
A Unique Immune-Related Gene Signature Represents Advanced Liver Fibrosis and Reveals Potential Therapeutic Targets

 , ,
, ,
Abstract
:1. Introduction
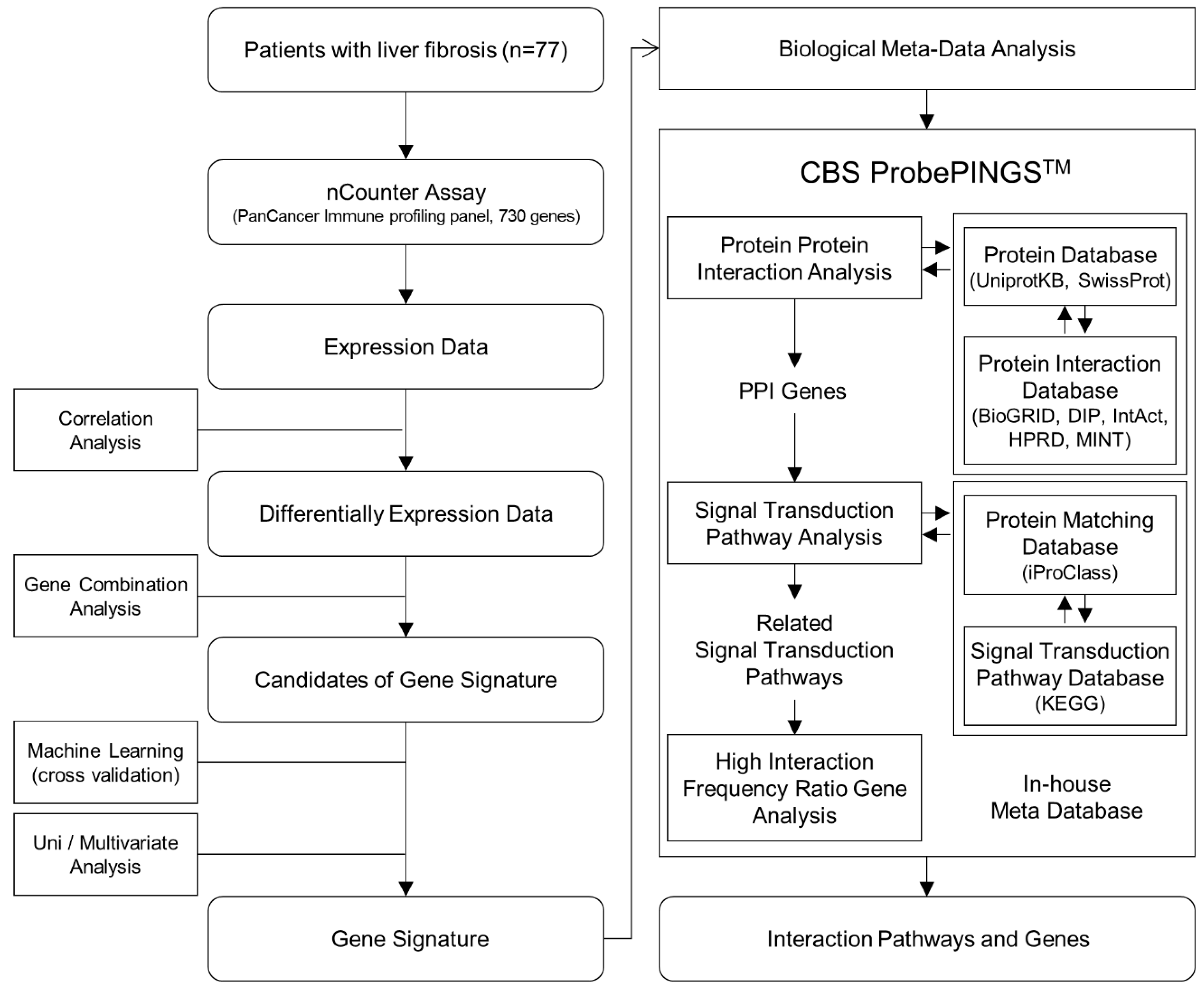
2. Methods
2.1. Patients and Tissue Samples
2.2. RNA Extraction
2.3. Gene Expression Assay
2.4. Gene Combination Analysis
2.5. Cross-Validation as Pre-Validation of Candidate Gene Signatures
2.6. Biological Meta-Analysis
2.7. Statistical Analysis
3. Results
3.1. Patient Characteristics
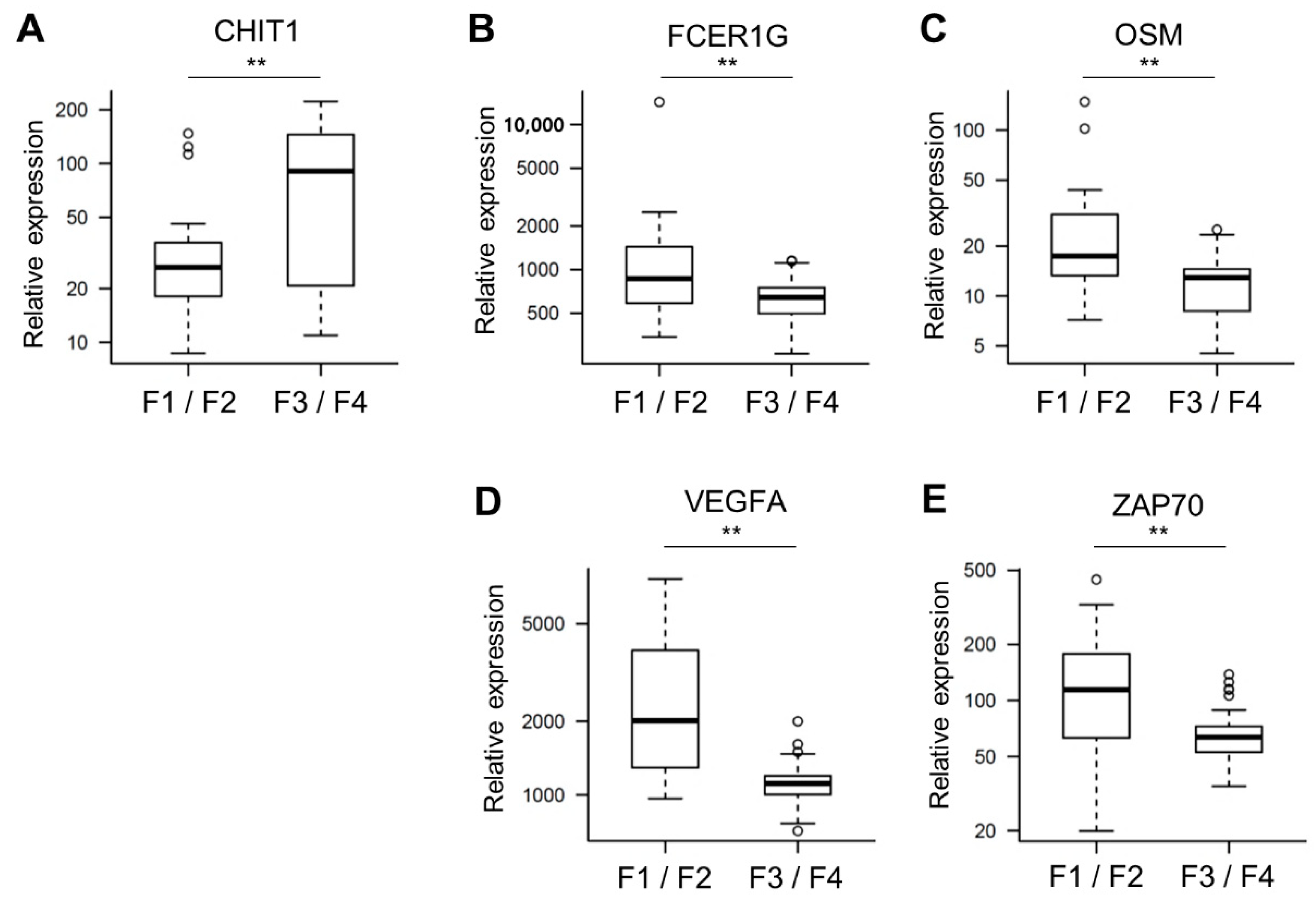
3.2. Expression Analysis-Derived Gene Signatures
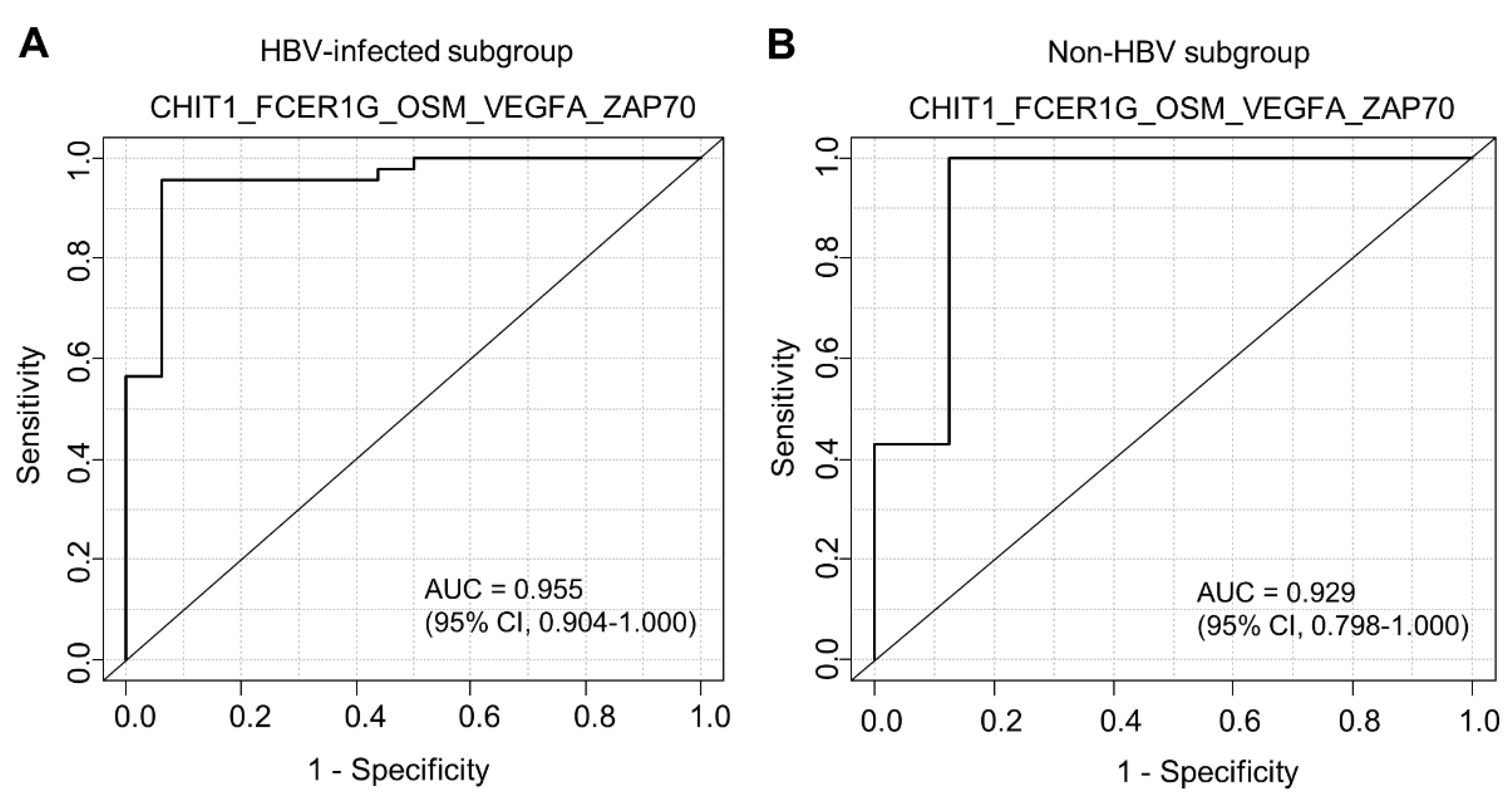
3.3. Factors Associated with Pathological High-Grade Fibrosis
3.4. Pathways and Genes Significantly Associated with the Gene Signature
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- Lesmana, C.R.A.; Raharjo, M.; Gani, R.A. Managing liver cirrhotic complications: Overview of esophageal and gastric varices. Clin. Mol. Hepatol. 2020, 26, 444–460. [Google Scholar] [CrossRef] [PubMed]
- Odagiri, N.; Matsubara, T.; Sato-Matsubara, M.; Fujii, H.; Enomoto, M.; Kawada, N. Anti-fibrotic treatments for chronic liver diseases: The present and the future. Clin. Mol. Hepatol. 2020, 27, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Hoshida, Y.; Villanueva, A.; Sangiovanni, A.; Sole, M.; Hur, C.; Andersson, K.L.; Chung, R.T.; Gould, J.; Kojima, K.; Gupta, S.; et al. Prognostic gene expression signature for patients with hepatitis C-related early-stage cirrhosis. Gastroenterology 2013, 144, 1024–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goytain, A.; Ng, T. NanoString nCounter Technology: High-Throughput RNA Validation. Methods Mol. Biol. 2020, 2079, 125–139. [Google Scholar] [CrossRef]
- Park, I.J.; Yu, Y.S.; Mustafa, B.; Park, J.Y.; Seo, Y.B.; Kim, G.D.; Kim, J.; Kim, C.M.; Noh, H.D.; Hong, S.M.; et al. A Nine-Gene Signature for Predicting the Response to Preoperative Chemoradiotherapy in Patients with Locally Advanced Rectal Cancer. Cancers 2020, 12, 800. [Google Scholar] [CrossRef] [Green Version]
- Sung, P.S. Crosstalk between tumor-associated macrophages and neighboring cells in hepatocellular carcinoma. Clin. Mol. Hepatol. 2021. [Google Scholar] [CrossRef]
- Gomez Perdiguero, E.; Klapproth, K.; Schulz, C.; Busch, K.; Azzoni, E.; Crozet, L.; Garner, H.; Trouillet, C.; de Bruijn, M.F.; Geissmann, F.; et al. Tissue-resident macrophages originate from yolk-sac-derived erythro-myeloid progenitors. Nature 2015, 518, 547–551. [Google Scholar] [CrossRef]
- Tacke, F.; Zimmermann, H.W. Macrophage heterogeneity in liver injury and fibrosis. J. Hepatol. 2014, 60, 1090–1096. [Google Scholar] [CrossRef] [Green Version]
- Carpino, G.; Del Ben, M.; Pastori, D.; Carnevale, R.; Baratta, F.; Overi, D.; Francis, H.; Cardinale, V.; Onori, P.; Safarikia, S.; et al. Increased Liver Localization of Lipopolysaccharides in Human and Experimental NAFLD. Hepatology 2020, 72, 470–485. [Google Scholar] [CrossRef]
- Ruf, B.; Heinrich, B.; Greten, T.F. Immunobiology and immunotherapy of HCC: Spotlight on innate and innate-like immune cells. Cell Mol. Immunol. 2021, 18, 112–127. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, P.; Dobie, R.; Wilson-Kanamori, J.R.; Dora, E.F.; Henderson, B.E.P.; Luu, N.T.; Portman, J.R.; Matchett, K.P.; Brice, M.; Marwick, J.A.; et al. Resolving the fibrotic niche of human liver cirrhosis at single-cell level. Nature 2019, 575, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Hong, S.J.; Park, J.Y.; Park, J.H.; Yu, Y.S.; Park, S.Y.; Lim, E.K.; Choi, K.Y.; Lee, E.K.; Paik, S.S.; et al. Epithelial-mesenchymal transition gene signature to predict clinical outcome of hepatocellular carcinoma. Cancer Sci. 2010, 101, 1521–1528. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, A.J.; Van Natta, M.L.; Clark, J.; Neuschwander-Tetri, B.A.; Diehl, A.; Dasarathy, S.; Loomba, R.; Chalasani, N.; Kowdley, K.; Hameed, B.; et al. Prospective Study of Outcomes in Adults with Nonalcoholic Fatty Liver Disease. N. Engl. J. Med. 2021, 385, 1559–1569. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Lambrecht, J.; Ju, C.; Tacke, F. Hepatic macrophages in liver homeostasis and diseases-diversity, plasticity and therapeutic opportunities. Cell Mol. Immunol. 2021, 18, 45–56. [Google Scholar] [CrossRef]
- Bernsmeier, C.; van der Merwe, S.; Perianin, A. Innate immune cells in cirrhosis. J. Hepatol. 2020, 73, 186–201. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Arndtz, K.; Webb, G.; Corrigan, M.; Akiror, S.; Liaskou, E.; Woodward, P.; Adams, D.H.; Weston, C.J.; Hirschfield, G.M. Intrahepatic macrophage populations in the pathophysiology of primary sclerosing cholangitis. JHEP Rep. 2019, 1, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Cheng, D.; Chai, J.; Wang, H.; Fu, L.; Peng, S.; Ni, X. Hepatic macrophages: Key players in the development and progression of liver fibrosis. Liver Int. 2021, 41, 2279–2294. [Google Scholar] [CrossRef]
- Jophlin, L.L.; Cao, S.; Shah, V.H. The Transcriptome of Hepatic Fibrosis Revealed by Single-Cell RNA Sequencing. Hepatology 2020, 71, 1865–1867. [Google Scholar] [CrossRef]
- Wan, J.; Weiss, E.; Ben Mkaddem, S.; Mabire, M.; Choinier, P.M.; Picq, O.; Thibault-Sogorb, T.; Hegde, P.; Pishvaie, D.; Bens, M.; et al. LC3-associated phagocytosis protects against inflammation and liver fibrosis via immunoreceptor inhibitory signaling. Sci. Transl. Med. 2020, 12, eaaw8523. [Google Scholar] [CrossRef]
- Morse, C.; Tabib, T.; Sembrat, J.; Buschur, K.L.; Bittar, H.T.; Valenzi, E.; Jiang, Y.; Kass, D.J.; Gibson, K.; Chen, W.; et al. Proliferating SPP1/MERTK-expressing macrophages in idiopathic pulmonary fibrosis. Eur. Respir. J. 2019, 54. [Google Scholar] [CrossRef]
- Kanneganti, M.; Kamba, A.; Mizoguchi, E. Role of chitotriosidase (chitinase 1) under normal and disease conditions. J. Epithel. Biol. Pharmacol. 2012, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, L.; Di Rosa, M.; Zambito, A.M.; dell’Ombra, N.; Di Marco, R.; Malaguarnera, M. Potential role of chitotriosidase gene in nonalcoholic fatty liver disease evolution. Am. J. Gastroenterol. 2006, 101, 2060–2069. [Google Scholar] [CrossRef]
- Logue, E.C.; Neff, C.P.; Mack, D.G.; Martin, A.K.; Fiorillo, S.; Lavelle, J.; Vandivier, R.W.; Campbell, T.B.; Palmer, B.E.; Fontenot, A.P. Upregulation of Chitinase 1 in Alveolar Macrophages of HIV-Infected Smokers. J. Immunol. 2019, 202, 1363–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bournazos, S.; Wang, T.T.; Ravetch, J.V. The Role and Function of Fcgamma Receptors on Myeloid Cells. Microbiol. Spectr. 2016, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Vallejo-Diaz, J.; Chagoyen, M.; Olazabal-Moran, M.; Gonzalez-Garcia, A.; Carrera, A.C. The Opposing Roles of PIK3R1/p85alpha and PIK3R2/p85beta in Cancer. Trends Cancer 2019, 5, 233–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deau, M.C.; Heurtier, L.; Frange, P.; Suarez, F.; Bole-Feysot, C.; Nitschke, P.; Cavazzana, M.; Picard, C.; Durandy, A.; Fischer, A.; et al. A human immunodeficiency caused by mutations in the PIK3R1 gene. J. Clin. Investig. 2014, 124, 3923–3928. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.W.; Kim, S.M.; Hur, W.; Kang, B.Y.; Lee, H.L.; Nam, H.; Yoo, S.H.; Sung, P.S.; Kwon, J.H.; Jang, J.W.; et al. Tenofovir disoproxil fumarate directly ameliorates liver fibrosis by inducing hepatic stellate cell apoptosis via downregulation of PI3K/Akt/mTOR signaling pathway. PLoS ONE 2021, 16, e0261067. [Google Scholar] [CrossRef]
- Zhao, J.; Qi, Y.F.; Yu, Y.R. STAT3: A key regulator in liver fibrosis. Ann. Hepatol. 2021, 21, 100224. [Google Scholar] [CrossRef] [PubMed]
- Ozturk Akcora, B.; Vassilios Gabriel, A.; Ortiz-Perez, A.; Bansal, R. Pharmacological inhibition of STAT3 pathway ameliorates acute liver injury in vivo via inactivation of inflammatory macrophages and hepatic stellate cells. FASEB Bioadv. 2020, 2, 77–89. [Google Scholar] [CrossRef]
- Bharadwaj, U.; Kasembeli, M.M.; Robinson, P.; Tweardy, D.J. Targeting Janus Kinases and Signal Transducer and Activator of Transcription 3 to Treat Inflammation, Fibrosis, and Cancer: Rationale, Progress, and Caution. Pharmacol. Rev. 2020, 72, 486–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newsome, P.N.; Buchholtz, K.; Cusi, K.; Linder, M.; Okanoue, T.; Ratziu, V.; Sanyal, A.J.; Sejling, A.S.; Harrison, S.A.; Investigators, N.N. A Placebo-Controlled Trial of Subcutaneous Semaglutide in Nonalcoholic Steatohepatitis. N. Engl. J. Med. 2021, 384, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, M.J.; Gaunt, P.; Aithal, G.P.; Barton, D.; Hull, D.; Parker, R.; Hazlehurst, J.M.; Guo, K.; LEAN trial team; Abouda, G.; et al. Liraglutide safety and efficacy in patients with non-alcoholic steatohepatitis (LEAN): A multicentre, double-blind, randomised, placebo-controlled phase 2 study. Lancet 2016, 387, 679–690. [Google Scholar] [CrossRef] [Green Version]
- Ipsen, D.H.; Rolin, B.; Rakipovski, G.; Skovsted, G.F.; Madsen, A.; Kolstrup, S.; Schou-Pedersen, A.M.; Skat-Rordam, J.; Lykkesfeldt, J.; Tveden-Nyborg, P. Liraglutide Decreases Hepatic Inflammation and Injury in Advanced Lean Non-Alcoholic Steatohepatitis. Basic Clin. Pharmacol. Toxicol. 2018, 123, 704–713. [Google Scholar] [CrossRef]
- Li, Z.; Feng, P.P.; Zhao, Z.B.; Zhu, W.; Gong, J.P.; Du, H.M. Liraglutide protects against inflammatory stress in non-alcoholic fatty liver by modulating Kupffer cells M2 polarization via cAMP-PKA-STAT3 signaling pathway. Biochem. Biophys. Res. Commun. 2019, 510, 20–26. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fibrosis Stage 1–2 (N = 24) | Fibrosis Stage 3–4 (N = 53) | p Value | |||
|---|---|---|---|---|---|
| No. | % | No. | % | ||
| Sex | 0.5556 ^ | ||||
| Male | 18 | 75.0 | 43 | 81.1 | |
| Female | 6 | 25.0 | 10 | 18.9 | |
| Mean age (±SD) | 53.3 (±12.3) | 51.0 (±7.9) | 0.4474 * | ||
| Hepatitis B | 0.0606 ^ | ||||
| No | 8 | 33.3 | 7 | 13.2 | |
| Yes | 16 | 66.7 | 46 | 86.8 | |
| Hepatitis C | 1.0000 ^ | ||||
| No | 24 | 100.0 | 51 | 96.2 | |
| Yes | 0 | 0.0 | 2 | 3.8 | |
| Diabetes | 0.4761 # | ||||
| No | 17 | 70.8 | 43 | 81.1 | |
| Yes | 7 | 29.2 | 10 | 18.9 | |
| BMI | 0.2546 # | ||||
| ≤25 kg/m2 | 18 | 75.0 | 31 | 58.5 | |
| >25 kg/m2 | 6 | 25.0 | 22 | 41.5 | |
| ALT | 0.7843 # | ||||
| <31 (F), <41 (M) IU/L | 16 | 66.7 | 32 | 60.4 | |
| ≥31 (F), ≥41 (M) IU/L | 8 | 33.3 | 21 | 39.6 | |
| AST | 0.2101 # | ||||
| <31 (F), <37 (M) IU/L | 10 | 41.7 | 13 | 24.5 | |
| ≥31 (F), ≥37 (M) IU/L | 14 | 58.3 | 40 | 75.5 | |
| Platelets | 0.0144 # | ||||
| <150 × 109/L | 4 | 16.7 | 26 | 49.1 | |
| ≥150 × 109/L | 20 | 83.3 | 27 | 50.9 | |
| Antiviral treatment | 0.0143 ^ | ||||
| No | 24 | 100.0 | 42 | 79.2 | |
| Yes | 0 | 0.0 | 11 | 20.8 | |
| Fibrosis | <0.0001 ^ | ||||
| Stage 1 | 12 | 50.0 | 0 | 0.0 | |
| Stage 2 | 12 | 50.0 | 0 | 0.0 | |
| Stage 3 | 0 | 0.0 | 25 | 47.2 | |
| Stage 4 | 0 | 0.0 | 28 | 52.8 | |
| Etiology | 0.0543 ^ | ||||
| HBV | 16 | 66.6 | 46 | 86.8 | |
| HCV | 0 | 0.0 | 2 | 3.8 | |
| Alcohol | 4 | 16.7 | 3 | 5.6 | |
| Others | 4 | 16.7 | 2 | 3.8 | |
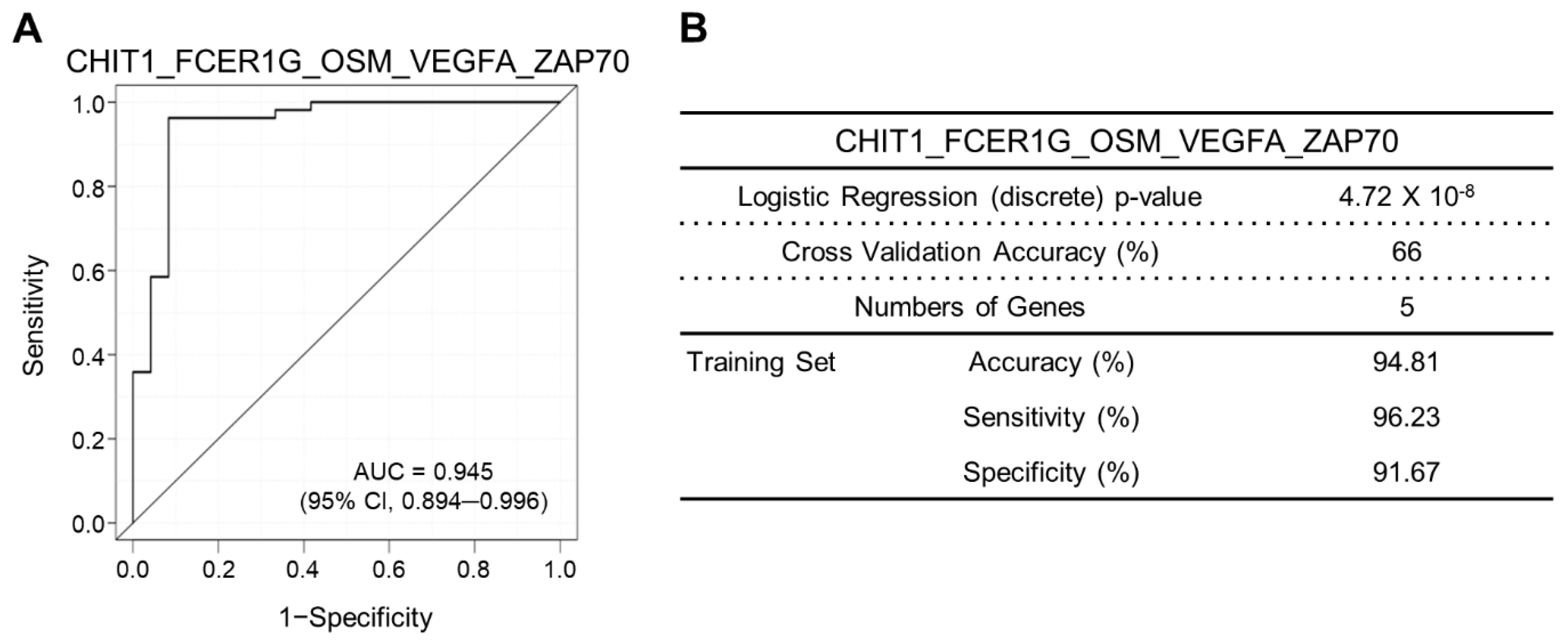
| Rank | Gene Signature | No. of Gene | Logistic Regression Continuous p-Value | ROC AUC | Threshold | Sensitivity | Specificity | Accuracy | PPV | NPV |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | CHIT1_FCER1G_OSM_VEGFA_ZAP70 | 5 | 3.45 × 10−4 | 0.945 | −11.152614 | 96.23 | 91.67 | 94.81 | 96.23 | 91.67 |
| 2 | FCER1G_LTB_MME_OSM_VEGFA | 5 | 9.21 × 10−4 | 0.954 | −11.766822 | 94.34 | 91.67 | 93.51 | 96.15 | 88.00 |
| 3 | CD1B_CHIT1_FCER1G_OSM_VEGFA | 5 | 6.44 × 10−4 | 0.951 | −9.544123 | 94.34 | 91.67 | 93.51 | 96.15 | 88.00 |
| 4 | HLADRB3_OSM_VEGFA_ZAP70 | 4 | 4.05 × 10−4 | 0.951 | −11.382897 | 94.34 | 91.67 | 93.51 | 96.15 | 88.00 |
| 5 | CHIT1_FCER1G_MAF_OSM_VEGFA | 5 | 2.08 × 10−4 | 0.950 | −11.392033 | 94.34 | 91.67 | 93.51 | 96.15 | 88.00 |
| 6 | CHIT1_OSM_VEGFA_ZAP70 | 4 | 1.01 × 10−3 | 0.950 | −8.781110 | 94.34 | 91.67 | 93.51 | 96.15 | 88.00 |
| 7 | CD1B_OSM_TAPBP_VEGFA_ZAP70 | 5 | 7.31 × 10−4 | 0.943 | −13.930554 | 94.34 | 91.67 | 93.51 | 96.15 | 88.00 |
| 8 | CCL25_CHIT1_OSM_VEGFA_ZAP70 | 5 | 7.92 × 10−4 | 0.943 | −9.519937 | 94.34 | 91.67 | 93.51 | 96.15 | 88.00 |
| 9 | CLEC4C_OSM_TAPBP_VEGFA_ZAP70 | 5 | 5.52 × 10−4 | 0.940 | −13.622690 | 94.34 | 91.67 | 93.51 | 96.15 | 88.00 |
| Univariable Logistic Regression | ||||||
|---|---|---|---|---|---|---|
| Variable | n | Coefficient | Odds Ratio (95% CI) | se (Coefficient) | z | p-Value |
| Gene signature (Low vs. High) | 77 | 5.6366 | 280.50 (37.11–2120.26) | 1.0320 | 5.462 | 4.72 × 10−8 |
| Age (≤55 years vs. >55 years) | 77 | −0.2364 | 0.79 (0.28–2.23) | 0.5296 | −0.446 | 0.655 |
| Sex (Male vs. Female) | 77 | −0.3600 | 0.70 (0.22–2.21) | 0.5878 | −0.612 | 0.540 |
| HBV (absent vs. present) | 77 | 1.1896 | 3.29 (1.03–10.51) | 0.5934 | 2.005 | 4.50 × 10−2 |
| BMI (≤25 kg/m2 vs. >25 kg/m2) | 77 | 0.7557 | 2.13 (0.73–6.23) | 0.5477 | 1.380 | 0.168 |
| Diabetes (absent vs. present) | 77 | −0.5713 | 0.56 (0.18–1.73) | 0.5700 | −1.002 | 0.316 |
| ALT (<31(F), <41(M) IU/L vs. ≥31(F), ≥41(M) IU/L) | 77 | 0.2719 | 1.31 (0.48–3.61) | 0.5161 | 0.527 | 0.598 |
| AST (<31(F), <37(M) IU/L vs. ≥31(F), ≥37(M) IU/L) | 77 | 0.7875 | 2.20 (0.79–6.12) | 0.5228 | 1.506 | 0.132 |
| Albumin (<4.0 g/dL vs. ≥4.0 g/dL) | 77 | 0.3314 | 1.39 (0.49–3.96) | 0.5335 | 0.621 | 0.534 |
| Platelets (<150 × 109/L vs. ≥150 × 109/L) | 77 | −1.5717 | 0.21 (0.06–0.69) | 0.6128 | −2.565 | 1.03 × 10−2 |
| Antiviral treatment (absent vs. present) | 77 | 18.0065 | 66085024.42 (0.00–Inf) | 1966.6495 | 0.009 | 0.993 |
| Simple steatosis (absent vs. present) | 77 | −035477 | 0.58 (0.16–2.05) | 0.6459 | −0.848 | 0.396 |
| Multivariable logistic regression | ||||||
| Variable | n | coefficient | Odds ratio (95% CI) | se (coefficient) | z | p-value |
| Gene signature (low vs. high) | 77 | 6.2559 | 521.08 (31.21–8700.42) | 1.4364 | 4.355 | 1.33 × 10−5 |
| HBV (absent vs. present) | 77 | 2.1565 | 8.64 (0.59-126.12) | 1.3678 | 1.577 | 0.115 |
| Platelets (<150 × 109/L vs. ≥150 × 109/L) | 77 | 0.5786 | 1.78 (0.13–24.46) | 1.3359 | 0.433 | 0.665 |
| High Interaction Pathways | High Interaction Genes |
|---|---|
| Kaposi sarcoma-associated herpesvirus infection Human papillomavirus infection Epstein–Barr virus infection | PIK3R1 STAT3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sung, P.-S.; Kim, C.-M.; Cha, J.-H.; Park, J.-Y.; Yu, Y.-S.; Wang, H.-J.; Kim, J.-K.; Bae, S.-H. A Unique Immune-Related Gene Signature Represents Advanced Liver Fibrosis and Reveals Potential Therapeutic Targets. Biomedicines 2022, 10, 180. https://doi.org/10.3390/biomedicines10010180
Sung P-S, Kim C-M, Cha J-H, Park J-Y, Yu Y-S, Wang H-J, Kim J-K, Bae S-H. A Unique Immune-Related Gene Signature Represents Advanced Liver Fibrosis and Reveals Potential Therapeutic Targets. Biomedicines. 2022; 10(1):180. https://doi.org/10.3390/biomedicines10010180
Chicago/Turabian StyleSung, Pil-Soo, Chang-Min Kim, Jung-Hoon Cha, Jin-Young Park, Yun-Suk Yu, Hee-Jung Wang, Jin-Kyeoung Kim, and Si-Hyun Bae. 2022. "A Unique Immune-Related Gene Signature Represents Advanced Liver Fibrosis and Reveals Potential Therapeutic Targets" Biomedicines 10, no. 1: 180. https://doi.org/10.3390/biomedicines10010180