
Cytochrome P450 Surface Domains Prevent the β-Carotene Monohydroxylase CYP97H1 of Euglena gracilis from Acting as a Dihydroxylase
 , , , and
, , , and
Abstract
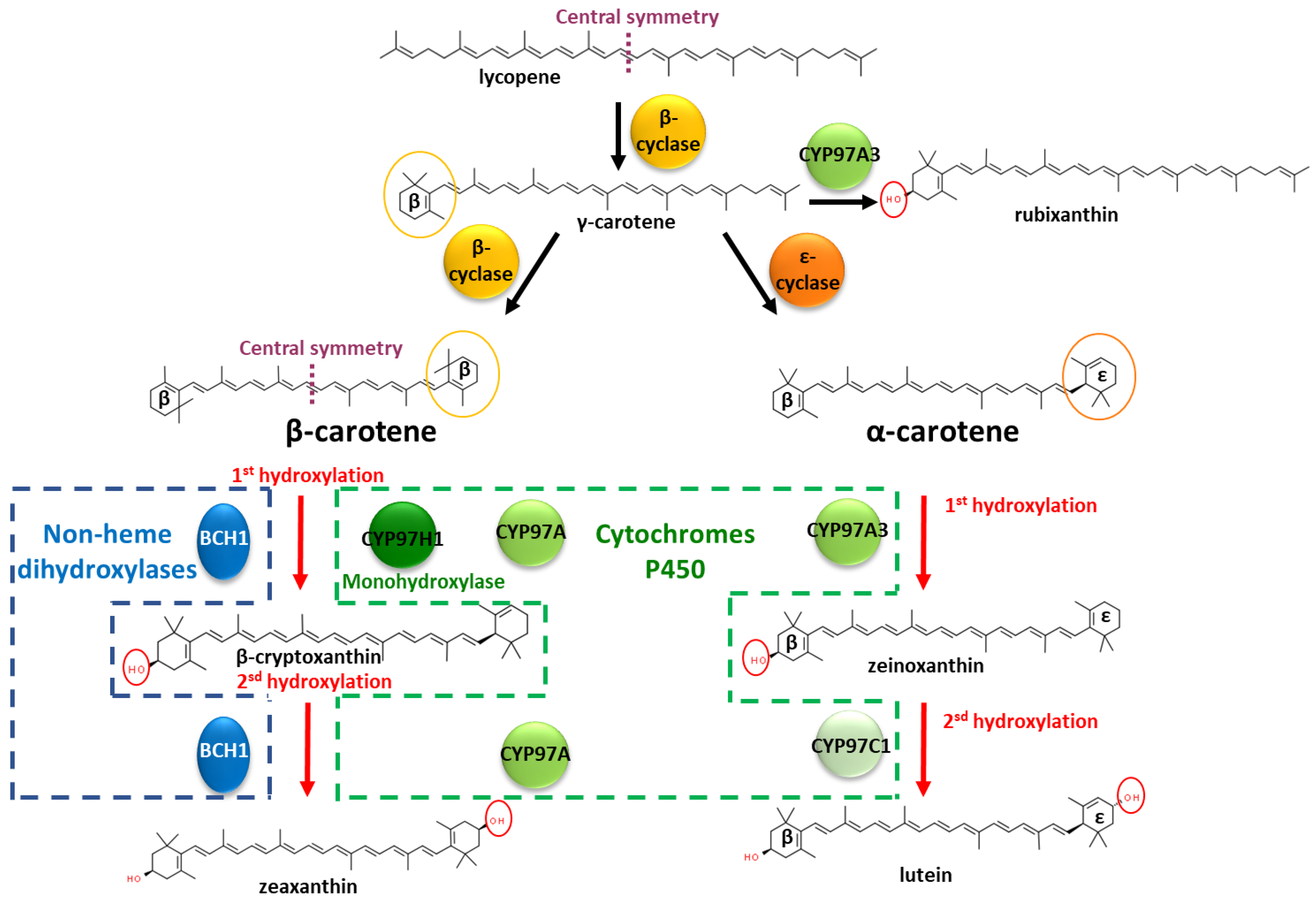
:1. Introduction
2. Materials and Methods
2.1. Strain and Plasmid Construction
2.2. P450 Structure Modelling and Substrate Docking
2.3. Culture of the E. coli Strains
2.4. Extraction of Carotenoids
2.5. Quantification of Carotenoids
3. Results
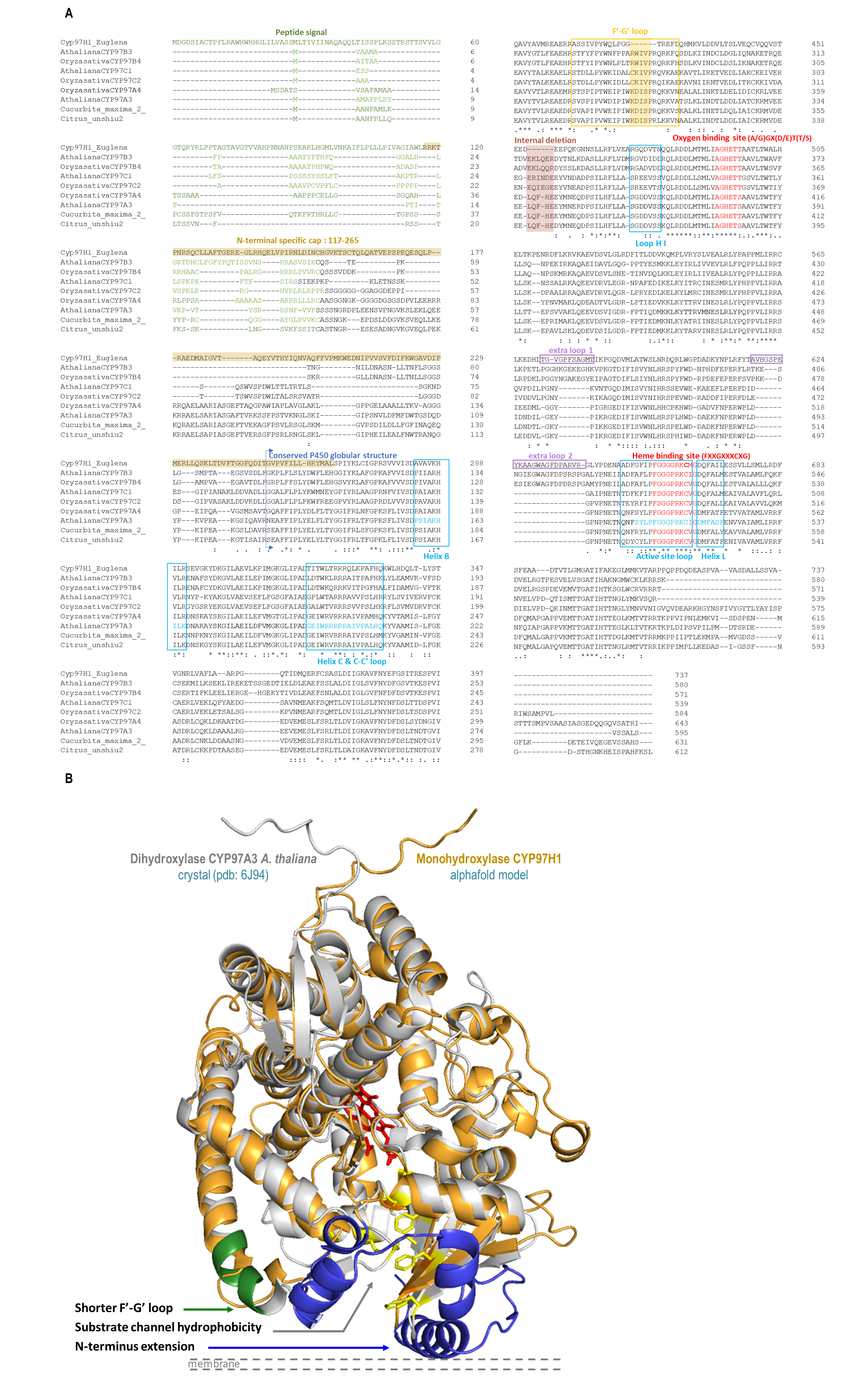
3.1. In Silico Identification of the Structural Specificities of CYP97H1 Compared to CYP97 Members
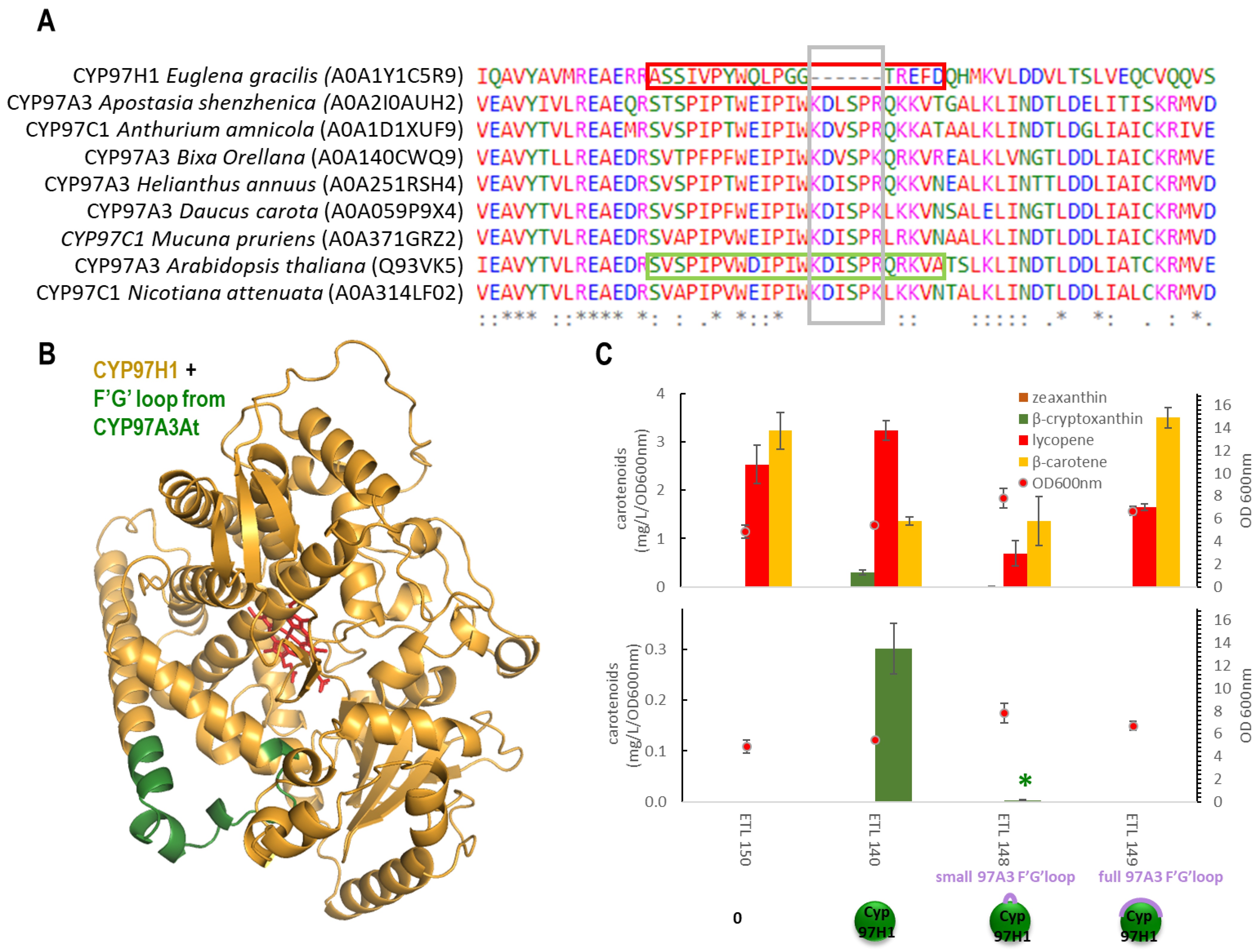
3.2. The F′-G′ Loop of CYP97H1 Is Shorter Compared to the Ones of CYP97 Members
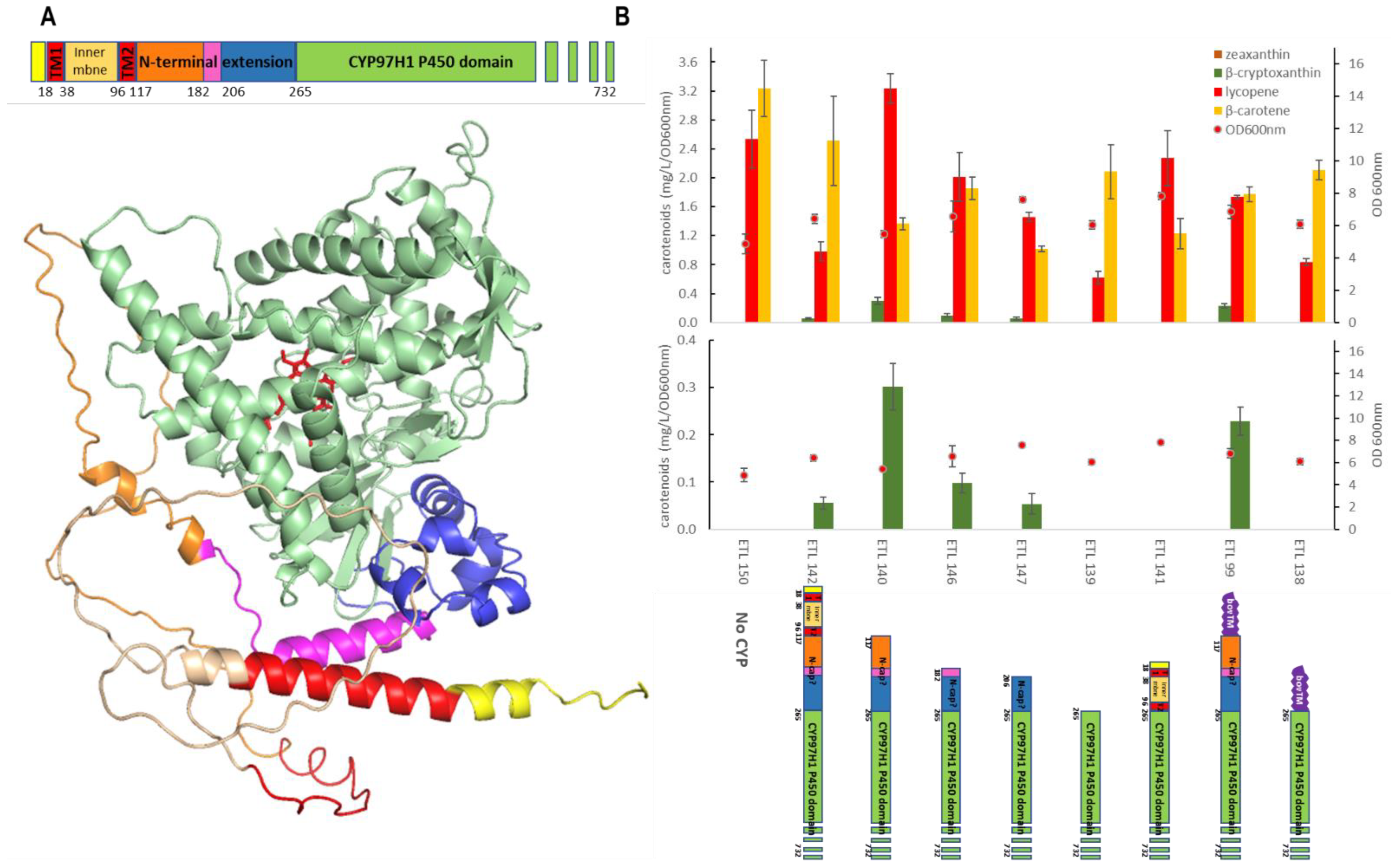
3.3. The N-Terminal Additional Domain Is Required for Full CYP97H1 Activity
3.4. The Substrate Channel Entrance Harbours a Hydrophobic Patch Involved in the Monoterminal Activity of CYP97H1
4. Discussion
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biosynthesis of Carotenoids and Apocarotenoids by Microorganisms and Their Industrial Potential|IntechOpen. Available online: https://www.intechopen.com/chapters/62140 (accessed on 7 November 2022).
- Avalos, J.; Carmen Limón, M. Biological roles of fungal carotenoids. Curr. Genet. 2015, 61, 309–324. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids as natural functional pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandmann, G. Diversity and origin of carotenoid biosynthesis: Its history of coevolution towards plant photosynthesis. New Phytol. 2021, 232, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Smith, J.J.; Tian, L.; Dellapenna, D. The evolution and function of carotenoid hydroxylases in Arabidopsis. Plant Cell Physiol. 2009, 50, 463–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, C.C.; Nelson, D.R.; Møller, B.L.; Werck-Reichhart, D. Plant cytochrome P450 plasticity and evolution. Mol. Plant 2021, 14, 1244–1265. [Google Scholar] [CrossRef]
- Nelson, D.; Werck-Reichhart, D. A P450-centric view of plant evolution: P450-centric evolution. Plant J. 2011, 66, 194–211. [Google Scholar] [CrossRef]
- Sun, Z.; Gantt, E.; Cunningham, F.X. Cloning and Functional Analysis of the β-Carotene Hydroxylase of Arabidopsis thaliana. J. Biol. Chem. 1996, 271, 24349–24352. [Google Scholar] [CrossRef] [Green Version]
- Arango, J.; Jourdan, M.; Geoffriau, E.; Beyer, P.; Welsch, R. Carotene Hydroxylase Activity Determines the Levels of Both α-Carotene and Total Carotenoids in Orange Carrots. Plant Cell 2014, 26, 2223–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, G.; Guo, Q.; Wang, J.; Zhao, S.; He, Y.; Liu, L. Structural basis for plant lutein biosynthesis from α-carotene. Proc. Natl. Acad. Sci. USA. 2020, 117, 14150–14157. [Google Scholar] [CrossRef]
- Kim, J.; DellaPenna, D. Defining the primary route for lutein synthesis in plants: The role of Arabidopsis carotenoid β-ring hydroxylase CYP97A3. Proc. Natl. Acad. Sci. USA 2006, 103, 3474–3479. [Google Scholar] [CrossRef]
- Quinlan, R.F.; Jaradat, T.T.; Wurtzel, E.T. Escherichia coli as a platform for functional expression of plant P450 carotene hydroxylases. Arch. Biochem. Biophys. 2007, 458, 146–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamaki, S.; Kato, S.; Shinomura, T.; Ishikawa, T.; Imaishi, H. Physiological role of β-carotene monohydroxylase (CYP97H1) in carotenoid biosynthesis in Euglena gracilis. Plant Sci. 2019, 278, 80–87. [Google Scholar] [CrossRef]
- Zhang, C.; Seow, V.Y.; Chen, X.; Too, H.-P. Multidimensional heuristic process for high-yield production of astaxanthin and fragrance molecules in Escherichia coli. Nat. Commun. 2018, 9, 1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Chen, X.; Too, H.-P. Production of Carotenoids and Apocarotenoids. U.S. Patent Application 16/488,504, 5 December 2019. Available online: https://patents.google.com/patent/US20190367928A1/en (accessed on 18 September 2020).
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Ciccarelli, F.D.; Doerks, T.; von Mering, C.; Creevey, C.J.; Snel, B.; Bork, P. Toward automatic reconstruction of a highly resolved tree of life. Science 2006, 311, 1283–1287. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Land, H.; Humble, M.S. YASARA: A Tool to Obtain Structural Guidance in Biocatalytic Investigations. Methods Mol. Biol. 2018, 1685, 43–67. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef] [PubMed]
- PyMOL | pymol.org. Available online: https://pymol.org/2/ (accessed on 12 April 2022).
- De Ritter, E.; Purcell, A.E. Carotenoid Analytical Methods. In Carotenoids as Colorants and Vitamin A Precursors; Elsevier: Amsterdam, The Netherlands, 1981; pp. 815–923. ISBN 978-0-12-082850-0. [Google Scholar]
- Rivera, S.M.; Christou, P.; Canela-Garayoa, R. Identification of carotenoids using mass spectrometry: IDENTIFICATION OF CAROTENOIDS. Mass Spec. Rev. 2014, 33, 353–372. [Google Scholar] [CrossRef] [Green Version]
- Lautier, T.; Urban, P.; Loeper, J.; Jezequel, L.; Pompon, D.; Truan, G. Ordered chimerogenesis applied to CYP2B P450 enzymes. Biochim. Biophys. Acta 2016, 1860, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.Y.; Eachus, R.A.; Trieu, W.; Ro, D.-K.; Keasling, J.D. Engineering Escherichia coli for production of functionalized terpenoids using plant P450s. Nat. Chem. Biol. 2007, 3, 274–277. [Google Scholar] [CrossRef]
- Ortiz de Montellano, P.R. (Ed.) Cytochrome P450: Structure, Mechanism, and Biochemistry, 3rd ed.; Springer: New York, NY, USA, 2005; ISBN 978-0-387-27447-8. [Google Scholar]
- Wang, A.; Stout, C.D.; Zhang, Q.; Johnson, E.F. Contributions of Ionic Interactions and Protein Dynamics to Cytochrome P450 2D6 (CYP2D6) Substrate and Inhibitor Binding. J. Biol. Chem. 2015, 290, 5092–5104. [Google Scholar] [CrossRef] [Green Version]
- Behera, R.K.; Mazumdar, S. Roles of two surface residues near the access channel in the substrate recognition by cytochrome P450cam. Biophys. Chem. 2008, 135, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Urban, P.; Lautier, T.; Pompon, D.; Truan, G. Ligand Access Channels in Cytochrome P450 Enzymes: A Review. Int. J. Mol. Sci. 2018, 19, 1617. [Google Scholar] [CrossRef] [Green Version]
- Laursen, T.; Borch, J.; Knudsen, C.; Bavishi, K.; Torta, F.; Martens, H.J.; Silvestro, D.; Hatzakis, N.S.; Wenk, M.R.; Dafforn, T.R.; et al. Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum. Science 2016, 354, 890–893. [Google Scholar] [CrossRef]
- Urban, P.; Pompon, D. Confrontation of AlphaFold models with experimental structures enlightens conformational dynamics supporting CYP102A1 functions. Sci. Rep. 2022, 12, 15982. [Google Scholar] [CrossRef]
- Mukherjee, G.; Nandekar, P.P.; Wade, R.C. An electron transfer competent structural ensemble of membrane-bound cytochrome P450 1A1 and cytochrome P450 oxidoreductase. Commun. Biol. 2021, 4, 55. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
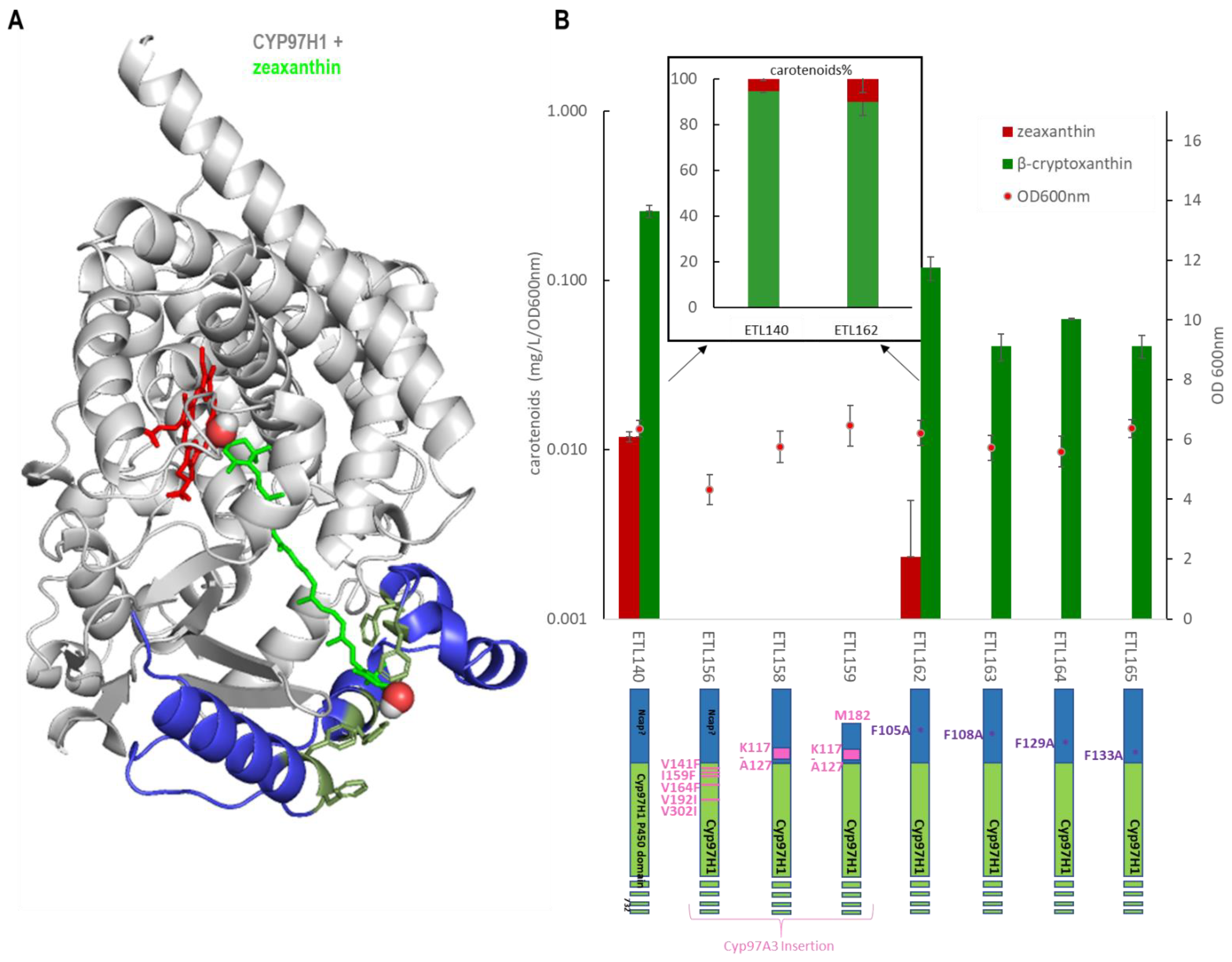{kind=link}
| Strain | “Hydroxylase” Plasmid | Specific CYP Sequence | Source |
|---|---|---|---|
| ETL99 | pTL45 | bov-CYP97H1 | this work |
| ETL138 | pTL70 | bov-capless-CYP97H1 | this work |
| ETL139 | pTL71 | supertruncated-CYP97H1 | this work |
| ETL140 | pTL72 | tCYP97H1 | this work |
| ETL141 | pTL73 | capless CYP97H1 | this work |
| ETL142 | pTL68 | full-length CYP97H1 | this work |
| ETL146 | pTL79 | M182-CYP97H1 | this work |
| ETL147 | pTL80 | M206-CYP97H1 | this work |
| ETL148 | pTL82 | tCYP97H1-6At: 6 F′-G′ loop residues from CYP97A3At | this work |
| ETL149 | pTL81 | tCYP97H1-fullAt: full F′-G′ loop from CYP97A3At | this work |
| ETL150 | p15A-amp-CrtY | no CYP | Zhang et al., 2018, [14] |
| ETL154 | pTL83 | tCYP97A3At | this work |
| ETL155 | pTL84 | Nterm97H1-CYP97A3At | this work |
| ETL156 | pTL88 | tCYP97H1 with substrate channel from CYP97AAt | this work |
| ETL158 | pTL95 | K117-A127At-CYP97H1 | this work |
| ETL159 | pTL96 | M182-K117-A127At-CYP97H1 | this work |
| ETL160 | pTL98 | MBP-CYP97A3At | this work |
| ETL162 | pTL91 | tCYP97H1 F105A | this work |
| ETL163 | pTL92 | tCYP97H1 F108A | this work |
| ETL164 | pTL93 | tCYP97H1 F129A | this work |
| ETL165 | pTL94 | tCYP97H1 F133A | this work |
| No. | RT (min) | Compound Name | Molecular Formula | Precursor Ion (m/z) | Key Fragments (m/z) |
|---|---|---|---|---|---|
| 1 | 3.45 | zeaxanthin | C40H56O2 | 568.4180 | 476.3476 |
| 2 | 4.99 | rubixanthin | C40H56O | 552.4341 | 460.3706, 391.2997 |
| 3 | 5.21 | β-cryptoxanthin | C40H56O | 552.4347 | 460.3705 |
| 4 | 8.10 | lycopene | C40H56 | 536.4164 | 467.3490, 444.3568 |
| 5 | 9.45 | β-carotene | C40H56 | 536.4137 | 444.3602 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lautier, T.; Smith, D.J.; Yang, L.K.; Chen, X.; Zhang, C.; Truan, G.; Lindley, N.D. Cytochrome P450 Surface Domains Prevent the β-Carotene Monohydroxylase CYP97H1 of Euglena gracilis from Acting as a Dihydroxylase. Biomolecules 2023, 13, 366. https://doi.org/10.3390/biom13020366
Lautier T, Smith DJ, Yang LK, Chen X, Zhang C, Truan G, Lindley ND. Cytochrome P450 Surface Domains Prevent the β-Carotene Monohydroxylase CYP97H1 of Euglena gracilis from Acting as a Dihydroxylase. Biomolecules. 2023; 13(2):366. https://doi.org/10.3390/biom13020366
Chicago/Turabian StyleLautier, Thomas, Derek J. Smith, Lay Kien Yang, Xixian Chen, Congqiang Zhang, Gilles Truan, and Nic D Lindley. 2023. "Cytochrome P450 Surface Domains Prevent the β-Carotene Monohydroxylase CYP97H1 of Euglena gracilis from Acting as a Dihydroxylase" Biomolecules 13, no. 2: 366. https://doi.org/10.3390/biom13020366