Maternal Metformin Treatment Improves Developmental and Metabolic Traits of IUGR Fetuses

 , , , , and
, , , , and
Abstract
:1. Introduction
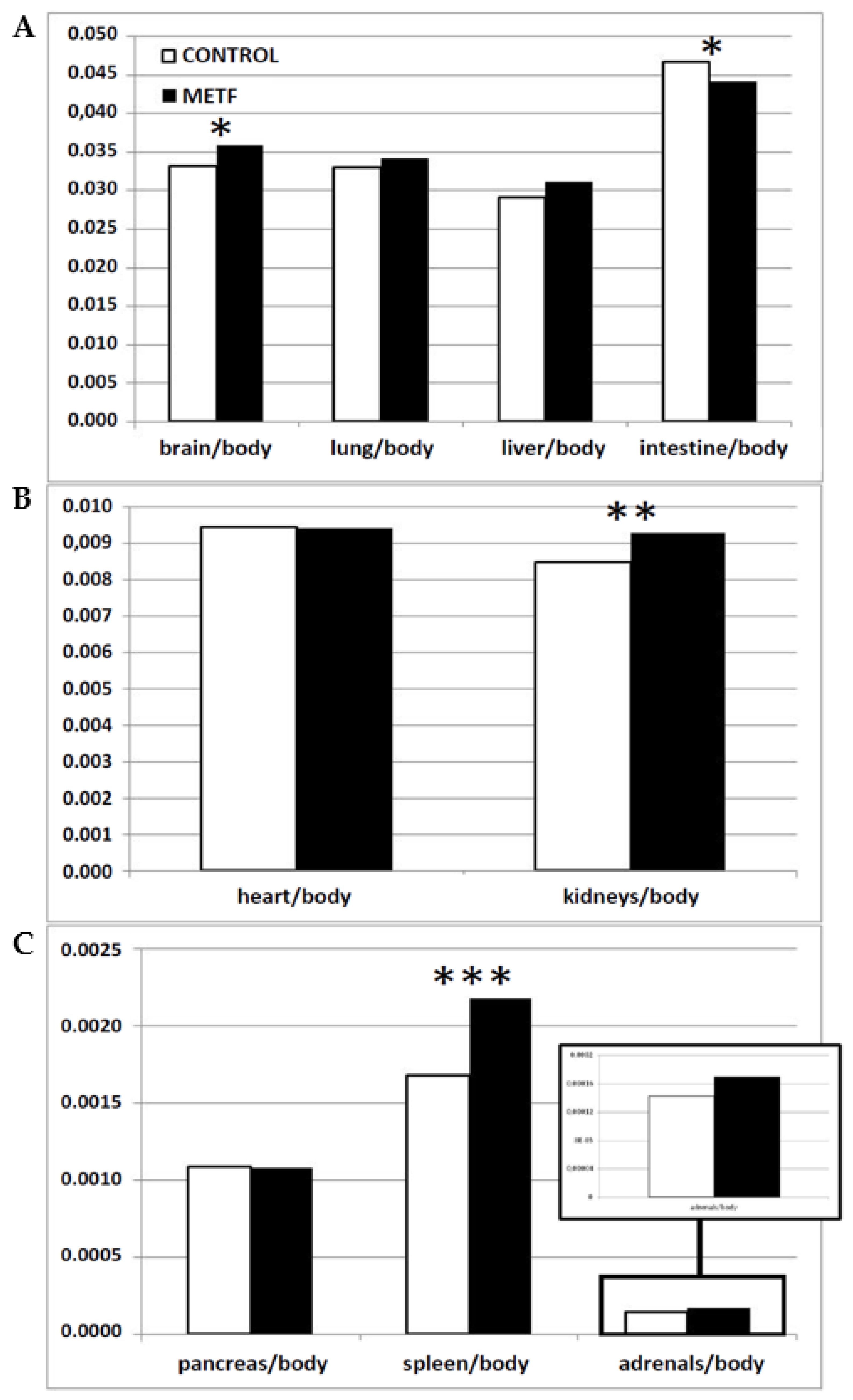
2. Results
2.1. Effects of Metformin Treatment on Maternal Weight and Metabolism
2.2. Effects of Metformin Treatment on Fetal Size and Metabolism
3. Discussion
4. Material and Methods
4.1. Animals and Ethics Statement
4.2. Animal Handling and Experimental Procedure
4.3. Measuring, Weighting and Sampling of Fetuses
4.4. Evaluation of Maternal and Fetal Metabolic Status
4.5. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bailey, C.J. Metformin: historical overview. Diabetologia 2017, 60, 1566–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsay, R.S.; Loeken, M.R. Metformin use in pregnancy: Promises and uncertainties. Diabetologia 2017, 60, 1612–1619. [Google Scholar] [CrossRef]
- Hyer, S.; Balani, J.; Shehata, H. Metformin in pregnancy: Mechanisms and clinical applications. Int. J. Mol. Sci. 2018, 19, 1954. [Google Scholar] [CrossRef]
- Charles, B.; Norris, R.; Xiao, X.; Hague, W. Population pharmacokinetics of metformin in late pregnancy. Ther. Drug Monit. 2006, 28, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Simmons, D. Safety considerations with pharmacological treatment of gestational diabetes mellitus. Drug Saf. 2015, 38, 65–78. [Google Scholar] [CrossRef]
- Stumvoll, M.; Nurjhan, N.; Perriello, G.; Dailey, G.; Gerich, J.E. Metabolic effects of metformin in non-insulin-dependent diabetes mellitus. N. Engl. J. Med. 1995, 333, 550–554. [Google Scholar] [CrossRef] [PubMed]
- Astiz, S.; Gonzalez-Bulnes, A.; Astiz, I.; Barbero, A.; Perez-Solana, M.L.; Garcia-Real, I. Advanced onset of puberty after metformin therapy in swine with thrifty genotype. Exp. Physiol. 2014, 99, 1241–1252. [Google Scholar] [CrossRef] [Green Version]
- Scarpello, J.H.; Howlett, H.C. Metformin therapy and clinical uses. Diab. Vasc. Dis. Res. 2008, 5, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Sadava, D.; Frykman, P.; Harris, E.; Majerus, D.; Mustard, J.; Bernard, B. Development of enzymes of glycolysis and gluconeogenesis in human fetal liver. Biol. Neonate 1992, 62, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Bulnes, A.; Astiz, S.; Vazquez-Gomez, M.; Garcia-Contreras, C. Developmental origins of metabolic disorders: The need for biomarker candidates and therapeutic targets from adequate preclinical models. EuPA Open Proteom. 2016, 10, 50–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Bulnes, A.; Ovilo, C.; Lopez-Bote, C.J.; Astiz, S.; Ayuso, M.; Perez-Solana, M.L.; Sanchez-Sanchez, R.; Torres-Rovira, L. Gender-specific early postnatal catch-up growth after intrauterine growth retardation by food restriction in swine with obesity/leptin resistance. Reproduction 2012, 144, 269–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Type 2 diabetes and metformin. First choice for monotherapy: weak evidence of efficacy but well-known and acceptable adverse effects. Prescrire Int. 2014, 23, 269–272. Available online: https://www.ncbi.nlm.nih.gov/pubmed/25954799 (accessed on 28 April 2019).
- Mahan, D.C.; Watts, M.R.; St-Pierre, N. Macro- and micromineral composition of fetal pigs and their accretion rates during fetal development. J. Anim. Sci. 2009, 87, 2823–2832. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, A.M. The fetal circulation and its response to stress. J. Dev. Physiol. 1984, 6, 11–19. [Google Scholar] [PubMed]
- Peeling, A.N.; Smart, J.L. Review of literature showing that undernutrition affects the growth rate of all processes in the brain to the same extent. Metab. Brain Dis. 1994, 9, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Salihagic-Kadic, A.; Medic, M.; Jugovic, D.; Kos, M.; Latin, V.; Kusan Jukic, M.; Arbeille, P. Fetal cerebrovascular response to chronic hypoxia—Implications for the prevention of brain damage. J. Matern. Fetal Neonatal Med. 2006, 19, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Perry, B.D. Childhood experience and the expression of genetic potential: What childhood neglect tells us about nature and nurture. Brain Mind 2002, 3, 79–100. [Google Scholar] [CrossRef]
- Latini, G.; De Mitri, B.; Del Vecchio, A.; Chitano, G.; De Felice, C.; Zetterstrom, R. Foetal growth of kidneys, liver and spleen in intrauterine growth restriction: “Programming” causing “metabolic syndrome” in adult age. Acta Paediatr. 2004, 93, 1635–1639. [Google Scholar] [CrossRef] [PubMed]
- Haugen, G.; Hanson, M.; Kiserud, T.; Crozier, S.; Inskip, H.; Godfrey, K.M. Fetal liver-sparing cardiovascular adaptations linked to mother’s slimness and diet. Circ. Res. 2005, 96, 12–14. [Google Scholar] [CrossRef]
- Dawkins, M.J. Biochemical aspects of developing function in newborn mammalian liver. Br. Med. Bull. 1966, 22, 27–33. [Google Scholar] [CrossRef]
- Shelley, H.J. Glycogen rfserves and their changes at birth and in anoxia. Br. Med. Bull. 1961, 17, 137–143. [Google Scholar] [CrossRef]
- McMillen, I.C.; Adams, M.B.; Ross, J.T.; Coulter, C.L.; Simonetta, G.; Owens, J.A.; Robinson, J.S.; Edwards, L.J. Fetal growth restriction: adaptations and consequences. Reproduction 2001, 122, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Pike, K.; Jane Pillow, J.; Lucas, J.S. Long term respiratory consequences of intrauterine growth restriction. Semin. Fetal Neonatal Med. 2012, 17, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Briana, D.D.; Malamitsi-Puchner, A. Small for gestational age birth weight: impact on lung structure and function. Paediatr. Respir. Rev. 2013, 14, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Puddu, M.; Fanos, V.; Podda, F.; Zaffanello, M. The kidney from prenatal to adult life: perinatal programming and reduction of number of nephrons during development. Am. J. Nephrol 2009, 30, 162–170. [Google Scholar] [CrossRef]
- Ritz, E.; Amann, K.; Koleganova, N.; Benz, K. Prenatal programming-effects on blood pressure and renal function. Nat. Rev. Nephrol. 2011, 7, 137–144. [Google Scholar] [CrossRef]
- Dotsch, J.; Alejandre-Alcazar, M.; Janoschek, R.; Nusken, E.; Weber, L.T.; Nusken, K.D. Perinatal programming of renal function. Curr. Opin. Pediatr. 2016, 28, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Paixao, A.D.; Alexander, B.T. How the kidney is impacted by the perinatal maternal environment to develop hypertension. Biol. Reprod. 2013, 89, 144. [Google Scholar] [CrossRef]
- Kolacek, S.; Puntis, J.W.; Lloyd, D.R.; Brown, G.A.; Booth, I.W. Ontogeny of pancreatic exocrine function. Arch. Dis. Child. 1990, 65, 178–181. [Google Scholar] [CrossRef]
- D’Inca, R.; Kloareg, M.; Gras-Le Guen, C.; Le Huerou-Luron, I. Intrauterine growth restriction modifies the developmental pattern of intestinal structure, transcriptomic profile, and bacterial colonization in neonatal pigs. J. Nutr. 2010, 140, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Bakhashab, S.; Ahmed, F.; Schulten, H.J.; Ahmed, F.W.; Glanville, M.; Al-Qahtani, M.H.; Weaver, J.U. Proangiogenic effect of metformin in endothelial cells is via upregulation of VEGFR1/2 and their signaling under hyperglycemia-hypoxia. Int. J. Mol. Sci. 2018, 19, 293. [Google Scholar] [CrossRef]
- Gargett, C.E.; Rogers, P.A. Human endometrial angiogenesis. Reproduction 2001, 121, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Przala, J.; Gregoraszczuk, E.L.; Kotwica, G.; Stefanczyk-Krzymowska, S.; Ziecik, A.J.; Blitek, A.; Ptak, A.; Rak, A.; Wojtowicz, A.; Kaminski, T.; et al. Mechanisms ensuring optimal conditions of implantation and embryo development in the pig. Reprod. Biol. 2006, 6, 59–87. [Google Scholar] [PubMed]
- Shelley, H.J.; Bassett, J.M.; Milner, R.D. Control of carbohydrate metabolism in the fetus and newborn. Br. Med. Bull. 1975, 31, 37–43. [Google Scholar] [CrossRef]
- Szabo, A.J.; De Lellis, R.; Grimaldi, R.D. Triglyceride synthesis by the human placenta. I. Incorporation of labeled palmitate into placental triglycerides. Am. J. Obstetr. Gynecol. 1973, 115, 257–262. [Google Scholar] [CrossRef]
- Coleman, R.A.; Haynes, E.B. Microsomal and lysosomal enzymes of triacylglycerol metabolism in rat placenta. Biochem. J. 1984, 217, 391–397. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association. Management of diabetes in pregnancy: Standards of medical care in diabetes 2019. Diabetes Care 2019, 42, S165–S172. [Google Scholar] [CrossRef]
- Hanem, L.G.E.; Stridsklev, S.; Júlíusson, P.B.; Salvesen, Ø.; Roelants, M.; Carlsen, S.M.; Ødegård, R.; Vanky, E. Metformin use in PCOS pregnancies increases the risk of offspring overweight at 4 years of age: Follow-up of two RCTs. J. Clin. Endocrinol. Metab. 2018, 103, 1612–1621. [Google Scholar] [CrossRef]
- Rowan, J.A.; Rush, E.C.; Plank, L.D.; Lu, J.; Obolonkin, V.; Coat, S.; Hague, W.M. Metformin in gestational diabetes: the offspring follow-up (MiG TOFU): Body composition and metabolic outcomes at 7–9 years of age. BMJ Open Diabetes Res. Care 2018, 6, e000456. [Google Scholar] [CrossRef]
- Garbarino, V.R.; Santos, T.A.; Nelson, A.R.; Zhang, W.Q.; Smolik, C.M.; Javors, M.A.; Daws, L.C.; Gould, G.G. Prenatal metformin exposure or organic cation transporter 3 knock-out curbs social interaction preference in male mice. Pharmacol. Res. 2019, 140, 21–32. [Google Scholar] [CrossRef]
- Forcato, S.; Novi, D.R.B.D.S.; Costa, N.O.; Borges, L.I.; Góes, M.L.M.; Ceravolo, G.S.; Gerardin, D.C.C. In utero and lactational exposure to metformin induces reproductive alterations in male rat offspring. Reprod. Toxicol. 2017, 74, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Walsh Hentges, L.S.; Martin, R.J. Serum and lipoprotein lipids of fetal pigs and their dams during gestation as compared with man. Biol. Neonate 1987, 52, 127–134. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Group C | Group METF | ||
|---|---|---|---|
| Body morphometry | Body weight | 111.2 ± 1.1 | 113.8 ± 2.1 |
| Back-fat depth | 21.7 ± 5.0 | 24.4 ± 5.0 | |
| Glucose metabolisms | Glucose | 90.3 ± 0.8 | 85.3 ± 1.2 |
| Fructosamine | 302.1 ± 4.0 | 266.7 ± 2.6 | |
| Lipid metabolisms | Triglycerides | 52.3 ± 2.4 | 57.7 ± 2.1 |
| Total cholesterol | 58.6 ± 1.7 | 54.9 ± 1.0 | |
| HDL-cholesterol | 25.6 ± 1.0 | 23.2 ± 0.7 | |
| LDL-cholesterol | 31.2 ± 1.5 | 33.1 ± 1.6 |
| Group C | Group METF | Increase (%) | |
|---|---|---|---|
| Body weight (g) | 756.9 ± 28.1 | 806.7 ± 25.8 | 6.6 |
| Body length (cm) | 21.4 ± 0.3 | 21.1 ± 0.4 | 0.0 |
| Occipito-nasal length (cm) | 11.1 ± 0.2 | 11.1 ± 0.3 | 0.0 |
| Biparietal diameter (cm) | 4.4 ± 0.1 | 4.5 ± 0.1 | 2.3 |
| Thoracic circumference (cm) | 18.3 ± 0.3 | 17.8 ± 0.3 | 0.0 |
| Abdominal circumference (cm) | 14.0 ± 0.3 | 13.9 ± 0.3 | 0.0 |
| Head weight (g) | 171.6 ± 5.1 | 178.1 ± 5.3 | 3.8 |
| Carcass weight (g) | 428.6 ± 17.4 | 445.3 ± 15.9 | 3.9 |
| Brain weight (g) | 25.5 ± 0.5 g | 28.1 ± 0.4 h | 10.2 |
| Total viscera weight (g) | 122.0 ± 6.7 c | 137.5 ± 4.7 d | 12.7 |
| Heart weight (g) | 7.2 ± 0.4 | 7.4 ± 0.3 | 2.8 |
| Lungs weight (g) | 24.9 ± 1.1 a | 27.7 ± 1.1 b | 11.2 |
| Liver weight (g) | 22.2 ± 1.0 c | 25.4 ± 1.2 d | 14.4 |
| Intestine weight (g) | 35.4 ± 1.5 | 35.1 ± 1.3 | 0.0 |
| Kidneys weight (g) | 6.4 ± 0.3 c | 7.4 ± 0.3 d | 15.6 |
| Spleen weight (mg) | 129.1 ± 7.7 g | 172.1 ± 7.4 h | 33.3 |
| Pancreas weight (mg) | 834.9 ± 5.3 | 862.6 ± 5.5 | 3.3 |
| Adrenal glands (mg) | 105.5 ± 5.5 c | 134.4 ± 1.0 d | 27.4 |
| Placental weight (g) | 253.2 ± 12.7 a | 286.2 ± 12.6 b | 13.0 |
| Group C | Group METF | |
|---|---|---|
| Glucose | 176.1 ± 3.6 e | 260.3 ± 2.4 f |
| Fructosamine | 126.1 ± 5.1 g | 189.8 ± 7.1 h |
| Triglycerides | 48.6 ± 3.8 g | 67.5 ± 4.3 h |
| Total cholesterol | 53.7 ± 3.1 | 56.9 ± 2.3 |
| HDL-cholesterol | 25.2 ± 2.7 | 17.5 ± 0.7 |
| LDL-cholesterol | 33.5 ± 2.1 | 34.2 ± 1.6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Contreras, C.; Vazquez-Gomez, M.; Pesantez-Pacheco, J.L.; Torres-Rovira, L.; Heras-Molina, A.; Encinas, T.; Astiz, S.; Gonzalez-Bulnes, A. Maternal Metformin Treatment Improves Developmental and Metabolic Traits of IUGR Fetuses. Biomolecules 2019, 9, 166. https://doi.org/10.3390/biom9050166
Garcia-Contreras C, Vazquez-Gomez M, Pesantez-Pacheco JL, Torres-Rovira L, Heras-Molina A, Encinas T, Astiz S, Gonzalez-Bulnes A. Maternal Metformin Treatment Improves Developmental and Metabolic Traits of IUGR Fetuses. Biomolecules. 2019; 9(5):166. https://doi.org/10.3390/biom9050166
Chicago/Turabian StyleGarcia-Contreras, Consolación, Marta Vazquez-Gomez, José Luis Pesantez-Pacheco, Laura Torres-Rovira, Ana Heras-Molina, Teresa Encinas, Susana Astiz, and Antonio Gonzalez-Bulnes. 2019. "Maternal Metformin Treatment Improves Developmental and Metabolic Traits of IUGR Fetuses" Biomolecules 9, no. 5: 166. https://doi.org/10.3390/biom9050166