
Potential Protective Effect of Vitamin C on Qunalphos-Induced Cardiac Toxicity: Histological and Tissue Biomarker Assay

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Design
2.3. Specimen’s Processing and Staining for Light Microscopy (LM)
2.4. Tissue Preparation for Transmission Electron Microscopy (TEM)
2.5. Parameters Processing and Estimation
2.6. The Antioxidant Situation of Heart Homogenate
2.7. TNF-α and Il-6 Levels in the Blood
2.8. Statistical Analysis
3. Results
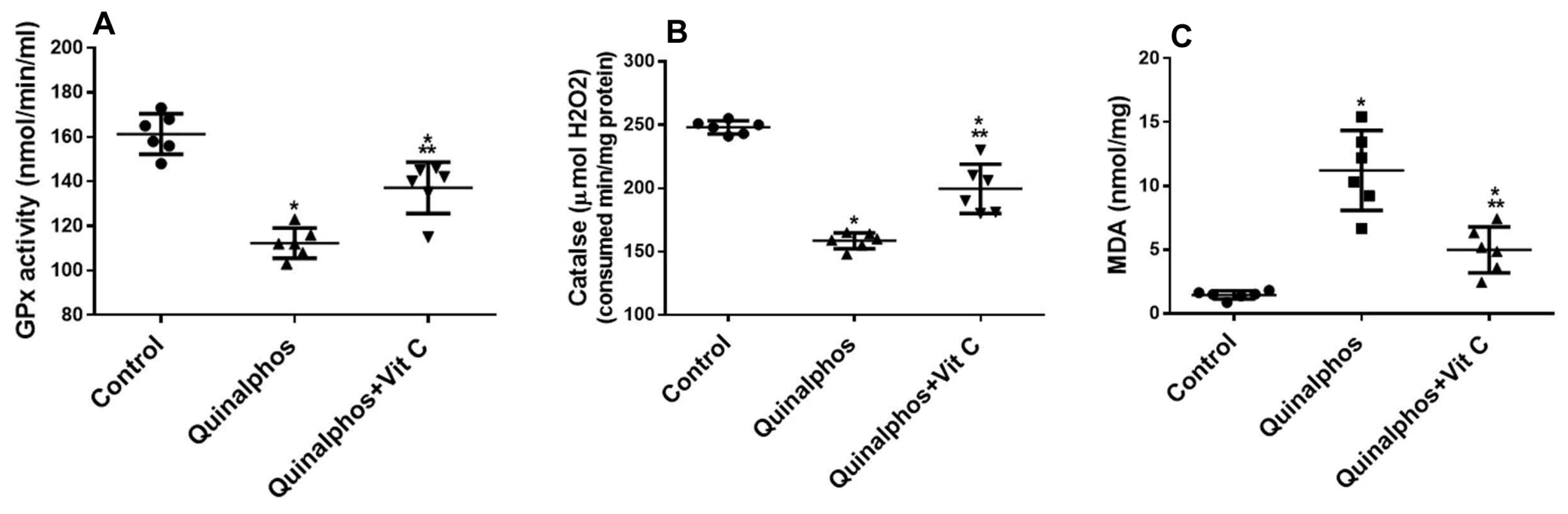
3.1. Biochemical and Statistical Analysis
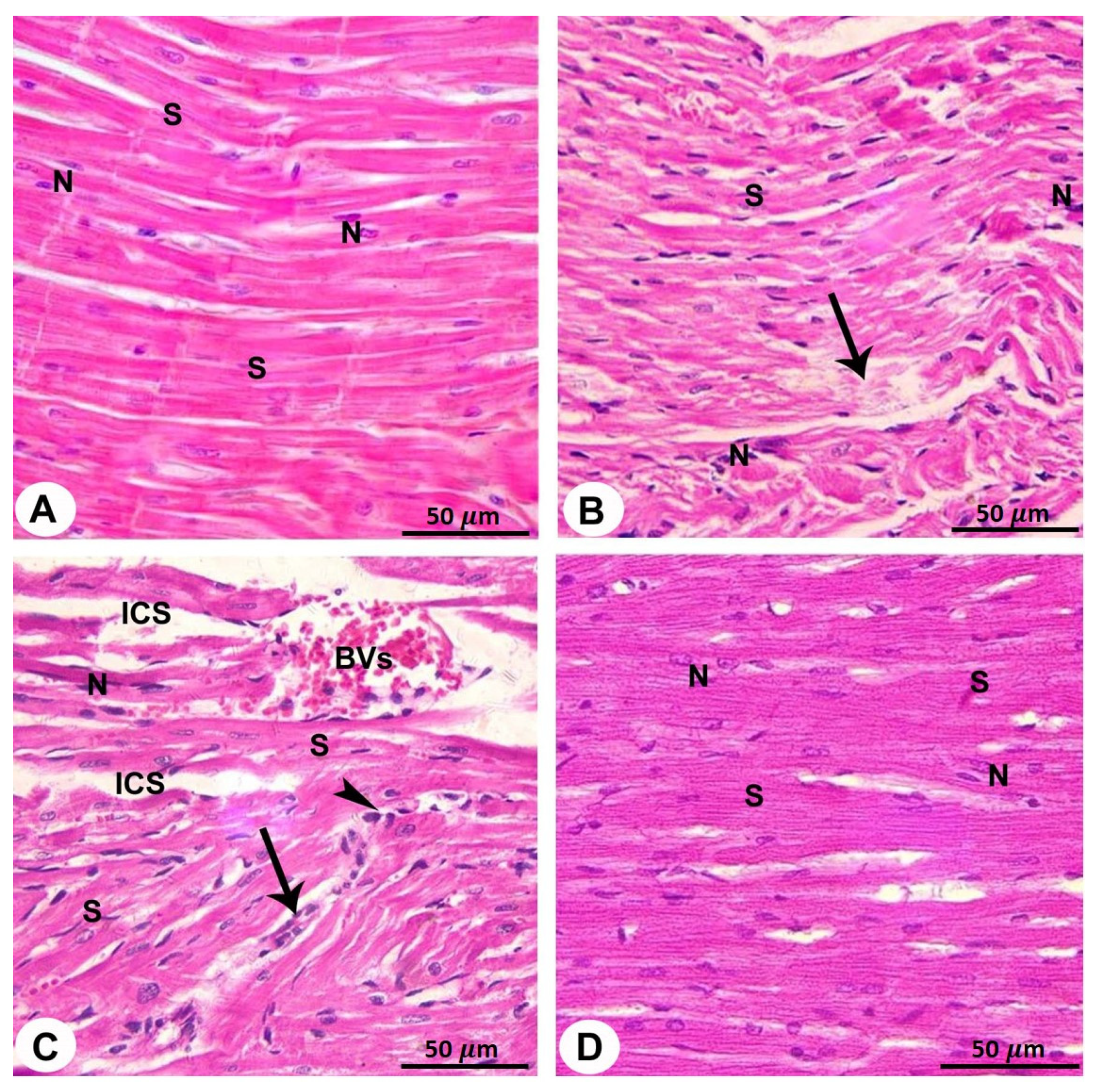
3.2. Light Microscopy
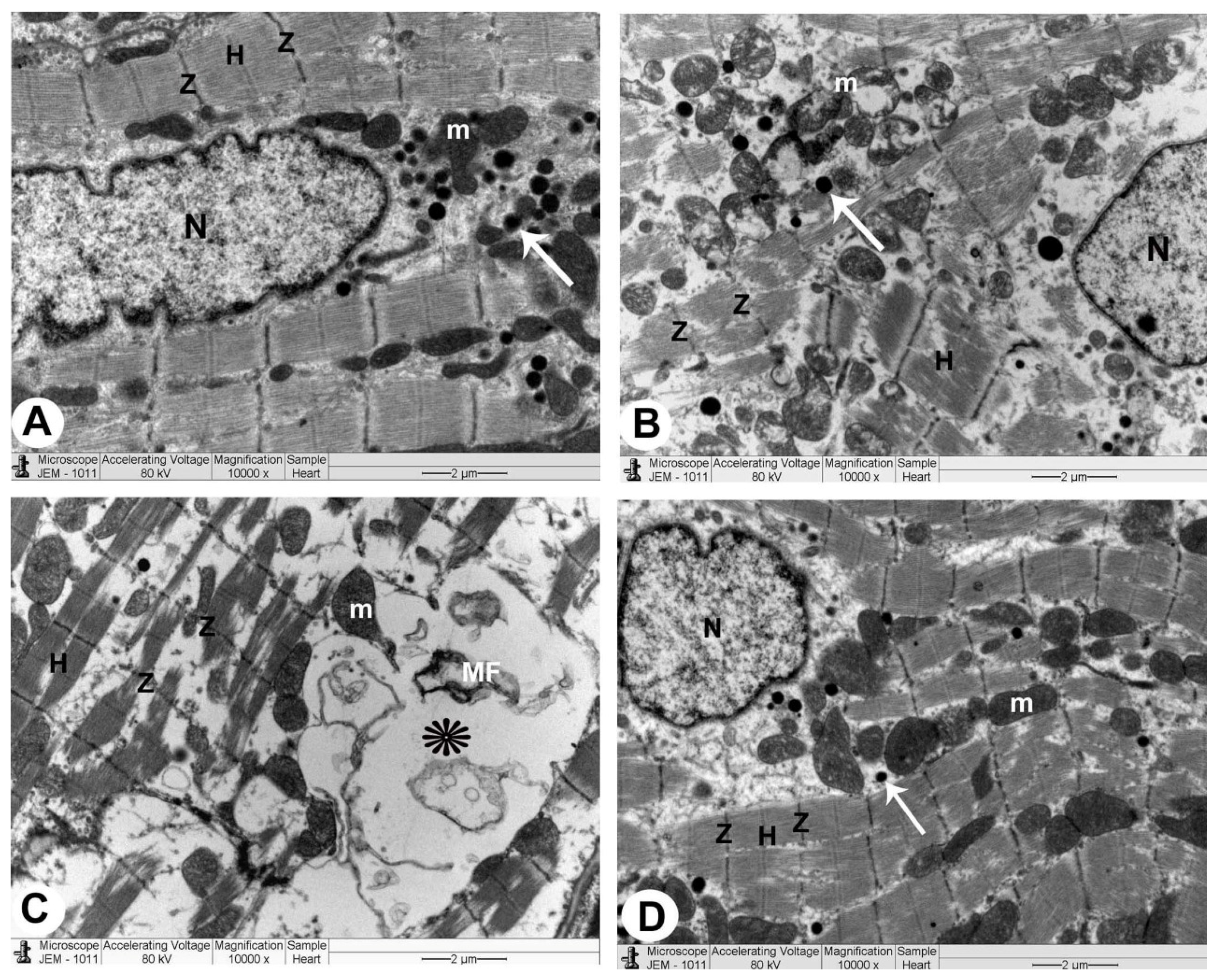
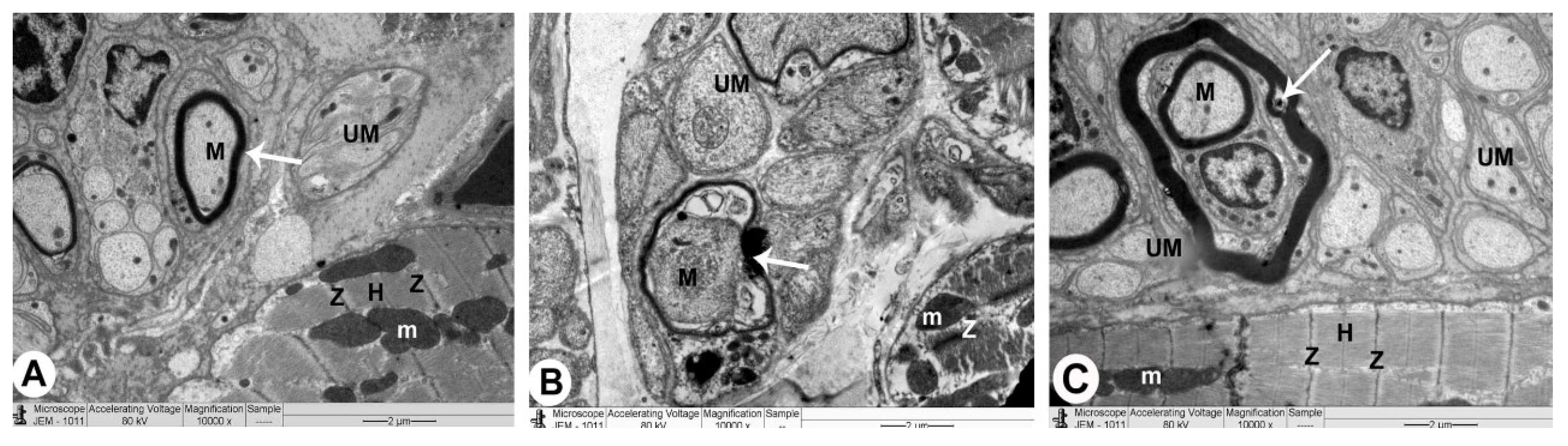
3.3. Electron Microscopy
3.3.1. Control Group
3.3.2. QP-Treated Group
3.3.3. QP- and Vit-C-Treated Group
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abdollahi, M.; Ranjbar, A.; Shadnia, S.; Nikfar, S.; Rezaie, A. Pesticides and oxidative stress: A review. Med. Sci. Monit. 2004, 10, RA141–RA147. [Google Scholar] [PubMed]
- El-Demerdash, F.M. Lipid peroxidation, oxidative stress and acetylcholinesterase in rat brain exposed to organophosphate and pyrethroid insecticides. Food Chem. Toxicol. 2011, 49, 1346–1352. [Google Scholar] [CrossRef]
- El-Demerdash, F.M.; Yousef, M.I.; Kedwany, F.S.; Baghdadi, H.H. Role of α-Tocopherol and β-Carotene in Ameliorating the Fenvalerate-Induced Changes in Oxidative Stress, Hemato-Biochemical Parameters, and Semen Quality of Male Rats. J. Environ. Sci. Health Part B 2004, 39, 443–459. [Google Scholar] [CrossRef] [PubMed]
- Oruç, E.; Üner, N. Combined effects of 2,4-D and azinphosmethyl on antioxidant enzymes and lipid peroxidation in liver of Oreochromis niloticus. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 2000, 127, 291–296. [Google Scholar] [CrossRef]
- Gupta, B.; Rani, M.; Kumar, R.; Dureja, P. Decay profile and metabolic pathways of quinalphos in water, soil and plants. Chemosphere 2011, 85, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, R.; Mohanakumar, K.P.; Chowdhury, M. Effects of an organophosphate pesticide, quinalphos, on the hypothalamo-pituitary-gonadal axis in adult male rats. J. Reprod. Fertil. 2000, 118, 29–38. [Google Scholar] [CrossRef]
- Singh, P.; Verma, P.K.; Raina, R.; Sood, S.; Sharma, P. Maximum contaminant level of arsenic in drinking water potentiates quinalphos-induced renal damage on co-administration of both arsenic and quinalphos in Wistar rats. Environ. Sci. Pollut. Res. 2020, 27, 21331–21340. [Google Scholar] [CrossRef]
- Goodman, J.; Hochstein, P. Generation of free radicals and lipid peroxidation by redox cycling of adriamycin and daunomycin. Biochem. Biophys. Res. Commun. 1977, 77, 797–803. [Google Scholar] [CrossRef]
- Rajagopalan, S.; Landrigan, P.J. Pollution and the Heart. New Engl. J. Med. 2021, 385, 1881–1892. [Google Scholar] [CrossRef]
- Heistad, D.D. Oxidative Stress and Vascular Disease. Arter. Thromb. Vasc. Biol. 2006, 26, 689–695. [Google Scholar] [CrossRef] [Green Version]
- Budin, S.B.; Han, K.J.; Jayusman, P.A.; Taib, I.S.; Ghazali, A.R.; Mohamed, J. Antioxidant Activity of Tocotrienol Rich Fraction Prevents Fenitrothion-induced Renal Damage in Rats. J. Toxicol. Pathol. 2013, 26, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shireen, K.; Pace, R.; Mahboob, M.; Khan, A. Effects of dietary vitamin E, C and soybean oil supplementation on antioxidant enzyme activities in liver and muscles of rats. Food Chem. Toxicol. 2008, 46, 3290–3294. [Google Scholar] [CrossRef]
- Sauberlich, H.E. Pharmacology of Vitamin C. Annu. Rev. Nutr. 1994, 14, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Naidu, K.A. Vitamin C in human health and disease is still a mystery? An overview. Nutr. J. 2003, 2, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carr, A.; Frei, B. Does vitamin C act as a pro-oxidant under physiological conditions? FASEB J. 1999, 13, 1007–1024. [Google Scholar] [CrossRef] [Green Version]
- Sim, M.; Hong, S.; Jung, S.; Kim, J.-S.; Goo, Y.-T.; Chun, W.Y.; Shin, D.-M. Vitamin C supplementation promotes mental vitality in healthy young adults: Results from a cross-sectional analysis and a randomized, double-blind, placebo-controlled trial. Eur. J. Nutr. 2021, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kammon, A.; Brar, R.; Sodhi, S.; Banga, H.; Singh, J.; Nagra, N. Chlorpyrifos chronic toxicity in broilers and effect of vitamin C. Open Vet. J. 2011, 1, 21–27. [Google Scholar]
- Ambali, S.; Akanbi, D.; Igbokwe, N.; Shittu, M.; Kawu, M.; Ayo, J. Evaluation of subchronic chlorpyrifos poisoning on hematological and serum biochemical changes in mice and protective effect of vitamin C. J. Toxicol. Sci. 2007, 32, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raina, R.; Baba, N.A.; Verma, P.K.; Sultana, M.; Singh, M. Hepatotoxicity Induced by Subchronic Exposure of Fluoride and Chlorpyrifos in Wistar Rats: Mitigating Effect of Ascorbic Acid. Biol. Trace Element Res. 2015, 166, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Akpa, A.R.; Ayo, J.O.; Mika’Il, H.G.; Zakari, F.O. Protective effect of fisetin against subchronic chlorpyrifos-induced toxicity on oxidative stress biomarkers and neurobehavioral parameters in adult male albino mice. Toxicol. Res. 2020, 37, 163–171. [Google Scholar] [CrossRef]
- Khalaf, H.A.; El-Mansy, A.A.E.-R. The possible alleviating effect of saffron on chlorpyrifos experimentally induced cardiotoxicity: Histological, immunohistochemical and biochemical study. Acta Histochem. 2019, 121, 472–483. [Google Scholar] [CrossRef]
- Eid, R.A.; Zaki, M.S.A.; Al-Shraim, M.; Eldeen, M.A.; Haidara, M.A. Grape seed extract protects against amiodarone—induced nephrotoxicity and ultrastructural alterations associated with the inhibition of biomarkers of inflammation and oxidative stress in rats. Ultrastruct. Pathol. 2021, 45, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Eid, R.A.; Bin-Meferij, M.M.; El-Kott, A.F.; Eleawa, S.M.; Zaki, M.S.A.; Al-Shraim, M.; El-Sayed, F.; Eldeen, M.A.; Alkhateeb, M.A.; Alharbi, S.A.; et al. Exendin-4 Protects Against Myocardial Ischemia-Reperfusion Injury by Upregulation of SIRT1 and SIRT3 and Activation of AMPK. J. Cardiovasc. Transl. Res. 2020, 14, 619–635. [Google Scholar] [CrossRef]
- Khalili, F.; Vaisi-Raygani, A.; Shakiba, E.; Kohsari, M.; Dehbani, M.; Naseri, R.; Asadi, S.; Rahimi, Z.; Rahimi, M.; Rahimi, Z. Oxidative stress parameters and keap 1 variants in T2DM: Association with T2DM, diabetic neuropathy, diabetic retinopathy, and obesity. J. Clin. Lab. Anal. 2021, e24163. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Haidara, M.A.; Yassin, H.Z.; Zakula, Z.; Mikhailidis, D.P.; Isenovic, E.R. Diabetes and antioxidants: Myth or reality? Curr. Vasc. Pharmacol. 2010, 8, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Araujo, A.S.R.; Schenkel, P.; Enzveiler, A.T.; Fernandes, T.R.G.; Partata, W.A.; Llesuy, S.; Ribeiro, M.F.M.; Khaper, N.; Singal, P.K.; Belló-Klein, A. The role of redox signaling in cardiac hypertrophy induced by experimental hyperthyroidism. J. Mol. Endocrinol. 2008, 41, 423–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yavuz, O.; Cam, M.; Bukan, N.; Guven, A.; Silan, F. Protective effect of melatonin on β-cell damage in streptozotocin-induced diabetes in rats. Acta Histochem. 2003, 105, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, B.; Kribbs, S.B.; ClubbJr, F.J.; Michael, L.H.; Didenko, V.V.; Hornsby, P.J.; Seta, Y.; Oral, H.; Spinale, F.G.; Mann, D.L. Pathophysiologically Relevant Concentrations of Tumor Necrosis Factor-α Promote Progressive Left Ventricular Dysfunction and Remodeling in Rats. Circulation 1998, 97, 1382–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapadia, S.; Lee, J.; Torre-Amione, G.; Birdsall, H.H.; Ma, T.S.; Mann, D.L. Tumor necrosis factor-alpha gene and protein expression in adult feline myocardium after endotoxin administration. J. Clin. Investig. 1995, 96, 1042–1052. [Google Scholar] [CrossRef] [Green Version]
- Takada, M.; Nadeau, K.C.; Hancock, W.W.; MacKenzie, H.S.; Shaw, G.D.; Waaga, A.M.; Chandraker, A.; Sayegh, M.H.; Tilney, N.L. Effects of explosive brain death on cytokine activation of peripheral organs in the RAT1. Transplantation 1998, 65, 1533–1542. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Starkov, A.; Froberg, M.K.; Leino, R.L.; Wallace, K.B. Cumulative and irreversible cardiac mitochondrial dysfunction induced by doxorubicin. Cancer Res. 2001, 61, 771–777. [Google Scholar] [PubMed]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelkka, T.; Kienhöfer, D.; Hoffmann, M.; Linja, M.; Wing, K.; Sareila, O.; Hultqvist, M.; Laajala, E.; Chen, Z.; Vasconcelos, J.; et al. Reactive Oxygen Species Deficiency Induces Autoimmunity with Type 1 Interferon Signature. Antioxid. Redox Signal. 2014, 21, 2231–2245. [Google Scholar] [CrossRef] [Green Version]
- Janssen-Heininger, Y.M.; Mossman, B.T.; Heintz, N.H.; Forman, H.J.; Kalyanaraman, B.; Finkel, T.; Stamler, J.S.; Rhee, S.G.; van der Vliet, A. Redox-based regulation of signal transduction: Principles, pitfalls, and promises. Free. Radic. Biol. Med. 2008, 45, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Wilson, K.P.; Black, J.-A.F.; Thomson, J.A.; Kim, E.E.; Griffith, J.P.; Navia, M.A.; Murcko, M.A.; Chambers, S.P.; Aldape, R.A.; Raybuck, S.A.; et al. Structure and mechanism of interleukin-lβ converting enzyme. Nature 1994, 370, 270–275. [Google Scholar] [CrossRef]
- Zankov, D.P.; Shimizu, A.; Tanaka-Okamoto, M.; Miyoshi, J.; Ogita, H. Protective effects of intercalated disk protein afadin on chronic pressure overload-induced myocardial damage. Sci. Rep. 2017, 7, 39335. [Google Scholar] [CrossRef] [Green Version]
- Werner, N.; Nickenig, G.; Sinning, J.M. Complex PCI procedures: Challenges for the interventional cardiologist. Clin. Res. Cardiol. 2018, 107, 64–73. [Google Scholar] [CrossRef]
- Li, H.; Jia, J.-P.; Xu, M.; Zhang, L. Changes in the blood-nerve barrier after sciatic nerve cold injury: Indications supporting early treatment. Neural Regen. Res. 2015, 10, 419–424. [Google Scholar] [CrossRef]
- Cantin, M.; Genest, J. The Heart and the Atrial Natriuretic Factor. Endocr. Rev. 1985, 6, 107–127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Moalem, J.; Tse, J.; Scholz, P.M.; Weiss, H.R. Effects of natriuretic peptides on ventricular myocyte contraction and role of cyclic GMP signaling. Eur. J. Pharmacol. 2005, 510, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Hirose, M.; Furukawa, Y.; Kurogouchi, F.; Nakajima, K.; Miyashita, Y.; Chiba, S. C-type natriuretic peptide increases myocardial contractility and sinus rate mediated by guanylyl cyclase-linked natriuretic peptide receptors in isolated, blood-perfused dog heart preparations. J. Pharmacol. Exp. Ther. 1998, 286, 70–76. [Google Scholar] [PubMed]
- Haidara, M.A.; Khloussy, H.; Ammar, H.; Kassem, L.A.A. Impact of alpha-tocopherol and vitamin C on endothelial markers in rats with streptozotocin-induced diabetes. Med. Sci. Monit. 2004, 10, BR41–BR46. [Google Scholar]
- Giampieri, F.; Alvarez-Suarez, J.M.; Mazzoni, L.; Forbes-Hernandez, T.Y.; Gasparrini, M.; Gonzàlez-Paramàs, A.M.; Santos-Buelga, C.; Quiles, J.L.; Bompadre, S.; Mezzetti, B.; et al. Polyphenol-Rich Strawberry Extract Protects Human Dermal Fibroblasts against Hydrogen Peroxide Oxidative Damage and Improves Mitochondrial Functionality. Molecules 2014, 19, 7798–7816. [Google Scholar] [CrossRef] [Green Version]
- Lynch, S.M.; Gaziano, J.M.; Frei, B. Ascorbic Acid and Atherosclerotic Cardiovascular Disease. Subcell. Biochem. 1996, 25, 331–367. [Google Scholar] [CrossRef]
- Grandjean, P.; Landrigan, P.J. Developmental neurotoxicity of industrial chemicals: A silent pandemic. Lancet 2006, 368, 2167–2178. [Google Scholar] [CrossRef]
- Guan, Z.-Z.; Yu, W.-F.; Nordberg, A. Dual effects of nicotine on oxidative stress and neuroprotection in PC12 cells. Neurochem. Int. 2003, 43, 243–249. [Google Scholar] [CrossRef]
- Olanow, C.W.; Arendash, G.W. Metals and free radicals in neurodegeneration. Curr. Opin. Neurol. 1994, 7, 548–558. [Google Scholar] [CrossRef]
- Slotkin, T.A.; Seidler, F.J. Oxidative stress from diverse developmental neurotoxicants: Antioxidants protect against lipid peroxidation without preventing cell loss. Neurotoxicol. Teratol. 2010, 32, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Karasu-Minareci, E.; Gunay, N.; Minareci, K.; Sadan, G.; Ozbey, G. What may be happen after an organophosphate exposure: Acute myocardial infarction? J. Forensic Leg. Med. 2011, 19, 94–96. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Aspects | Control Rats | QP-Treated Rats | QP + Vit-C-Treated Rats |
|---|---|---|---|---|
| 1 | Atrionatriuretic factor (ANF) granules | Small electron-dense granules focused mainly in the centre of the cell, between both plasma membrane and peripheral muscle fibres, and close to the nucleus. | Only a minority and most of the granules of various sizes are disturbed. | Normal distribution and normal size granules |
| 2 | Intercalated discs | Normal | Damaged | Intact |
| • Fascia adherence | Healthy | Discrepancies | Normal | |
| • Macula adherence | Normal | Abnormalities | Normal | |
| • Gap junctions (Nexus) | Intact | Lost | Reconstruction | |
| 3 | Cardiomyocytes | Normal architecture | Degenerated myofibrils | Intact architecture |
| • Myofibrils | Preserved sarcomeres | Damaged sarcomeres | Normal sarcomeres | |
| • Z and H bands | Clear | Disrupted | Intact | |
| • Mitochondria | Intact | Disorganised | Preserved | |
| 4 | Nerve endings in interstitial areas | Normal | Damaged | Intact |
| • Myelinated fibres | Intact with intact myelin sheaths | Damaged with ruptured myelin sheaths | Normal with intact myelin sheaths | |
| • Unmyelinated fibres | Intact | Detached | Intact |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shati, A.A.; Zaki, M.S.A.; Alqahtani, Y.A.; Haidara, M.A.; Al-Shraim, M.; Dawood, A.F.; Eid, R.A. Potential Protective Effect of Vitamin C on Qunalphos-Induced Cardiac Toxicity: Histological and Tissue Biomarker Assay. Biomedicines 2022, 10, 39. https://doi.org/10.3390/biomedicines10010039
Shati AA, Zaki MSA, Alqahtani YA, Haidara MA, Al-Shraim M, Dawood AF, Eid RA. Potential Protective Effect of Vitamin C on Qunalphos-Induced Cardiac Toxicity: Histological and Tissue Biomarker Assay. Biomedicines. 2022; 10(1):39. https://doi.org/10.3390/biomedicines10010039
Chicago/Turabian StyleShati, Ayed A., Mohamed Samir A. Zaki, Youssef A. Alqahtani, Mohamed A. Haidara, Mubarak Al-Shraim, Amal F. Dawood, and Refaat A. Eid. 2022. "Potential Protective Effect of Vitamin C on Qunalphos-Induced Cardiac Toxicity: Histological and Tissue Biomarker Assay" Biomedicines 10, no. 1: 39. https://doi.org/10.3390/biomedicines10010039