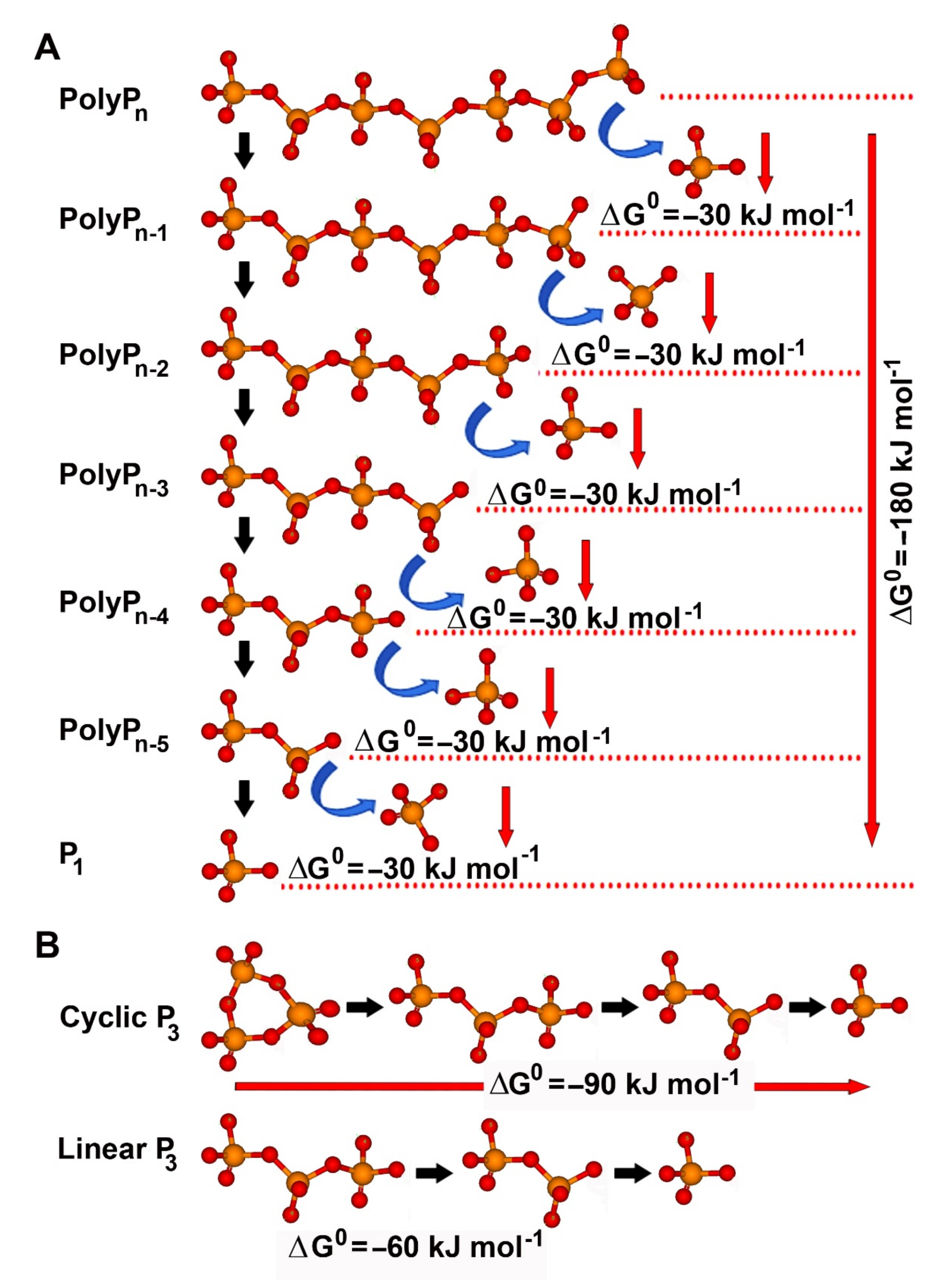{kind=link}
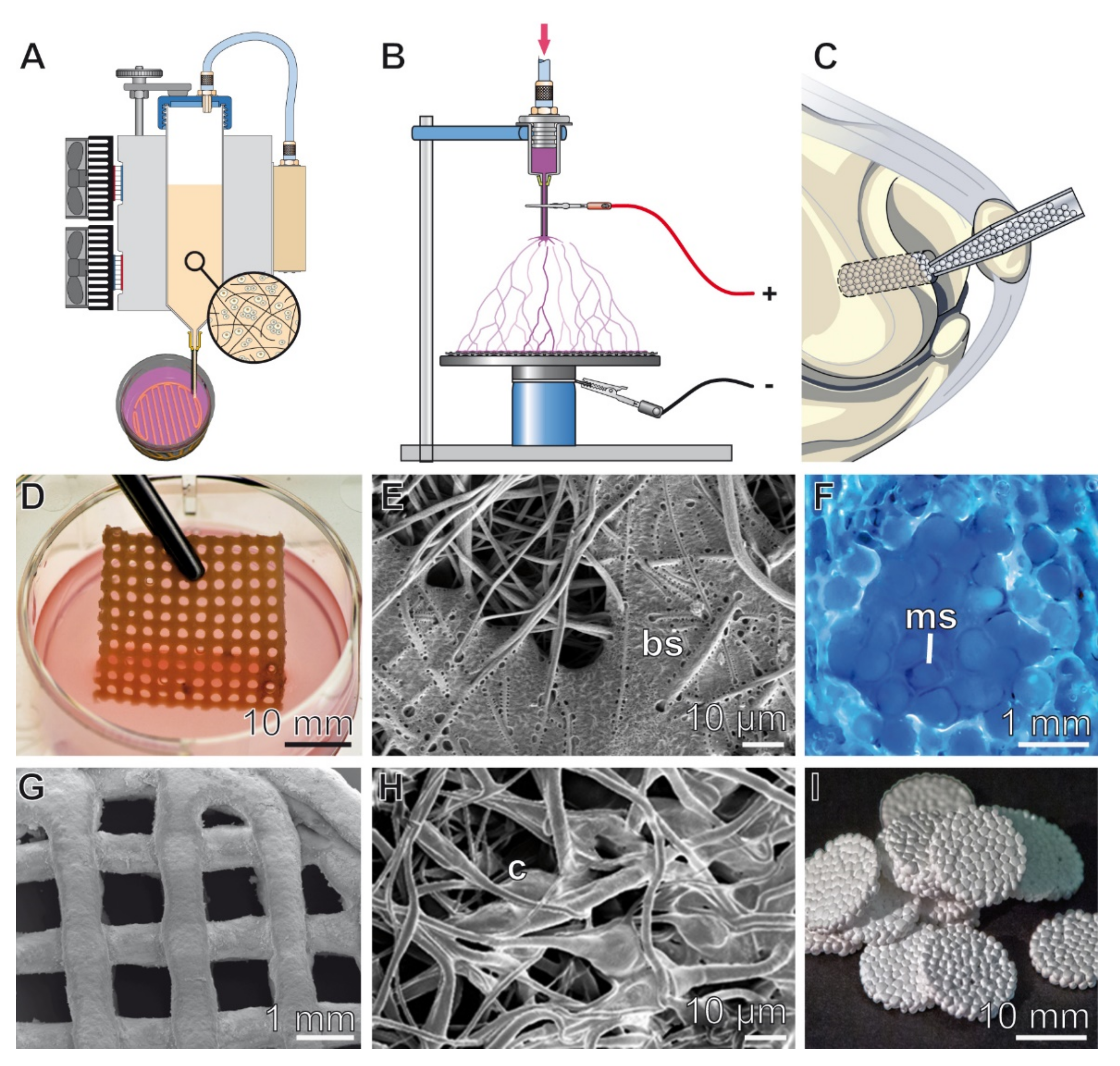{kind=link}
Abstract
:1. Introduction
2. Differential Characteristics of the Si-O-Si and P-O-P Linkages
3. Silicatein and Biosilica Formation
3.1. Mechanism of Silicatein Reaction
3.2. Silicatein Assembly
3.3. Biosynthesis and Processing of Silicatein
4. Alkaline Phosphatase and (Poly)Phosphate-Based Materials
4.1. Hydrolytic Cleavage of Polyphosphate
4.2. Phosphotransfer Reactions
4.3. Further Functions
5. Different Forms and Phases of Biosilica and Bioinorganic Polyphosphate
5.1. Silica
5.1.1. Gelation
5.1.2. Syneresis
5.1.3. Biosintering
5.2. PolyP–PolyP Nano/Microparticles
5.2.1. Calcium-PolyP
5.2.2. Magnesium-PolyP
5.2.3. Strontium-PolyP
5.2.4. Zinc-PolyP
5.2.5. Amorphous Ca-Phosphate Stabilized with PolyP
5.2.6. PolyP Coacervates
6. Properties of Amorphous Silica and PolyP as Regeneratively Active Inorganic Polymers
6.1. Morphogenetic Activity
6.2. Generation of Metabolic Energy
7. Routes of Administration of Biosilica and Polyphosphate for Tissue Regeneration/Repair
7.1. 3D Printing
7.2. Electrospinning
7.3. Microspheres
8. Biohybrid Formation with Hydrogel Forming Polymers
9. Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peacock, M. Phosphate metabolism in health and disease. Calcif. Tissue Int. 2021, 108, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Westheimer, F.H. Why nature chose phosphates. Science 1987, 235, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.G.; Schröder, H.C.; Wang, X.H. Inorganic polyphosphates as storage for and generator of metabolic energy in the extracellular matrix. Chem. Rev. 2019, 119, 12337–12374. [Google Scholar] [CrossRef] [PubMed]
- Moura, H.M.; Unterlass, M.M. Biogenic metal oxides. Biomimetics 2020, 5, 29. [Google Scholar] [CrossRef]
- Hildebrand, M.; Lerch, S.J.L.; Shrestha, R.P. Understanding diatom cell wall silicification—Moving forward. Front. Mar. Sci. 2018, 5, 125. [Google Scholar] [CrossRef] [Green Version]
- Guerriero, G.; Stokes, I.; Valle, N.; Hausman, J.F.; Exley, C. Visualising silicon in plants: Histochemistry, silica sculptures and elemental imaging. Cells 2020, 9, 1066. [Google Scholar] [CrossRef] [Green Version]
- Carlisle, E.M. Silicon as a trace nutrient. Sci. Total Environ. 1988, 73, 95–106. [Google Scholar] [CrossRef]
- Rondanelli, M.; Faliva, M.A.; Peroni, G.; Gasparri, C.; Perna, S.; Riva, A.; Petrangolini, G.; Tartara, A. Silicon: A neglected micronutrient essential for bone health. Exp. Biol. Med. 2021, 246, 1500–1511. [Google Scholar] [CrossRef]
- Weiner, S.; Mahamid, J.; Politi, Y.; Ma, Y.; Addadi, L. Overview of the amorphous precursor phase strategy in biomineralization. Front. Mater. Sci. China 2009, 3, 104. [Google Scholar] [CrossRef]
- Wang, X.H.; Schröder, H.C.; Müller, W.E.G. Enzyme-based biosilica and biocalcite: Biomaterials for the future in regenerative medicine. Trends Biotechnol. 2014, 32, 441–447. [Google Scholar] [CrossRef]
- Wang, X.H.; Schröder, H.C.; Müller, W.E.G. Amorphous polyphosphate, a smart bioinspired nano-/bio-material for bone and cartilage regeneration: Towards a new paradigm in tissue engineering. J. Mat. Chem. B 2018, 6, 2385–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krasko, A.; Lorenz, B.; Batel, R.; Schröder, H.C.; Müller, I.M.; Müller, W.E.G. Expression of silicatein and collagen genes in the marine sponge Suberites domuncula is controlled by silicate and myotrophin. Eur. J. Biochem. 2000, 267, 4878–4887. [Google Scholar] [CrossRef] [PubMed]
- Rao, N.N.; Gomez-Garcia, M.R.; Kornberg, A. Inorganic polyphosphate: Essential for growth and survival. Annu. Rev. Biochem. 2009, 78, 605–647. [Google Scholar] [CrossRef] [PubMed]
- Dietzel, M. Dissolution of silicates and the stability of polysilicic acid. Geochim. Cosmochim. Acta 2000, 64, 3275–3281. [Google Scholar] [CrossRef]
- De Jager, H.J.; Heyns, A.M. Kinetics of acid-catalyzed hydrolysis of a polyphosphate in water. J. Phys. Chem. A 1998, 102, 2838–2841. [Google Scholar] [CrossRef]
- Brady, P.V.; Walther, J.V. Kinetics of quartz dissolution at low temperatures. Chem. Geol. 1990, 82, 253–264. [Google Scholar] [CrossRef]
- Pelmenschikov, A.; Leszczynski, J.; Pettersson, L.G.M. Mechanism of dissolution of neutral silica surfaces: Including effect of self-healing. J. Phys. Chem. A 2001, 105, 9528–9532. [Google Scholar] [CrossRef]
- Trinh, T.T.; Jansen, A.P.; Santen, R.A. Mechanism of oligomerization reactions of silica. J. Phys. Chem. B 2006, 110, 23099–23106. [Google Scholar] [CrossRef]
- Liu, X.; Liu, C.; Meng, C. Oligomerization of silicic acids in neutral aqueous solution: A first-principles investigation. Int. J. Mol. Sci. 2019, 20, 3037. [Google Scholar] [CrossRef] [Green Version]
- Hunter, T. Why nature chose phosphate to modify proteins. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 2513–2516. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Schloßmacher, U.; Wang, X.H.; Boreiko, A.; Brandt, D.; Wolf, S.E.; Tremel, W.; Schröder, H.C. Poly(silicate)-metabolizing silicatein in siliceous spicules and silicasomes of demosponges comprises dual enzymatic activities (silica-polymerase and silica-esterase). FEBS J. 2008, 275, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Ojovan, M.I.; Tournier, R.F. On structural rearrangements near the glass transition temperature in amorphous silica. Materials 2021, 14, 5235. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Cha, J.; Stucky, G.D.; Morse, D.E. Silicatein alpha: Cathepsin L-like protein in sponge biosilica. Proc. Natl. Acad. Sci. USA 1998, 95, 6234–6238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, J.N.; Shimizu, K.; Zhou, Y.; Christianssen, S.C.; Chmelka, B.F.; Stucky, G.D.; Morse, D.E. Silicatein filaments and subunits from a marine sponge direct the polymerization of silica and silicones in vitro. Proc. Natl. Acad. Sci. USA 1999, 96, 361–365. [Google Scholar] [CrossRef] [Green Version]
- Schloßmacher, U.; Wiens, M.; Schröder, H.C.; Wang, X.H.; Jochum, K.P.; Müller, W.E.G. Silintaphin-1: Interaction with silicatein during structure-guiding biosilica formation. FEBS J. 2011, 278, 1145–1155. [Google Scholar] [CrossRef]
- Belton, D.J.; Deschaume, O.; Patwardhan, S.V.; Perry, C.C. A solution study of silica condensation and speciation with relevance to in vitro investigations of biosilicification. J. Phys. Chem. B 2010, 114, 9947–9955. [Google Scholar] [CrossRef] [Green Version]
- Belton, D.J.; Deschaume, O.; Perry, C.C. An overview of the fundamentals of the chemistry of silica with relevance to biosilicification and technological advances. FEBS J. 2012, 279, 1710–1720. [Google Scholar] [CrossRef]
- Schröder, H.C.; Wiens, M.; Schloßmacher, U.; Brandt, D.; Müller, W.E.G. Silicatein-mediated polycondensation of orthosilicic acid: Modeling of catalytic mechanism involving ring formation. Silicon 2012, 4, 33–38. [Google Scholar] [CrossRef]
- Zhou, Y.; Shimizu, K.; Cha, J.N.; Stucky, G.D.; Morse, D.E. Efficient catalysis of polysiloxane synthesis by silicatein a requires specific hydroxy and imidazole functionalities. Angew. Chem. Int. Ed. 1999, 38, 780–782. [Google Scholar] [CrossRef]
- Tabatabaei Dakhili, S.Y.; Caslin, S.A.; Faponle, A.S.; Quayle, P.; de Visser, S.P.; Wong, L.S. Recombinant silicateins as model biocatalysts in organosiloxane chemistry. Proc. Natl. Acad. Sci. USA 2017, 114, E5285–E5291. [Google Scholar] [CrossRef] [Green Version]
- Müller, W.E.G.; Schröder, H.C.; Muth, S.; Gietzen, S.; Korzhev, M.; Grebenjuk, V.; Wiens, M.; Schloßmacher, U.; Wang, X.H. The silicatein-propeptide acts as inhibitor/modulator of self-organization during spicule axial filament formation. FEBS J. 2013, 280, 1693–1708. [Google Scholar] [CrossRef] [PubMed]
- Schröder, H.C.; Wang, X.H.; Tremel, W.; Ushijima, H.; Müller, W.E.G. Biofabrication of biosilica-glass by living organisms. Nat. Prod. Rep. 2008, 25, 455–474. [Google Scholar] [CrossRef] [PubMed]
- Görlich, S.; Samuel, A.J.; Best, R.J.; Seidel, R.; Vacelet, J.; Leonarski, F.K.; Tomizaki, T.; Rellinghaus, B.; Pohl, D.; Zlotnikov, I. Natural hybrid silica/protein superstructure at atomic resolution. Proc. Natl. Acad. Sci. USA 2020, 117, 31088–31093. [Google Scholar] [CrossRef] [PubMed]
- Zlotnikov, I.; Werner, P.; Blumtritt, H.; Graff, A.; Dauphin, Y.; Zolotoyabko, E.; Fratzl, P. A perfectly periodic three-dimensional protein/silica mesoporous structure produced by an organism. Adv. Mater. 2014, 26, 1682–1687. [Google Scholar] [CrossRef]
- Sparkes, E.I.; Kettles, R.A.; Egedeuzu, C.S.; Stephenson, N.L.; Caslin, S.A.; Tabatabaei Dakhili, S.Y.; Wong, L.S. Improved production and biophysical analysis of recombinant silicatein-α. Biomolecules 2020, 10, 1209. [Google Scholar] [CrossRef]
- Godigamuwa, K.; Nakashima, K.; Okamoto, J.; Kawasaki, S. Biological route to fabricate silica on cellulose using immobilized silicatein fused with a carbohydrate-binding module. Biomacromolecules 2020, 21, 2922–2928. [Google Scholar] [CrossRef] [PubMed]
- Kulaev, I.S.; Vagabov, V.M.; Kulakovskaya, T.V. The Biochemistry of Inorganic Polyphosphates; John Wiley & Sons Ltd.: Chichester, England, 2004. [Google Scholar]
- Pavlov, E.; Aschar-Sobbi, R.; Campanella, M.; Turner, R.J.; Gómez-García, M.R.; Abramov, A.Y. Inorganic polyphosphate and energy metabolism in mammalian cells. J. Biol. Chem. 2010, 285, 9420–9428. [Google Scholar] [CrossRef] [Green Version]
- Angelova, P.R.; Baev, A.Y.; Berezhnov, A.V.; Abramov, A.Y. Role of inorganic polyphosphate in mammalian cells: From signal transduction and mitochondrial metabolism to cell death. Biochem. Soc. Trans. 2016, 44, 40–45. [Google Scholar] [CrossRef]
- Hothorn, M.; Neumann, H.; Lenherr, E.D.; Wehner, M.; Rybin, V.; Hassa, P.O.; Uttenweiler, A.; Reinhardt, M.; Schmidt, A.; Seiler, J.; et al. Catalytic core of a membrane-associated eukaryotic polyphosphate polymerase. Science 2009, 324, 513–516. [Google Scholar] [CrossRef]
- Docampo, R.; de Souza, W.; Miranda, K.; Rohloff, P.; Moreno, S.N. Acidocalcisomes—Conserved from bacteria to man. Nat. Rev. Microbiol. 2005, 3, 251–261. [Google Scholar] [CrossRef]
- Lorenz, B.; Schröder, H.C. Mammalian intestinal alkaline phosphatase acts as highly active exopolyphosphatase. Biochim. Biophys. Acta 2001, 1547, 254–261. [Google Scholar] [CrossRef]
- Omelon, S.; Georgiou, J.; Henneman, Z.J.; Wise, L.M.; Sukhu, B.; Hunt, T.; Wynnyckyj, C.; Holmyard, D.; Ryszard, B.; Grynpas, M.D. Control of vertebrate skeletal mineralization by polyphosphates. PLoS ONE 2009, 4, e5634. [Google Scholar] [CrossRef] [Green Version]
- Millán, J.L. Alkaline phosphatases—Structure, substrate specificity and functional relatedness to other members of a large superfamily of enzymes. Purinergic Signal. 2006, 2, 335–341. [Google Scholar] [CrossRef] [Green Version]
- Millán, J.L.; Whyte, M.P. Alkaline phosphatase and hypophosphatasia. Calcif. Tissue Int. 2016, 98, 398–416. [Google Scholar] [CrossRef] [Green Version]
- Müller, W.E.G.; Wang, S.F.; Neufurth, M.; Kokkinopoulou, M.; Feng, Q.L.; Schröder, H.C.; Wang, X.H. Polyphosphate as a donor of high-energy phosphate for the synthesis of ADP and ATP. J. Cell Sci. 2017, 130, 2747–2756. [Google Scholar] [CrossRef] [Green Version]
- Kornberg, A.; Rao, N.N.; Ault-Riché, D. Inorganic polyphosphate: A molecule of many functions. Ann. Rev. Biochem. 1999, 68, 89–125. [Google Scholar] [CrossRef]
- Ruiz, F.A.; Lea, C.R.; Oldfield, E.; Docampo, R. Human platelet dense granules contain polyphosphate and are similar to acidocalcisomes of bacteria and unicellular eukaryotes. J. Biol. Chem. 2004, 279, 44250–44257. [Google Scholar] [CrossRef] [Green Version]
- Morrissey, J.H.; Choi, S.H.; Smith, S.A. Polyphosphate: An ancient molecule that links platelets, coagulation, and inflammation. Blood 2012, 119, 5972–5979. [Google Scholar] [CrossRef] [Green Version]
- Bezold, D.; Dürr, T.; Singh, J.; Jessen, H.J. Cyclotriphosphate: A brief history, recent developments, and perspectives in synthesis. Chemistry 2020, 26, 2298–2308. [Google Scholar] [CrossRef] [Green Version]
- Jessen, H.J.; Dürr-Mayer, T.; Haas, T.M.; Ripp, A.; Cummins, C.C. Lost in condensation: Poly-, cyclo-, and ultraphosphates. Acc. Chem. Res. 2021, 54, 4036–4050. [Google Scholar] [CrossRef]
- Dürr-Mayer, T.; Qiu, D.; Eisenbeis, V.B.; Steck, N.; Häner, M.; Hofer, A.; Mayer, A.; Siegel, J.S.; Baldridge, K.K.; Jessen, H.J. The chemistry of branched condensed phosphates. Nat. Commun. 2021, 12, 5368. [Google Scholar] [CrossRef] [PubMed]
- Hoylaerts, M.F.; Manes, T.; Millán, J.L. Mammalian alkaline phosphatases are allosteric enzymes. J. Biol. Chem. 1997, 272, 22781–22787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engle, M.J.; Mahmood, A.; Alpers, D.H. Two rat intestinal alkaline phosphatase isoforms with different carboxyl-terminal peptides are both membrane-bound by a glycan phosphatidylinositol linkage. J. Biol. Chem. 1995, 270, 11935–11940. [Google Scholar] [CrossRef] [Green Version]
- Halling Linder, C.; Englund, U.H.; Narisawa, S.; Millán, J.L.; Magnusson, P. Isozyme profile and tissue-origin of alkaline phosphatases in mouse serum. Bone 2013, 53, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Eliakim, R.; Mahmood, A.; Alpers, D.H. Rat intestinal alkaline phosphatase secretion into lumen and serum is coordinately regulated. Biochim. Biophys. Acta 1991, 1091, 1–8. [Google Scholar] [CrossRef]
- De Broe, M.E.; Pollet, D.E. Multicenter evaluation of human placental alkaline phosphatase as a possible tumor-associated antigen in serum. Clin. Chem. 1988, 34, 1995–1999. [Google Scholar] [CrossRef]
- De Jong, L.; van der Kraan, I.; de Waal, A. The kinetics of the hydroxylation of procollagen by prolyl 4-hydroxylase. Proposal for a processive mechanism of binding of the dimeric hydroxylating enzyme in relation to the high kcat/Km ratio and a conformational requirement for hydroxylation of -X-Pro-Gly- sequences. Biochim. Biophys. Acta 1991, 1079, 103–111. [Google Scholar]
- Chaudhuri, G.; Chatterjee, S.; Venu-Babu, P.; Ramasamy, K.; Thilagaraj, W.R. Kinetic behaviour of calf intestinal alkaline phosphatase with pNPP. Indian J. Biochem. Biophys. 2013, 50, 64–71. [Google Scholar]
- Sun, Y.; Rahbani, J.F.; Jedrychowski, M.P.; Riley, C.L.; Vidoni, S.; Bogoslavski, D.; Hu, B.; Dumesic, P.A.; Zeng, X.; Wang, A.B.; et al. Mitochondrial TNAP controls thermogenesis by hydrolysis of phosphocreatine. Nature 2021, 593, 580–585. [Google Scholar] [CrossRef]
- Kazak, L.; Chouchani, E.T.; Jedrychowski, M.P.; Erickson, B.K.; Shinoda, K.; Cohen, P.; Vetrivelan, R.; Lu, G.Z.; Laznik-Bogoslavski, D.; Hasenfuss, S.C.; et al. A creatine-driven substrate cycle enhances energy expenditure and thermogenesis in beige fat. Cell 2015, 163, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Nam, H.K.; Crouch, S.; Hatch, N.E. Tissue nonspecific alkaline phosphatase function in bone and muscle progenitor cells: Control of mitochondrial respiration and ATP production. Int. J. Mol. Sci. 2021, 22, 1140. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.A.; Mutch, N.J.; Baskar, D.; Rohloff, P.; Docampo, R.; Morrissey, J.H. Polyphosphate modulates blood coagulation and fibrinolysis. Proc. Natl. Acad. Sci. USA 2006, 103, 903–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijeyewickrema, L.C.; Lameignere, E.; Hor, L.; Duncan, R.C.; Shiba, T.; Travers, R.J.; Kapopara, P.R.; Lei, V.; Smith, S.A.; Kim, H.; et al. Polyphosphate is a novel cofactor for regulation of complement by a serpin, C1 inhibitor. Blood 2016, 128, 1766–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, W.E.G.; Wang, X.H.; Diehl-Seifert, B.; Kropf, K.; Schloßmacher, U.; Lieberwirth, I.; Glasser, G.; Wiens, M.; Schröder, H.C. Inorganic polymeric phosphate/polyphosphate as an inducer of alkaline phosphatase and a modulator of intracellular Ca2+ level in osteoblasts (SaOS-2 cells) in vitro. Acta Biomater. 2011, 7, 2661–2671. [Google Scholar] [CrossRef]
- Leyhausen, G.; Lorenz, B.; Zhu, H.; Geurtsen, W.; Bohnensack, R.; Müller, W.E.G.; Schröder, H.C. Inorganic polyphosphate in human osteoblast-like cells. J. Bone Miner. Res. 1998, 13, 803–812. [Google Scholar] [CrossRef]
- Wang, X.H.; Schröder, H.C.; Wiens, M.; Ushijima, H.; Müller, W.E.G. Bio-silica and bio-polyphosphate: Applications in biomedicine (bone formation). Curr. Opin. Biotechnol. 2012, 23, 570–578. [Google Scholar] [CrossRef]
- Omelon, S.J.; Grynpas, M.D. Relationships between polyphosphate chemistry, biochemistry and apatite biomineralization. Chem. Rev. 2008, 108, 4694–4715. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Ackermann, M.; Wang, S.F.; Neufurth, M.; Muñoz-Espí, R.; Feng, Q.L.; Schröder, H.C.; Wang, X.H. Inorganic polyphosphate induces accelerated tube formation of HUVEC endothelial cells. Cell. Mol. Life Sci. 2018, 75, 21–32. [Google Scholar] [CrossRef]
- Wang, X.H.; Schloßmacher, U.; Schröder, H.C.; Müller, W.E.G. Biologically-induced transition of bio-silica sol to mesoscopic gelatinous flocs: A biomimetic approach to a controlled fabrication of bio-silica structures. Soft Matter 2013, 9, 654–664. [Google Scholar] [CrossRef]
- Wang, X.H.; Schröder, H.C.; Wang, K.; Kaandorp, J.A.; Müller, W.E.G. Genetic, biological and structural hierarchies during sponge spicule formation: From soft sol-gels to solid 3D silica composite structures. Soft Matter 2012, 8, 9501–9518. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Wang, X.H.; Burghard, Z.; Bill, J.; Krasko, A.; Boreiko, A.; Schloßmacher, U.; Schröder, H.C.; Wiens, M. Bio-sintering processes in hexactinellid sponges: Fusion of biosilica in giant basal spicules from Monorhaphis chuni. J. Struct. Biol. 2009, 168, 548–561. [Google Scholar] [CrossRef] [PubMed]
- Phosphoric Acid and Phosphates. In Kirk-Othmer Encyclopedia of Chemical Technology, 4th ed.; John Wiley & Sons: New York, NY, USA, 1991; Volume 18, pp. 669–718.
- Van Wazer, J.R. (Ed.) Phosphorus and Its Compounds: Chemistry; Interscience Publishers Inc.: New York, NY, USA, 1958; Volume 1. [Google Scholar]
- De Jager, H.; Heyns, A.M. Study of the hydrolysis of sodium polyphosphate in water using Raman spectroscopy. Appl. Spectrosc. 1998, 52, 808–814. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Tolba, E.; Schröder, H.C.; Wang, S.F.; Glaßer, G.; Muñoz-Espí, R.; Link, T.; Wang, X.H. A new polyphosphate calcium material with morphogenetic activity. Mater. Lett. 2015, 148, 163–166. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Tolba, E.; Ackermann, M.; Neufurth, M.; Wang, S.F.; Feng, Q.L.; Schröder, H.C.; Wang, X.H. Fabrication of amorphous strontium polyphosphate microparticles that induce mineralization of bone cells in vitro and in vivo. Acta Biomater. 2017, 50, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.G.; Schepler, H.; Tolba, E.; Wang, S.F.; Ackermann, M.; Muñoz-Espí, R.; Xiao, S.; Tan, R.; She, Z.; Neufurth, M.; et al. A physiologically active interpenetrating collagen network that supports growth and migration of epidermal keratinocytes: Zinc-polyP nanoparticles integrated into compressed collagen. J. Mater. Chem. B 2020, 8, 5892–5902. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhang, X. Receptor activator of nuclear factor-κB ligand (RANKL)/RANK/osteoprotegerin system in bone and other tissues (review). Mol. Med. Rep. 2015, 11, 3212–3218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.H.; Ackermann, M.; Wang, S.F.; Tolba, E.; Neufurth, M.; Feng, Q.L.; Schröder, H.C.; Müller, W.E.G. Amorphous polyphosphate/amorphous calcium carbonate implant material with enhanced bone healing efficacy in a critical-size-defect in rats. Biomed. Mater. 2016, 11, 035005. [Google Scholar] [CrossRef]
- Wang, X.H.; Ackermann, M.; Tolba, E.; Neufurth, M.; Wurm, F.; Feng, Q.L.; Wang, S.F.; Schröder, H.C.; Müller, W.E.G. Artificial cartilage bio-matrix formed of hyaluronic acid and Mg2+-polyphosphate. Europ. Cells Mater. 2016, 32, 271–283. [Google Scholar] [CrossRef]
- Malinauskas, T.; Jones, E.Y. Extracellular modulators of Wnt signalling. Curr. Opin. Struct. Biol. 2014, 29, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.F.; Wang, X.H.; Neufurth, M.; Tolba, E.; Schepler, H.; Xiao, S.; Schröder, H.C.; Müller, W.E.G. Biomimetic alginate/gelatin cross-linked hydrogels supplemented with polyphosphate for wound healing applications. Molecules 2020, 25, 5210. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Ackermann, M.; Al-Nawas, B.; Righesso, L.A.R.; Muñoz-Espí, R.; Tolba, E.; Neufurth, M.; Schröder, H.C.; Wang, X.H. Amplified morphogenetic and bone forming activity of amorphous versus crystalline calcium phosphate/polyphosphate. Acta Biomater. 2020, 118, 233–247. [Google Scholar] [CrossRef] [PubMed]
- Betts, F.; Posner, A.S. An X-ray radial distribution study of amorphous calcium phosphate. Mater. Res. Bull. 1974, 9, 353–360. [Google Scholar] [CrossRef]
- Kanzaki, N.; Treboux, G.; Onuma, K.; Tsutsumi, S.; Ito, A. Calcium phosphate clusters. Biomaterials 2001, 22, 2921–2929. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Wang, S.F.; Tolba, E.; Neufurth, M.; Ackermann, M.; Muñoz-Espí, R.; Lieberwirth, I.; Glasser, G.; Schröder, H.C.; Wang, X.H. Transformation of amorphous polyphosphate nanoparticles into coacervate complexes: An approach for the encapsulation of mesenchymal stem cells. Small 2018, 14, e1801170. [Google Scholar] [CrossRef]
- Franco, D.F.; De Oliveira Barud, H.G.; Barud, H.S.; Oliveira, O.B.; Meneguin, A.B.; de Oliveira, L.F.C.; Silva, M.A.P.; Ribeiro, S.J.L.; Nalin, M. A review on polyphosphate coacervates—Structural properties and bioapplications. J. Sol-Gel Sci. Technol. 2020, 94, 531–543. [Google Scholar] [CrossRef]
- Yewdall, N.A.; André, A.A.M.; Lu, T.; Spruijt, E. Coacervates as models of membrane-less organelles. Curr. Opin. Colloid Interface Sci. 2021, 52, 101416. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Schröder, H.C.; Feng, Q.L.; Schlossmacher, U.; Link, T.; Wang, X.H. Development of a morphogenetically active scaffold for three-dimensional growth of bone cells: Biosilica-alginate hydrogel for SaOS-2 cell cultivation. J. Tissue Eng. Regen. Med. 2015, 9, E39–E50. [Google Scholar] [CrossRef]
- Cicco, S.R.; Vona, D.; Leone, G.; Lo Presti, M.; Palumbo, F.; Altamura, E.; Ragni, R.; Farinola, G.M. From polydisperse diatomaceous earth to biosilica with specific morphologies by glucose gradient/dialysis: A natural material for cell growth. MRS Commun. 2017, 7, 214–220. [Google Scholar] [CrossRef]
- Cicco, S.R.; Vona, D.; Leone, G.; De Giglio, E.; Bonifacio, M.A.; Cometa, S.; Fiore, S.; Palumbo, F.; Ragni, R.; Farinola, G.M. In vivo functionalization of diatom biosilica with sodium alendronate as osteoactive material. Mater. Sci. Eng. C 2019, 104, 109897. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, S.; Pan, H.; Tang, R. Less is more: Silicate in the crystallization of hydroxyapatite in simulated body fluids. CrystEngComm 2016, 18, 379–383. [Google Scholar] [CrossRef]
- Kellermeier, M.; Melero-García, E.; Glaab, F.; Klein, R.; Drechsler, M.; Rachel, R.; García-Ruiz, J.M.; Kunz, W. Stabilization of amorphous calcium carbonate in inorganic silica-rich environments. J. Am. Chem. Soc. 2010, 132, 17859–17866. [Google Scholar] [CrossRef] [PubMed]
- Sahai, N. Modeling apatite nucleation in the human body and in the geochemical environment. Am. J. Sci. 2000, 305, 661–672. [Google Scholar] [CrossRef]
- Sahai, N.; Tossell, J.A. Molecular orbital study of apatite (Ca5(PO4)3OH) nucleation at silica bioceramic surfaces. J. Phys. Chem. B 2000, 104, 4321–4322. [Google Scholar] [CrossRef]
- Salih, E.; Flückiger, R. Complete topographical distribution of both the in vivo and in vitro phosphorylation sites of bone sialoprotein and their biological significance. J. Biol. Chem. 2004, 279, 19808–19815. [Google Scholar] [CrossRef] [Green Version]
- Müller, W.E.G.; Tolba, E.; Schröder, H.C.; Neufurth, M.; Wang, S.; Link, T.; Al-Nawas, B.; Wang, X.H. A new printable and durable N,O-carboxymethyl chitosan-Ca2+-polyphosphate complex with morphogenetic activity. J. Mat. Chem. B 2015, 3, 1722–1730. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Relkovic, D.; Ackermann, M.; Wang, S.F.; Neufurth, M.; Paravic-Radicevic, A.; Ushijima, H.; Schröder, H.C.; Wang, X.H. Enhancement of wound healing in normal and diabetic mice by topical application of amorphous polyphosphate—Superior effect of the host-guest composite material composed of collagen (host) and polyphosphate (guest). Polymers 2017, 9, 300. [Google Scholar] [CrossRef] [Green Version]
- Müller, W.E.G.; Tolba, E.; Feng, Q.L.; Schröder, H.C.; Markl, J.S.; Kokkinopoulou, M.; Wang, X.H. Amorphous Ca2+ polyphosphate nanoparticles regulate ATP level in bone-like SaOS-2 cells. J. Cell Sci. 2015, 128, 2202–2207. [Google Scholar] [CrossRef] [Green Version]
- Gibard, C.; Bhowmik, S.; Karki, M.; Kim, E.K.; Krishnamurthy, R. Phosphorylation, oligomerization and self-assembly in water under potential prebiotic conditions. Nat. Chem. 2018, 10, 212–217. [Google Scholar] [CrossRef]
- Britvin, S.N.; Murashko, M.N.; Vapnik, Y.; Vlasenko, N.S.; Krzhizhanovskaya, M.G.; Vereshchagin, O.S.; Bocharov, V.N.; Lozhkin, M.S. Cyclophosphates, a new class of native phosphorus compounds, and some insights into prebiotic phosphorylation on early Earth. Geology 2021, 49, 382–386. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell. Sci. 2010, 123 Pt 24, 4195–4200. [Google Scholar] [CrossRef] [Green Version]
- Theocharis, A.D.; Manou, D.; Karamanos, N.K. The extracellular matrix as a multitasking player in disease. FEBS J. 2019, 286, 2830–2869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagliabracci, V.S.; Engel, J.L.; Wen, J.; Wiley, S.E.; Worby, C.A.; Kinch, L.N.; Xiao, J.; Grishin, N.V.; Dixon, J.E. Secreted kinase phosphorylates extracellular proteins that regulate biomineralization. Science 2012, 336, 1150–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagliabracci, V.S.; Pinna, L.A.; Dixon, J.E. Secreted protein kinases. Trends Biochem. Sci. 2013, 38, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poon, S.; Easterbrook-Smith, S.B.; Rybchyn, M.S.; Carver, J.A.; Wilson, M.R. Clusterin is an ATP-independent chaperone with very broad substrate specificity that stabilizes stressed proteins in a folding-competent state. Biochemistry 2000, 39, 15953–15960. [Google Scholar] [CrossRef] [PubMed]
- Heppel, L.A.; Harkness, D.R.; Hilmoe, R.J. A study of the substrate specificity and other properties of the alkaline phosphatase of Escherichia coli. J. Biol. Chem. 1962, 237, 841–846. [Google Scholar] [CrossRef]
- Kohn, G.; Delvaux, D.; Lakaye, B.; Servais, A.C.; Scholer, G.; Fillet, M.; Elias, B.; Derochette, J.M.; Crommen, J.; Wins, P.; et al. High inorganic triphosphatase activities in bacteria identification of the enzymes involved. PLoS ONE 2012, 7, e43879. [Google Scholar] [CrossRef]
- Neufurth, M.; Wang, X.H.; Wang, S.F.; Steffen, R.; Ackermann, M.; Haep, N.D.; Schröder, H.C.; Müller, W.E.G. 3D printing of hybrid (bio)materials for bone tissue engineering: Calcium-polyphosphate microparticles encapsulated by polycaprolactone. Acta Biomater. 2017, 64, 377–388. [Google Scholar] [CrossRef]
- Neufurth, M.; Wang, X.H.; Schröder, H.C.; Feng, Q.L.; Diehl-Seifert, B.; Ziebart, T.; Steffen, R.; Wang, S.F.; Müller, W.E.G. Engineering a morphogenetically active hydrogel for bioprinting of bioartificial tissue derived from human osteoblast-like SaOS-2 cells. Biomaterials 2014, 35, 8810–8819. [Google Scholar] [CrossRef]
- Neufurth, M.; Wang, S.F.; Schröder, H.C.; Al-Nawas, B.; Wang, X.H.; Müller, W.E.G. 3D bioprinting of tissue units with mesenchymal stem cells; retaining their proliferative and differentiating potential, in polyphosphate-containing bio-ink. Biofabrication 2021, in press. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Tolba, E.; Schröder, H.C.; Diehl-Seifert, B.; Link, T.; Wang, X.H. Biosilica-loaded poly(ϵ-caprolactone) nanofibers mats provide a morphogenetically active surface scaffold for the growth and mineralization of the osteoclast-related SaOS-2 cells. Biotechnol. J. 2014, 9, 1312–1321. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Tolba, E.; Dorweiler, B.; Schröder, H.C.; Diehl-Seifert, B.; Wang, X.H. Electrospun bioactive mats enriched with Ca-polyphosphate/retinol nanospheres as potential wound dressing. Biochem. Biophys. Rep. 2015, 3, 150–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.F.; Wang, X.H.; Draenert, F.G.; Albert, O.; Schröder, H.C.; Mailänder, V.; Mitov, G.; Müller, W.E.G. Bioactive and biodegradable silica biomaterial for bone regeneration. Bone 2014, 67, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Wang, S.; He, F.; Tolba, E.; Schröder, H.C.; Diehl-seifert, B.; Müller, W.E.G. Polyphosphate as a bioactive and biodegradable implant material: Induction of bone regeneration in rats. Adv. Eng. Mater. 2016, 18, 1406–1417. [Google Scholar] [CrossRef]
- Wiens, M.; Elkhooly, T.A.; Schröder, H.C.; Mohamed, T.H.; Müller, W.E.G. Characterization and osteogenic activity of a silicatein/biosilica-coated chitosan-graft-polycaprolactone. Acta Biomater. 2014, 10, 4456–4464. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.G.; Neufurth, M.; Ackermann, M.; Tolba, E.; Wang, S.F.; Feng, Q.L.; Schröder, H.C.; Wang, X.H. Fabrication of a new physiological macroporous hybrid biomaterial/bioscaffold material based on polyphosphate and collagen by freeze-extraction. J. Mat. Chem. B 2017, 5, 3823–3835. [Google Scholar] [CrossRef]
- Tolba, E.; Wang, X.H.; Ackermann, M.; Neufurth, M.; Muñoz-Espí, R.; Schröder, H.C.; Müller, W.E.G. In-situ polyphosphate nanoparticle formation in hybrid poly(vinyl alcohol,/karaya gum-hydrogels: A porous scaffold inducing infiltration of mesenchymal stem cells. Adv. Sci. 2018, 2018, 1801452. [Google Scholar] [CrossRef]
- Van Loocke, M.; Lyons, C.G.; Simms, C.K. Viscoelastic properties of passive skeletal muscle in compression: Stress-relaxation behaviour and constitutive modelling. J. Biomech. 2008, 41, 1555–1566. [Google Scholar] [CrossRef]
- Kocen, R.; Gasik, M.; Gantar, A.; Novak, S. Viscoelastic behaviour of hydrogel-based composites for tissue engineering under mechanical load. Biomed. Mater. 2017, 12, 025004. [Google Scholar] [CrossRef]
- Bjarnsholt, T.; Kirketerp-Møller, K.; Jensen, P.Ø.; Madsen, K.G.; Phipps, R.; Krogfelt, K.; Høiby, N.; Givskov, M. Why chronic wounds will not heal: A novel hypothesis. Wound Repair Regen. 2008, 16, 2–10. [Google Scholar] [CrossRef]
- Schepler, H.; Neufurth, M.; Wang, S.F.; She, Z.; Schröder, H.C.; Wang, X.H.; Müller, W.E.G. Acceleration of chronic wound healing by bio-inorganic polyphosphate: In vitro studies and first clinical applications. Theranostics 2022, 12, 18–34. [Google Scholar] [CrossRef]
- Rice, J.J.; Martino, M.M.; De Laporte, L.; Tortelli, F.; Briquez, P.S.; Hubbell, J.A. Engineering the regenerative microenvironment with biomaterials. Adv. Health Mater. 2013, 2, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Walsh, K.; Hoff, B.L.; Camci-Unal, G. Mineralization of biomaterials for bone tissue engineering. Bioengineering 2020, 7, 132. [Google Scholar] [CrossRef] [PubMed]
- Alkaabi, S.A.; Kalla, D.S.N.; Alsabri, G.A.; Fauzi, A.; Jansen, N.; Tajrin, A.; Nurrahma, R.; Müller, W.E.G.; Schröder, H.C.; Xiaohong, X.H.; et al. Safety and feasibility study of using polyphosphate (PolyP) in alveolar cleft repair: A pilot study. Pilot Feasibility Stud. 2021, 7, 199. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schröder, H.C.; Wang, X.; Neufurth, M.; Wang, S.; Tan, R.; Müller, W.E.G. Inorganic Polymeric Materials for Injured Tissue Repair: Biocatalytic Formation and Exploitation. Biomedicines 2022, 10, 658. https://doi.org/10.3390/biomedicines10030658
Schröder HC, Wang X, Neufurth M, Wang S, Tan R, Müller WEG. Inorganic Polymeric Materials for Injured Tissue Repair: Biocatalytic Formation and Exploitation. Biomedicines. 2022; 10(3):658. https://doi.org/10.3390/biomedicines10030658
Chicago/Turabian StyleSchröder, Heinz C., Xiaohong Wang, Meik Neufurth, Shunfeng Wang, Rongwei Tan, and Werner E. G. Müller. 2022. "Inorganic Polymeric Materials for Injured Tissue Repair: Biocatalytic Formation and Exploitation" Biomedicines 10, no. 3: 658. https://doi.org/10.3390/biomedicines10030658