
Transcriptomic Effects on the Mouse Heart Following 30 Days on the International Space Station
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Flight and Control Conditions
2.2. Tissue Collection and RNA Extraction for Sequencing
2.3. RNA Sequencing and Analysis
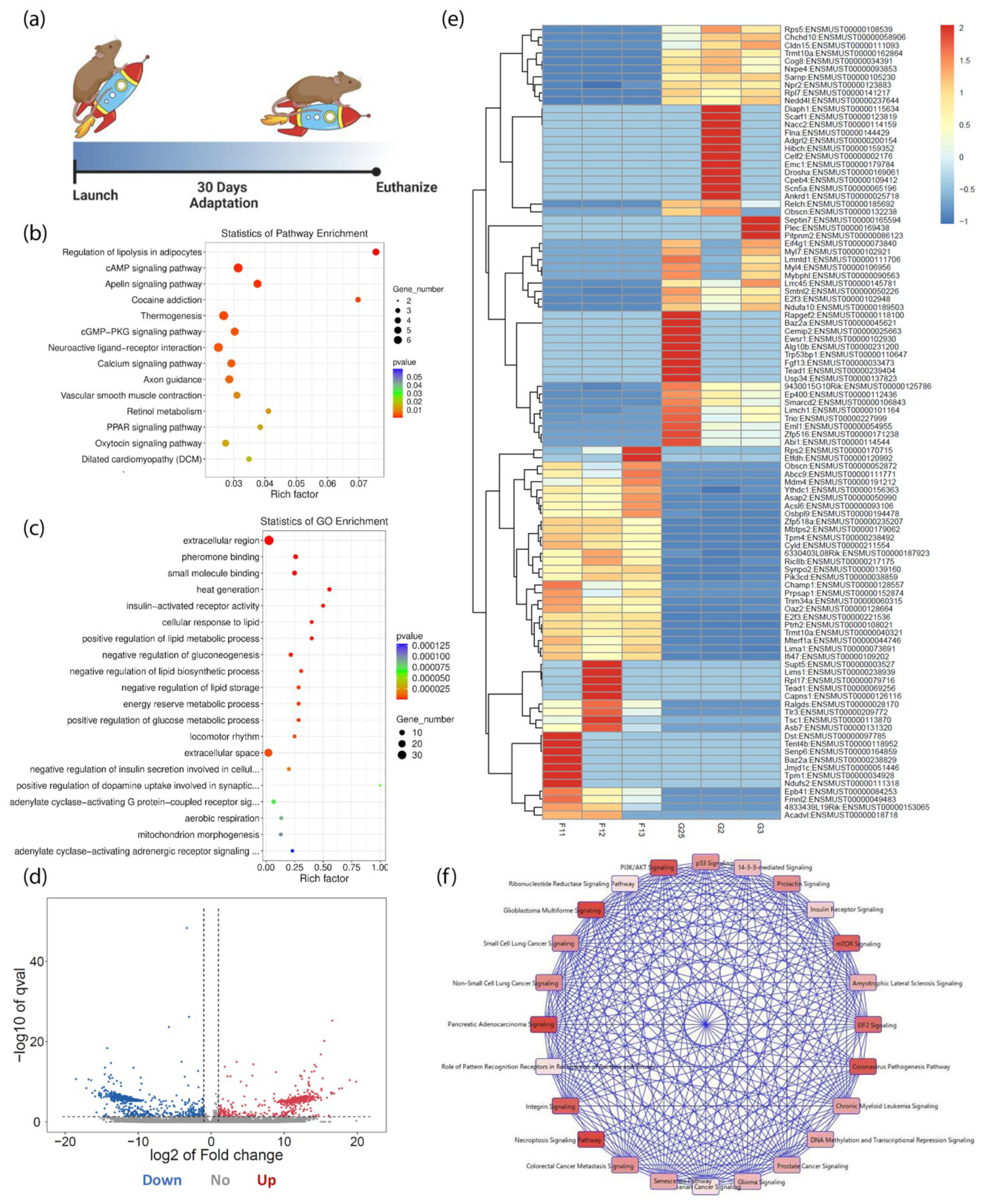
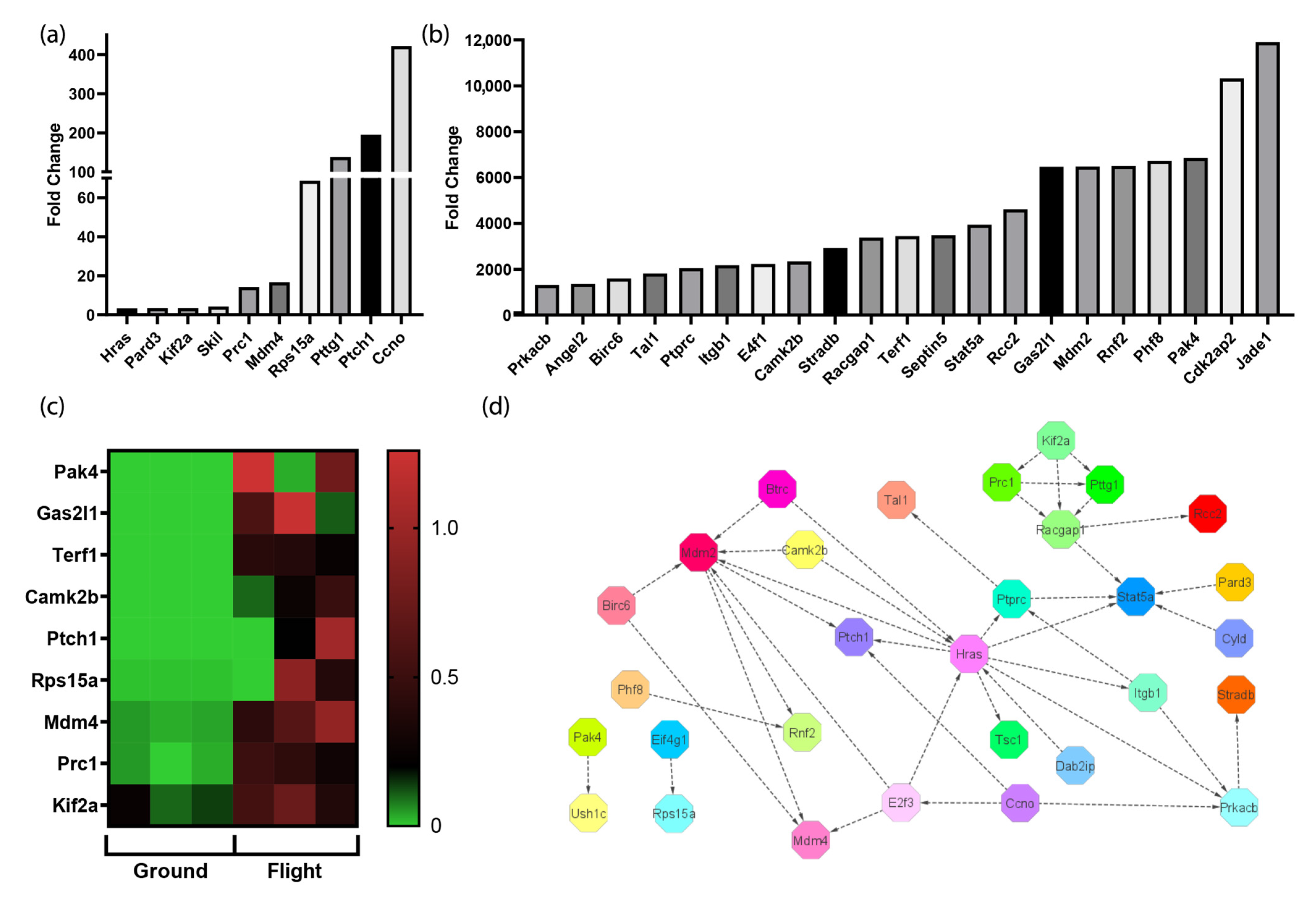
3. Results
3.1. Cardiac Transcriptome Alterations Induced by 30 Days in Spaceflight
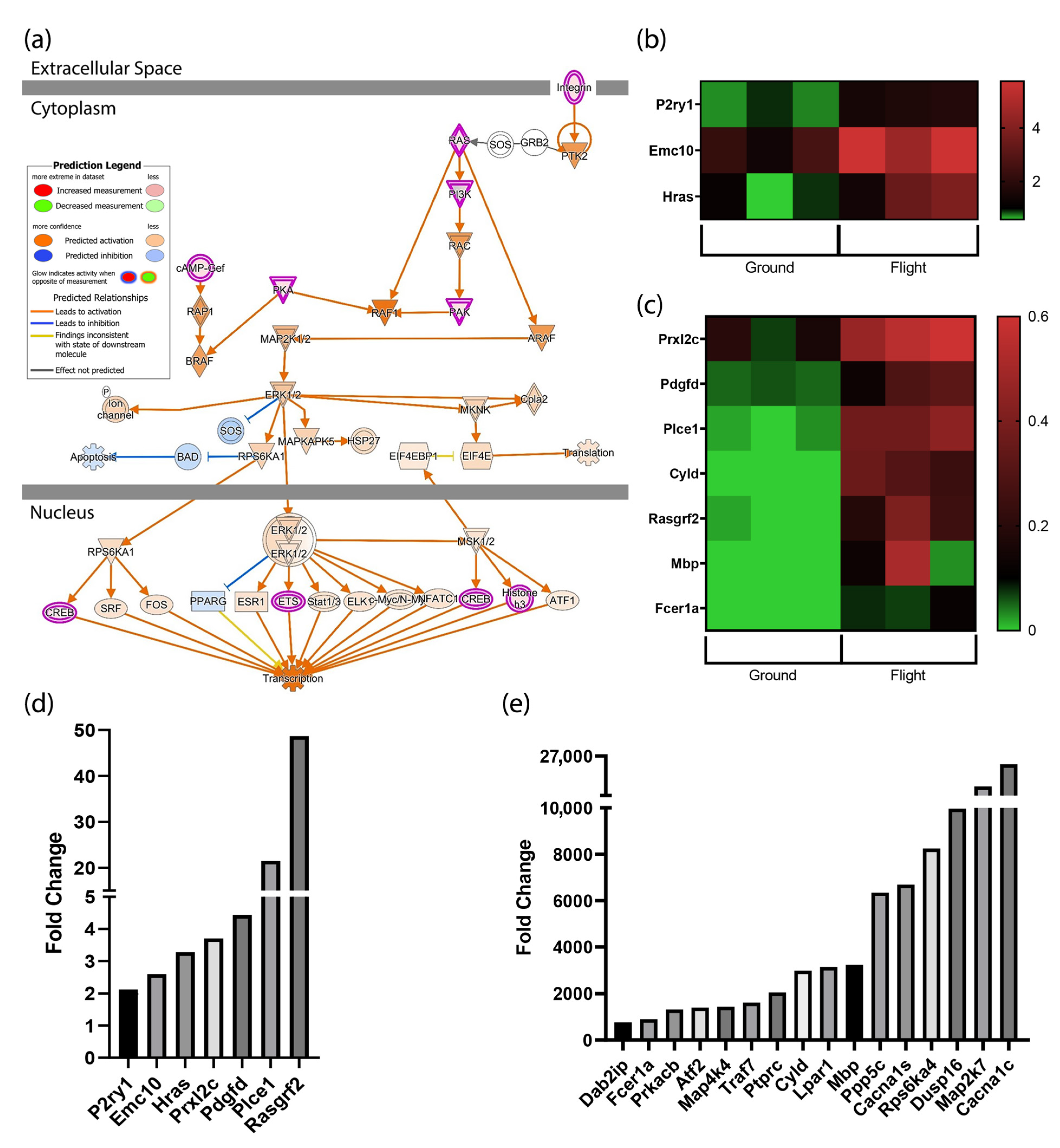
3.2. MAPK Signaling Is Activated by Spaceflight
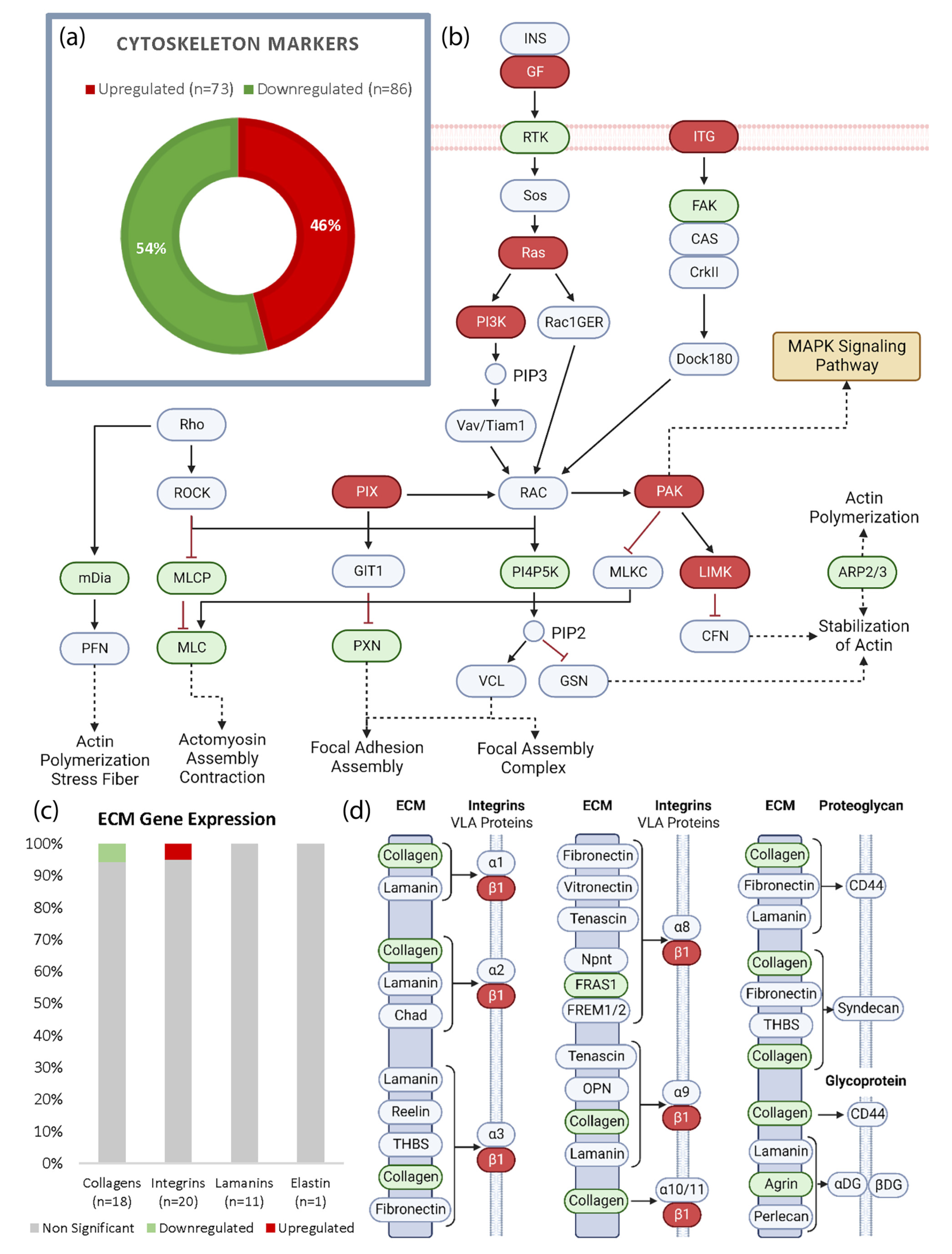
3.3. Spaceflight Modifies Actin but Has Limited Effect on the ECM in the Heart
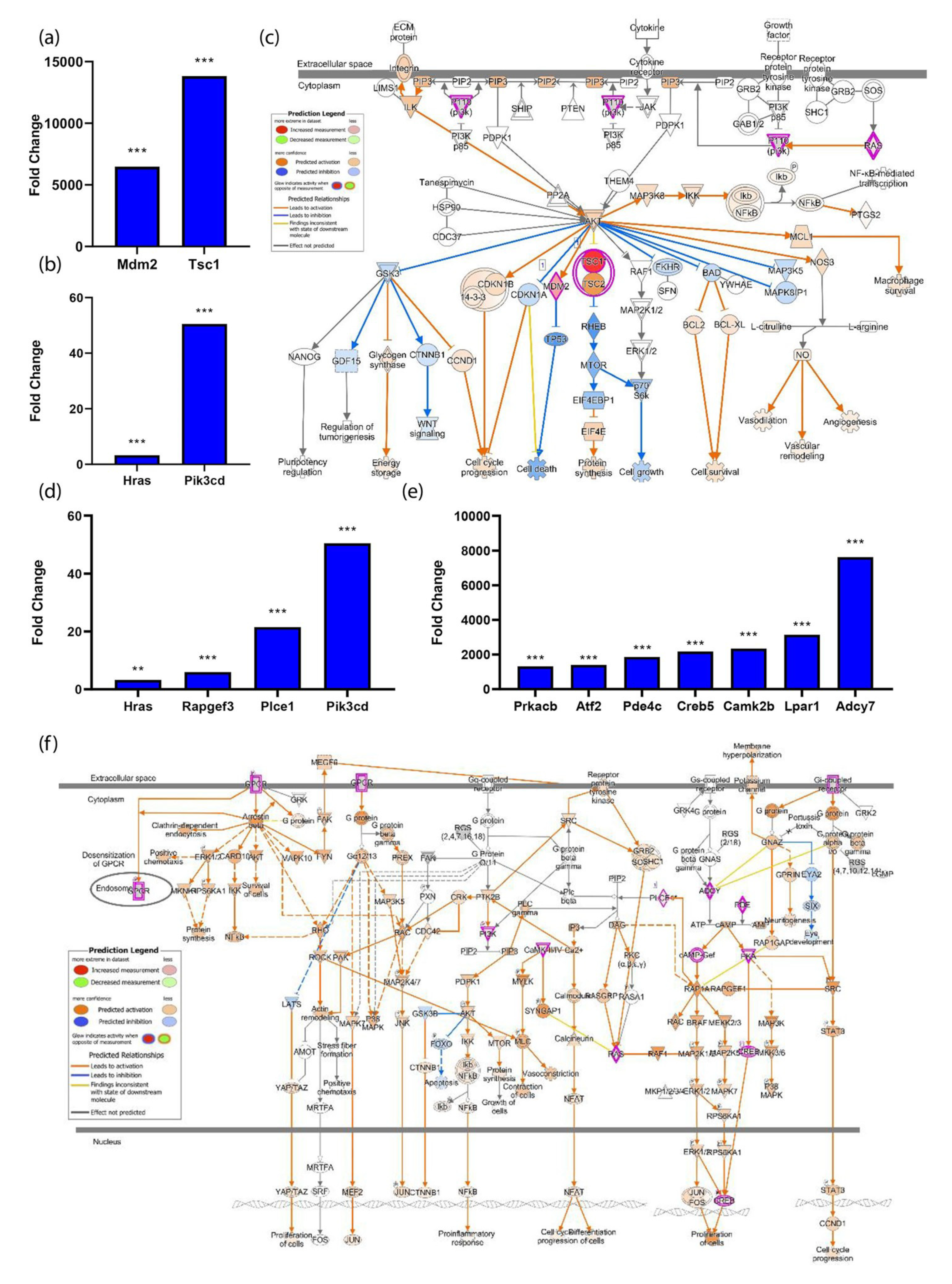
3.4. The PI3K-Akt Pathway Is Induced by Spaceflight
3.5. The Spaceflight Environment Activates Transcripts Associated with the Cell Cycle
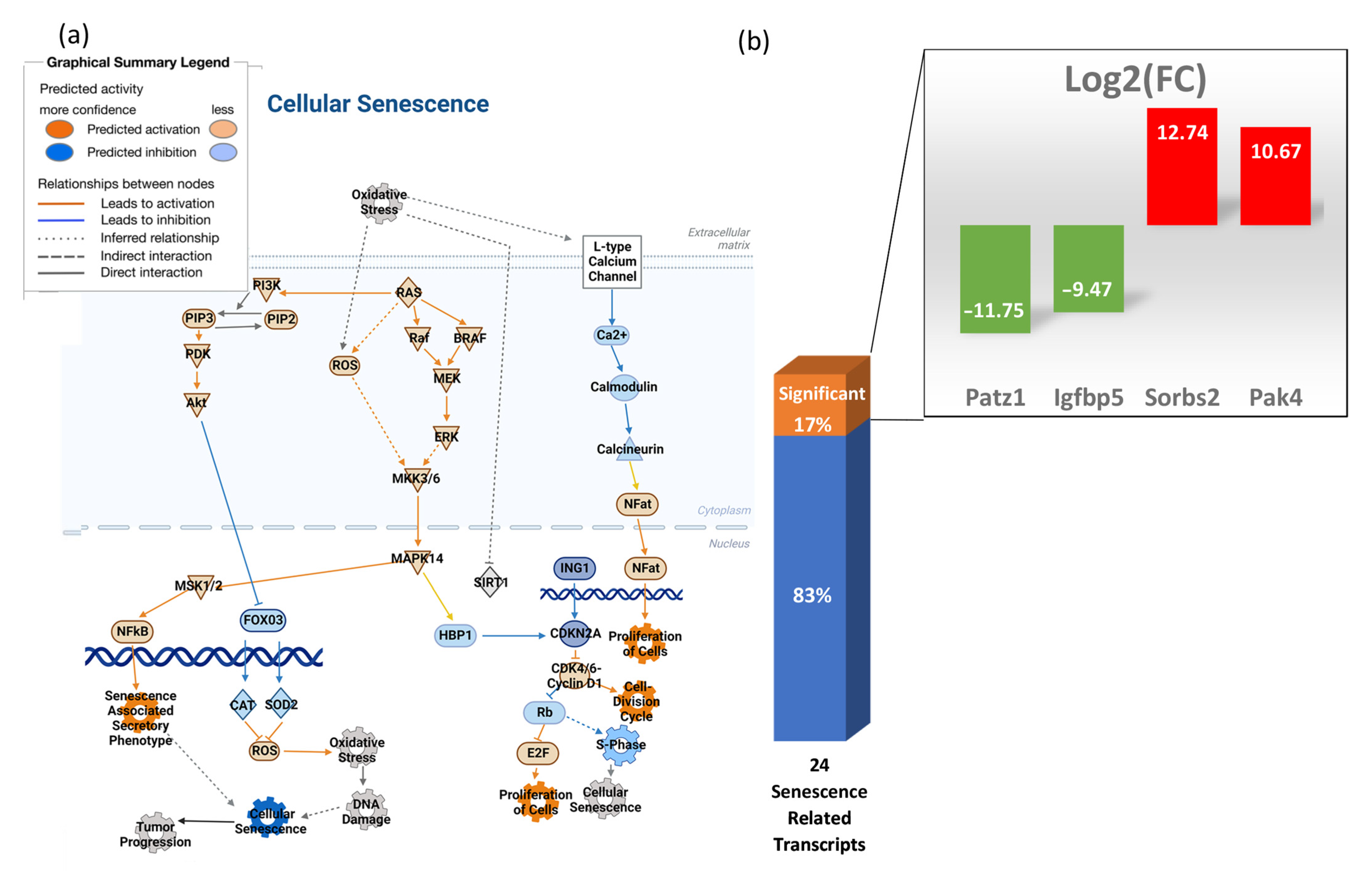
3.6. Transcripts Associated with Cellular Senescence Are Not Significantly Elevated in the Heart
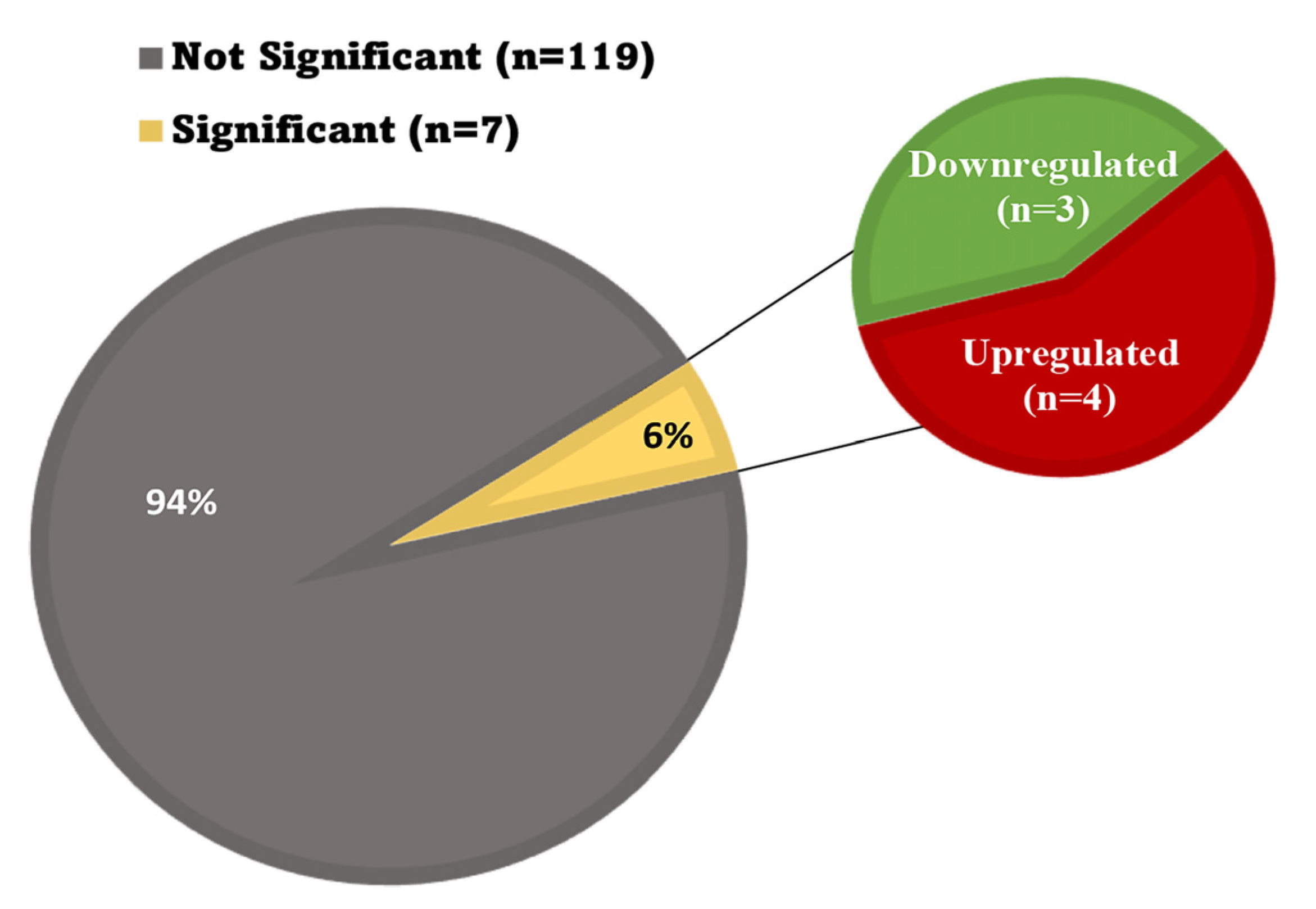
3.7. Oxidative Stress in the Heart
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marshall-Goebel, K.; Macias, B.R.; Laurie, S.S.; Lee, S.M.C.; Ebert, D.J.; Kemp, D.T.; Miller, A.; Greenwald, S.H.; Martin, D.S.; Young, M.; et al. Mechanical countermeasures to headward fluid shifts. J. Appl. Physiol. 2021, 130, 1766–1777. [Google Scholar] [CrossRef]
- Kandarpa, K.; Schneider, V.; Ganapathy, K. Human health during space travel: An overview. Neurol. India 2019, 67, S176–S181. [Google Scholar] [CrossRef]
- Baisch, F.; Beck, L.; Blomqvist, G.; Wolfram, G.; Drescher, J.; Rome, J.L.; Drummer, C. Cardiovascular response to lower body negative pressure stimulation before, during, and after space flight. Eur. J. Clin. Investig. 2000, 30, 1055–1065. [Google Scholar] [CrossRef] [PubMed]
- Khine, H.W.; Steding-Ehrenborg, K.; Hastings, J.L.; Kowal, J.; Daniels, J.D.; Page, R.L.; Goldberger, J.J.; Ng, J.; Adams-Huet, B.; Bungo, M.W.; et al. Effects of Prolonged Spaceflight on Atrial Size, Atrial Electrophysiology, and Risk of Atrial Fibrillation. Circ. Arrhythm. Electrophysiol. 2018, 11, e005959. [Google Scholar] [CrossRef]
- Scott, J.M.; Stoudemire, J.; Dolan, L.; Downs, M. Leveraging Spaceflight to Advance Cardiovascular Research on Earth. Circ. Res. 2022, 130, 942–957. [Google Scholar] [CrossRef] [PubMed]
- Hargens, A.R.; Richardson, S. Cardiovascular adaptations, fluid shifts, and countermeasures related to space flight. Respir. Physiol. Neurobiol. 2009, 169 (Suppl. S1), S30–S33. [Google Scholar] [CrossRef]
- Garrett-Bakelman, F.E.; Darshi, M.; Green, S.J.; Gur, R.C.; Lin, L.; Macias, B.R.; McKenna, M.J.; Meydan, C.; Mishra, T.; Nasrini, J.; et al. The NASA Twins Study: A multidimensional analysis of a year-long human spaceflight. Science 2019, 364, eaau8650. [Google Scholar] [CrossRef] [PubMed]
- Da Silveira, W.A.; Fazelinia, H.; Rosenthal, S.B.; Laiakis, E.C.; Kim, M.S.; Meydan, C.; Kidane, Y.; Rathi, K.S.; Smith, S.M.; Stear, B.; et al. Comprehensive Multi-omics Analysis Reveals Mitochondrial Stress as a Central Biological Hub for Spaceflight Impact. Cell 2020, 183, 1185–1201.e1120. [Google Scholar] [CrossRef]
- Wnorowski, A.; Sharma, A.; Chen, H.; Wu, H.; Shao, N.Y.; Sayed, N.; Liu, C.; Countryman, S.; Stodieck, L.S.; Rubins, K.H.; et al. Effects of Spaceflight on Human Induced Pluripotent Stem Cell-Derived Cardiomyocyte Structure and Function. Stem Cell Rep. 2019, 13, 960–969. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Tahimic, C.G.T.; Almeida, E.A.C.; Globus, R.K. Spaceflight Modulates the Expression of Key Oxidative Stress and Cell Cycle Related Genes in Heart. Int. J. Mol. Sci. 2021, 22, 88. [Google Scholar] [CrossRef]
- Walls, S.; Diop, S.; Birse, R.; Elmen, L.; Gan, Z.; Kalvakuri, S.; Pineda, S.; Reddy, C.; Taylor, E.; Trinh, B.; et al. Prolonged Exposure to Microgravity Reduces Cardiac Contractility and Initiates Remodeling in Drosophila. Cell Rep. 2020, 33, 108445. [Google Scholar] [CrossRef]
- Martin, M. CUTADAPT Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet. J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. Stringtie Enables Improved Reconstruction of a Transcriptome from RNA-Seq Reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with deseq2. Genome Biol. 2014, 15, 12. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Krämer, A.; Green, J.; Pollard, J.; Tugendreich, S. Causal Analysis Approaches in Ingenuity Pathway Analysis. Bioinformatics 2013, 30, 523–530. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. String V10: Protein–Protein Interaction Networks, Integrated over the Tree of Life. Nucleic. Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. David: A Web Server for Functional Enrichment Analysis and Functional Annotation of Gene Lists (2021 Update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using David Bioinformatics Resources. Nat. Protoc. 2008, 4, 44–57. [Google Scholar] [CrossRef]
- Deshpande, A.M.; Khalid, O.; Kim, J.J.; Kim, Y.; Lindgren, A.; Clark, A.T.; Wong, D.T.W. Cdk2ap2 Is a Novel Regulator for Self-Renewal of Murine Embryonic Stem Cells. Stem Cells Dev. 2012, 21, 3010–3018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Won, S.-Y.; Park, J.-J.; Shin, E.-Y.; Kim, E.-G. Pak4 Signaling in Health and Disease: Defining the pak4–Creb Axis. Exp. Mol. Med. 2019, 51, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, H.-J.; Dimova, N.V.; Tan, M.-K.M.; Sigoillot, F.D.; King, R.W.; Shi, Y. The G2/M Regulator Histone Demethylase PHF8 Is Targeted for Degradation by the Anaphase-Promoting Complex Containing cdc20. Mol. Cell. Biol. 2013, 33, 4166–4180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panchenko, M.V. Structure, Function and Regulation of Jade Family Phd Finger 1 (JADE1). Gene 2016, 589, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, T.; Horie, K.; Hinoi, E.; Hiraiwa, M.; Kato, A.; Maekawa, Y.; Takahashi, A.; Furukawa, S. How Does Spaceflight Affect the Acquired Immune System? NPJ Microgravity 2020, 6, 14. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, J.; Zhang, C.; Wang, P.; Huang, C.; Peng, H. MDM2 Promotes the Proliferation and Inhibits the Apoptosis of Pituitary Adenoma Cells by Directly Interacting with p53. Endokrynol. Pol. 2020, 71, 425–431. [Google Scholar] [CrossRef]
- Córdova-Alarcón, E.; Centeno, F.; Reyes-Esparza, J.; García-Carrancá, A.; Garrido, E. Effects of HRAS Oncogene on Cell Cycle Progression in a Cervical Cancer-Derived Cell Line. Arch. Med. Res. 2005, 36, 311–316. [Google Scholar] [CrossRef]
- Ingber, D.E. Cellular Tensegrity: Defining New Rules of Biological Design That Govern the Cytoskeleton. J. Cell Sci. 1993, 104, 613–627. [Google Scholar] [CrossRef]
- Wu, X.T.; Yang, X.; Tian, R.; Li, Y.H.; Wang, C.Y.; Fan, Y.B.; Sun, L.W. Cells Respond to Space Microgravity through Cytoskeleton Reorganization. FASEB J. 2022, 36, e22114. [Google Scholar] [CrossRef]
- Tian, J.; Pecaut, M.J.; Slater, J.M.; Gridley, D.S. Spaceflight Modulates Expression of Extracellular Matrix, Adhesion, and Profibrotic Molecules in Mouse Lung. J. Appl. Physiol. 2010, 108, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Ludtka, C.; Silberman, J.; Moore, E.; Allen, J.B. Macrophages in Microgravity: The Impact of Space on Immune Cells. NPJ Microgravity 2021, 7, 13. [Google Scholar] [CrossRef]
- Sun, Y.; Kuang, Y.; Zuo, Z. The Emerging Role of Macrophages in Immune System Dysfunction under Real and Simulated Microgravity Conditions. Int. J. Mol. Sci. 2021, 22, 2333. [Google Scholar] [CrossRef]
- Thiel, C.S.; Tauber, S.; Lauber, B.; Polzer, J.; Seebacher, C.; Uhl, R.; Neelam, S.; Zhang, Y.; Levine, H.; Ullrich, O. Rapid Morphological and Cytoskeletal Response to Microgravity in Human Primary Macrophages. Int. J. Mol. Sci. 2019, 20, 2402. [Google Scholar] [CrossRef] [Green Version]
- Goldsmith, Z.G.; Dhanasekaran, D.N. G Protein Regulation of MAPK Networks. Oncogene 2007, 26, 3122–3142. [Google Scholar] [CrossRef] [Green Version]
- Ravingerova, T.; Barancik, M.; Strniskova, M. Mitogen-Activated Protein Kinases: A New Therapeutic Target in Cardiac Pathology. Mol. Cell. Biochem. 2003, 247, 127–138. [Google Scholar] [CrossRef]
- Kumei, Y.; Shimokawa, H.; Ohya, K.; Katano, H.; Akiyama, H.; Hirano, M.; Morita, S. Small GTPase Ras and Rho Expression in Rat Osteoblasts during Spaceflight. Ann. N. Y. Acad. Sci. 2007, 1095, 292–299. [Google Scholar] [CrossRef]
- Rezatabar, S.; Karimian, A.; Rameshknia, V.; Parsian, H.; Majidinia, M.; Kopi, T.A.; Bishayee, A.; Sadeghinia, A.; Yousefi, M.; Monirialamdari, M.; et al. Ras/MAPK Signaling Functions in Oxidative Stress, DNA Damage Response and Cancer Progression. J. Cell. Physiol. 2019, 234, 14951–14965. [Google Scholar] [CrossRef]
- Wise, K.C.; Manna, S.K.; Yamauchi, K.O.; Ramesh, V.; Wilson, B.L.; Thomas, R.L.; Sarkar, S.; Kulkarni, A.D.; Pellis, N.R.; Ramesh, G. Activation of Nuclear Transcription Factor–ΚB in Mouse Brain Induced by a Simulated Microgravity Environment. Vitr. Cell. Dev. Biol. Anim. 2005, 41, 118–123. [Google Scholar] [CrossRef]
- Toker, A.; Cantley, L.C. Signalling through the lipid products of phosphoinositide-3-OH kinase. Nature 1997, 387, 673–676. [Google Scholar] [CrossRef]
- Hemmings, B.A.; Restuccia, D.F. PI3K-PKB/Akt pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a011189. [Google Scholar] [CrossRef] [Green Version]
- Qin, W.; Cao, L.; Massey, I.Y. Role of PI3K/Akt signaling pathway in cardiac fibrosis. Mol. Cell Biochem. 2021, 476, 4045–4059. [Google Scholar] [CrossRef] [PubMed]
- Brand, C.S.; Lighthouse, J.K.; Trembley, M.A. Protective transcriptional mechanisms in cardiomyocytes and cardiac fibroblasts. J. Mol. Cell Cardiol. 2019, 132, 1–12. [Google Scholar] [CrossRef] [PubMed]
- McMullen, J.R.; Amirahmadi, F.; Woodcock, E.A.; Schinke-Braun, M.; Bouwman, R.D.; Hewitt, K.A.; Mollica, J.P.; Zhang, L.; Zhang, Y.; Shioi, T.; et al. Protective effects of exercise and phosphoinositide 3-kinase(p110alpha) signaling in dilated and hypertrophic cardiomyopathy. Proc. Natl. Acad. Sci. USA 2007, 104, 612–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemi, O.J.; Ceci, M.; Wisloff, U.; Grimaldi, S.; Gallo, P.; Smith, G.L.; Condorelli, G.; Ellingsen, O. Activation or inactivation of cardiac Akt/mTOR signaling diverges physiological from pathological hypertrophy. J. Cell Physiol. 2008, 214, 316–321. [Google Scholar] [CrossRef]
- Walkowski, B.; Kleibert, M.; Majka, M.; Wojciechowska, M. Insight into the Role of the PI3K/Akt Pathway in Ischemic Injury and Post-Infarct Left Ventricular Remodeling in Normal and Diabetic Heart. Cells 2022, 11, 1553. [Google Scholar] [CrossRef]
- McMullen, J.R.; Shioi, T.; Huang, W.Y.; Zhang, L.; Tarnavski, O.; Bisping, E.; Schinke, M.; Kong, S.; Sherwood, M.C.; Brown, J.; et al. The insulin-like growth factor 1 receptor induces physiological heart growth via the phosphoinositide 3-kinase(p110alpha) pathway. J. Biol. Chem. 2004, 279, 4782–4793. [Google Scholar] [CrossRef] [Green Version]
- Kumei, Y.; Morita, S.; Nakamura, H.; Akiyama, H.; Katano, H.; Shimokawa, H.; Ohya, K. Platelet-activating factor receptor signals in rat osteoblasts during spaceflight. Ann. N. Y. Acad. Sci. 2004, 1030, 116–120. [Google Scholar] [CrossRef]
- Acharya, A.; Nemade, H.; Papadopoulos, S.; Hescheler, J.; Neumaier, F.; Schneider, T.; Rajendra Prasad, K.; Khan, K.; Hemmersbach, R.; Gusmao, E.G.; et al. Microgravity-induced stress mechanisms in human stem cell-derived cardiomyocytes. iScience 2022, 25, 104577. [Google Scholar] [CrossRef]
- Wang, M.; Wang, Y.; Weil, B.; Abarbanell, A.; Herrmann, J.; Tan, J.; Kelly, M.; Meldrum, D.R. Estrogen receptor beta mediates increased activation of PI3K/Akt signaling and improved myocardial function in female hearts following acute ischemia. Am. J. Physiol. Regul Integr. Comp. Physiol. 2009, 296, R972–R978. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.P.; Tran, P.H.; Kim, K.-S.; Yang, S.-G. The Effects of Real and Simulated Microgravity on Cellular Mitochondrial Function. NPJ Microgravity 2021, 7, 44. [Google Scholar] [CrossRef]
- Zhao, R.Z.; Jiang, S.; Zhang, L.; Yu, Z.B. Mitochondrial Electron Transport Chain, Ros Generation and Uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.P.; Shin, S.; Shin, K.-J.; Tran, P.H.; Park, H.; De Tran, Q.; No, M.-H.; Sun, J.S.; Kim, K.W.; Kwak, H.-B.; et al. Protective Effect of TPP-Niacin on Microgravity-Induced Oxidative Stress and Mitochondrial Dysfunction of Retinal Epithelial Cells. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2023, 1870, 119384. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, H.; Zhou, J.; Shao, Q. Glutathione Peroxidase GPX1 and Its Dichotomous Roles in Cancer. Cancers 2022, 14, 2560. [Google Scholar] [CrossRef]
- Mathien, S.; Tesnière, C.; Meloche, S. Regulation of Mitogen-Activated Protein Kinase Signaling Pathways by the Ubiquitin-Proteasome System and Its Pharmacological Potential. Pharmacol. Rev. 2021, 73, 1434–1467. [Google Scholar] [CrossRef]
- Kriegenburg, F.; Poulsen, E.G.; Koch, A.; Krüger, E.; Hartmann-Petersen, R. Redox Control of the Ubiquitin-Proteasome System: From Molecular Mechanisms to Functional Significance. Antioxid. Redox Signal. 2011, 15, 2265–2299. [Google Scholar] [CrossRef]
- Mao, X.W.; Nishiyama, N.C.; Byrum, S.D.; Stanbouly, S.; Jones, T.; Holley, J.; Sridharan, V.; Boerma, M.; Tackett, A.J.; Willey, J.S.; et al. Spaceflight Induces Oxidative Damage to Blood-Brain Barrier Integrity in a Mouse Model. FASEB J. 2020, 34, 15516–15530. [Google Scholar] [CrossRef]
- Morabito, C.; Guarnieri, S.; Cucina, A.; Bizzarri, M.; Mariggiò, M.A. Antioxidant Strategy to Prevent Simulated Microgravity-Induced Effects on Bone Osteoblasts. Int. J. Mol. Sci. 2020, 21, 3638. [Google Scholar] [CrossRef]
- Kumari, R.; Jat, P. Mechanisms of Cellular Senescence: Cell Cycle Arrest and Senescence Associated Secretory Phenotype. Front. Cell Dev. Biol. 2021, 9, 645593. [Google Scholar] [CrossRef]
- Barros, P.R.; Costa, T.J.; Akamine, E.H.; Tostes, R.C. Vascular Aging in Rodent Models: Contrasting Mechanisms Driving the Female and Male Vascular Senescence. Front. Aging 2021, 2, 43. [Google Scholar] [CrossRef]
- Mark, S.; Scott, G.B.I.; Donoviel, D.B.; Leveton, L.B.; Mahoney, E.; Charles, J.B.; Siegel, B. The Impact of Sex and Gender on Adaptation to Space: Executive Summary. J. Women’s Health 2014, 23, 941–947. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | Fold Change |
|---|---|---|
| Cat | Catalase | 1.13 |
| Fancc | Fanconi anemia, complementation group C | 0.82 |
| Gpx1 | Glutathione Peroxidase 1 | 0.78 |
| Gpx3 | Glutathione Peroxidase 3 | 0.27 |
| Gsr | Glutathione reductase | 1.00 |
| Gstp1 | Glutathione S-transferase, pi 1 | 0.83 |
| Ncf2 | Neutrophil cytosolic factor 2 | 0.78 |
| Nox1 | NADPH oxidase 1 | 0.65 |
| Nox4 | NADPH oxidase 4 | 0.49 |
| Prdx1 | Peroxiredoxin 1 | 1.06 |
| Prdx6 | Peroxiredoxin 6 | 1.05 |
| Prnp | Prion protein | 1.02 |
| Ptgs2 | Prostaglandin-endoperoxide synthase 2 | 0.22 |
| Slc41a3 | Solute carrier family 41, member 3 | 1.62 |
| Sod1 | Superoxide dismutase 1 | 0.87 |
| Sod2 | Superoxide dismutase 2 | 1.05 |
| Sod3 | Superoxide dismutase 3 | 0.78 |
| Tpo | Thyroid peroxidase | 1.00 |
| Txnip | Thioredoxin interacting protein | 0.88 |
| Txnrd3 | Thioredoxin reductase 3 | 0.98 |
| Xpa | Xeroderma pigmentosum, complementation group A | 0.72 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veliz, A.L.; Mamoun, L.; Hughes, L.; Vega, R.; Holmes, B.; Monteon, A.; Bray, J.; Pecaut, M.J.; Kearns-Jonker, M. Transcriptomic Effects on the Mouse Heart Following 30 Days on the International Space Station. Biomolecules 2023, 13, 371. https://doi.org/10.3390/biom13020371
Veliz AL, Mamoun L, Hughes L, Vega R, Holmes B, Monteon A, Bray J, Pecaut MJ, Kearns-Jonker M. Transcriptomic Effects on the Mouse Heart Following 30 Days on the International Space Station. Biomolecules. 2023; 13(2):371. https://doi.org/10.3390/biom13020371
Chicago/Turabian StyleVeliz, Alicia L., Lana Mamoun, Lorelei Hughes, Richard Vega, Bailey Holmes, Andrea Monteon, Jillian Bray, Michael J. Pecaut, and Mary Kearns-Jonker. 2023. "Transcriptomic Effects on the Mouse Heart Following 30 Days on the International Space Station" Biomolecules 13, no. 2: 371. https://doi.org/10.3390/biom13020371