
Accumulation of Fat Not Responsible for Femoral Head Necrosis, Revealed by Single-Cell RNA Sequencing: A Preliminary Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Obtaining Bone Marrow Samples of Femoral Head Necrosis
2.2. scRNA-seq Data Alignment and Sample Aggregation
2.3. Dimensionality Reduction and Clustering Analysis
2.4. Cell Type Annotation and Cluster Marker Identification
2.5. Gene Functional Annotation
3. Results
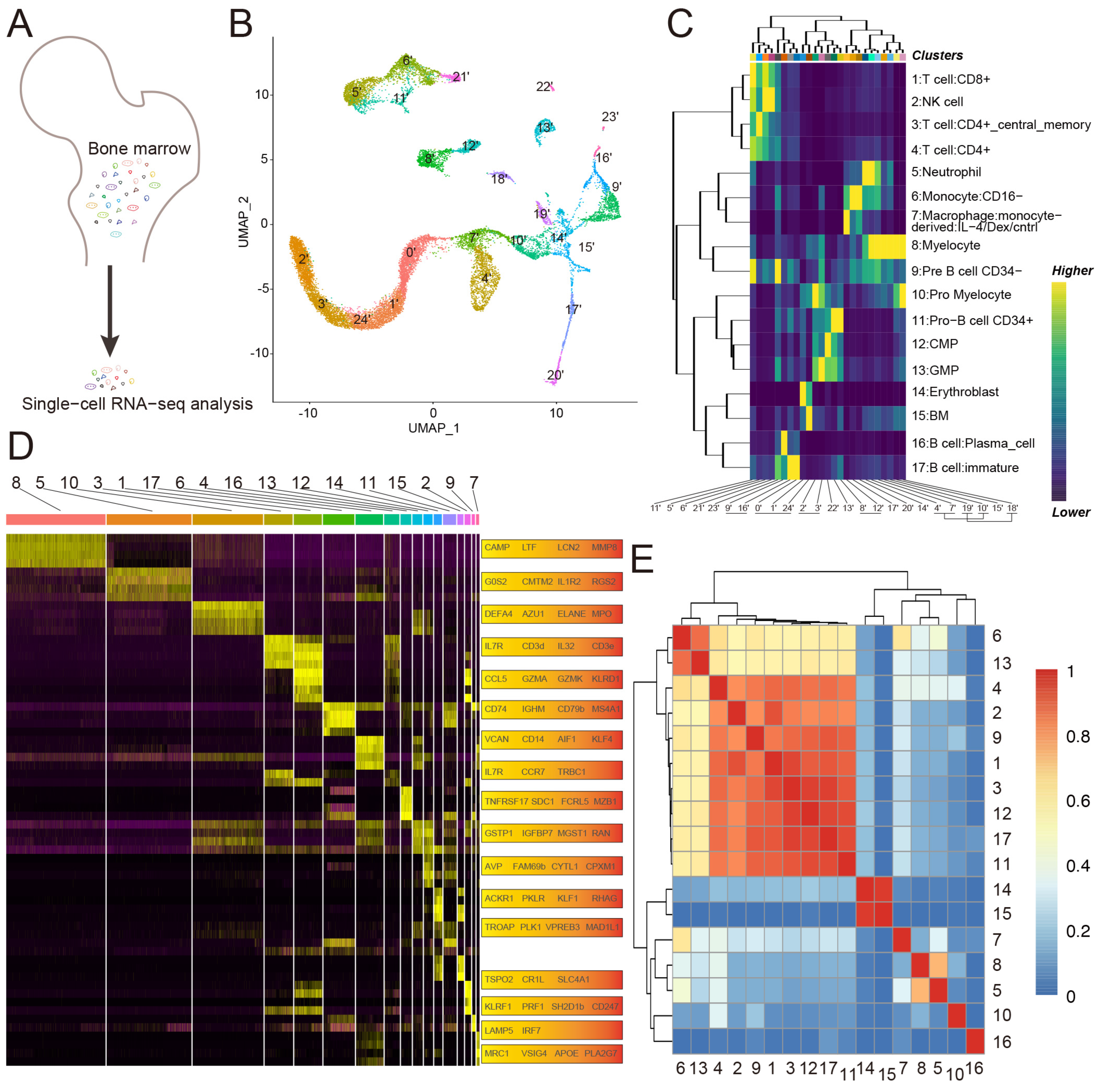
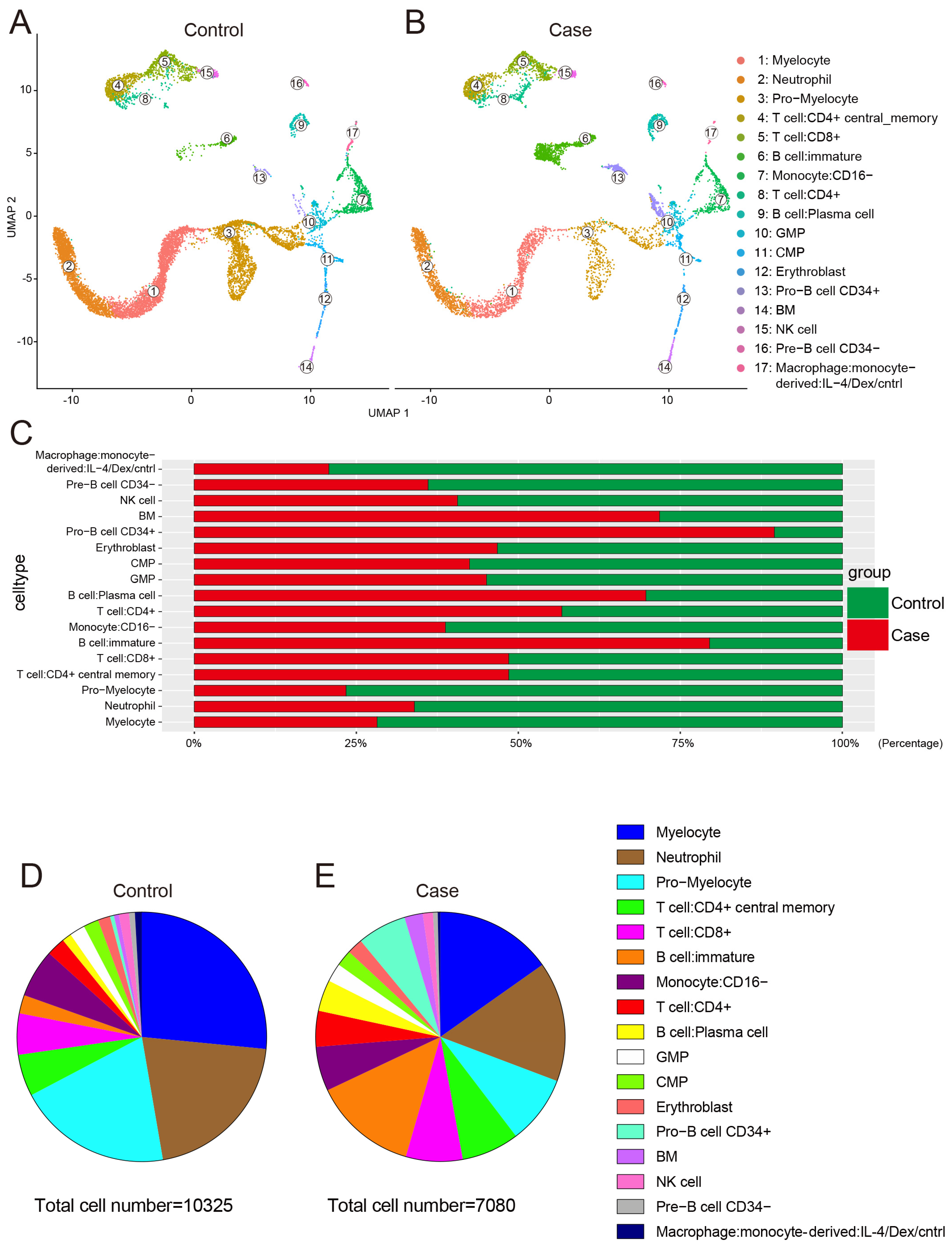
3.1. Single-Cell Profiling of Human Bone Marrow
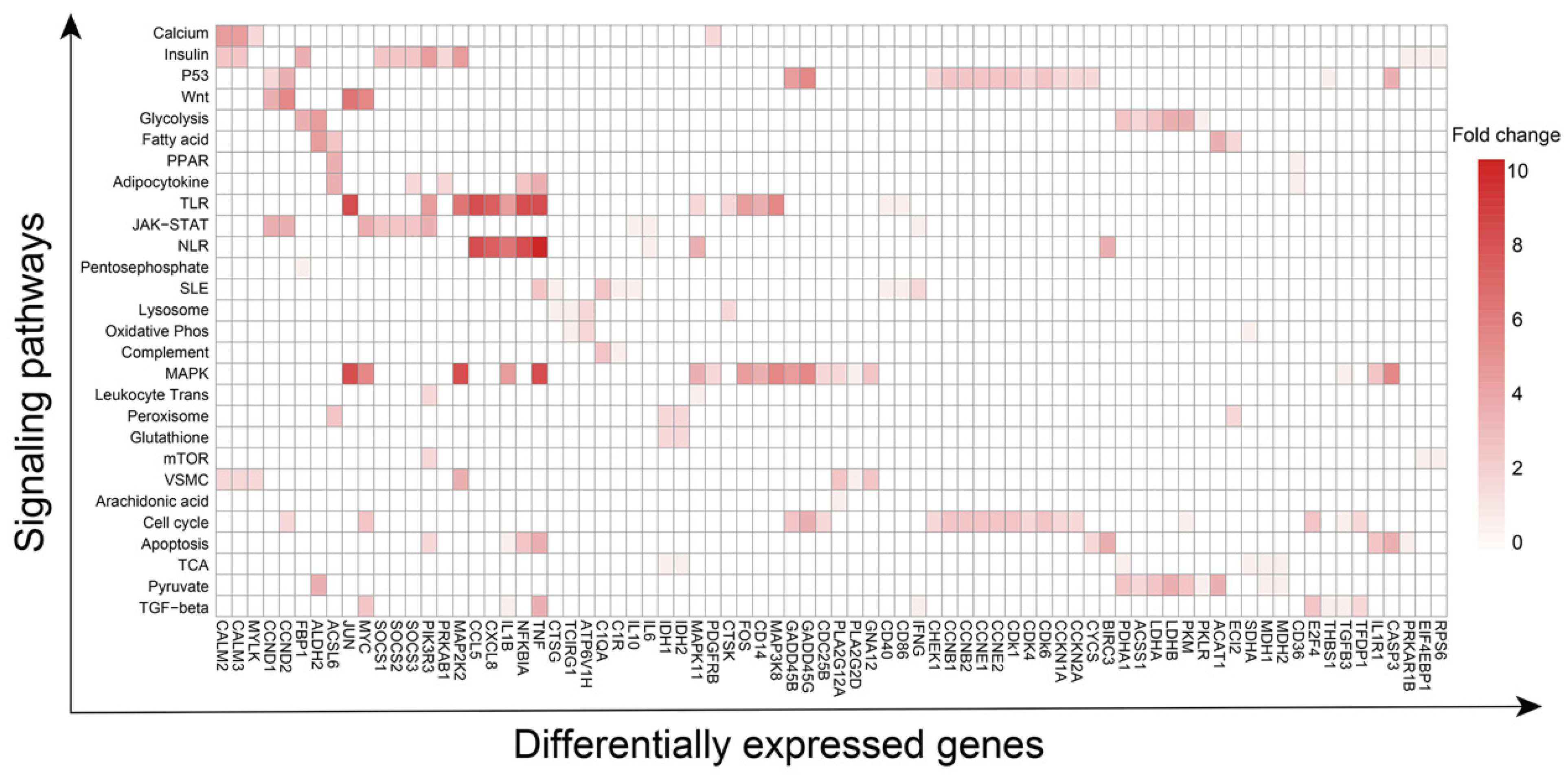
3.2. Expression of Glucocorticoid Therapy-Related Genes in Various Cell Types
3.3. Pathways and Corresponding Key Genes in Various Cell Types
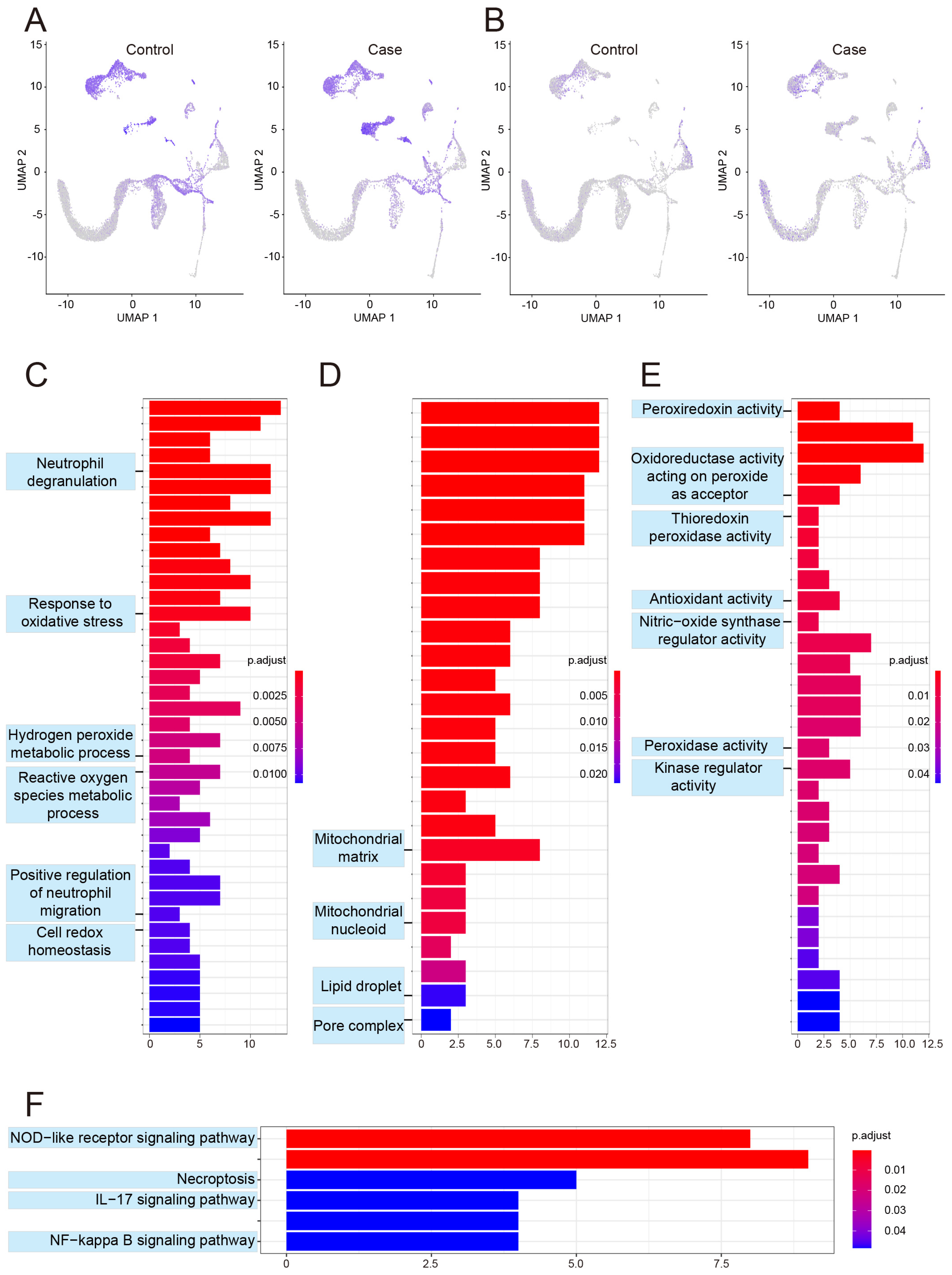
3.4. Changes in Metabolic Profiling in the Bone Marrow
3.5. Changes in Inflammatory-Response Profiling in Bone Marrow
3.6. Potential Effects of SLE Combined with Glucocorticoids on Blood Vessel and Bone Metabolism
3.7. Potential Effects of SLE Combined with Glucocorticoids on Cell Proliferation and Apoptosis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, M.; Zhao, Y.; Zhang, Z.; Huang, C.; Liu, Y.; Gu, J.; Zhang, X.; Xu, H.; Li, X.; Zeng, X.; et al. Chinese guidelines for the diagnosis and treatment of systemic lupus erythematosus. Rheumatol. Immunol. Res. 2020, 59, 172–185. [Google Scholar] [CrossRef]
- Becht, E.; McInnes, L.; Healy, J.; Dutertre, C.-A.; Kwok, I.W.H.; Ng, L.G.; Ginhoux, F.; Newell, E.W. Dimensionality reduction for visualizing single-cell data using UMAP. Nat. Biotechnol. 2018, 37, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Bertsias, G.; A Ioannidis, J.P.; Boletis, J.N.; Bombardieri, S.; Cervera, R.; Dostal, C.; Font, J.; Gilboe, I.M.; Houssiau, F.; Huizinga, T.; et al. EULAR recommendations for the management of systemic lupus erythematosus. Report of a Task Force of the EULAR Standing Committee for International Clinical Studies Including Therapeutics. Ann. Rheum. Dis. 2008, 67, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Boskey, A.L.; Raggio, C.L.; Bullough, P.G.; Kinnett, J.G. Changes in the bone tissue lipids in persons with steroid- and alcohol-induced osteonecrosis. Clin. Orthop. Relat. Res. 1983, 172, 289–295. [Google Scholar] [CrossRef]
- Calder, J.D.F.; Buttery, L.; Revell, P.A.; Pearse, M.; Polak, J.M. Apoptosis--a significant cause of bone cell death in osteonecrosis of the femoral head. J. Bone Jt. Surg. 2004, 86, 1209–1213. [Google Scholar] [CrossRef] [Green Version]
- Cárcamo-Orive, I.; Gaztelumendi, A.; Delgado, J.; Tejados, N.; Dorronsoro, A.; Fernández-Rueda, J.; Pennington, D.J.; Trigueros, C. Regulation of human bone marrow stromal cell proliferation and differentiation capacity by glucocorticoid receptor and AP-1 crosstalk. J. Bone Miner. Res. 2010, 25, 2115–2125. [Google Scholar] [CrossRef] [Green Version]
- Chalayer, E.; Costedoat-Chalumeau, N.; Beyne-Rauzy, O.; Ninet, J.; Durupt, S.; Tebib, J.; Asli, B.; Lambotte, O.; Ffrench, M.; Vasselon, C.; et al. Bone marrow involvement in systemic lupus erythematosus. Qjm Int. J. Med. 2017, 110, 701–711. [Google Scholar] [CrossRef] [Green Version]
- Chalayer, É.; Ffrench, M.; Cathébras, P. Aplastic anemia as a feature of systemic lupus erythematosus: A case report and literature review. Rheumatol. Int. 2015, 35, 1073–1082. [Google Scholar] [CrossRef]
- Chang, C.; Greenspan, A.; Gershwin, M.E. The pathogenesis, diagnosis and clinical manifestations of steroid-induced osteonecrosis. J. Autoimmun. 2020, 110, 102460. [Google Scholar] [CrossRef]
- Chang, M.K.; Raggatt, L.-J.; Alexander, K.A.; Kuliwaba, J.S.; Fazzalari, N.L.; Schroder, K.; Maylin, E.R.; Ripoll, V.M.; Hume, D.A.; Pettit, A.R. Osteal tissue macrophages are intercalated throughout human and mouse bone lining tissues and regulate osteoblast function in vitro and in vivo. J. Immunol. 2008, 181, 1232–1244. [Google Scholar] [CrossRef]
- Chmielnicka, M.; Woźniacka, A.; Torzecka, J.D. The influence of corticosteroid treatment on the OPG/RANK/RANKL pathway and osteocalcin in patients with pemphigus. Adv. Dermatol. Allergol. 2014, 31, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, A.; Huggins, M.; Ahmed, J.; Hashimoto, D.; Lucas, D.; Kunisaki, Y.; Pinho, S.; Leboeuf, M.; Noizat, C.; van Rooijen, N.; et al. CD169+ macrophages provide a niche promoting erythropoiesis under homeostasis and stress. Nat. Med. 2013, 19, 429–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, H.-Y.; Chung, J.-W.; Kim, H.-A.; Yun, J.-M.; Jeon, J.-Y.; Ye, Y.-M.; Kim, S.-H.; Park, H.-S.; Suh, C.-H. Cytokine IL-6 and IL-10 as Biomarkers in Systemic Lupus Erythematosus. J. Clin. Immunol. 2007, 27, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Zhuang, Q.; Lin, J.; Jin, J.; Zhang, K.; Cao, L.; Lin, J.; Yan, S.; Guo, W.; He, W.; et al. Multicentric epidemiologic study on six thousand three hundred and ninety five cases of femoral head osteonecrosis in China. Int. Orthop. 2016, 40, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Culemann, S.; Grüneboom, A.; Nicolás-Ávila, J.Á.; Weidner, D.; Lämmle, K.F.; Rothe, T.; Quintana, J.A.; Kirchner, P.; Krljanac, B.; Eberhardt, M.; et al. Locally renewing resident synovial macrophages provide a protective barrier for the joint. Nature 2019, 572, 670–675. [Google Scholar] [CrossRef] [PubMed]
- de Waal Malefyt, R.; Abrams, J.; Bennett, B.; Figdor, C.G.; De Vries, J.E. Interleukin 10(IL-10) inhibits cytokine synthesis by human monocytes: An autoregulatory role of IL-10 produced by monocytes. J. Exp. Med. 1991, 174, 1209–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degirmenci, U.; Wang, M.; Hu, J. Targeting Aberrant RAS/RAF/MEK/ERK Signaling for Cancer Therapy. Cells 2020, 9, 198. [Google Scholar] [CrossRef] [Green Version]
- Donnarumma, M.; Regis, S.; Tappino, B.; Rosano, C.; Assereto, S.; Corsolini, F.; Di Rocco, M.; Filocamo, M. Molecular analysis and characterization of nine novel CTSK mutations in twelve patients affected by pycnodysostosis. Mutation in brief #961. Hum. Mutat. 2007, 28, 524. [Google Scholar] [CrossRef]
- Dörner, T.; Furie, R. Novel paradigms in systemic lupus erythematosus. Lancet 2019, 393, 2344–2358. [Google Scholar] [CrossRef]
- Dorrity, M.W.; Saunders, L.M.; Queitsch, C.; Fields, S.; Trapnell, C. Dimensionality reduction by UMAP to visualize physical and genetic interactions. Nat. Commun. 2020, 11, 1537. [Google Scholar] [CrossRef]
- Dubois, E.L.; Cozen, L. Avascular (Aseptic) Bone Necrosis Associated with Systemic Lupus Erythematosus. JAMA 1960, 174, 966–971. [Google Scholar] [CrossRef] [PubMed]
- Eggold, J.T.; Rankin, E.B. Erythropoiesis, EPO, macrophages, and bone. Bone 2019, 119, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Faggiano, A.; Pivonello, R.; Spiezia, S.; De Martino, M.C.; Filippella, M.; Di Somma, C.; Lombardi, G.; Colao, A. Cardiovascular Risk Factors and Common Carotid Artery Caliber and Stiffness in Patients with Cushing’s Disease during Active Disease and 1 Year after Disease Remission. J. Clin. Endocrinol. Metab. 2003, 88, 2527–2533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagiani, E.; Bill, R.; Pisarsky, L.; Ivanek, R.; Rüegg, C.; Christofori, G. An immature B cell population from peripheral blood serves as surrogate marker for monitoring tumor angiogenesis and anti-angiogenic therapy in mouse models. Angiogenesis 2015, 18, 327–345. [Google Scholar] [CrossRef] [Green Version]
- Fanouriakis, A.; Kostopoulou, M.; Alunno, A.; Aringer, M.; Bajema, I.; Boletis, J.N.; Cervera, R.; Doria, A.; Gordon, C.; Govoni, M.; et al. 2019 update of the EULAR recommendations for the management of systemic lupus erythematosus. Ann. Rheum. Dis. 2019, 78, 736–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanouriakis, A.; Tziolos, N.; Bertsias, G.; Boumpas, D.T. Update οn the diagnosis and management of systemic lupus erythematosus. Ann. Rheum. Dis. 2021, 80, 14–25. [Google Scholar] [CrossRef]
- Farberov, S.; Meidan, R. Functions and Transcriptional Regulation of Thrombospondins and Their Interrelationship with Fibroblast Growth Factor-2 in Bovine Luteal Cells1. Biol. Reprod. 2014, 91, 58. [Google Scholar] [CrossRef]
- Faustman, D.; Davis, M. TNF receptor 2 pathway: Drug target for autoimmune diseases. Nat. Rev. Drug Discov. 2010, 9, 482–493. [Google Scholar] [CrossRef]
- Fukushima, W.; Fujioka, M.; Kubo, T.; Tamakoshi, A.; Nagai, M.; Hirota, Y. Nationwide Epidemiologic Survey of Idiopathic Osteonecrosis of the Femoral Head. Clin. Orthop. Relat. Res. 2010, 468, 2715–2724. [Google Scholar] [CrossRef] [Green Version]
- Gordon, C.; Amissah-Arthur, M.-B.; Gayed, M.; Brown, S.; Bruce, I.N.; D’Cruz, D.; Empson, B.; Griffiths, B.; Jayne, D.; Khamashta, M.; et al. The British Society for Rheumatology guideline for the management of systemic lupus erythematosus in adults. Rheumatology 2018, 57, e1–e45. [Google Scholar] [CrossRef]
- Gui, C.-P.; Liao, B.; Luo, C.-G.; Chen, Y.-H.; Tan, L.; Tang, Y.-M.; Li, J.-Y.; Hou, Y.; Song, H.-D.; Lin, H.-S.; et al. circCHST15 is a novel prognostic biomarker that promotes clear cell renal cell carcinoma cell proliferation and metastasis through the miR-125a-5p/EIF4EBP1 axis. Mol. Cancer 2021, 20, 169. [Google Scholar] [CrossRef]
- Han, L.; Wang, B.; Wang, R.; Gong, S.; Chen, G.; Xu, W. The shift in the balance between osteoblastogenesis and adipogenesis of mesenchymal stem cells mediated by glucocorticoid receptor. Stem Cell Res. Ther. 2019, 10, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, S.-I.; Komuro, I.; Yamada, M.; Akagawa, K.S. IL-10 Inhibits Granulocyte-Macrophage Colony-Stimulating Factor-Dependent Human Monocyte Survival at the Early Stage of the Culture and Inhibits the Generation of Macrophages. J. Immunol. 2001, 167, 3619–3625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haziot, A.; Ferrero, E.; Köntgen, F.; Hijiya, N.; Yamamoto, S.; Silver, J.; Stewart, C.L.; Goyert, S.M. Resistance to Endotoxin Shock and Reduced Dissemination of Gram-Negative Bacteria in CD14-Deficient Mice. Immunity 1996, 4, 407–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heideveld, E.; Akker, E.V.D. Digesting the role of bone marrow macrophages on hematopoiesis. Immunobiology 2017, 222, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.W.; Bishop, E.; Karchenko, P.V.; Nègre, N.; White, K.P.; Park, P.J. ChIP-chip versus ChIP-seq: Lessons for experimental design and data analysis. BMC Genom. 2011, 12, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosaka, K.; Yang, Y.; Seki, T.; Du, Q.; Jing, X.; He, X.; Wu, J.; Zhang, Y.; Morikawa, H.; Nakamura, M.; et al. Therapeutic paradigm of dual targeting VEGF and PDGF for effectively treating FGF-2 off-target tumors. Nat. Commun. 2020, 11, 3704. [Google Scholar] [CrossRef]
- Ichiseki, T.; Matsumoto, T.; Nishino, M.; Kaneuji, A.; Katsuda, S. Oxidative stress and vascular permeability in steroid-induced osteonecrosis model. J. Orthop. Sci. 2004, 9, 509–515. [Google Scholar] [CrossRef]
- Jin, Y.; Dai, M.-S.; Lu, S.Z.; Xu, Y.; Luo, Z.; Zhao, Y.; Lu, H. 14-3-3γ binds to MDMX that is phosphorylated by UV-activated Chk1, resulting in p53 activation. EMBO J. 2006, 25, 1207–1218. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.P. Epidemiological risk factors for non-traumatic osteonecrosis. Orthopade2000 2000, 29, 0370–0379, (Epidemiologische Risikofaktoren für die nichttraumatische Osteonekrose). [Google Scholar] [CrossRef]
- Jones, L.C.; A Mont, M.; Le, T.B.; Petri, M.; Hungerford, D.S.; Wang, P.; Glueck, C.J. Procoagulants and osteonecrosis. J. Rheumatol. 2003, 30, 783–791. [Google Scholar] [PubMed]
- Kamura, T.; Maenaka, K.; Kotoshiba, S.; Matsumoto, M.; Kohda, D.; Conaway, R.C.; Conaway, J.W.; Nakayama, K.I. VHL-box and SOCS-box domains determine binding specificity for Cul2-Rbx1 and Cul5-Rbx2 modules of ubiquitin ligases. Genes Dev. 2004, 18, 3055–3065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Bronson, S.; Pal-Nath, D.; Miller, T.; Soto-Pantoja, D.; Roberts, D. Functions of Thrombospondin-1 in the Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 4570. [Google Scholar] [CrossRef] [PubMed]
- Kawai, K.; Tamaki, A.; Hirohata, K. Steroid-induced accumulation of lipid in the osteocytes of the rabbit femoral head. A histochemical and electron microscopic study. J. Bone Jt. Surg. 1985, 67, 755–763. [Google Scholar] [CrossRef]
- Kesarwani, P.; Prabhu, A.; Kant, S.; Chinnaiyan, P. Metabolic remodeling contributes towards an immune-suppressive phenotype in glioblastoma. Cancer Immunol. Immunother. 2019, 68, 1107–1120. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Giaccone, G. MEK inhibitors under development for treatment of non-small-cell lung cancer. Expert Opin. Investig. Drugs 2018, 27, 17–30. [Google Scholar] [CrossRef]
- Kim, H.; Lee, K.; Ko, C.-Y.; Kim, H.-S.; Shin, H.-I.; Kim, T.; Lee, S.H.; Jeong, D. The role of nacreous factors in preventing osteoporotic bone loss through both osteoblast activation and osteoclast inactivation. Biomaterials 2012, 33, 7489–7496. [Google Scholar] [CrossRef]
- Kim, T.-W.; Shin, J.-S.; Chung, K.-S.; Lee, Y.-G.; Baek, N.-I.; Lee, K.T. Anti-Inflammatory Mechanisms of Koreanaside A, a Lignan Isolated from the Flower of Forsythia koreana, against LPS-Induced Macrophage Activation and DSS-Induced Colitis Mice: The Crucial Role of AP-1, NF-κB, and JAK/STAT Signaling. Cells 2019, 8, 1163. [Google Scholar] [CrossRef] [Green Version]
- Koçak, G.; Güzel, B.N.; Mıhçı, E.; Küpesiz, O.A.; Yalçın, K.; Manguoğlu, A.E. TCIRG1 and SNX10 gene mutations in the patients with autosomal recessive osteopetrosis. Gene 2019, 702, 83–88. [Google Scholar] [CrossRef]
- Kubo, S.; Nakayamada, S.; Yoshikawa, M.; Miyazaki, Y.; Sakata, K.; Nakano, K.; Hanami, K.; Iwata, S.; Miyagawa, I.; Saito, K.; et al. Peripheral Immunophenotyping Identifies Three Subgroups Based on T Cell Heterogeneity in Lupus Patients. Arthritis Rheumatol. 2017, 69, 2029–2037. [Google Scholar] [CrossRef]
- Lan, H.-C.; Li, H.-J.; Lin, G.; Lai, P.-Y.; Chung, B.-C. Cyclic AMP Stimulates SF-1-Dependent CYP11A1 Expression through Homeodomain-Interacting Protein Kinase 3-Mediated Jun N-Terminal Kinase and c-Jun Phosphorylation. Mol. Cell. Biol. 2007, 27, 2027–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurent, C.; Nicolae, A.; Laurent, C.; Le Bras, F.; Haioun, C.; Fataccioli, V.; Amara, N.; Adélaïde, J.; Guille, A.; Schiano, J.-M.; et al. Gene alterations in epigenetic modifiers and JAK-STAT signaling are frequent in breast implant-associated ALCL. Blood 2020, 135, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kuchen, S.; Fischer, R.; Chang, S.; Lipsky, P.E. Identification and characterization of a human CD5+ pre-naive B cell population. J. Immunol. 2009, 182, 4116–4126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.S.; Hsieh, P.-H.; Chang, Y.-H.; Chan, Y.-S.; Agrawal, S.; Ueng, S.W.N. Elevated intraosseous pressure in the intertrochanteric region is associated with poorer results in osteonecrosis of the femoral head treated by multiple drilling. J. Bone Jt. Surg. 2008, 90, 852–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.-J.; Ke, F.-Y.; Lin, C.-C.; Lu, M.-Y.; Kuo, Y.-H.; Wang, Y.-P.; Liang, K.-H.; Lin, S.-C.; Chang, Y.-H.; Chen, H.-Y.; et al. ENO1 Promotes Lung Cancer Metastasis via HGFR and WNT Signaling–Driven Epithelial-to-Mesenchymal Transition. Cancer Res 2021, 81, 4094–4109. [Google Scholar] [CrossRef]
- Li, L.; Yao, Y.-C.; Gu, X.-Q.; Che, D.; Ma, C.-Q.; Dai, Z.-Y.; Li, C.; Zhou, T.; Cai, W.-B.; Yang, Z.-H.; et al. Plasminogen kringle 5 induces endothelial cell apoptosis by triggering a voltage-dependent anion channel 1 (VDAC1) positive feedback loop. J. Biol. Chem. 2014, 289, 32628–32638. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Ma, Y.-C.; Sheng, X.-Y.; Dong, H.-T.; Han, H.; Wang, J.; Xia, Y.-Y. Cyclic fluid shear stress promotes osteoblastic cells proliferation through ERK5 signaling pathway. Mol. Cell. Biochem. 2012, 364, 321–327. [Google Scholar] [CrossRef]
- Li, T.; Fu, Y.; Guo, Z.; Zhu, H.; Liao, H.; Niu, X.; Zhou, L.; Fu, S.; Li, Y.; Li, S.; et al. A new cell-free therapeutic strategy for liver regeneration: Human placental mesenchymal stem cell-derived extracellular vesicles. J. Tissue Eng. 2022, 13, 20417314221132093. [Google Scholar] [CrossRef]
- Li, W.; Mao, L.; Shu, X.; Liu, R.; Hao, F.; Li, J.; Liu, M.; Yang, L.; Zhang, W.; Sun, M.; et al. Transcriptome analysis reveals differential immune related genes expression in bovine viral diarrhea virus-2 infected goat peripheral blood mononuclear cells (PBMCs). BMC Genom. 2019, 20, 516. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, Y.; Luo, J.; Fu, X.; Liu, P.; Liu, S.; Pan, Y. FAM126A interacted with ENO1 mediates proliferation and metastasis in pancreatic cancer via PI3K/AKT signaling pathway. Cell Death Discov. 2022, 8, 248. [Google Scholar] [CrossRef]
- Lim, S.S.; Drenkard, C.; McCune, W.J.; Helmick, C.G.; Gordon, C.; DeGuire, P.; Bayakly, R.; Somers, E.C. Population-based lupus registries: Advancing our epidemiologic understanding. Arthritis Rheum. 2009, 61, 1462–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotinun, S.; Ishihara, Y.; Nagano, K.; Kiviranta, R.; Carpentier, V.T.; Neff, L.; Parkman, V.; Ide, N.; Hu, D.; Dann, P.; et al. Cathepsin K–deficient osteocytes prevent lactation-induced bone loss and parathyroid hormone suppression. J. Clin. Investig. 2019, 129, 3058–3071. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Xin, D.E.; Du, J.K.; Zou, Q.C.; Wu, Q.; Zhang, Y.S.; Deng, W.; Yue, J.; Fan, X.S.; Zeng, Y.; et al. Loss of LOXL2 Promotes Uterine Hypertrophy and Tumor Progression by Enhancing H3K36ac-Dependent Gene Expression. Cancer Res. 2022, 82, 4400–4413. [Google Scholar] [CrossRef] [PubMed]
- Mancini, T.; Kola, B.; Mantero, F.; Boscaro, M.; Arnaldi, G. High cardiovascular risk in patients with Cushing’s syndrome according to 1999 WHO/ISH guidelines. Clin. Endocrinol. 2004, 61, 768–777. [Google Scholar] [CrossRef]
- Mao, L.; Liao, C.; Qin, J.; Gong, Y.; Zhou, Y.; Li, S.; Liu, Z.; Deng, H.; Deng, W.; Sun, Q.; et al. Phosphorylation of SNX27 by MAPK11/14 links cellular stress–signaling pathways with endocytic recycling. J. Cell Biol. 2021, 220, 202010048. [Google Scholar] [CrossRef]
- Mistry, P.; Nakabo, S.; O’Neil, L.; Goel, R.R.; Jiang, K.; Carmona-Rivera, C.; Gupta, S.; Chan, D.W.; Carlucci, P.M.; Wang, X.; et al. Transcriptomic, epigenetic, and functional analyses implicate neutrophil diversity in the pathogenesis of systemic lupus erythematosus. Proc. Natl. Acad. Sci. USA 2019, 116, 25222–25228. [Google Scholar] [CrossRef]
- Mo, H.-Y.; Wei, J.C.C.; Chen, X.-H.; Chen, H.-H. Increased risk of systemic lupus erythematosus in patients with autoimmune haemolytic anaemia: A nationwide population-based cohort study. Ann. Rheum. Dis. 2020, 80, 403–404. [Google Scholar] [CrossRef]
- Mont, M.A.; Pivec, R.; Banerjee, S.; Issa, K.; Elmallah, R.K.; Jones, L.C. High-Dose Corticosteroid Use and Risk of Hip Osteonecrosis: Meta-Analysis and Systematic Literature Review. J. Arthroplast. 2015, 30, 1506–1512.e1505. [Google Scholar] [CrossRef]
- Monte, M.; Benetti, R.; Buscemi, G.; Sandy, P.; Del Sal, G.; Schneider, C. The Cell Cycle-regulated Protein Human GTSE-1 Controls DNA Damage-induced Apoptosis by Affecting p53 Function. J. Biol. Chem. 2003, 278, 30356–30364. [Google Scholar] [CrossRef] [Green Version]
- Monte, M.; Benetti, R.; Collavin, L.; Marchionni, L.; Del Sal, G.; Schneider, C. hGTSE-1 Expression Stimulates Cytoplasmic Localization of p53. J. Biol. Chem. 2004, 279, 11744–11752. [Google Scholar] [CrossRef]
- Motomura, G.; Yamamoto, T.; Miyanishi, K.; Yamashita, A.; Sueishi, K.; Iwamoto, Y. Bone marrow fat-cell enlargement in early steroid-induced osteonecrosis—A histomorphometric study of autopsy cases. Pathol. Res. Pr. 2005, 200, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, H.; Song, J.; Sugimoto, M.; Hagihara, K.; Kishimoto, T.; Yoshizaki, K.; Nishimoto, N. Anti-interleukin-6 receptor antibody therapy reduces vascular endothelial growth factor production in rheumatoid arthritis. Arthritis Rheum. 2003, 48, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Narayan, A.; Berger, B.; Cho, H. Assessing single-cell transcriptomic variability through density-preserving data visualization. Nat. Biotechnol. 2021, 39, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Nehar-Belaid, D.; Hong, S.; Marches, R.; Chen, G.; Bolisetty, M.; Baisch, J.; Walters, L.; Punaro, M.; Rossi, R.J.; Chung, C.-H.; et al. Mapping systemic lupus erythematosus heterogeneity at the single-cell level. Nat. Immunol. 2020, 21, 1094–1106. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Wang, Z.; Chai, G.; Xiong, Y.; Li, B.; Zhang, H.; Xin, R.; Qian, X.; Tang, Z.; Wu, J.; et al. Dehydrocostus Lactone Suppresses LPS-induced Acute Lung Injury and Macrophage Activation through NF-κB Signaling Pathway Mediated by p38 MAPK and Akt. Molecules 2019, 24, 1510. [Google Scholar] [CrossRef] [Green Version]
- Oinuma, K.; Harada, Y.; Nawata, Y.; Takabayashi, K.; Abe, I.; Kamikawa, K.; Moriya, H. Osteonecrosis in patients with systemic lupus erythematosus develops very early after starting high dose corticosteroid treatment. Ann. Rheum. Dis. 2001, 60, 1145–1148. [Google Scholar] [CrossRef] [Green Version]
- Otaify, G.A.; Abdel-Hamid, M.S.; Mehrez, M.I.; Aboul-Ezz, E.; Zaki, M.S.; Aglan, M.S.; Temtamy, S.A. Genetic study of eight Egyptian patients with pycnodysostosis: Identification of novel CTSK mutations and founder effect. Osteoporos. Int. 2018, 29, 1833–1841. [Google Scholar] [CrossRef]
- Ou, Y.-H.; Chung, P.-H.; Sun, T.-P.; Shieh, S.-Y. p53 C-Terminal Phosphorylation by CHK1 and CHK2 Participates in the Regulation of DNA-Damage-induced C-Terminal Acetylation. Mol. Biol. Cell 2005, 16, 1684–1695. [Google Scholar] [CrossRef] [Green Version]
- Petri, M. Musculoskeletal complications of systemic lupus erythematosus in the hopkins lupus cohort: An update. Arthritis Rheum. 1995, 8, 137–145. [Google Scholar] [CrossRef]
- Plotnikov, A.; Zehorai, E.; Procaccia, S.; Seger, R. The MAPK cascades: Signaling components, nuclear roles and mechanisms of nuclear translocation. Biochim. et Biophys. Acta (BBA) Mol. Cell Res. 2011, 1813, 1619–1633. [Google Scholar] [CrossRef]
- Pohl, S.; Angermann, A.; Jeschke, A.; Hendrickx, G.; A Yorgan, T.; Makrypidi-Fraune, G.; Steigert, A.; Kuehn, S.C.; Rolvien, T.; Schweizer, M.; et al. The Lysosomal Protein Arylsulfatase B Is a Key Enzyme Involved in Skeletal Turnover. J. Bone Miner. Res. 2018, 33, 2186–2201. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.; Buckley, J.A.; Li, X.; Liu, Y.; Fox, T.H.; Meares, G.; Yu, H.; Yan, Z.; Harms, A.; Li, Y.; et al. Inhibition of the JAK/STAT Pathway Protects Against α-Synuclein-Induced Neuroinflammation and Dopaminergic Neurodegeneration. J. Neurosci. 2016, 36, 5144–5159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raggi, C.; Taddei, M.L.; Sacco, E.; Navari, N.; Correnti, M.; Piombanti, B.; Pastore, M.; Campani, C.; Pranzini, E.; Iorio, J.; et al. Mitochondrial oxidative metabolism contributes to a cancer stem cell phenotype in cholangiocarcinoma. J. Hepatol. 2021, 74, 1373–1385. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Isenberg, D.A. Systemic Lupus Erythematosus. N. Engl. J. Med. 2008, 358, 929–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauch, A.; Seitz, S.; Baschant, U.; Schilling, A.F.; Illing, A.; Stride, B.; Kirilov, M.; Mandic, V.; Takacz, A.; Schmidt-Ullrich, R.; et al. Glucocorticoids suppress bone formation by attenuating osteoblast differentiation via the monomeric glucocorticoid receptor. Cell Metab. 2010, 11, 517–531. [Google Scholar] [CrossRef] [Green Version]
- Rees, F.; Doherty, M.; Grainge, M.; Davenport, G.; Lanyon, P.; Zhang, W. The incidence and prevalence of systemic lupus erythematosus in the UK, 1999–2012. Ann. Rheum. Dis. 2016, 75, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Rees, F.; Doherty, M.; Grainge, M.; Lanyon, P.; Zhang, W. The worldwide incidence and prevalence of systemic lupus erythematosus: A systematic review of epidemiological studies. Rheumatology 2017, 56, 1945–1961. [Google Scholar] [CrossRef] [Green Version]
- Shaath, H.; Vishnubalaji, R.; Elkord, E.; Alajez, N.M. Single-Cell Transcriptome Analysis Highlights a Role for Neutrophils and Inflammatory Macrophages in the Pathogenesis of Severe COVID-19. Cells 2020, 9, 2374. [Google Scholar] [CrossRef]
- Shi, Y.; Xu, X.; Zhang, Q.; Fu, G.; Mo, Z.; Wang, G.S.; Kishi, S.; Yang, X.-L. tRNA synthetase counteracts c-Myc to develop functional vasculature. eLife 2014, 3, e02349. [Google Scholar] [CrossRef]
- Shieh, S.-Y.; Ahn, J.; Tamai, K.; Taya, Y.; Prives, C. The human homologs of checkpoint kinases Chk1 and Cds1 (Chk2) phosphorylate p53 at multiple DNA damage-inducible sites. Genes Dev. 2000, 14, 289–300. [Google Scholar] [CrossRef]
- Tao, S.-C.; Yuan, T.; Rui, B.-Y.; Zhu, Z.-Z.; Guo, S.-C.; Zhang, C.-Q. Exosomes derived from human platelet-rich plasma prevent apoptosis induced by glucocorticoid-associated endoplasmic reticulum stress in rat osteonecrosis of the femoral head via the Akt/Bad/Bcl-2 signal pathway. Theranostics 2017, 7, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Titanji, K.; Ofotokun, I.; Weitzmann, M.N. Immature/transitional B-cell expansion is associated with bone loss in HIV-infected individuals with severe CD4+ T-cell lymphopenia. Aids 2020, 34, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, K.; Yoshimoto, T.; Torigoe, K.; Kurimoto, M.; Matsui, K.; Hada, T.; Okamura, H.; Nakanishi, K. IL-12 synergizes with IL-18 or IL-1β for IFN-γ production from human T cells. Int. Immunol. 2000, 12, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Trujillo-Martín, M.M.; de Larrinoa, I.R.-F.F.; Ruíz-Irastorza, G.; Pego-Reigosa, J.M.; Sánchez, J.M.S.; Serrano-Aguilar, P. Clinical practice guidelines for systemic lupus erythematosus: Recommendations for general clinical management. Med. Clin. 2016, 146, 413.e1–413.e14. [Google Scholar] [CrossRef]
- Verrier, F.; Mignotte, B.; Jan, G.; Brenner, C. Study of PTPC Composition during Apoptosis for Identification of Viral Protein Target. Ann. N. Y. Acad. Sci. 2003, 1010, 126–142. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-J.; Cui, Q.; Balian, G. The Nicolas Andry award. The pathogenesis and prevention of steroid induced osteonecrosis. Clin. Orthop. Relat. Res. 2000, 370, 295–310. [Google Scholar] [CrossRef]
- Wang, X.-S.; Zhuang, Q.-Y.; Weng, X.-S.; Lin, J.; Jin, J.; Qian, W.-W. Etiological and clinical analysis of osteonecrosis of the femoral head in Chinese patients. Chin. Med. J. 2013, 126, 290–295. [Google Scholar]
- Weinstein, R.S.; Chen, J.-R.; Powers, C.C.; Stewart, S.A.; Landes, R.D.; Bellido, T.; Jilka, R.L.; Parfitt, A.M.; Manolagas, S.C. Promotion of osteoclast survival and antagonism of bisphosphonate-induced osteoclast apoptosis by glucocorticoids. J. Clin. Investig. 2002, 109, 1041–1048. [Google Scholar] [CrossRef]
- Weinstein, R.S.; Nicholas, R.W.; Manolagas, S.C. Apoptosis of Osteocytes in Glucocorticoid-Induced Osteonecrosis of the Hip 1. J. Clin. Endocrinol. Metab. 2000, 85, 2907–2912. [Google Scholar] [CrossRef]
- Willems, F.; Marchant, A.; Delville, J.-P.; Gérard, C.; Delvaux, A.; Velu, T.; De Boer, M.; Goldman, M. Interleukin-10 inhibits B7 and intercellular adhesion molecule-1 expression on human monocytes. Eur. J. Immunol. 1994, 24, 1007–1009. [Google Scholar] [CrossRef]
- Winkler, I.G.; Sims, N.A.; Pettit, A.R.; Barbier, V.; Nowlan, B.; Helwani, F.; Poulton, I.J.; Van Rooijen, N.; Alexander, K.; Raggatt, L.J.; et al. Bone marrow macrophages maintain hematopoietic stem cell (HSC) niches and their depletion mobilizes HSCs. Blood 2010, 116, 4815–4828. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Li, Z.; Weng, X. The role of lncRNA RP11-154D6 in steroid-induced osteonecrosis of the femoral head through BMSC regulation. J. Cell. Biochem. 2019, 120, 18435–18445. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Li, Z.; Weng, X. Changed cellular functions and aberrantly expressed miRNAs and circRNAs in bone marrow stem cells in osteonecrosis of the femoral head. Int. J. Mol. Med. 2020, 45, 805–815. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Xu, T.; Xie, T.; Yang, L.; Wang, G.; Gao, Y.; Xi, G.; Zhang, X. CDC42EP3 promotes glioma progression via regulation of CCND1. Cell Death Dis. 2022, 13, 290. [Google Scholar] [CrossRef]
- Yoon, S.I.; Logsdon, N.J.; Sheikh, F.; Donnelly, R.P.; Walter, M.R. Conformational Changes Mediate Interleukin-10 Receptor 2 (IL-10R2) Binding to IL-10 and Assembly of the Signaling Complex. J. Biol. Chem. 2006, 281, 35088–35096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Q.Y.; Chen, R.; Darmawan, J.; Xiao, Z.Y.; Chen, S.B.; Wigley, R.; Le Chen, S.; Zhang, N.Z. Rheumatic Diseases in China. Thromb. Haemost. 2008, 10, R17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.-L.; Zhou, B.-W.; Yan, Z.-Z.; Zhao, J.; Zhao, B.-C.; Liu, W.-F.; Li, C.; Liu, K.-X. 6-Gingerol attenuates macrophages pyroptosis via the inhibition of MAPK signaling pathways and predicts a good prognosis in sepsis. Cytokine 2019, 125, 154854. [Google Scholar] [CrossRef]
- Zhang, N.-F.; Li, Z.R.; Wei, H.-Y.; Liu, Z.-H.; Hernigou, P. Steroid-induced osteonecrosis: The number of lesions is related to the dosage. J. Bone Jt. Surg. 2008, 90, 1239–1243. [Google Scholar] [CrossRef]
- Zhang, Y.; Ji, D.; Li, L.; Yang, S.; Zhang, H.; Duan, X. ClC-7 Regulates the Pattern and Early Development of Craniofacial Bone and Tooth. Theranostics 2019, 9, 1387–1400. [Google Scholar] [CrossRef]
- Zhao, S.; Kong, F.-Q.; Cai, W.; Xu, T.; Zhou, Z.-M.; Wang, Z.-B.; Xu, A.-D.; Yang, Y.-Q.; Chen, J.; Tang, P.-Y.; et al. GIT1 contributes to autophagy in osteoclast through disruption of the binding of Beclin1 and Bcl2 under starvation condition. Cell Death Dis. 2018, 9, 1195. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, W.; Guo, S.; Bai, R.; Yan, J. Mechanism of Methylprednisolone-Induced Primary Cilia Formation Disorder and Autophagy in Osteoblasts. Orthop. Surg. 2020, 12, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Zuo, R.; Kong, L.; Wang, M.; Wang, W.; Xu, J.; Chai, Y.; Guan, J.; Kang, Q. Exosomes derived from human CD34+ stem cells transfected with miR-26a prevent glucocorticoid-induced osteonecrosis of the femoral head by promoting angiogenesis and osteogenesis. Stem Cell Res. Ther. 2019, 10, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Myelocyte | Neutrophil | Pro-Myelocyte | T Cell: CD4+ Central_Memory | T Cell: CD8+ | B Cell: Immature | Monocyte: CD16− | T Cell: CD4+ | B Cell: Plasma Cell | GMP | CMP | Erythroblast | Pro B Cell: CD34+ | BM | NK Cell | PreB Cell CD34− | Macrophage: Monocyte Derived | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lysosome | √ | — | — | — | — | √ | — | — | — | √ | — | — | — | — | — | — | √ |
| Wnt signaling pathway | √ | √ | √ | — | √ | — | — | — | — | — | √ | √ | √ | — | √ | √ | √ |
| Pyruvate metabolism | √ | — | — | — | — | — | √ | √ | — | √ | — | √ | — | — | — | √ | — |
| Insulin signaling pathway | — | √ | — | — | √ | — | √ | √ | — | — | — | — | √ | — | — | — | √ |
| P53 signaling pathway | — | √ | — | √ | — | √ | — | — | — | √ | √ | √ | — | — | — | √ | — |
| Glycolysis and gluconeogenesis | — | √ | — | — | √ | — | √ | √ | — | √ | √ | √ | √ | — | √ | — | √ |
| Calcium signaling pathway | — | √ | √ | — | — | √ | — | √ | √ | — | — | — | — | — | — | √ | √ |
| NLR signaling pathway | — | — | √ | √ | √ | √ | — | √ | √ | √ | √ | — | √ | — | √ | — | √ |
| MAPK signaling pathway | — | — | √ | √ | √ | √ | √ | √ | √ | √ | √ | — | — | √ | — | √ | √ |
| Complement and coagulation cascades | — | — | √ | √ | — | √ | — | √ | — | √ | — | — | — | — | — | √ | √ |
| Oxidative phosphorylation | — | — | √ | √ | — | √ | — | — | — | — | — | √ | — | — | — | — | √ |
| TLR signaling pathway | — | — | — | √ | √ | √ | — | √ | √ | √ | — | — | √ | — | √ | — | — |
| TGF-Beta signaling pathway | — | — | — | √ | — | — | — | √ | — | — | — | √ | — | — | — | √ | — |
| Apoptosis | — | — | — | √ | √ | — | — | √ | — | — | √ | — | — | — | — | — | — |
| Peroxisome | — | — | — | √ | — | — | √ | — | — | — | — | √ | — | — | √ | — | √ |
| Systemic lupus erythematosus | — | — | √ | — | — | √ | — | — | — | — | — | — | — | — | — | — | √ |
| JAK-STAT signaling pathway | — | — | — | — | — | √ | — | — | — | — | — | — | √ | — | — | — | √ |
| Vascular smooth muscle contraction | — | — | — | — | — | √ | — | — | — | — | — | — | — | √ | √ | √ | — |
| Leukocyte trans-endothelial migration | — | — | — | — | — | √ | — | — | — | √ | — | √ | — | — | — | — | √ |
| PPAR signaling pathway | — | — | — | √ | — | — | √ | — | — | √ | — | √ | √ | — | — | √ | — |
| Fatty acid metabolism | — | — | — | — | — | — | √ | √ | √ | — | — | √ | √ | — | — | √ | — |
| Cell cycle | — | — | — | — | — | — | — | √ | — | — | √ | √ | — | — | — | √ | — |
| Glutathione metabolism | — | — | — | — | — | — | — | — | — | — | — | √ | — | — | — | √ | √ |
| Adipocytokine signaling pathway | — | — | — | √ | — | — | — | — | — | — | — | √ | √ | — | — | — | — |
| mTOR signaling pathway | — | — | — | — | √ | √ | — | — | — | — | — | — | — | — | — | √ | — |
| NOTCH signaling pathway | — | — | — | — | — | — | √ | — | √ | — | — | — | — | — | — | — | √ |
| Citrate cycle TCA cycle | — | — | — | — | — | — | — | — | — | — | — | √ | — | — | — | — | — |
| Pentose phosphate pathway | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — | √ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, D.; Chen, H.; Li, Z.; Feng, B.; Weng, X. Accumulation of Fat Not Responsible for Femoral Head Necrosis, Revealed by Single-Cell RNA Sequencing: A Preliminary Study. Biomolecules 2023, 13, 171. https://doi.org/10.3390/biom13010171
Wang Y, Li D, Chen H, Li Z, Feng B, Weng X. Accumulation of Fat Not Responsible for Femoral Head Necrosis, Revealed by Single-Cell RNA Sequencing: A Preliminary Study. Biomolecules. 2023; 13(1):171. https://doi.org/10.3390/biom13010171
Chicago/Turabian StyleWang, Yingjie, Dandan Li, Haijia Chen, Zhuolin Li, Bin Feng, and Xisheng Weng. 2023. "Accumulation of Fat Not Responsible for Femoral Head Necrosis, Revealed by Single-Cell RNA Sequencing: A Preliminary Study" Biomolecules 13, no. 1: 171. https://doi.org/10.3390/biom13010171